
Single-Cell Oils from Oleaginous Microorganisms as Green Bio-Lubricants: Studies on Their Tribological Performance

 ,
, 
 , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Strains and Cultivation Conditions
2.1.1. Microalgae Cultivation
2.1.2. Thraustochytrids Cultivation
2.1.3. Yeast Cultivation
2.2. Monitoring of Lipid Accumulation in Cultures
2.3. Lipid Extraction from Biomass
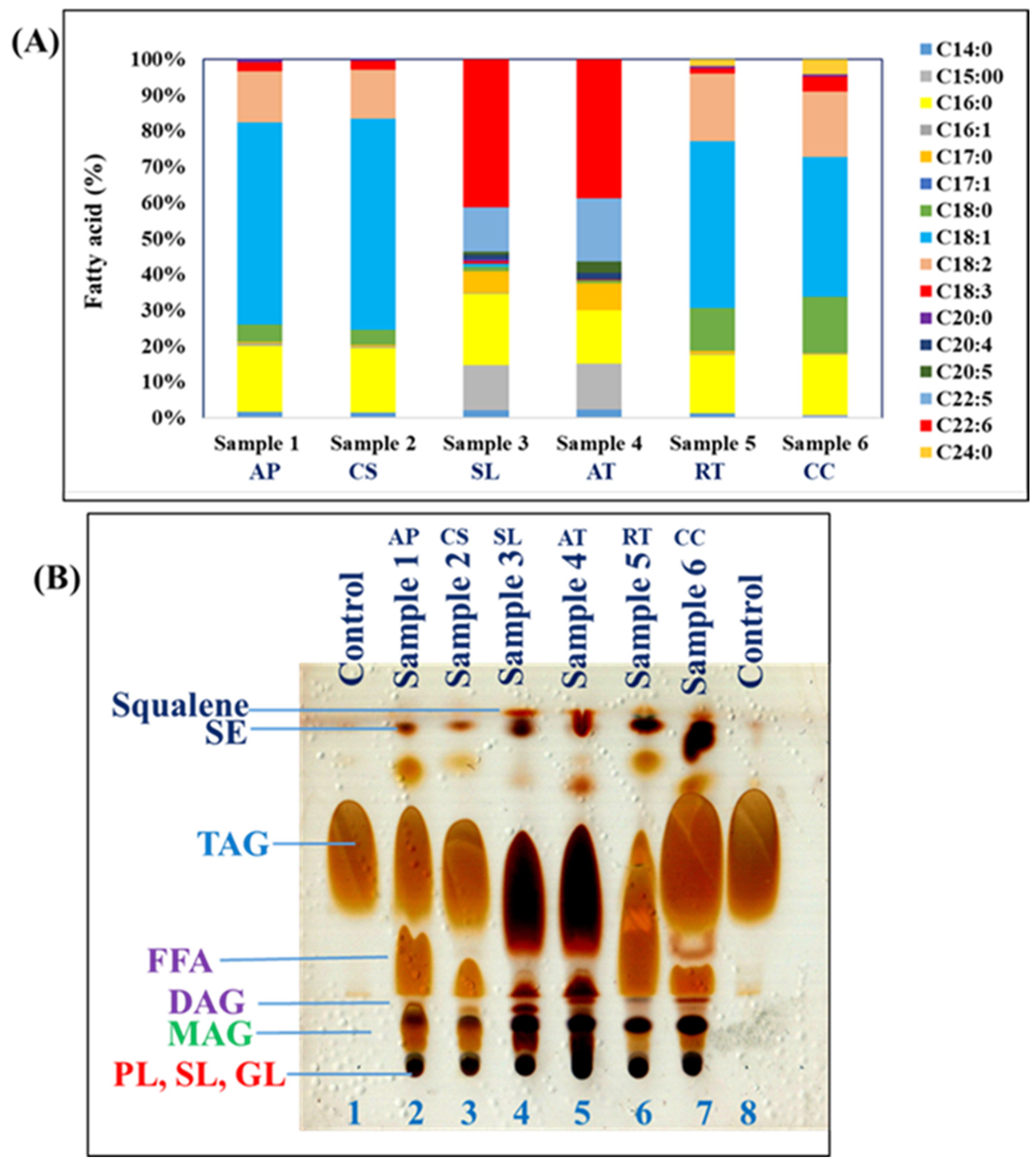
2.4. Determination of Fatty Acid Profiles by Gas Chromatography-Mass Spectrometry (GC-MS) and Thin-Layer Chromatography (TLC)
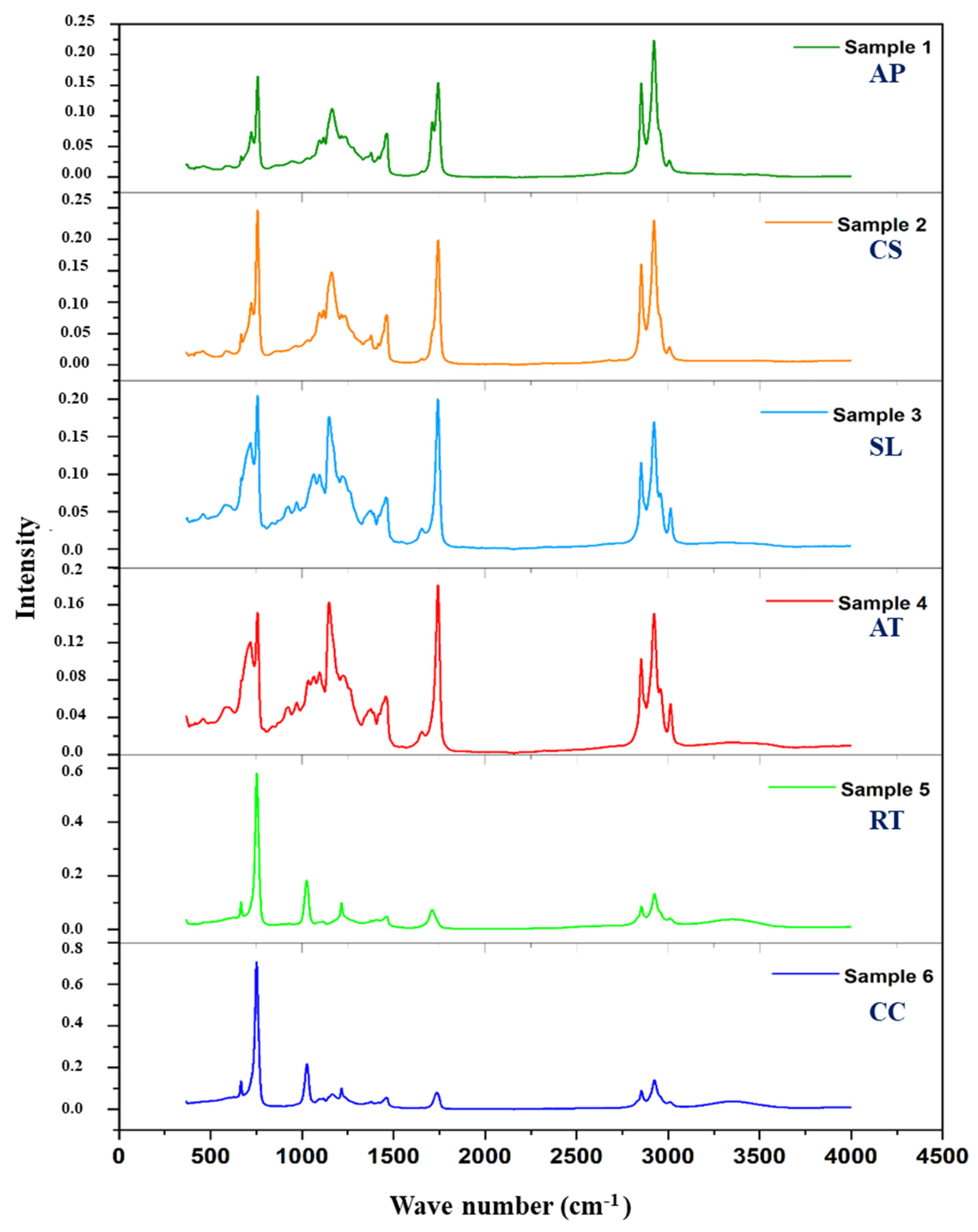
2.5. Infrared Spectroscopy (IR) Analysis of Microbial Oils
2.6. Rheological Properties of Microbial Oils
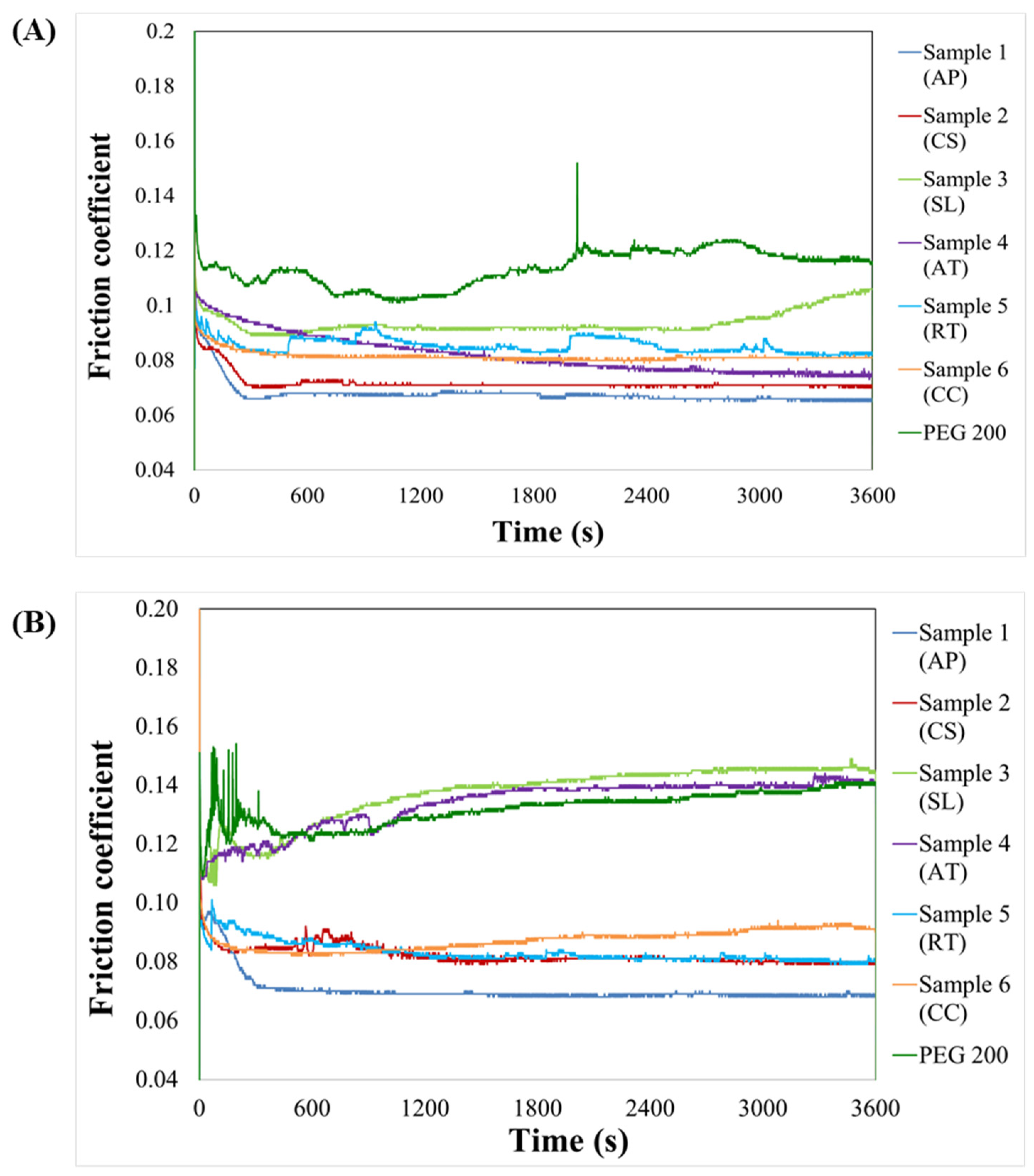
2.7. Determination of Lubrication Properties
2.8. Thermogravimetric Analysis (TGA) of Microbial Oils
2.9. Statistical Analysis
3. Results and Discussion
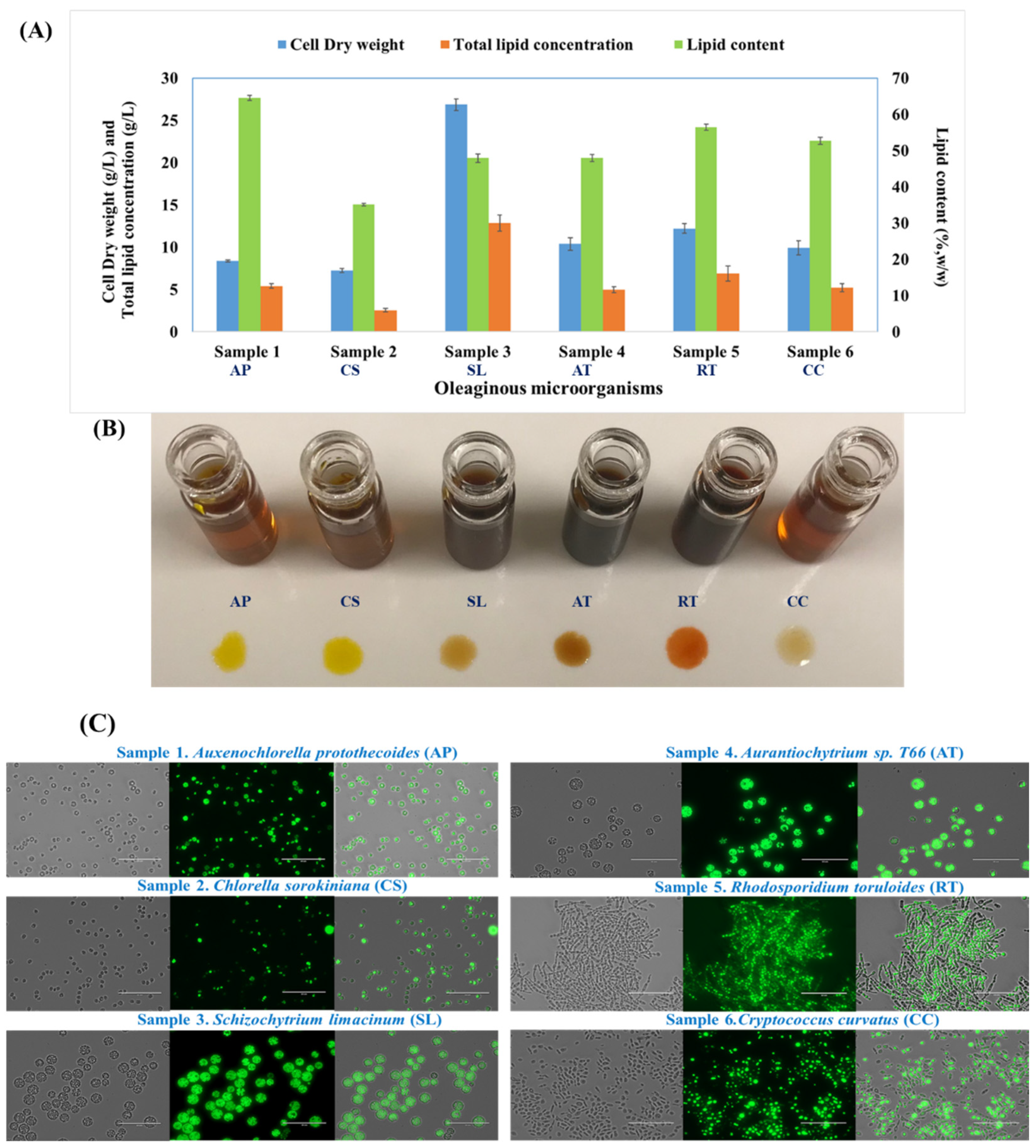
3.1. Biomass and Lipid Production by Oleaginous Microalgae, Yeasts, and Thraustochytrids
3.2. Fatty Acid Profile of Oleaginous Microorganisms
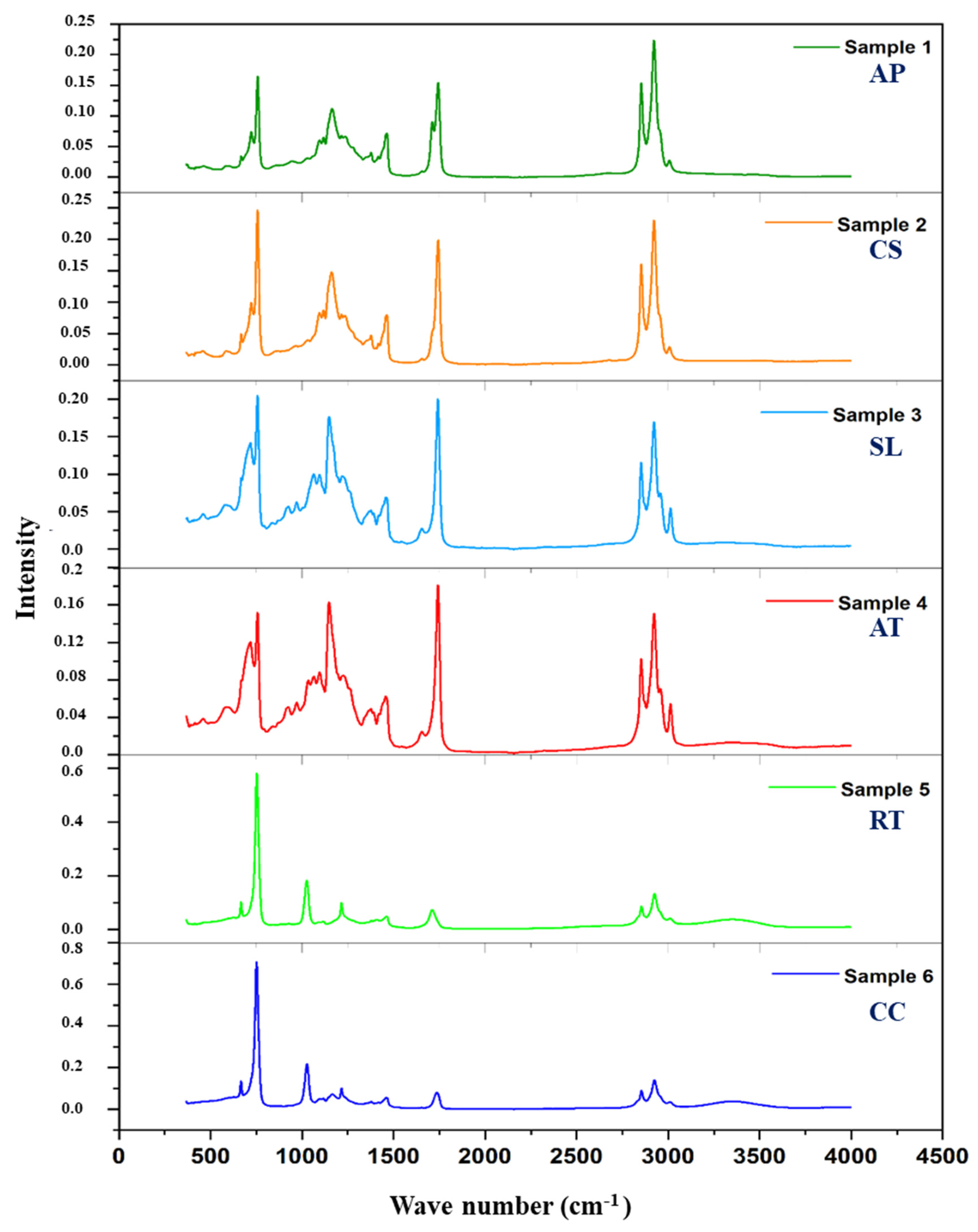
3.3. IR
3.4. Tribological Studies of Microbial Lipids
3.4.1. Friction Coefficient
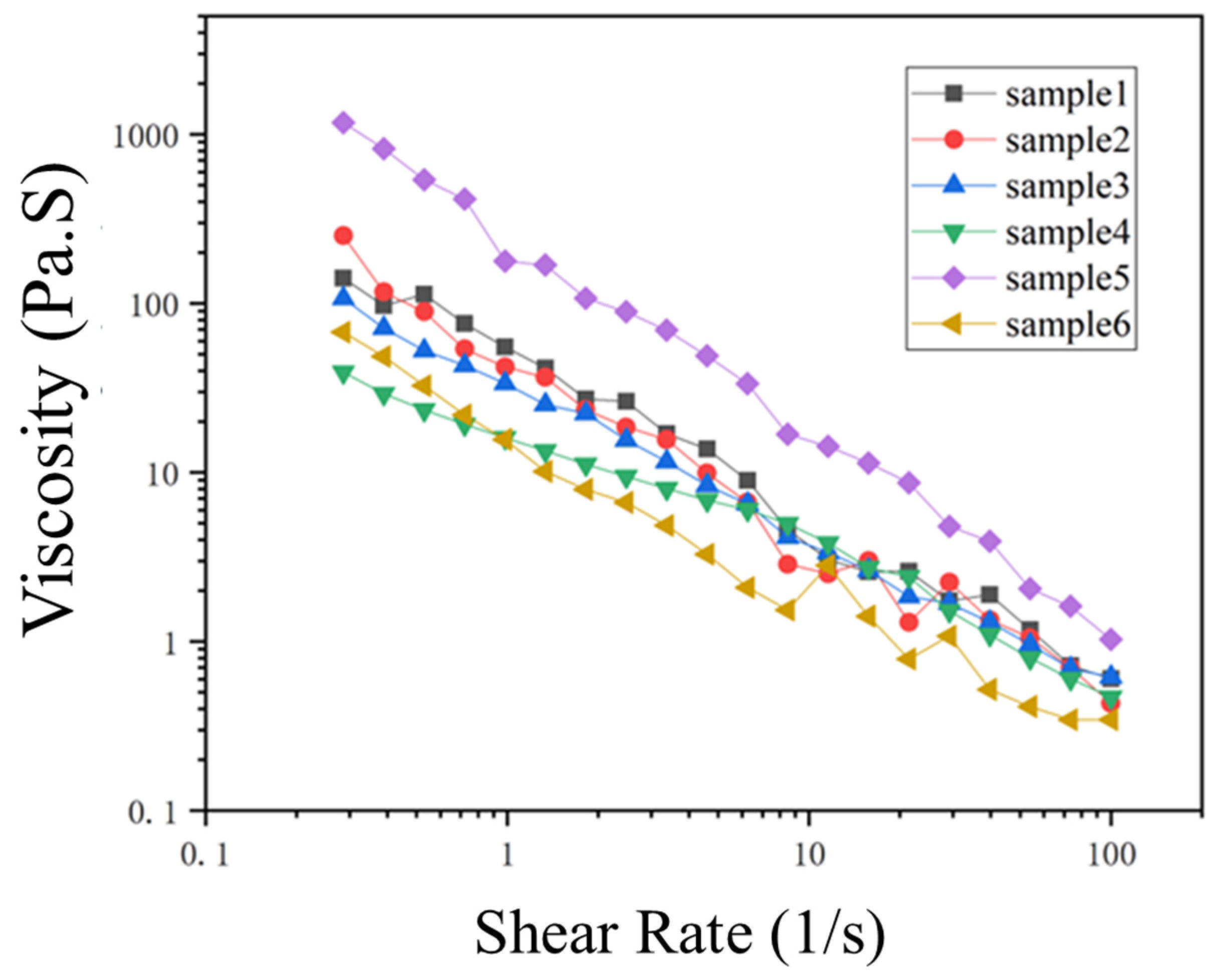
3.4.2. Viscosity
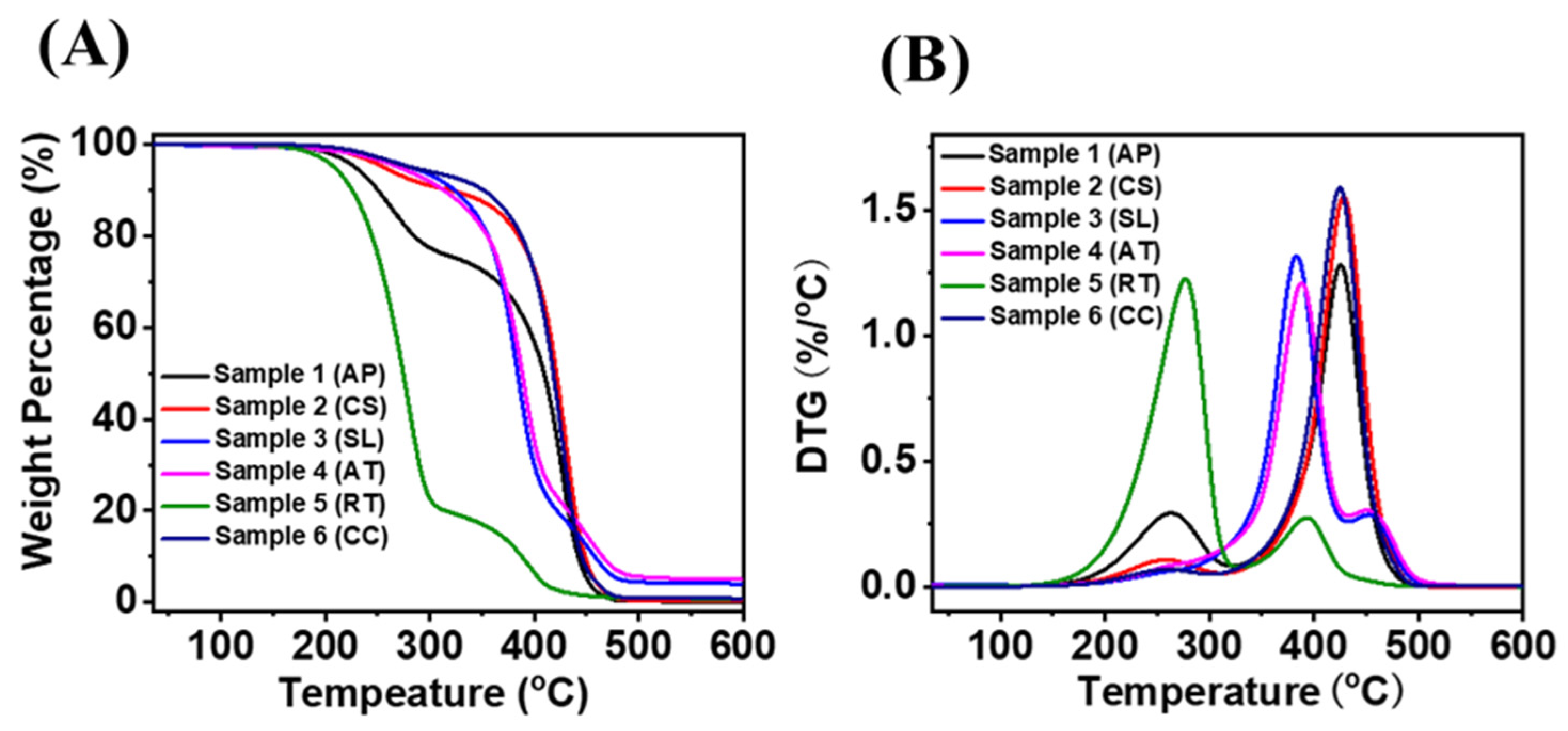
3.4.3. TGA of Microbial Oils
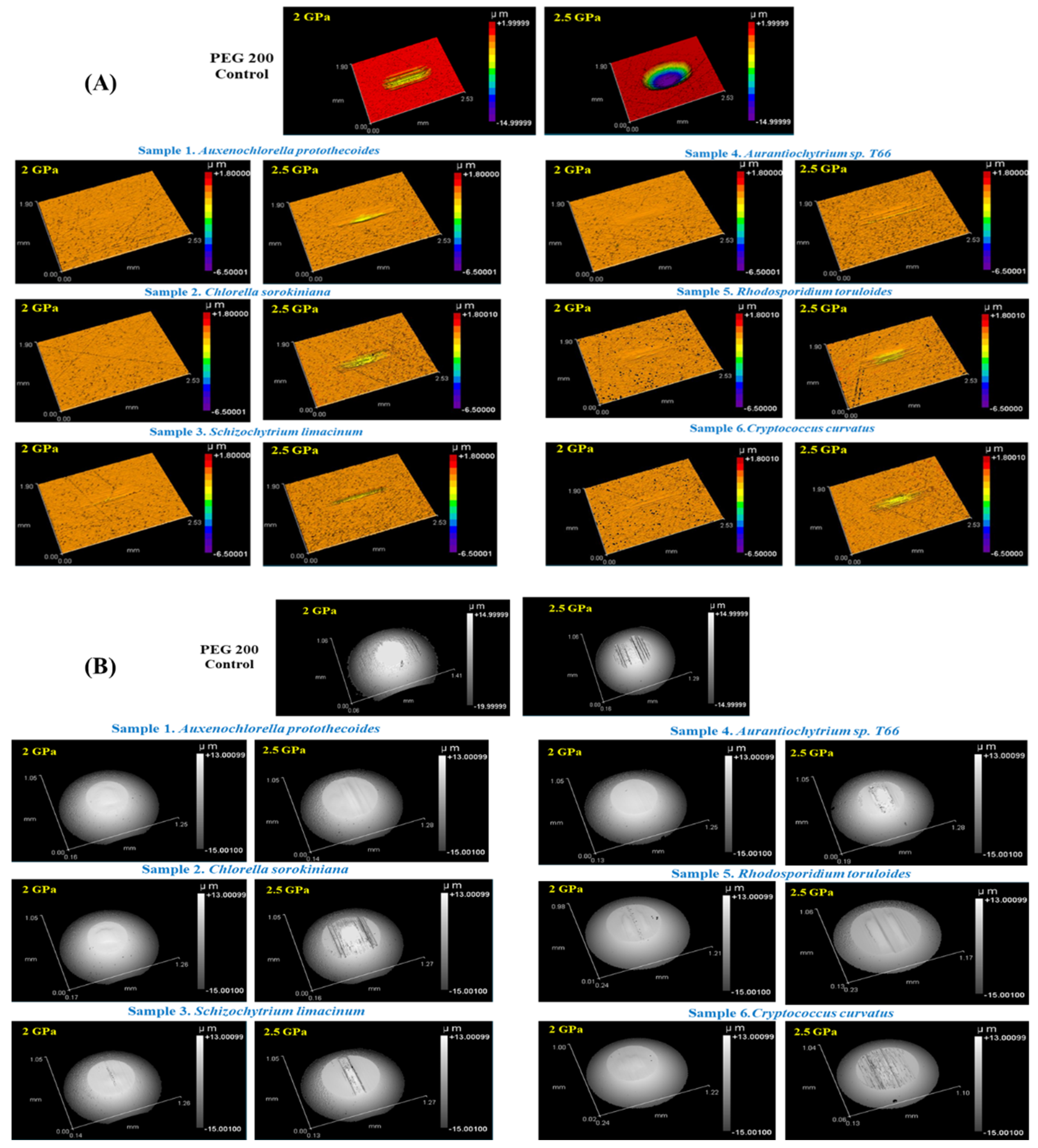
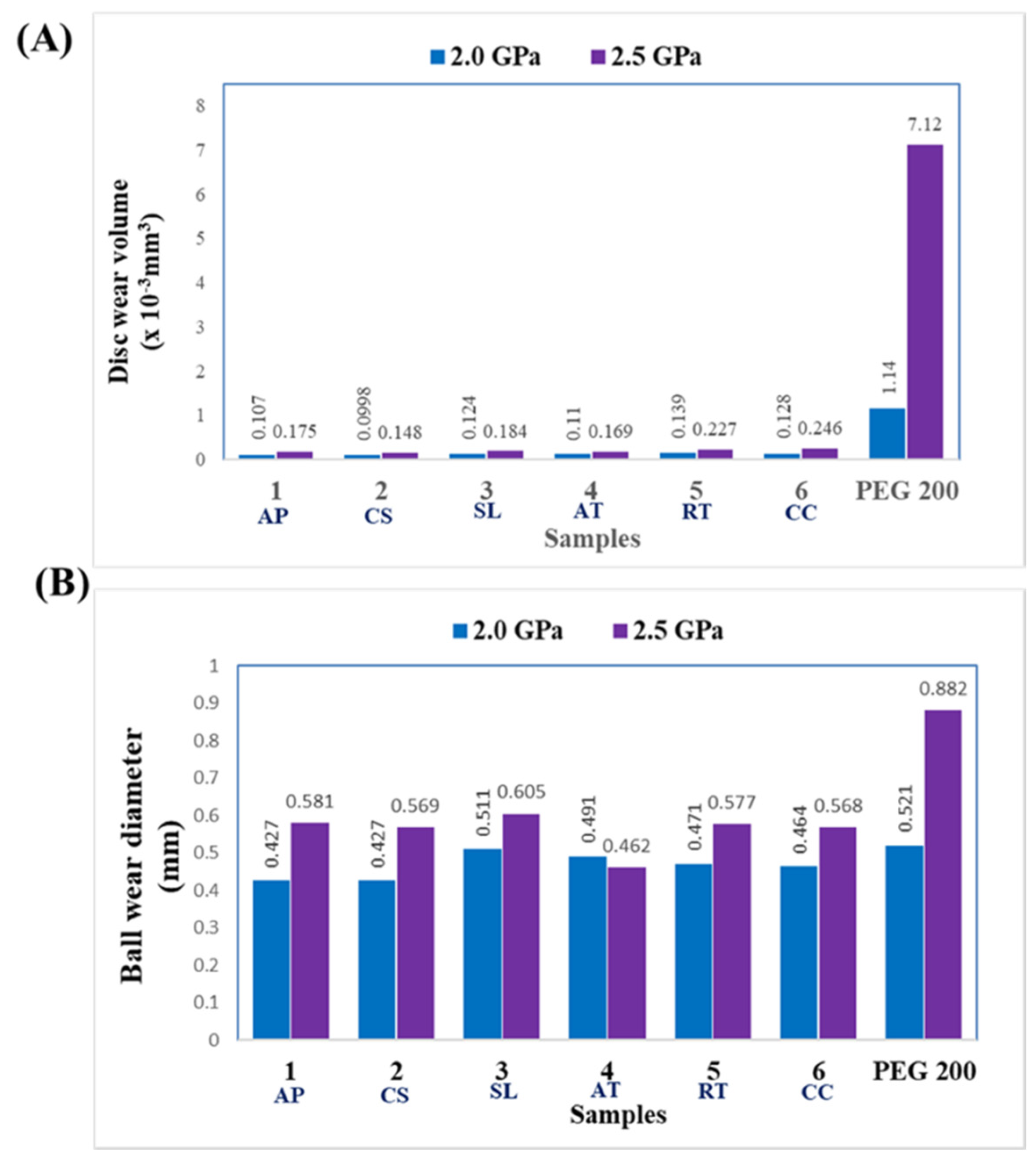
3.4.4. Wear Characteristic
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ASTM | American Society for Testing and Materials |
| BBM | Bold’s basal medium |
| CDW | Cell dry weight |
| ATCC | American Type Culture Collection |
| C/N | Carbon/nitrogen |
| DAG | Diacylglycerols |
| DHA | Docosahexaenoic acid |
| FAMEs | Fatty acid methyl esters |
| FC | Friction coefficient |
| FFA | Free fatty acids |
| GC-MS | Gas chromatography-mass spectrometry |
| h | Hour(s) |
| LD | Lipid droplets |
| MAG | Monoacylglycerols |
| MUFAs | Mono-unsaturated fatty acids |
| NCYC | National collection of yeast cultures |
| NREL | National Renewable Energy Laboratory |
| OD | Optical density |
| PEG | Polyethylene glycol |
| PL | Phospholipids |
| PUFAs | Poly-unsaturated fatty acids |
| SFAs | Saturated fatty acids |
| SL | Sphingolipids |
| TAGs | Triacylglycerols |
| TLC | Thin layer chromatography |
| YPD | Yeast extract peptone dextrose |
References
- Sheldon, R.A. The Road to Biorenewables: Carbohydrates to Commodity Chemicals. ACS Sustain. Chem. Eng. 2018, 6, 4464–4480. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, R.A. Metrics of Green Chemistry and Sustainability: Past, Present, and Future. ACS Sustain. Chem. Eng. 2018, 6, 32–48. [Google Scholar] [CrossRef] [Green Version]
- Hamdan, S.H.; Chong, W.W.F.; Ng, J.H.; Chong, C.T.; Zhang, H. Nano-tribological characterisation of palm oil-based trimethylolpropane ester for application as boundary lubricant. Tribol. Int. 2018, 127, 1–9. [Google Scholar] [CrossRef]
- Schneider, M.P. Plant-oil-based lubricants and hydraulic fluids. J. Sci. Food Agric. 2006, 86, 1769–1780. [Google Scholar] [CrossRef]
- Singh, Y.; Farooq, A.; Raza, A.; Mahmood, M.A.; Jain, S. Sustainability of a non-edible vegetable oil based bio-lubricant for automotive applications: A review. Process Saf. Environ. Prot. 2017, 111, 701–713. [Google Scholar] [CrossRef]
- Shah, R.; Woydt, M.; Zhang, S. The Economic and Environmental Significance of Sustainable Lubricants. Lubricants 2021, 9, 21. [Google Scholar] [CrossRef]
- Nagendramma, P.; Kaul, S. Development of ecofriendly/biodegradable lubricants: An overview. Renew. Sustain. Energy Rev. 2012, 16, 764–774. [Google Scholar] [CrossRef]
- Cecilia, J.A.; Plata, D.B.; Saboya, R.M.A.; de Luna, F.M.T.; Cavalcante, C.L.; Rodríguez-Castellón, E. An overview of the biolubricant production process: Challenges and future perspectives. Processes 2020, 8, 257. [Google Scholar] [CrossRef] [Green Version]
- Lovell, M.; Higgs, C.F.; Deshmukh, P.; Mobley, A. Increasing formability in sheet metal stamping operations using environmentally friendly lubricants. J. Mater. Process. Technol. 2006, 177, 87–90. [Google Scholar] [CrossRef]
- Patel, A.; Rova, U.; Christakopoulos, P.; Matsakas, L. Assessment of Fatty Acids Profile and Omega-3 Polyunsaturated Fatty Acid Production by the Oleaginous Marine Thraustochytrid Aurantiochytrium sp. T66 Cultivated on Volatile Fatty Acids. Biomolecules 2020, 10, 694. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part II: Technology and potential applications. Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073. [Google Scholar] [CrossRef]
- Patel, A.; Arora, N.; Sartaj, K.; Pruthi, V.; Pruthi, P.A. Sustainable biodiesel production from oleaginous yeasts utilizing hydrolysates of various non-edible lignocellulosic biomasses. Renew. Sustain. Energy Rev. 2016, 62, 836–855. [Google Scholar] [CrossRef]
- Patel, A.; Pruthi, V.; Pruthi, P.A. Innovative screening approach for the identification of triacylglycerol accumulating oleaginous strains. Renew. Energy 2019, 135, 936–944. [Google Scholar] [CrossRef]
- Ratledge, C. Fatty acid biosynthesis in microorganisms being used for Single Cell Oil production. Biochimie 2004, 86, 807–815. [Google Scholar] [CrossRef]
- Patel, A.; Matsakas, L.; Hrůzová, K.; Rova, U.; Christakopoulos, P. Biosynthesis of Nutraceutical Fatty Acids by the Oleaginous Marine Microalgae Phaeodactylum tricornutum Utilizing Hydrolysates from Organosolv-Pretreated Birch and Spruce Biomass. Mar. Drugs 2019, 17, 119. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.; Matsakas, L.; Rova, U.; Christakopoulos, P. Heterotrophic cultivation of Auxenochlorella protothecoides using forest biomass as a feedstock for sustainable biodiesel production. Biotechnol. Biofuels 2018, 11, 169. [Google Scholar] [CrossRef]
- Patel, A.; Liefeldt, S.; Rova, U.; Christakopoulos, P.; Matsakas, L. Co-production of DHA and squalene by thraustochytrid from forest biomass. Sci. Rep. 2020, 10, 1992. [Google Scholar] [CrossRef]
- Patel, A.; Pruthi, V.; Singh, R.P.; Pruthi, P.A. Synergistic effect of fermentable and non-fermentable carbon sources enhances TAG accumulation in oleaginous yeast Rhodosporidium kratochvilovae HIMPA1. Bioresour. Technol. 2015, 188, 136–144. [Google Scholar] [CrossRef]
- Patel, A.; Mikes, F.; Bühler, S.; Matsakas, L. Valorization of brewers’ spent grain for the production of lipids by oleaginous yeast. Molecules 2018, 23, 3052. [Google Scholar] [CrossRef] [Green Version]
- Van Wychen, S.; Ramirez, K.; Laurens, L.M. Determination of Total Lipids as Fatty Acid Methyl Esters (FAME) by In Situ Transesterification (NREL/TP-5100-60958); Laboratory Analytical Procedure (LAP), National Renewable Energy Laboratory: Golden, CO, USA, 2013; Volume 303.
- Patel, A.; Rova, U.; Christakopoulos, P.; Matsakas, L. Simultaneous production of DHA and squalene from Aurantiochytrium sp. grown on forest biomass hydrolysates. Biotechnol. Biofuels 2019, 12, 255. [Google Scholar] [CrossRef]
- Patel, A.; Matsakas, L. A comparative study on de novo and ex novo lipid fermentation by oleaginous yeast using glucose and sonicated waste cooking oil. Ultrason. Sonochem. 2019, 52, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Wu, Q. High yield bio-oil production from fast pyrolysis by metabolic controlling of Chlorella protothecoides. J. Biotechnol. 2004, 110, 85–93. [Google Scholar] [CrossRef]
- Xu, H.; Miao, X.; Wu, Q. High quality biodiesel production from a microalga Chlorella protothecoides by heterotrophic growth in fermenters. J. Biotechnol. 2006, 126, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.K.A.K.; Sahoo, P.K.P.K.; Singhal, S.; Patel, A. Impact of various media and organic carbon sources on biofuel production potential from Chlorella spp. 3 Biotech 2016, 6, 116. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Slininger, P.J.; Dien, B.S.; Waghmode, S.; Moser, B.R.; Orjuela, A.; Sousa, L.D.C.; Balan, V. Microbial lipid-based lignocellulosic biorefinery: Feasibility and challenges. Trends Biotechnol. 2015, 33, 43–54. [Google Scholar] [CrossRef]
- Barclay, W.R.; Meager, K.M.; Abril, J.R. Heterotrophic production of long chain omega-3 fatty acids utilizing algae and algae-like microorganisms. J. Appl. Phycol. 1994, 6, 123–129. [Google Scholar] [CrossRef]
- Erhan, S.Z.; Sharma, B.K.; Liu, Z.; Adhvaryu, A. Lubricant base stock potential of chemically modified vegetable oils. J. Agric. Food Chem. 2008, 56, 8919–8925. [Google Scholar] [CrossRef] [PubMed]
- Mosarof, M.H.; Kalam, M.A.; Masjuki, H.H.; Alabdulkarem, A.; Ashraful, A.M.; Arslan, A.; Rashedul, H.K.; Monirul, I.M. Optimization of performance, emission, friction and wear characteristics of palm and Calophyllum inophyllum biodiesel blends. Energy Convers. Manag. 2016, 118, 119–134. [Google Scholar] [CrossRef]
- Jayadas, N.H.; Nair, K.P. Coconut oil as base oil for industrial lubricants-evaluation and modification of thermal, oxidative and low temperature properties. Tribol. Int. 2006, 39, 873–878. [Google Scholar] [CrossRef]
- Cermak, S.C.; Biresaw, G.; Isbell, T.A.; Evangelista, R.L.; Vaughn, S.F.; Murray, R. New crop oils-Properties as potential lubricants. Ind. Crops Prod. 2013, 44, 232–239. [Google Scholar] [CrossRef]
- Habibullah, M.; Masjuki, H.H.; Kalam, M.A.; Zulkifli, N.W.M.; Masum, B.M.; Arslan, A.; Gulzar, M. Friction and wear characteristics of Calophyllum inophyllum biodiesel. Ind. Crops Prod. 2015, 76, 188–197. [Google Scholar] [CrossRef]
- Fox, N.J.; Tyrer, B.; Stachowiak, G.W. Boundary lubrication performance of free fatty acids in sunflower oil. Tribol. Lett. 2004, 16, 275–281. [Google Scholar] [CrossRef]
- Saboya, R.M.A.; Cecilia, J.A.; García-Sancho, C.; Sales, A.V.; de Luna, F.M.T.; Rodríguez-Castellón, E.; Cavalcante, C.L. Assessment of commercial resins in the biolubricants production from free fatty acids of castor oil. Catal. Today 2017, 279, 274–285. [Google Scholar] [CrossRef]
- Saboya, R.M.A.; Cecilia, J.A.; García-Sancho, C.; Luna, F.M.T.; Rodríguez-Castellón, E.; Cavalcante, C.L. WO3-based catalysts supported on porous clay heterostructures (PCH) with Si-Zr pillars for synthetic esters production. Appl. Clay Sci. 2016, 124–125, 69–78. [Google Scholar] [CrossRef]
- Kalam, M.A.; Masjuki, H.H.; Shahabuddin, M.; Mofijur, M. Tribological characteristics of amine phosphate and octylated/butylated diphenylamine additives infused bio-lubricant. Energy Educ. Sci. Technol. Part A Energy Sci. Res. 2012, 30, 123–136. [Google Scholar]
- Slatter, T.; Lewis, P.R. Thesis Investigation into Tribological Performance of Vegetable Oils as Biolubricants at Severe Contact Conditions. Ph.D. Theis, University of Sheffield, Sheffield, UK, 2017. [Google Scholar]
- Marx, N.; Fernández, L.; Barceló, F.; Spikes, H. Shear Thinning and Hydrodynamic Friction of Viscosity Modifier-Containing Oils. Part I: Shear Thinning Behaviour. Tribol. Lett. 2018, 66, 92. [Google Scholar] [CrossRef] [Green Version]
- Ashrafi, N. Effects of temperature on rheology of olive oils. Appl. Rheol. 2012, 22, 342031–342037. [Google Scholar] [CrossRef]
- Garcia, C.C.; Franco, P.I.B.M.; Zuppa, T.O.; Filho, N.R.A.; Leles, M.I.G. Thermal stability studies of some cerrado plant oils. J. Therm. Anal. Calorim. 2007, 87, 645–648. [Google Scholar] [CrossRef]
- Santos, J.C.O.; Lima, L.N.; Santos, I.M.G.; Souza, A.G. Thermal, spectroscopic and rheological study of mineral base lubricating oils. J. Therm. Anal. Calorim. 2007, 87, 639–643. [Google Scholar] [CrossRef]
- Dasari, S.R.; Goud, V.V. Effect of pre-treatment on solvents extraction and physico-chemical properties of castor seed oil. J. Renew. Sustain. Energy 2014, 6, 063108. [Google Scholar] [CrossRef]
- Garcia-Perez, M.; Adams, T.T.; Goodrum, J.W.; Geller, D.; Das, K.C. Production and fuel properties of pine chip bio-oil/biodiesel blends. Energy Fuels 2007, 21, 2363–2372. [Google Scholar] [CrossRef]
- Mosarof, M.H.; Kalam, M.A.; Masjuki, H.H.; Arslan, A.; Monirul, I.M.; Ruhul, A.M.; Shahir, S.A.; Khuong, L.S. Analysis of thermal stability and lubrication characteristics of: Millettia pinnata oil. RSC Adv. 2016, 6, 81414–81425. [Google Scholar] [CrossRef]
- Luo, S.; Feng, J.; Ng, K.M. Effect of fatty acid on the formation of ITO nanocrystals via one-pot pyrolysis reaction. CrystEngComm 2015, 17, 1168–1172. [Google Scholar] [CrossRef]
- Shi, Y.; Larsson, R. Non-corrosive and Biomaterials Protic Ionic Liquids with High Lubricating Performance. Tribol. Lett. 2016, 63, 1. [Google Scholar] [CrossRef]
- Wu, Y.; Li, W.; Zhang, M.; Wang, X. Oxidative degradation of synthetic ester and its influence on tribological behavior. Tribol. Int. 2013, 64, 16–23. [Google Scholar] [CrossRef]
- Jayadas, N.H.; Nair, K.P. Study of the anti-wear properties of coconut oil using quantum chemical calculations and tribological tests. J. Tribol. 2006, 128, 654–659. [Google Scholar] [CrossRef]
- Hu, J.Q.; Wei, X.Y.; Dai, G.L.; Fei, Y.W.; Xie, F.; Zong, Z.M. Tribological behaviors and mechanism of sulfur- and phosphorus-free organic molybdate ester with zinc dialkyldithiophosphate. Tribol. Int. 2008, 41, 549–555. [Google Scholar] [CrossRef]
- Zeng, X.; Wu, H.; Yi, H.; Ren, T. Tribological behavior of three novel triazine derivatives as additives in rapeseed oil. Wear 2007, 262, 718–726. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patel, A.; Mu, L.; Shi, Y.; Rova, U.; Christakopoulos, P.; Matsakas, L. Single-Cell Oils from Oleaginous Microorganisms as Green Bio-Lubricants: Studies on Their Tribological Performance. Energies 2021, 14, 6685. https://doi.org/10.3390/en14206685
Patel A, Mu L, Shi Y, Rova U, Christakopoulos P, Matsakas L. Single-Cell Oils from Oleaginous Microorganisms as Green Bio-Lubricants: Studies on Their Tribological Performance. Energies. 2021; 14(20):6685. https://doi.org/10.3390/en14206685
Chicago/Turabian StylePatel, Alok, Liwen Mu, Yijun Shi, Ulrika Rova, Paul Christakopoulos, and Leonidas Matsakas. 2021. "Single-Cell Oils from Oleaginous Microorganisms as Green Bio-Lubricants: Studies on Their Tribological Performance" Energies 14, no. 20: 6685. https://doi.org/10.3390/en14206685
APA StylePatel, A., Mu, L., Shi, Y., Rova, U., Christakopoulos, P., & Matsakas, L. (2021). Single-Cell Oils from Oleaginous Microorganisms as Green Bio-Lubricants: Studies on Their Tribological Performance. Energies, 14(20), 6685. https://doi.org/10.3390/en14206685