

Circadian Rhythms of Body Temperature and Locomotor Activity in Spontaneously Hypertensive Rats under Frequent Changes in Light Conditions

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Environmental Conditions
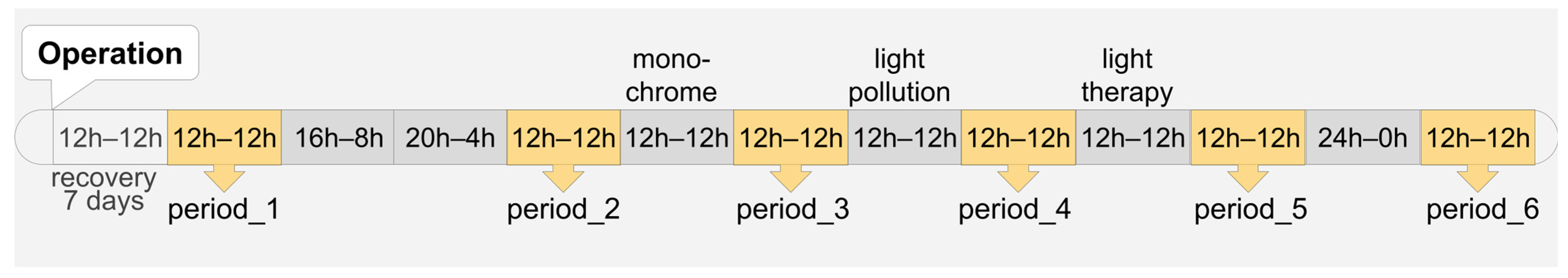
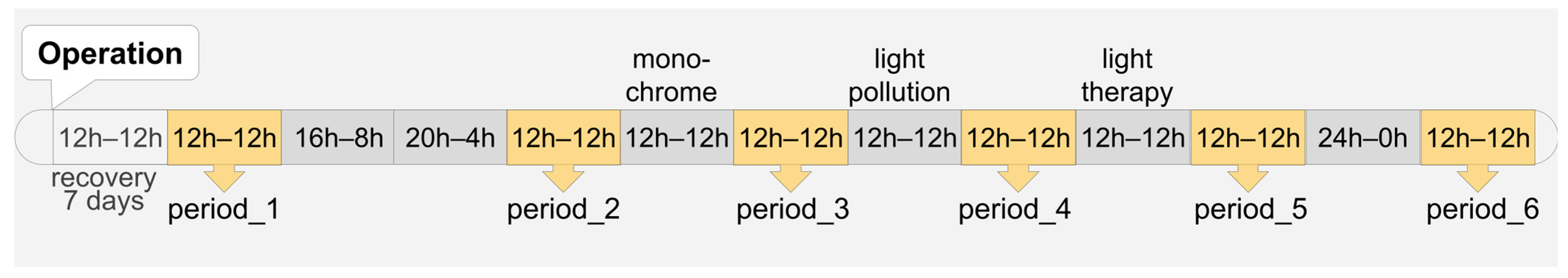
2.2. Study Design
2.3. Core Body Temperature and Activity Telemetry
2.4. Data Analysis
2.4.1. Linear Data Analysis
2.4.2. Circadian Rhythm Assessment
- Mesor (midline estimating statistic of rhythm)—the average level of the data in the 24 h period, calculated as a sum of minimum value and a half of a difference between maximum and minimum parameter value in the 24 h period;
- Amplitude—maximum deviation of the corresponding indicator from the mesor, calculated as a half of a difference between maximum and minimum parameter value in the 24 h period;
- Acrophase—a measure of the time of the general high values of the function repeated in each cycle, expressed in (negative) degrees with respect to a reference time set at 0°, with a period corresponding to 360° (2π or 6.283185 radian); in multicomponent analysis, it is defined as the first peak of the cycle;
- Ortophase (from lat. orto—rise)—phase of the cycle with the maximum value of the function (maximum peak), used in multicomponent analysis; may be the same value as acrophase.
2.4.3. Statistics
2.4.4. Software
3. Results
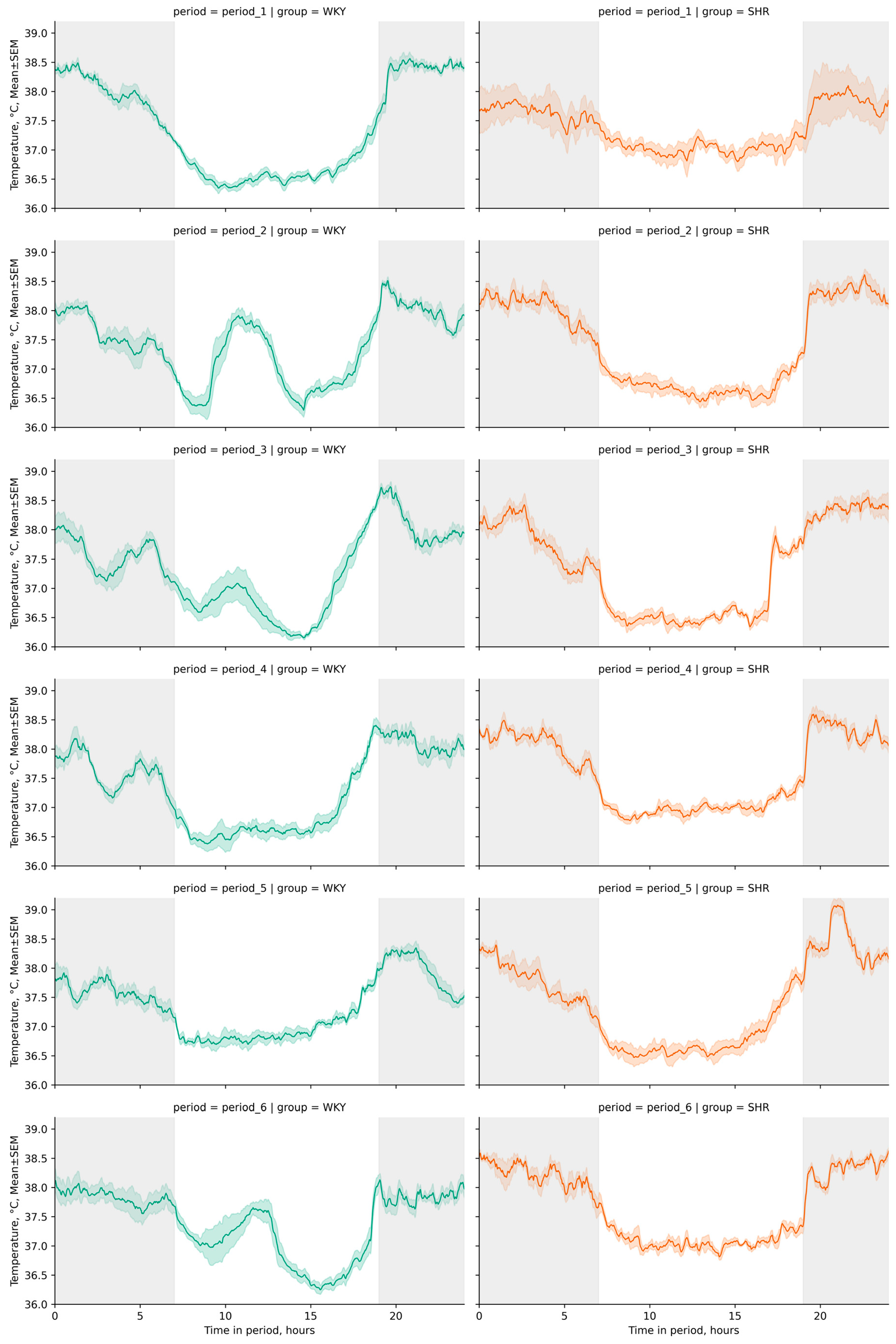
3.1. Tcore Telemetry
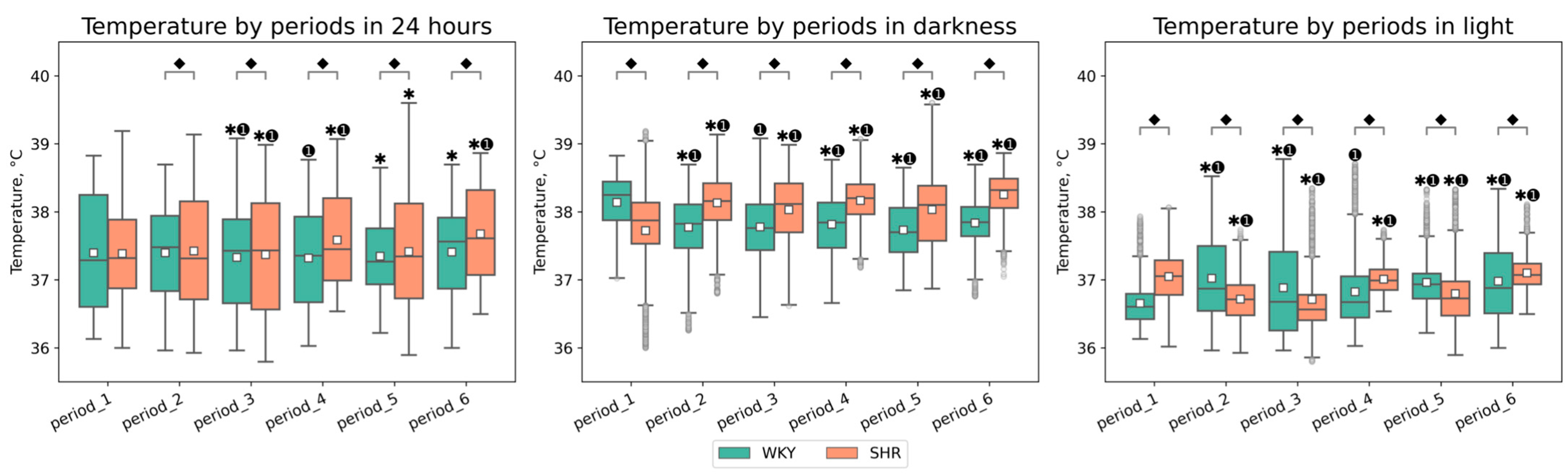
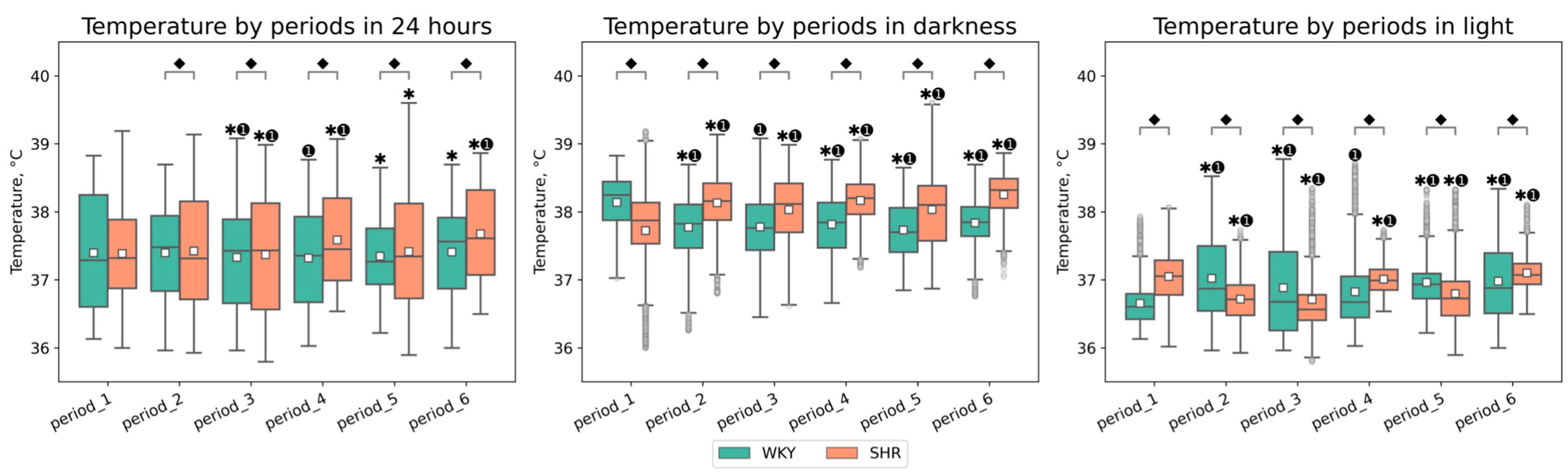
3.1.1. General Analysis of Tcore Data
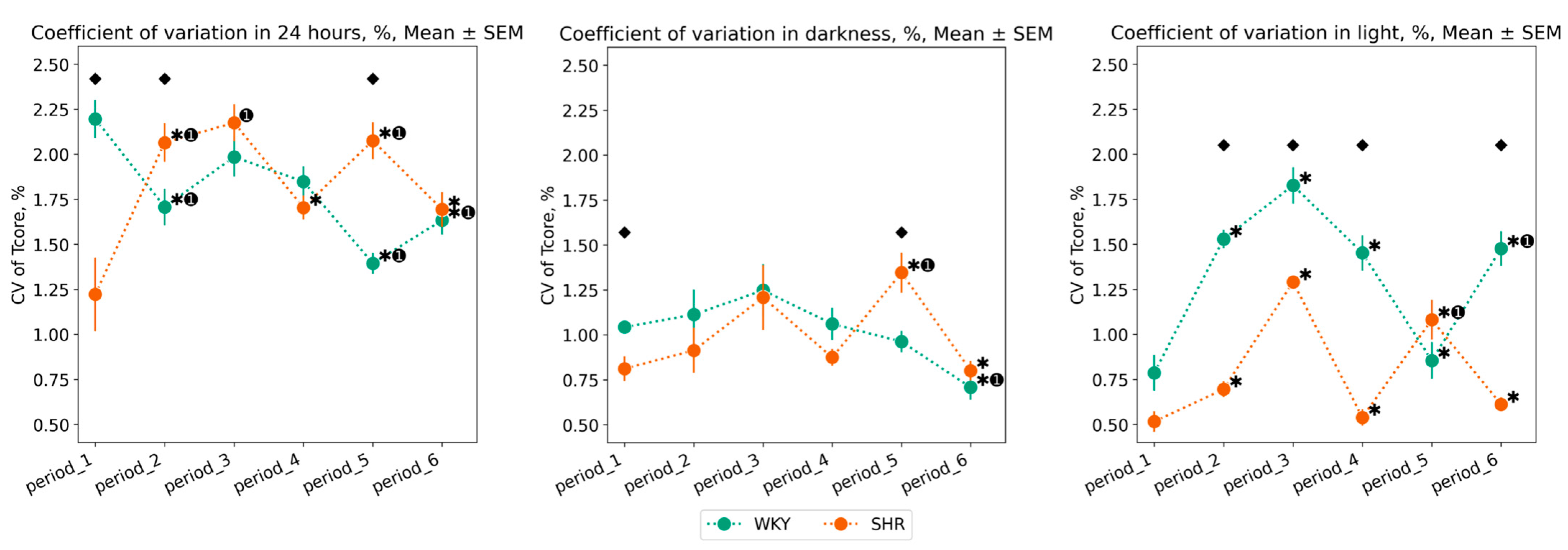
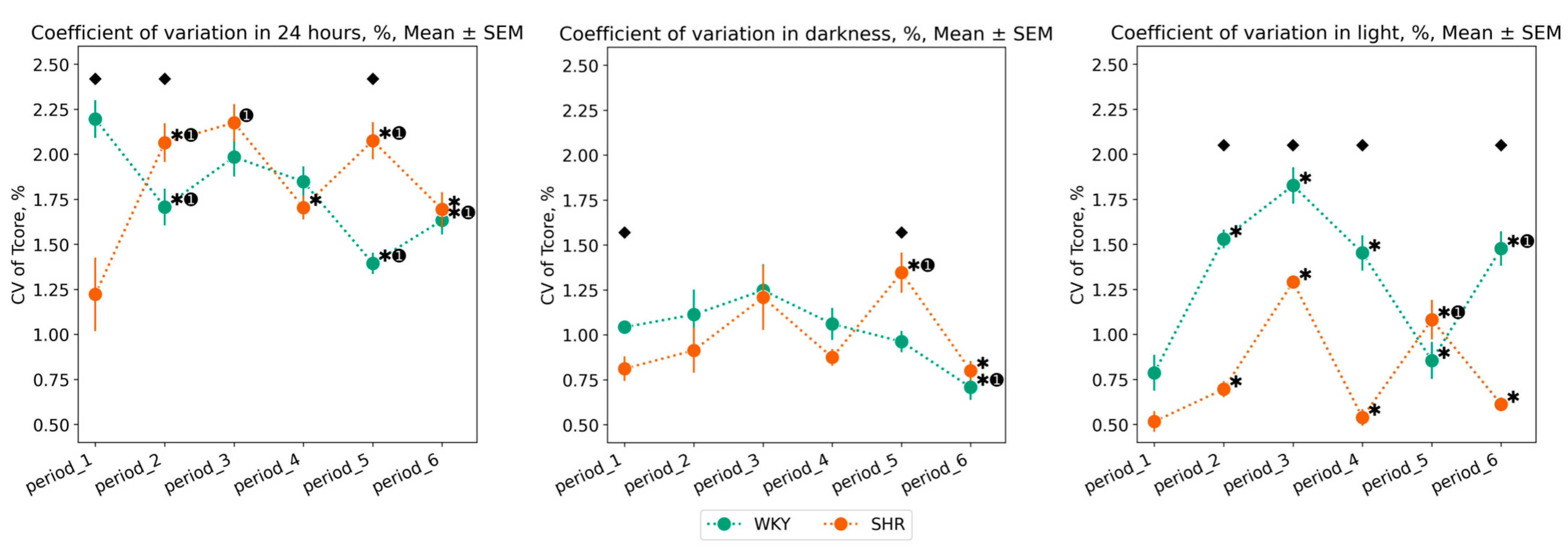
3.1.2. Variation of Tcore Data
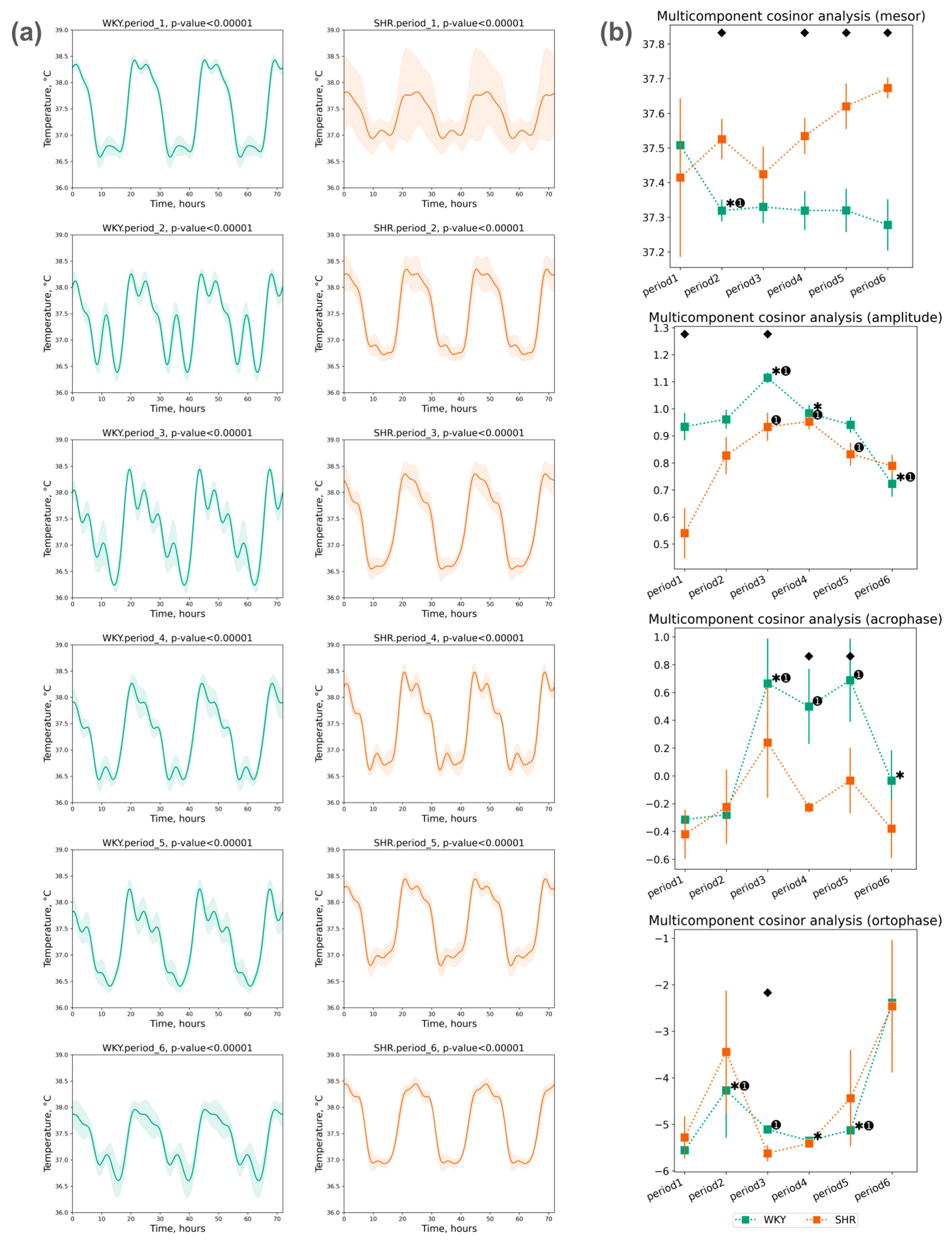
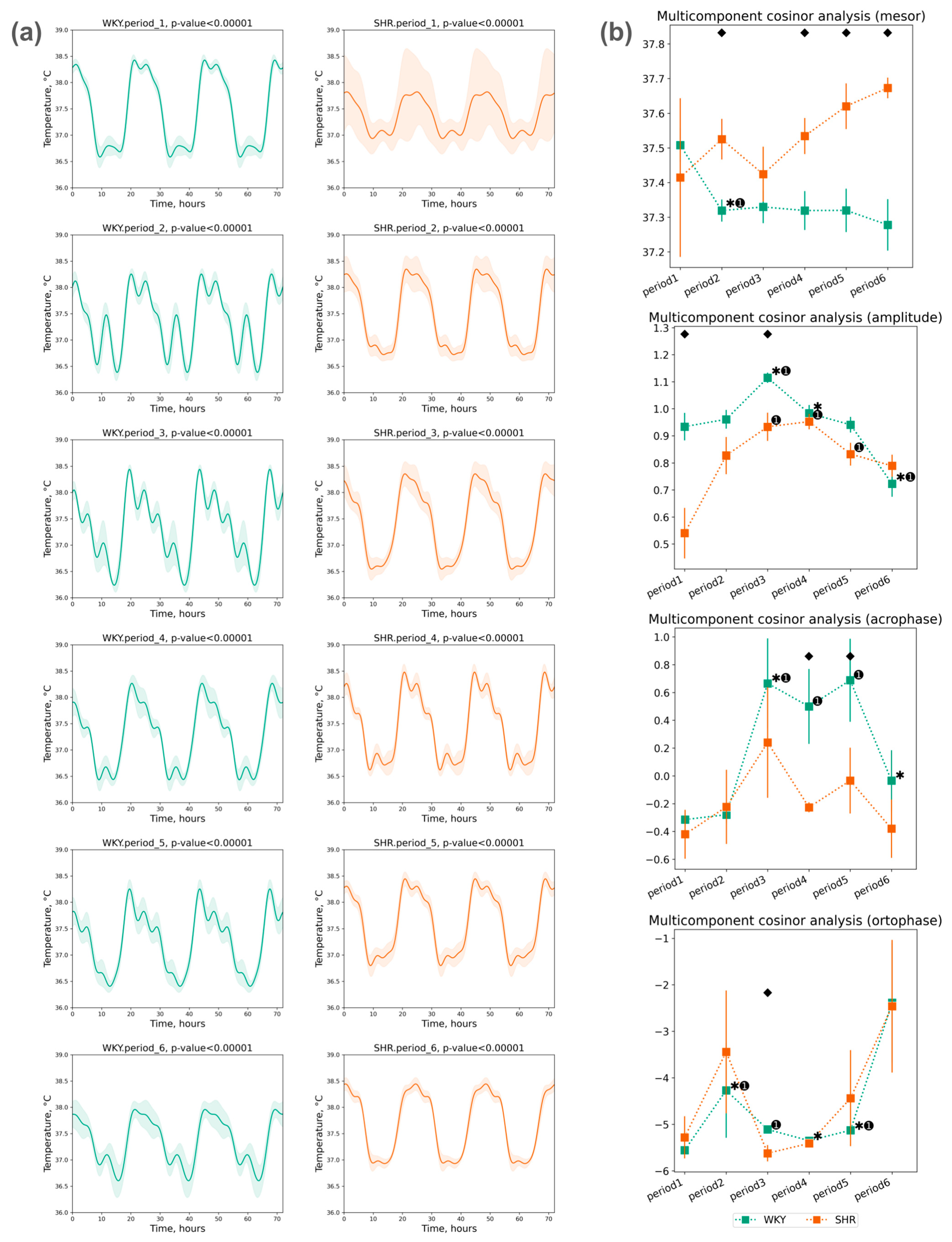
3.1.3. Cosinor Analysis of Tcore Data
- The acrophase shift of Tcore was noted during the third, fourth, and fifth periods compared to the initial (first) period in WKY, while no significant acrophase shift was observed in SHR;
- The ortophase shift of Tcore was also noted only in WKY during the second to fifth periods (Figure 5b).
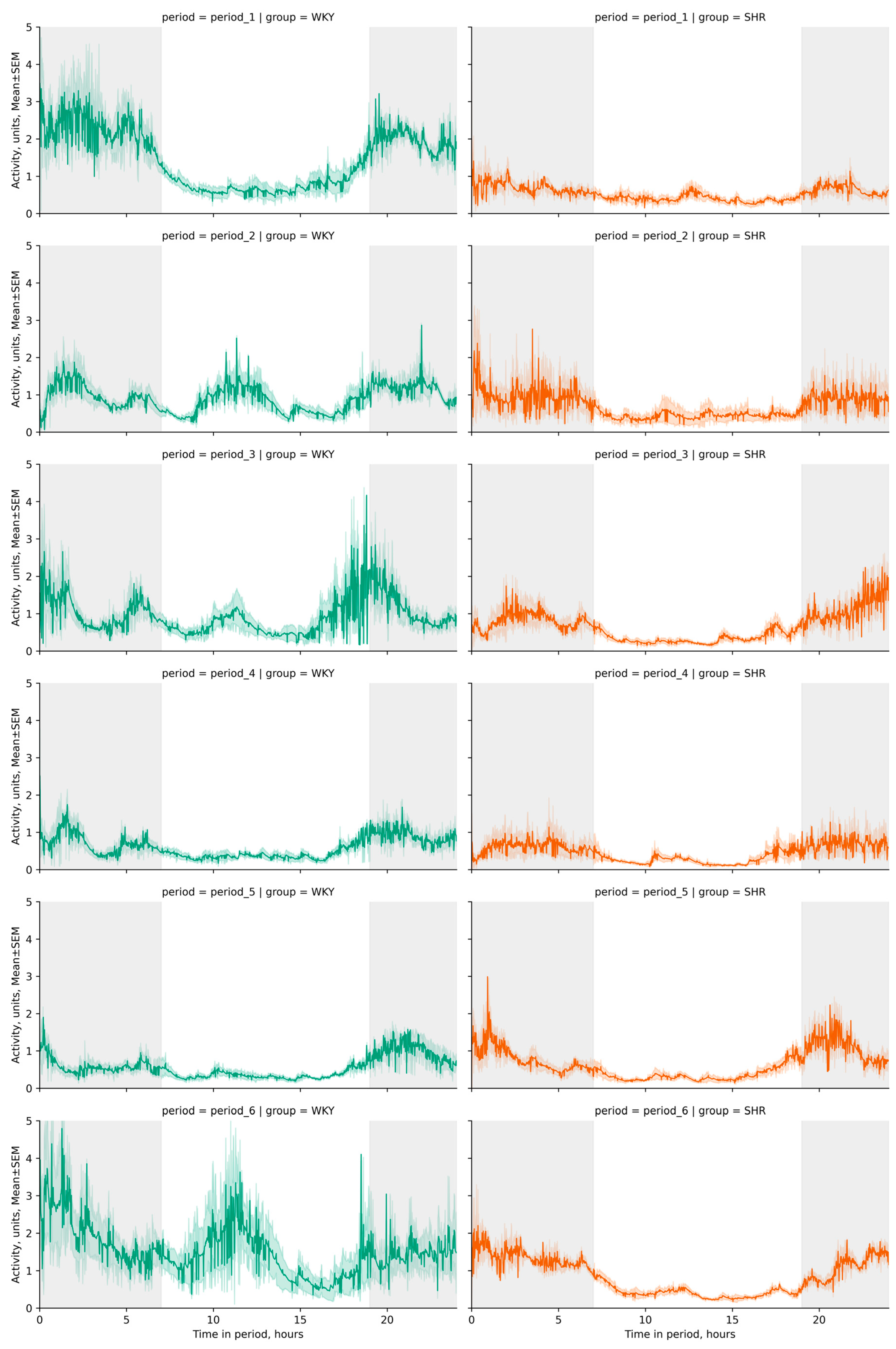
3.2. Activity Telemetry
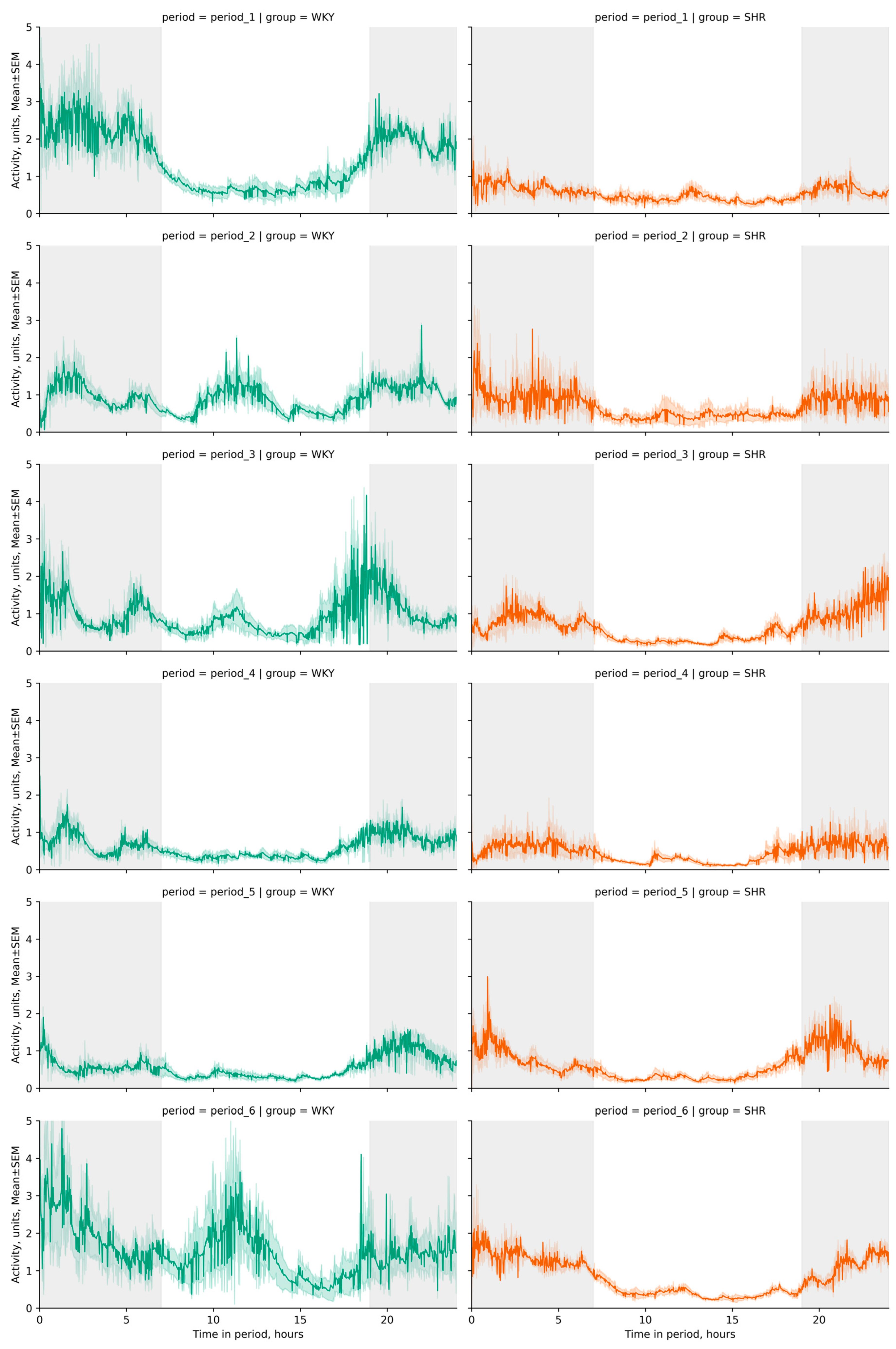
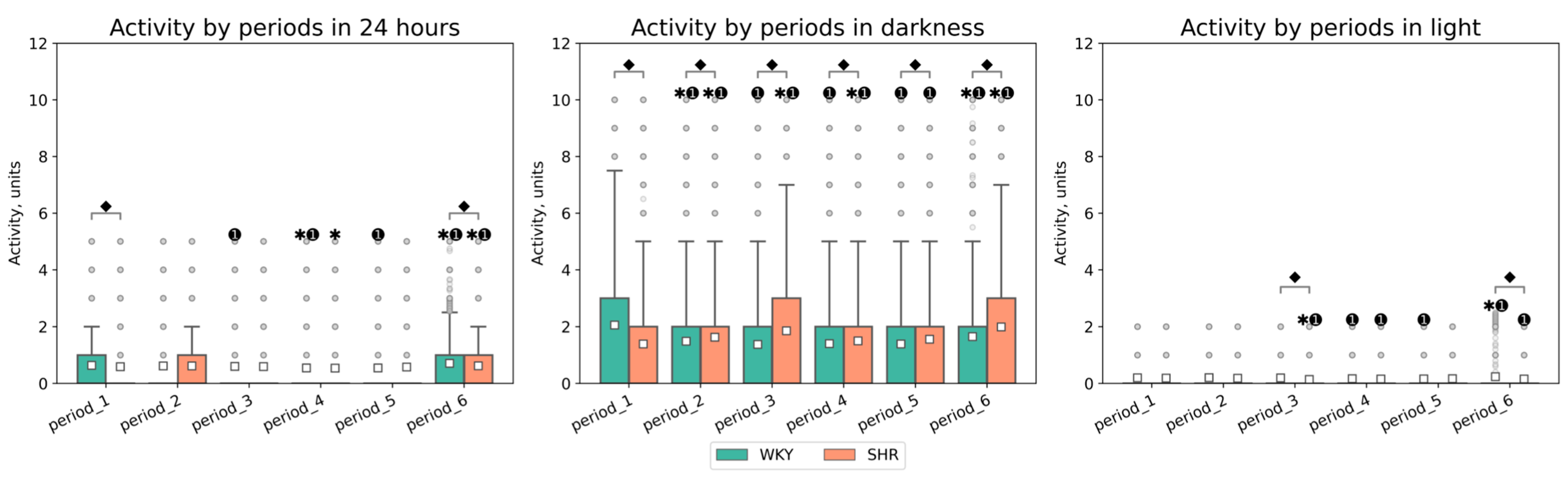
3.2.1. General Analysis of Activity Data
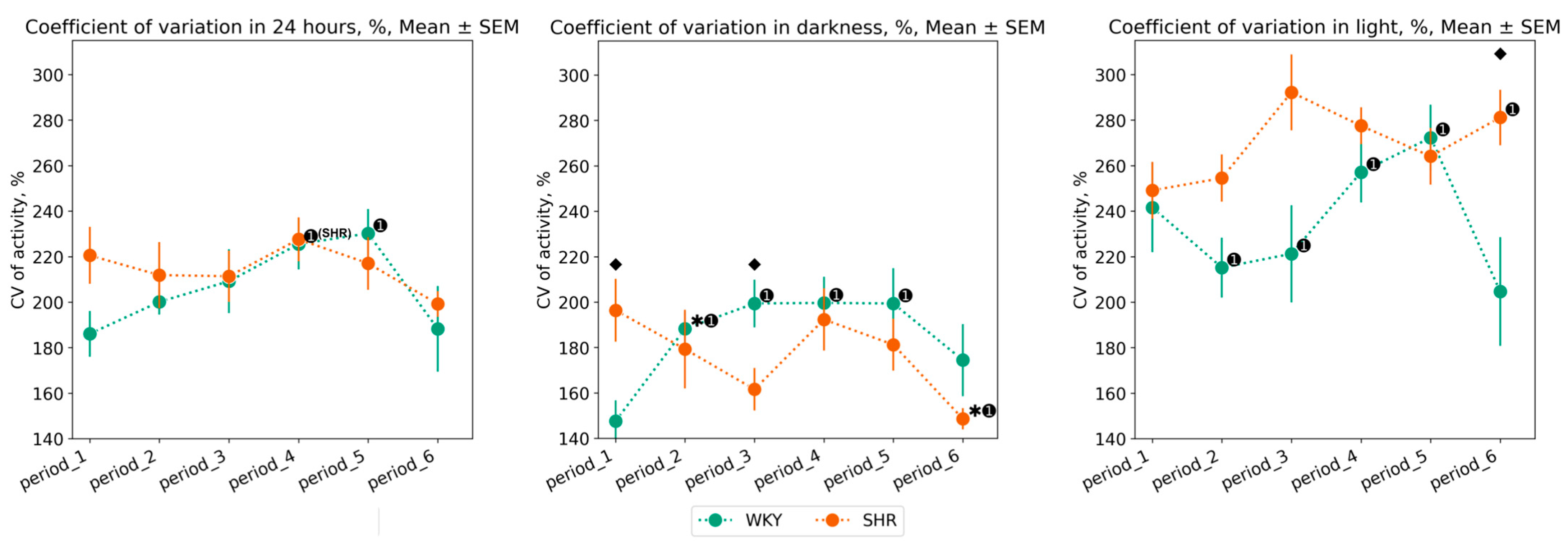
3.2.2. Variation of Activity Data
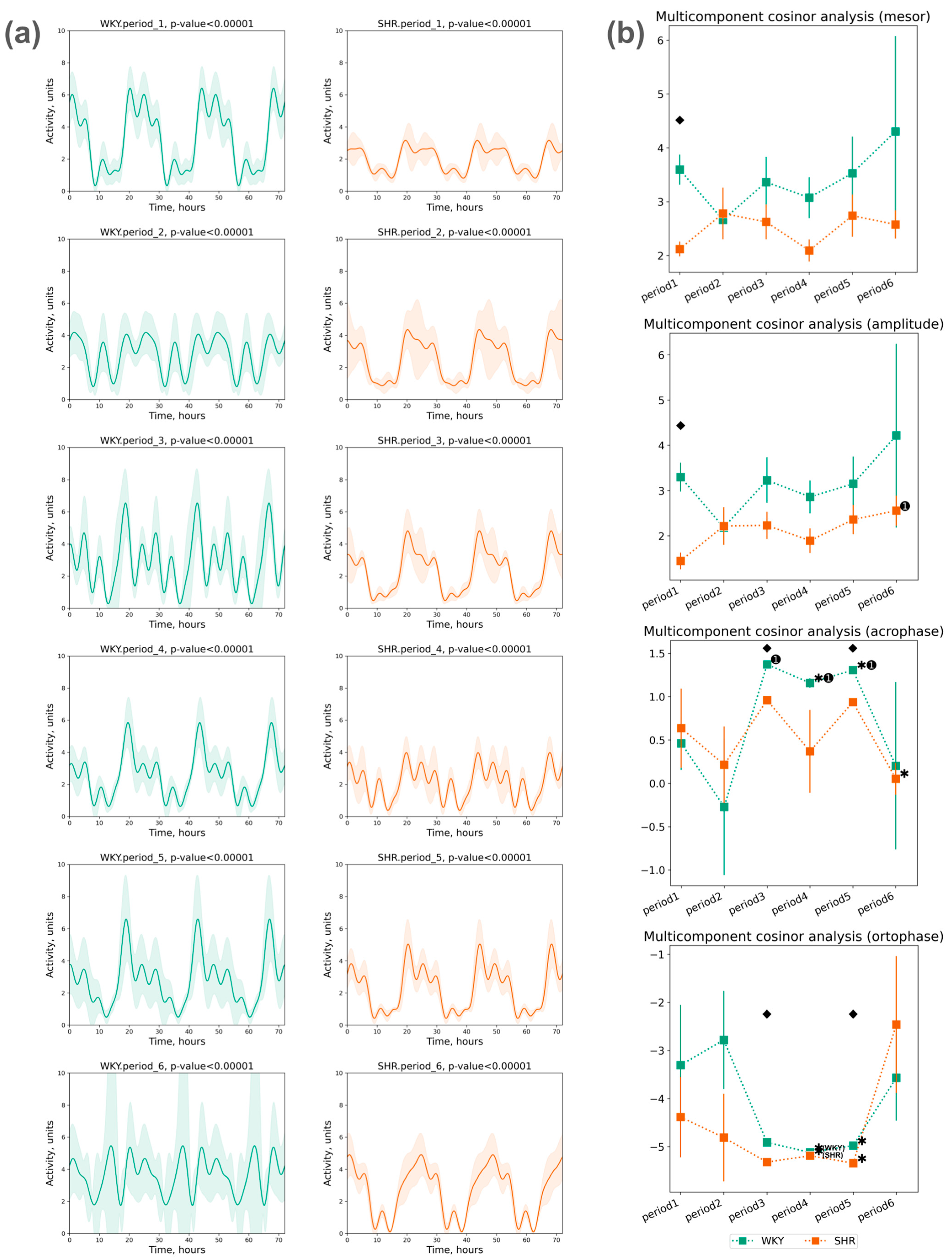
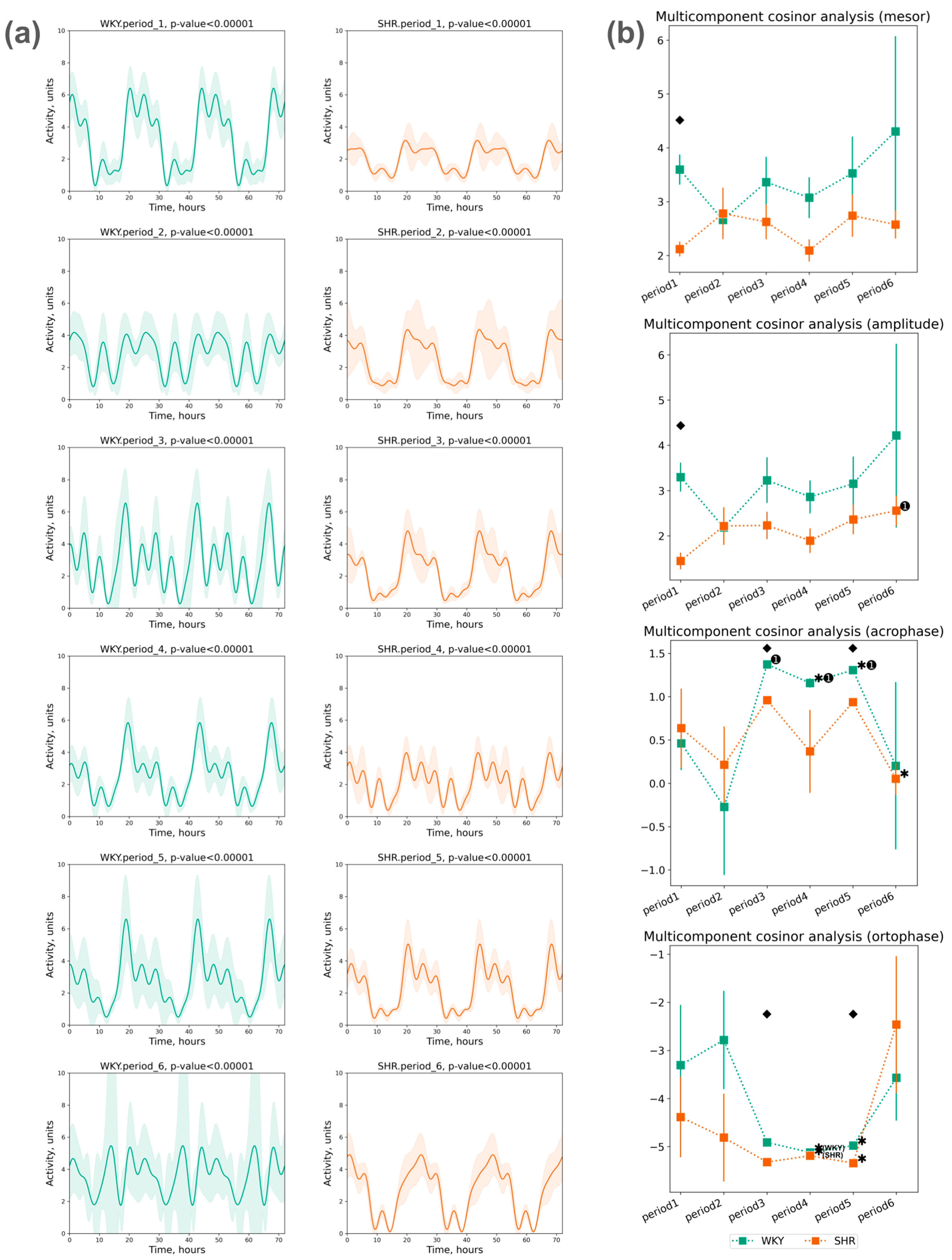
3.2.3. Cosinor Analysis of Activity Data
- The acrophase shift of activity was noted during the third, fourth, and fifth periods compared to the initial (first) period in WKY, while no significant acrophase shift was observed in SHR, except for a deviation in the last period compared to the previous one;
- The ortophase shift of activity was noted in WKY and SHR during the fourth and fifth periods compared to previous periods (Figure 9b).
3.3. Correlation of Tcore and Activity Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fagiani, F.; Di Marino, D.; Romagnoli, A.; Travelli, C.; Voltan, D.; Di Cesare Mannelli, L.; Racchi, M.; Govoni, S.; Lanni, C. Molecular Regulations of Circadian Rhythm and Implications for Physiology and Diseases. Signal Transduct. Target. Ther. 2022, 7, 41. [Google Scholar] [CrossRef]
- Neves, A.R.; Albuquerque, T.; Quintela, T.; Costa, D. Circadian Rhythm and Disease: Relationship, New Insights, and Future Perspectives. J. Cell. Physiol. 2022, 237, 3239–3256. [Google Scholar] [CrossRef]
- Wu, T.; Jin, Y.; Kato, H.; Fu, Z. Light and Food Signals Cooperate to Entrain the Rat Pineal Circadian System. J. Neurosci. Res. 2008, 86, 3246–3255. [Google Scholar] [CrossRef]
- Cambras, T.; Weller, J.R.; Anglès-Pujoràs, M.; Lee, M.L.; Christopher, A.; Díez-Noguera, A.; Krueger, J.M.; de la Iglesia, H.O. Circadian Desynchronization of Core Body Temperature and Sleep Stages in the Rat. Proc. Natl. Acad. Sci. USA 2007, 104, 7634–7639. [Google Scholar] [CrossRef]
- Refinetti, R. Metabolic Heat Production, Heat Loss and the Circadian Rhythm of Body Temperature in the Rat. Exp. Physiol. 2003, 88, 423–429. [Google Scholar] [CrossRef]
- Daan, S.; Honma, S.; Honma, K. Body Temperature Predicts the Direction of Internal Desynchronization in Humans Isolated from Time Cues. J. Biol. Rhythm. 2013, 28, 403–411. [Google Scholar] [CrossRef]
- Coiffard, B.; Diallo, A.B.; Mezouar, S.; Leone, M.; Mege, J.-L. A Tangled Threesome: Circadian Rhythm, Body Temperature Variations, and the Immune System. Biology 2021, 10, 65. [Google Scholar] [CrossRef]
- Refinetti, R. Circadian Rhythmicity of Body Temperature and Metabolism. Temperature 2020, 7, 321–362. [Google Scholar] [CrossRef]
- de Rezende, L.M.T.; Brito, L.C.; Moura, A.G.; Costa, A.J.L.D.; Leal, T.F.; Favarato, E.S.; Favarato, L.S.C.; Natali, A.J.; Coimbra, C.C.; Prímola-Gomes, T.N. Core Temperature Circadian Rhythm across Aging in Spontaneously Hypertensive Rats. J. Therm. Biol. 2021, 97, 102807. [Google Scholar] [CrossRef]
- Benstaali, C.; Mailloux, A.; Bogdan, A.; Auzéby, A.; Touitou, Y. Circadian Rhythms of Body Temperature and Motor Activity in Rodents Their Relationships with the Light-Dark Cycle. Life Sci. 2001, 68, 2645–2656. [Google Scholar] [CrossRef]
- Cesarovic, N.; Jirkof, P.; Rettich, A.; Arras, M. Implantation of radiotelemetry transmitters yielding data on ECG, heart rate, core body temperature and activity in free-moving laboratory mice. J. Vis. Exp. JoVE 2011, 57, 3260. [Google Scholar] [CrossRef]
- Bishop, B.; Silva, G.; Krasney, J.; Nakano, H.; Roberts, A.; Farkas, G.; Rifkin, D.; Shucard, D. Ambient Temperature Modulates Hypoxic-Induced Changes in Rat Body Temperature and Activity Differentially. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R1190–R1196. [Google Scholar] [CrossRef]
- Tsushima, H.; Yamada, K. Effects of Adipokine Administration to the Hypothalamic Preoptic Area on Body Temperature in Rats. J. Pharmacol. Sci. 2020, 144, 61–68. [Google Scholar] [CrossRef]
- Calonne, J.; Arsenijevic, D.; Scerri, I.; Miles-Chan, J.L.; Montani, J.-P.; Dulloo, A.G. Low 24-Hour Core Body Temperature as a Thrifty Metabolic Trait Driving Catch-up Fat during Weight Regain after Caloric Restriction. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E699–E709. [Google Scholar] [CrossRef]
- Teixeira-Coelho, F.; Fonseca, C.G.; Vaz, F.F.; Barbosa, N.H.S.; Soares, D.D.; Pires, W.; Wanner, S.P. Physical Exercise-Induced Thermoregulatory Responses in Trained Rats: Effects of Manipulating the Duration and Intensity of Aerobic Training Sessions. J. Therm. Biol. 2021, 97, 102878. [Google Scholar] [CrossRef]
- Rodovalho, G.V.; Drummond, L.R.; Coimbra, C.C. Involvement of Brainstem Noradrenergic System in Cutaneous Heat Loss during Exercise. Brain Res. Bull. 2020, 164, 372–379. [Google Scholar] [CrossRef]
- Machado, F.S.M.; Fóscolo, D.R.C.; Poletini, M.O.; Coimbra, C.C. Influence of Time-of-Day on Maximal Exercise Capacity Is Related to Daily Thermal Balance but Not to Induced Neuronal Activity in Rats. Front. Physiol. 2016, 7, 464. [Google Scholar] [CrossRef]
- Thompson, R.S.; Christianson, J.P.; Maslanik, T.M.; Maier, S.F.; Greenwood, B.N.; Fleshner, M. Effects of Stressor Controllability on Diurnal Physiological Rhythms. Physiol. Behav. 2013, 112, 32–39. [Google Scholar] [CrossRef]
- Schwimmer, H.; Mursu, N.; Haim, A. Effects of Light and Melatonin Treatment on Body Temperature and Melatonin Secretion Daily Rhythms in a Diurnal Rodent, the Fat Sand Rat. Chronobiol. Int. 2010, 27, 1401–1419. [Google Scholar] [CrossRef]
- Huang, S.; Jiao, X.; Lu, D.; Pei, X.; Qi, D.; Li, Z. Recent advances in modulators of circadian rhythms: An update and perspective. J. Enzym. Inhib. Med. Chem. 2020, 35, 1267–1286. [Google Scholar] [CrossRef]
- Deprés-Brummer, P.; Lévi, F.; Metzger, G.; Touitou, Y. Light-Induced Suppression of the Rat Circadian System. Am. J. Physiol. 1995, 268, R1111–R1116. [Google Scholar] [CrossRef]
- Honma, S.; Kanematsu, N.; Katsuno, Y.; Honma, K. Persistence of Circadian Oscillation While Locomotor Activity and Plasma Melatonin Levels Became Aperiodic under Prolonged Continuous Light in the Rat. Neurosci. Lett. 1996, 216, 49–52. [Google Scholar] [CrossRef]
- Sauvet, F.; Gomez-Merino, D.; Dorey, R.; Ciret, S.; Gallopin, T.; Drogou, C.; Arnal, P.J.; Chennaoui, M. Lengthening of the Photoperiod Influences Sleep Characteristics before and during Total Sleep Deprivation in Rat. J. Sleep Res. 2019, 28, e12709. [Google Scholar] [CrossRef] [PubMed]
- Rumanova, V.S.; Okuliarova, M.; Foppen, E.; Kalsbeek, A.; Zeman, M. Exposure to Dim Light at Night Alters Daily Rhythms of Glucose and Lipid Metabolism in Rats. Front. Physiol. 2022, 13, 973461. [Google Scholar] [CrossRef]
- Masís-Vargas, A.; Ritsema, W.I.G.R.; Mendoza, J.; Kalsbeek, A. Metabolic Effects of Light at Night Are Time- and Wavelength-Dependent in Rats. Obesity 2020, 28, S114–S125. [Google Scholar] [CrossRef]
- Guan, Q.; Li, Y.; Wang, Z.; Cao, J.; Dong, Y.; Ren, F.; Chen, Y. Monochromatic Light Pollution Exacerbates High-Fat Diet-Induced Adipocytic Hypertrophy in Mice. Cells 2022, 11, 3808. [Google Scholar] [CrossRef]
- Lin, H.; Wang, H.; Xu, Y.; Lin, Z.; Kang, D.; Zheng, S.; Yao, P. Lower Body Temperature Independently Predicts Delayed Cerebral Infarction in the Elderly With Ruptured Intracranial Aneurysm. Front. Neurol. 2022, 12, 763471. [Google Scholar] [CrossRef]
- Barnes, J.N.; Charkoudian, N. Integrative Cardiovascular Control in Women: Regulation of Blood Pressure, Body Temperature and Cerebrovascular Responsiveness. FASEB J. 2021, 35, e21143. [Google Scholar] [CrossRef] [PubMed]
- Sabino, J.P.J.; Soriano, R.N.; Donatti, A.F.; Fernandez, R.R.; Kwiatkoski, M.; Francescato, H.D.C.; Coimbra, T.M.; Branco, L.G.S. Involvement of Endogenous Central Hydrogen Sulfide (H2S) in Hypoxia-Induced Hypothermia in Spontaneously Hypertensive Rats. Can. J. Physiol. Pharmacol. 2017, 95, 157–162. [Google Scholar] [CrossRef]
- Anishchenko, A.M.; Aliev, O.I.; Sidekhmenova, A.V.; Shamanaev, A.Y.; Plotnikov, M.B. Dynamics of Blood Pressure Elevation and Endothelial Dysfunction in SHR Rats During the Development of Arterial Hypertension. Bull. Exp. Biol. Med. 2015, 159, 591–593. [Google Scholar] [CrossRef] [PubMed]
- Bingham, C.; Arbogast, B.; Guillaume, G.C.; Lee, J.K.; Halberg, F. Inferential Statistical Methods for Estimating and Comparing Cosinor Parameters. Chronobiologia 1982, 9, 397–439. [Google Scholar]
- Cornelissen, G. Cosinor-Based Rhythmometry. Theor. Biol. Med. Model. 2014, 11, 16. [Google Scholar] [CrossRef]
- Moškon, M. CosinorPy: A Python Package for Cosinor-Based Rhythmometry. BMC Bioinform. 2020, 21, 485. [Google Scholar] [CrossRef]
- Moškon, M. Identification and Characterisation of Variable Rhythms Using CosinorPy. bioRxiv 2022. [Google Scholar] [CrossRef]
- López-Canul, M.; Min, S.H.; Posa, L.; De Gregorio, D.; Bedini, A.; Spadoni, G.; Gobbi, G.; Comai, S. Melatonin MT1 and MT2 Receptors Exhibit Distinct Effects in the Modulation of Body Temperature across the Light/Dark Cycle. Int. J. Mol. Sci. 2019, 20, 2452. [Google Scholar] [CrossRef]
- Hankenson, F.C.; Marx, J.O.; Gordon, C.J.; David, J.M. Effects of Rodent Thermoregulation on Animal Models in the Research Environment. Comp. Med. 2018, 68, 425–438. [Google Scholar] [CrossRef]
- Campos, H.O.; Leite, L.H.R.; Drummond, L.R.; Cunha, D.N.Q.; Coimbra, C.C.; Natali, A.J.; Prímola-Gomes, T.N. Temperature Control of Hypertensive Rats during Moderate Exercise in Warm Environment. J. Sports Sci. Med. 2014, 13, 695–701. [Google Scholar] [PubMed]
- Drummond, L.R.; Campos, H.O.; de Andrade Lima, P.M.; da Fonseca, C.G.; Kunstetter, A.C.; Rodrigues, Q.T.; Szawka, R.E.; Natali, A.J.; Prímola-Gomes, T.N.; Wanner, S.P.; et al. Impaired thermoregulation in spontaneously hypertensive rats during physical exercise is related to reduced hypothalamic neuronal activation. Pflug. Arch. Eur. J. Physiol. 2020, 472, 1757–1768. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, J.; Drust, B.; Weinert, D.; Edwards, B.; Gregson, W.; Atkinson, G.; Kao, S.; Aizawa, S.; Reilly, T. The Circadian Rhythm of Core Temperature: Origin and Some Implications for Exercise Performance. Chronobiol. Int. 2005, 22, 207–225. [Google Scholar] [CrossRef] [PubMed]
- Jing, J.-N.; Wu, Z.-T.; Li, M.-L.; Wang, Y.-K.; Tan, X.; Wang, W.-Z. Constant Light Exerted Detrimental Cardiovascular Effects Through Sympathetic Hyperactivity in Normal and Heart Failure Rats. Front. Neurosci. 2020, 14, 248. [Google Scholar] [CrossRef] [PubMed]
- Kenny, G.P.; Yardley, J.; Brown, C.; Sigal, R.J.; Jay, O. Heat Stress in Older Individuals and Patients with Common Chronic Diseases. CMAJ 2010, 182, 1053–1060. [Google Scholar] [CrossRef] [PubMed]
- Montaruli, A.; Castelli, L.; Mulè, A.; Scurati, R.; Esposito, F.; Galasso, L.; Roveda, E. Biological Rhythm and Chronotype: New Perspectives in Health. Biomolecules 2021, 11, 487. [Google Scholar] [CrossRef] [PubMed]
- Gehrman, P.; Marler, M.; Martin, J.L.; Shochat, T.; Corey-Bloom, J.; Ancoli-Israel, S. The Timing of Activity Rhythms in Patients With Dementia Is Related to Survival. J. Gerontol. Ser. A 2004, 59, M1050–M1055. [Google Scholar] [CrossRef] [PubMed]
- Brooks, T.G.; Lahens, N.F.; Grant, G.R.; Sheline, Y.I.; FitzGerald, G.A.; Skarke, C. Diurnal Rhythms of Wrist Temperature Are Associated with Future Disease Risk in the UK Biobank. Nat. Commun. 2023, 14, 5172. [Google Scholar] [CrossRef]
- Conti, B.; Sanchez-Alavez, M.; Winsky-Sommerer, R.; Morale, M.C.; Lucero, J.; Brownell, S.; Fabre, V.; Huitron-Resendiz, S.; Henriksen, S.; Zorrilla, E.P.; et al. Transgenic Mice with a Reduced Core Body Temperature Have an Increased Life Span. Science 2006, 314, 825–828. [Google Scholar] [CrossRef]
- Beauchamp, M.T.; Lundgren, J.D. A Systematic Review of Bright Light Therapy for Eating Disorders. Prim. Care Companion CNS Disord. 2016, 18, 26718. [Google Scholar] [CrossRef]
- Fifel, K.; Videnovic, A. Light Therapy in Parkinson’s Disease: Towards Mechanism-Based Protocols. Trends Neurosci. 2018, 41, 252–254. [Google Scholar] [CrossRef]
- Penders, T.M.; Stanciu, C.N.; Schoemann, A.M.; Ninan, P.T.; Bloch, R.; Saeed, S.A. Bright Light Therapy as Augmentation of Pharmacotherapy for Treatment of Depression: A Systematic Review and Meta-Analysis. Prim. Care Companion CNS Disord. 2016, 18, 322–326. [Google Scholar] [CrossRef]
- Komarov, F.I.; Rapoport, S.I.; Breus, T.K.; Chibisov, S.M. Desynchronization of Biological Rhythms in Response to Environmental Factors. Klin. Med. 2017, 95, 502–512. [Google Scholar] [CrossRef]
- van Soest, I.; Del Olmo, M.; Schmal, C.; Herzel, H. Nonlinear phenomena in models of the circadian clock. J. R. Soc. Interface 2020, 17, 20200556. [Google Scholar] [CrossRef]
- Satoh, Y.; Kawai, H.; Kudo, N.; Kawashima, Y.; Mitsumoto, A. Temperature rhythm reentrains faster than locomotor rhythm after a light phase shift. Physiol. Behav. 2006, 88, 404–410. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Periods | Groups | Temperature in 24 h (Mean ± SEM, °C) | Temperature in Darkness (Mean ± SEM, °C) | Temperature in Light (Mean ± SEM, °C) |
|---|---|---|---|---|
| period_1 | WKY | 37.397 ± 0.010 | 38.140 ± 0.007 ⬥ | 36.655 ± 0.005 ⬥ |
| period_1 | SHR | 37.380 ± 0.007 | 37.717 ± 0.011 ⬥ | 37.044 ± 0.005 ⬥ |
| period_2 | WKY | 37.398 ± 0.008 ⬥ | 37.774 ± 0.007 ⬥✱➊ | 37.022 ± 0.010 ⬥✱➊ |
| period_2 | SHR | 37.423 ± 0.009 ⬥ | 38.132 ± 0.007 ⬥✱➊ | 36.715 ± 0.005 ⬥✱➊ |
| period_3 | WKY | 37.339 ± 0.009 ⬥✱➊ | 37.778 ± 0.008 ⬥➊ | 36.884 ± 0.011 ⬥✱➊ |
| period_3 | SHR | 37.371 ± 0.010 ⬥✱➊ | 38.031 ± 0.008 ⬥✱➊ | 36.711 ± 0.008 ⬥✱➊ |
| period_4 | WKY | 37.320 ± 0.008 ⬥➊ | 37.816 ± 0.007 ⬥✱➊ | 36.824 ± 0.009 ⬥➊ |
| period_4 | SHR | 37.587 ± 0.008 ⬥✱➊ | 38.165 ± 0.006 ⬥✱➊ | 37.010 ± 0.004 ⬥✱➊ |
| period_5 | WKY | 37.348 ± 0.006 ⬥✱ | 37.734 ± 0.007 ⬥✱➊ | 36.963 ± 0.006 ⬥✱➊ |
| period_5 | SHR | 37.415 ± 0.009 ⬥✱ | 38.033 ± 0.009 ⬥✱➊ | 36.798 ± 0.008 ⬥✱➊ |
| period_6 | WKY | 37.408 ± 0.008 ⬥✱ | 37.836 ± 0.006 ⬥✱➊ | 36.981 ± 0.010 ⬥✱➊ |
| period_6 | SHR | 37.677 ± 0.008 ⬥✱➊ | 38.255 ± 0.005 ⬥✱➊ | 37.101 ± 0.004 ⬥✱➊ |
| Periods | Groups | Activity in 24 h (Mean ± SEM, Units) | Activity in Darkness (Mean ± SEM, Units) | Activity in Light (Mean ± SEM, Units) |
|---|---|---|---|---|
| period_1 | WKY | 0.638 ± 0.017 ⬥ | 2.050 ± 0.055 ⬥ | 0.198 ± 0.009 |
| period_1 | SHR | 0.586 ± 0.016 ⬥ | 1.388 ± 0.043 ⬥ | 0.189 ± 0.009 |
| period_2 | WKY | 0.614 ± 0.016 | 1.484 ± 0.046 ⬥✱➊ | 0.203 ± 0.01 |
| period_2 | SHR | 0.615 ± 0.016 | 1.628 ± 0.046 ⬥✱➊ | 0.183 ± 0.009 |
| period_3 | WKY | 0.599 ± 0.016 ➊ | 1.372 ± 0.044 ⬥➊ | 0.190 ± 0.01 ⬥ |
| period_3 | SHR | 0.592 ± 0.016 | 1.853 ± 0.049 ⬥✱➊ | 0.140 ± 0.008 ⬥✱➊ |
| period_4 | WKY | 0.540 ± 0.015 ✱➊ | 1.400 ± 0.044 ⬥➊ | 0.162 ± 0.008 ➊ |
| period_4 | SHR | 0.539 ± 0.015 ✱ | 1.494 ± 0.044 ⬥✱➊ | 0.154 ± 0.008 ➊ |
| period_5 | WKY | 0.542 ± 0.015 | 1.388 ± 0.044 ⬥➊ | 0.159 ± 0.008 ➊ |
| period_5 | SHR | 0.578 ± 0.016 | 1.556 ± 0.045 ⬥➊ | 0.165 ± 0.008 |
| period_6 | WKY | 0.705 ± 0.017 ⬥✱➊ | 1.649 ± 0.048 ⬥✱➊ | 0.237 ± 0.011 ⬥✱➊ |
| period_6 | SHR | 0.619 ± 0.016 ⬥✱➊ | 1.988 ± 0.051 ⬥✱➊ | 0.155 ± 0.008 ⬥➊ |
| Day Phase | Periods | Groups | Spearman Test | p_Value |
|---|---|---|---|---|
| Darkness | period_1 | SHR | 0.02 | 0.40 |
| Darkness | period_1 | WKY | 0.13 | 6.14 × 10−13 |
| Darkness | period_2 | SHR | 0.18 | 1.80 × 10−21 |
| Darkness | period_2 | WKY | 0.24 | 2.93 × 10−42 |
| Darkness | period_3 | SHR | 0.23 | 6.25 × 10−34 |
| Darkness | period_3 | WKY | 0.18 | 2.32 × 10−22 |
| Darkness | period_4 | SHR | 0.18 | 6.43 × 10−21 |
| Darkness | period_4 | WKY | 0.21 | 1.15 × 10−30 |
| Darkness | period_5 | SHR | 0.27 | 1.91 × 10−47 |
| Darkness | period_5 | WKY | 0.31 | 1.34 × 10−62 |
| Darkness | period_6 | SHR | 0.26 | 5.91 × 10−43 |
| Darkness | period_6 | WKY | 0.31 | 1.25 × 10−70 |
| Light | period_1 | SHR | -0.02 | 0.31 |
| Light | period_1 | WKY | 0.12 | 3.20 × 10−12 |
| Light | period_2 | SHR | 0.03 | 0.14 |
| Light | period_2 | WKY | 0.24 | 9.08 × 10−43 |
| Light | period_3 | SHR | 0.11 | 3.60 × 10−10 |
| Light | period_3 | WKY | 0.23 | 5.94 × 10−39 |
| Light | period_4 | SHR | 0.10 | 3.92 × 10−08 |
| Light | period_4 | WKY | 0.17 | 4.33 × 10−22 |
| Light | period_5 | SHR | 0.15 | 2.80 × 10−19 |
| Light | period_5 | WKY | 0.12 | 2.69 × 10−11 |
| Light | period_6 | SHR | 0.08 | 6.43 × 10−07 |
| Light | period_6 | WKY | 0.36 | 2.80 × 10−100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryabinina, A.Y.; Bryk, A.A.; Blagonravov, M.L.; Goryachev, V.A.; Mozhaev, A.A.; Ovechkina, V.S. Circadian Rhythms of Body Temperature and Locomotor Activity in Spontaneously Hypertensive Rats under Frequent Changes in Light Conditions. Pathophysiology 2024, 31, 127-146. https://doi.org/10.3390/pathophysiology31010010
Ryabinina AY, Bryk AA, Blagonravov ML, Goryachev VA, Mozhaev AA, Ovechkina VS. Circadian Rhythms of Body Temperature and Locomotor Activity in Spontaneously Hypertensive Rats under Frequent Changes in Light Conditions. Pathophysiology. 2024; 31(1):127-146. https://doi.org/10.3390/pathophysiology31010010
Chicago/Turabian StyleRyabinina, Anna Yu., Anna A. Bryk, Mikhail L. Blagonravov, Vyacheslav A. Goryachev, Andrey A. Mozhaev, and Vera S. Ovechkina. 2024. "Circadian Rhythms of Body Temperature and Locomotor Activity in Spontaneously Hypertensive Rats under Frequent Changes in Light Conditions" Pathophysiology 31, no. 1: 127-146. https://doi.org/10.3390/pathophysiology31010010
APA StyleRyabinina, A. Y., Bryk, A. A., Blagonravov, M. L., Goryachev, V. A., Mozhaev, A. A., & Ovechkina, V. S. (2024). Circadian Rhythms of Body Temperature and Locomotor Activity in Spontaneously Hypertensive Rats under Frequent Changes in Light Conditions. Pathophysiology, 31(1), 127-146. https://doi.org/10.3390/pathophysiology31010010