
Effects of Different Land Use Types and Soil Depths on Soil Mineral Elements, Soil Enzyme Activity, and Fungal Community in Karst Area of Southwest China


 , , , , and
, , , , and 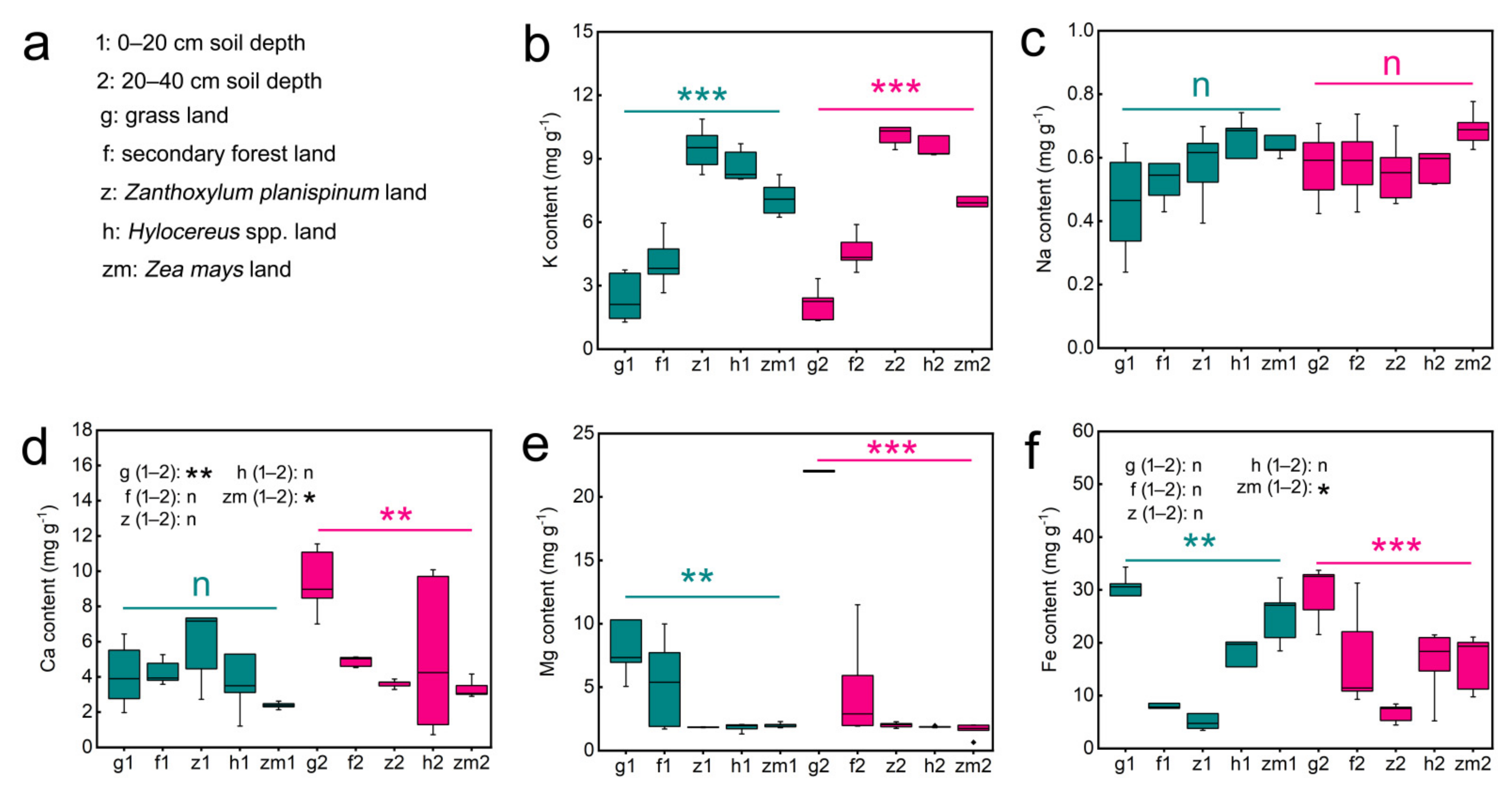{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Soil Sampling
2.2. Soil Mineral Elements Content Assay
2.3. DNA Extraction and Illumina Sequencing
2.4. Sequencing Data Processing and Analysis
2.5. Soil Enzymes Assay
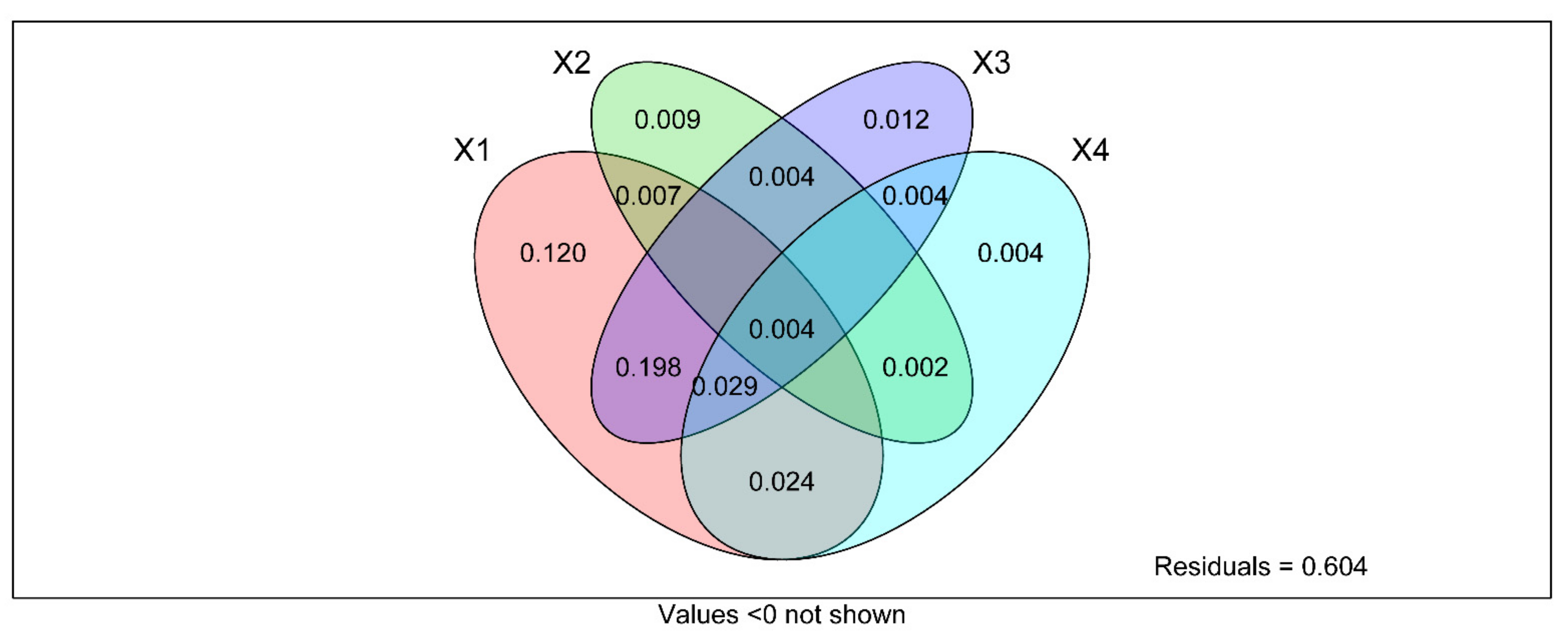
2.6. Statistical Analyses
3. Results
3.1. Differences in Content of Soil K, Na, Ca, Mg and Fe in Different Land Use Types and Soil Depths
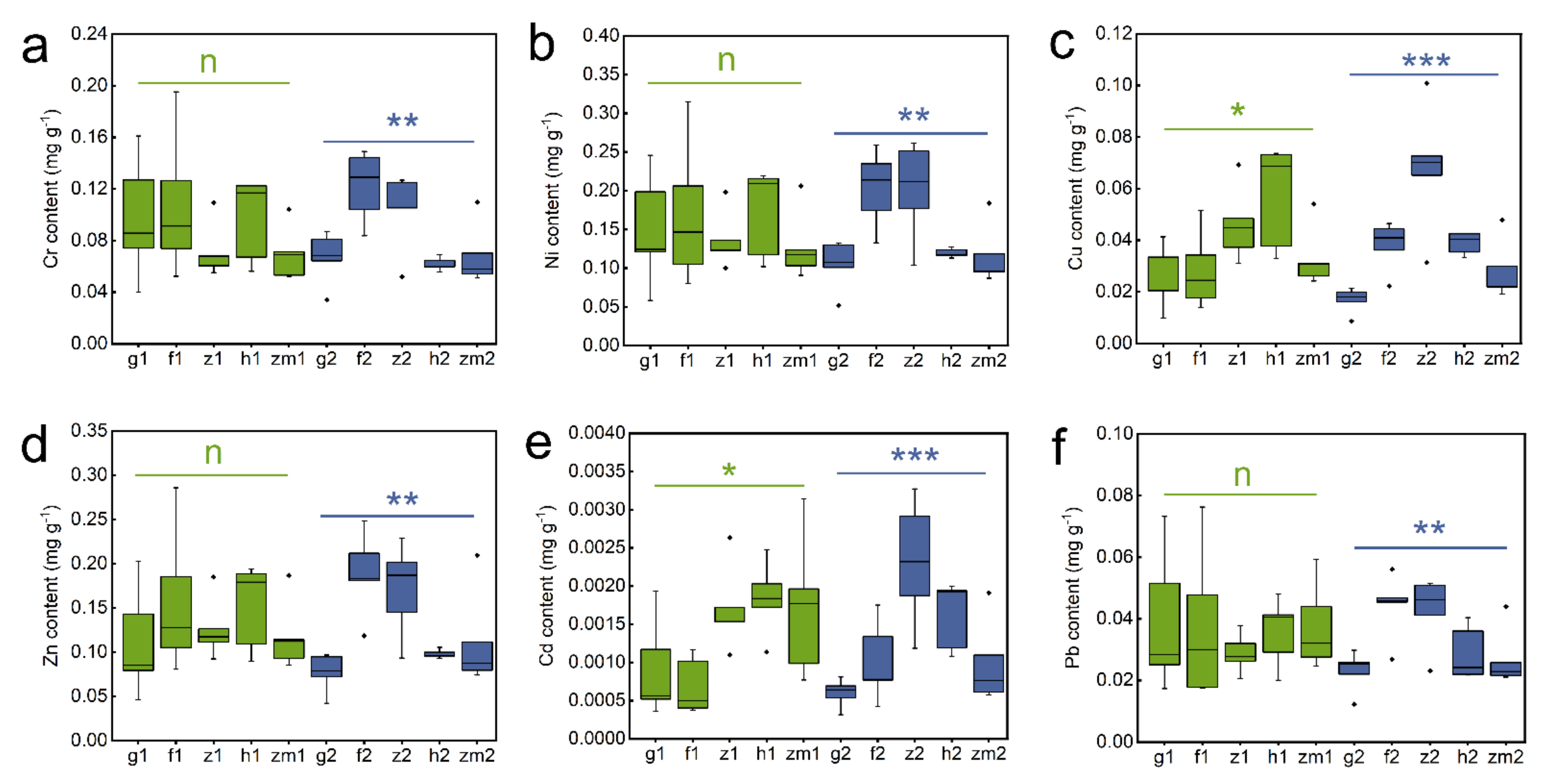
3.2. Differences in Content of soil Cr, Ni, Cu, Zn, Cd and Pb in Different Land Use Types and Soil Depths
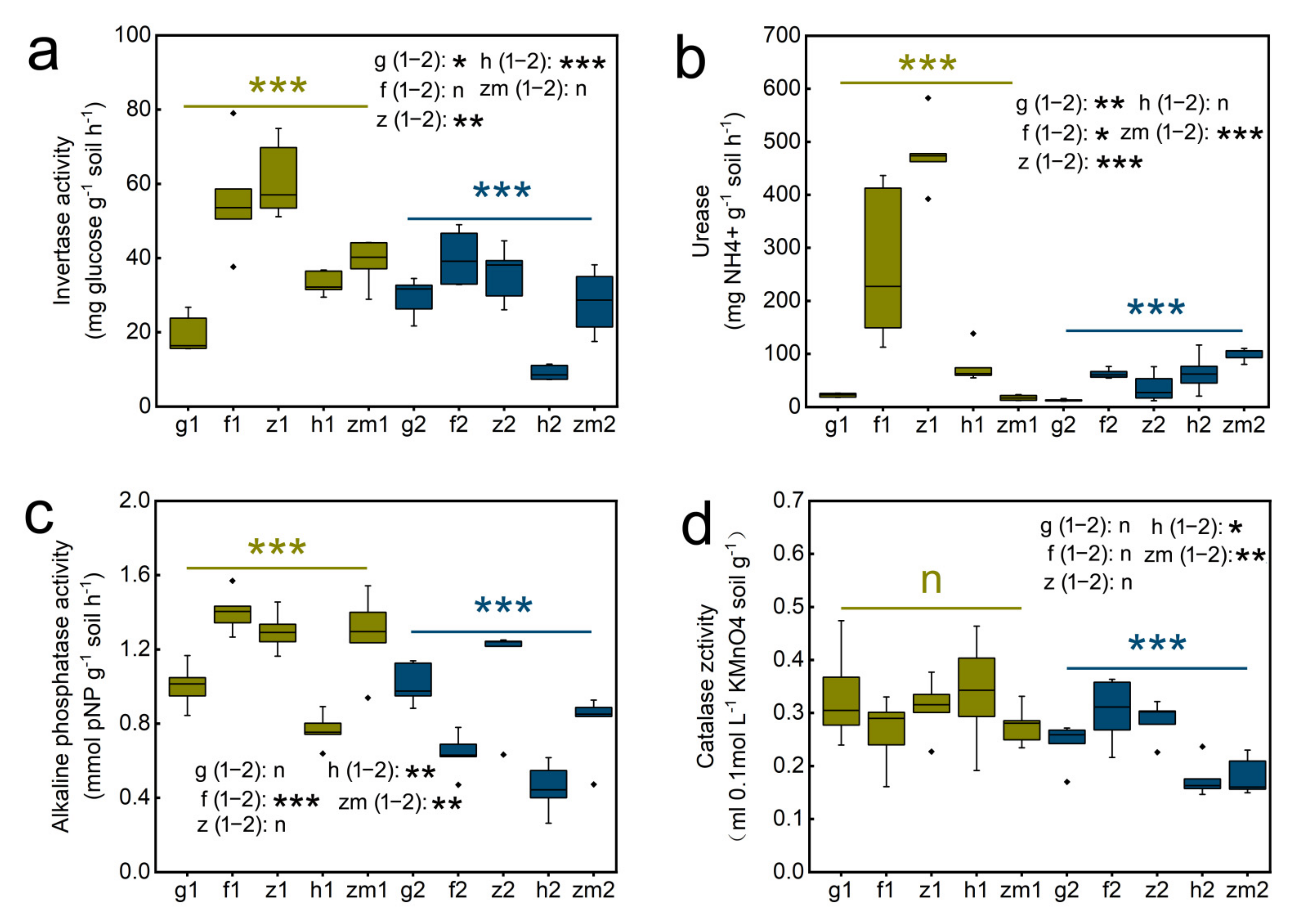
3.3. Differences in Soil Enzyme Activity in Different Land Use Types and Soil Depths
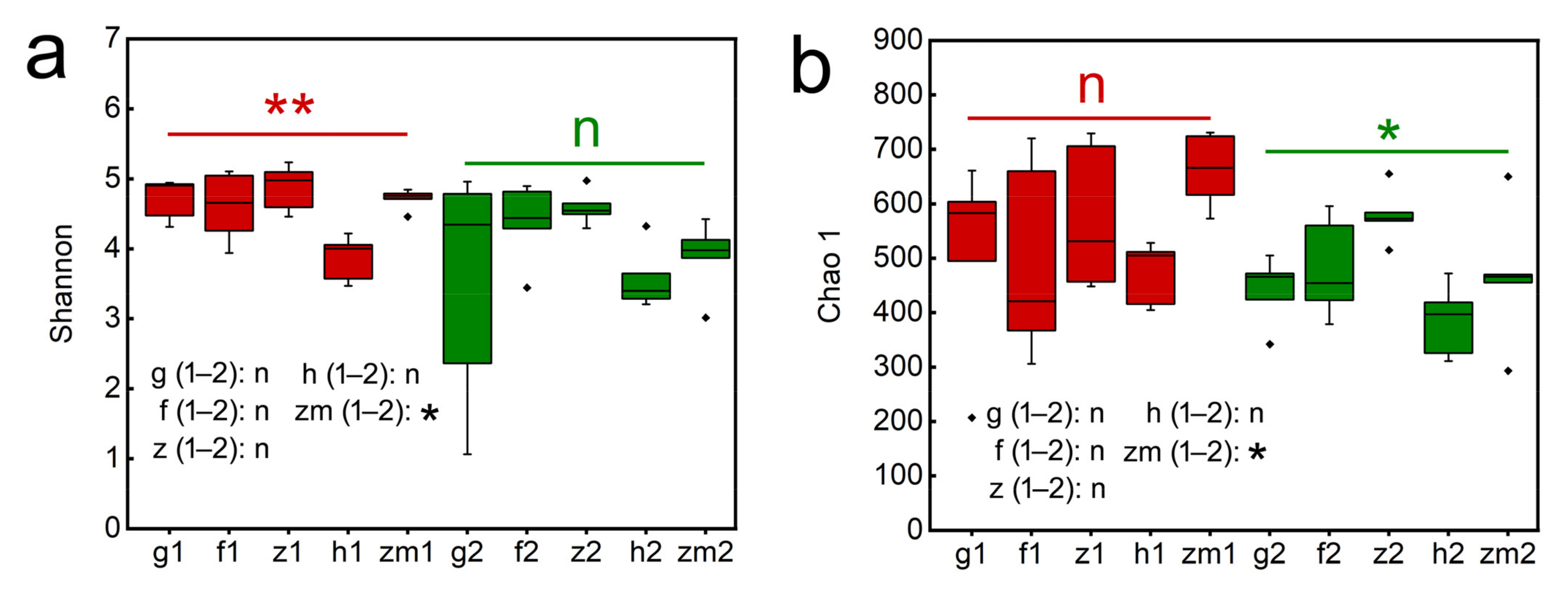
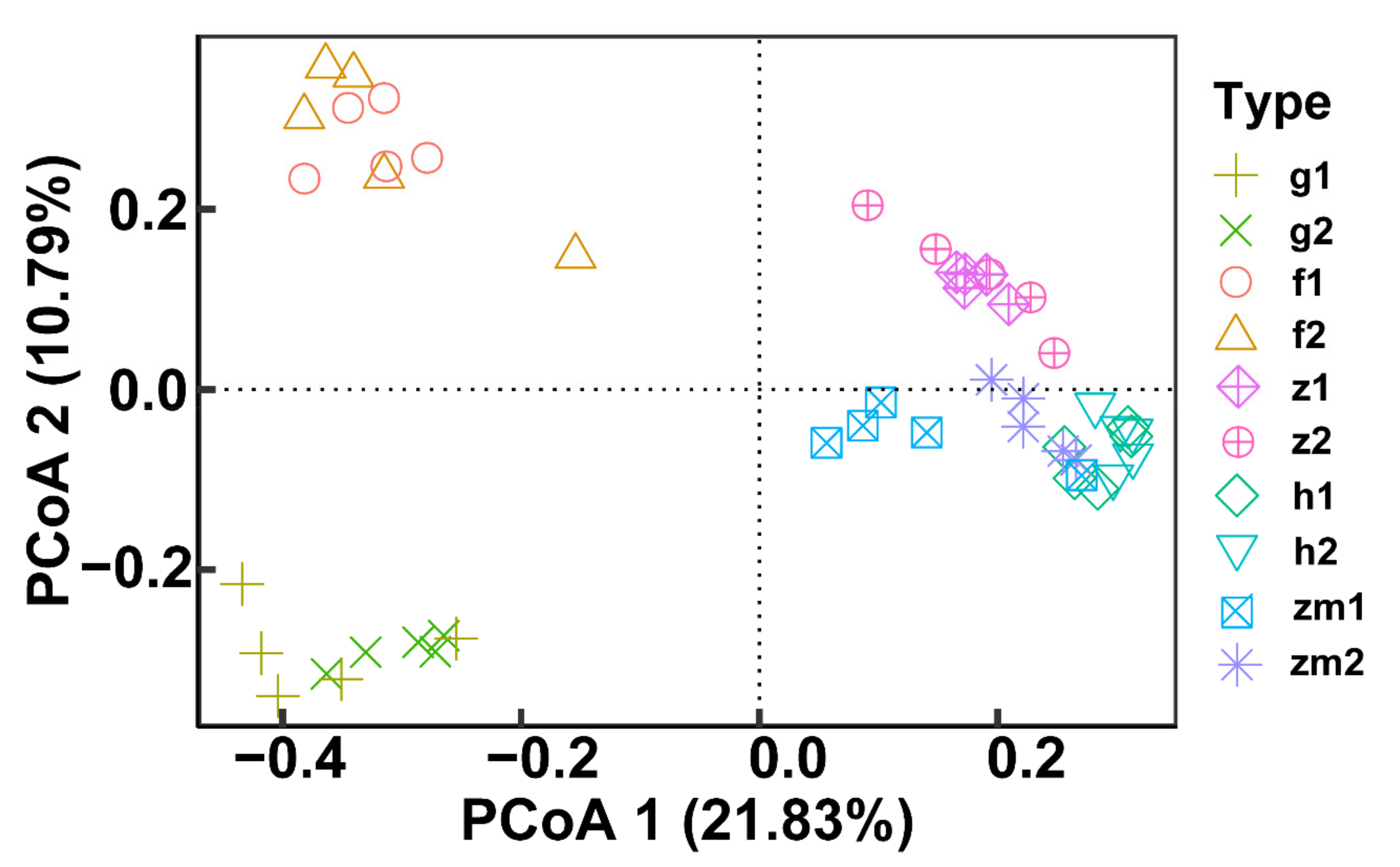
3.4. The Richness and Diversity of Soil Fungal Community
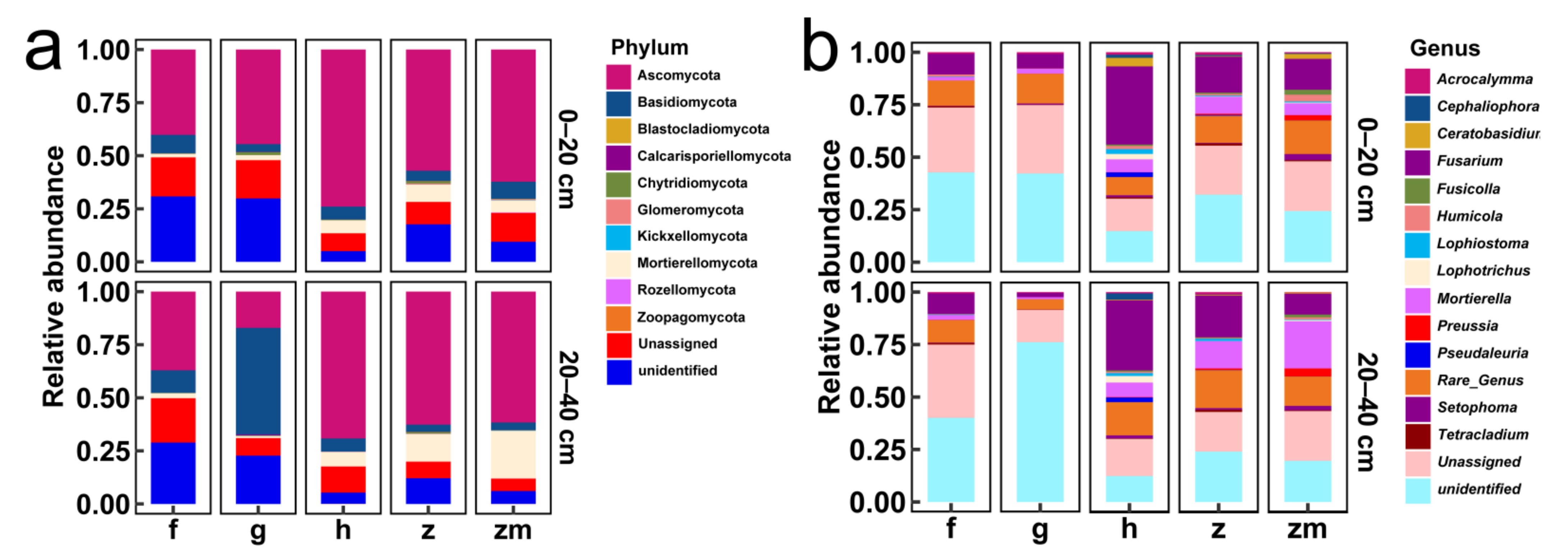
3.5. Relative Abundance of Soil Fungal Community
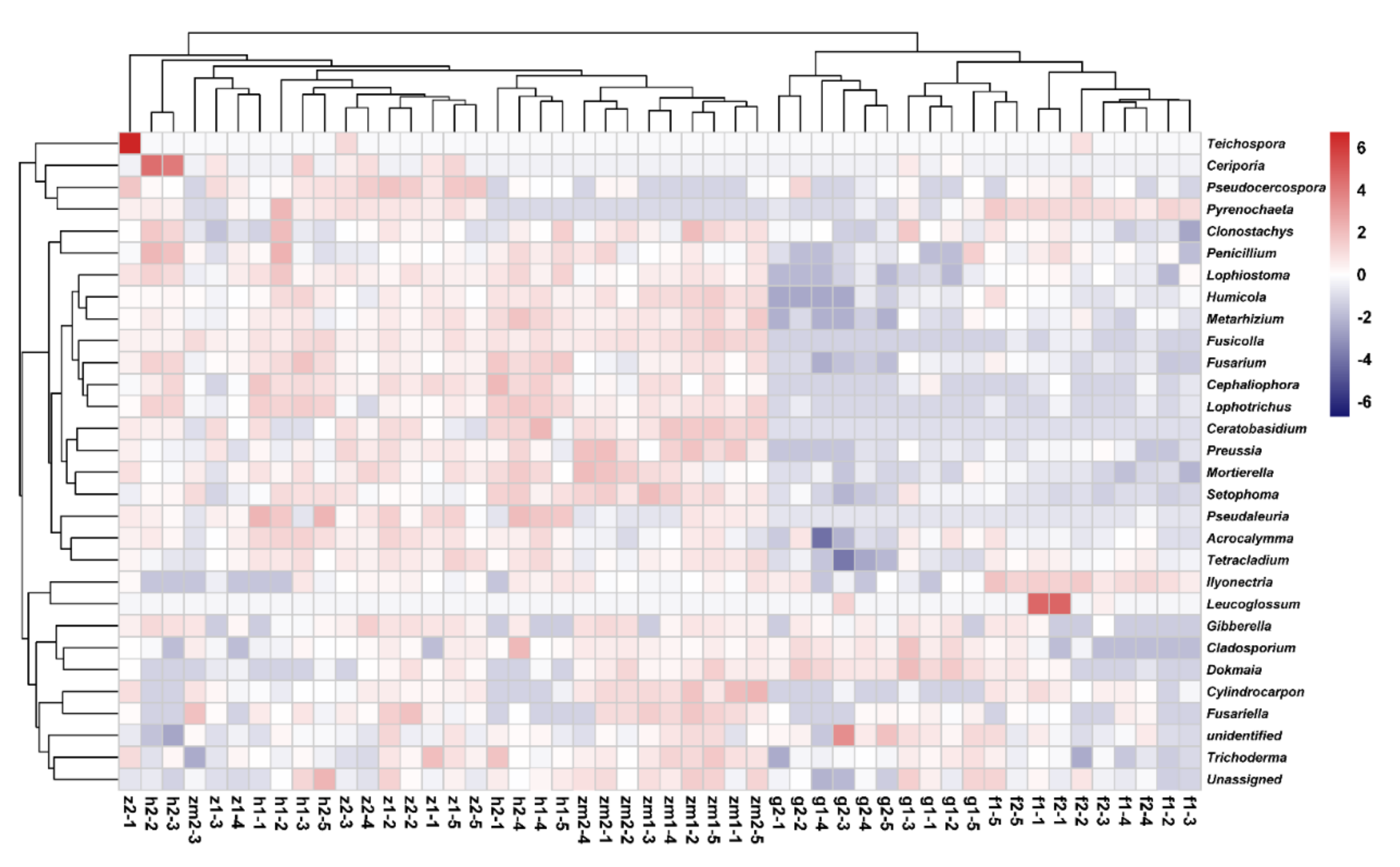
3.6. Comparison of Different Fungal Species at 0–20 cm and 20–40 cm Soil Depth in Different Land Use Types
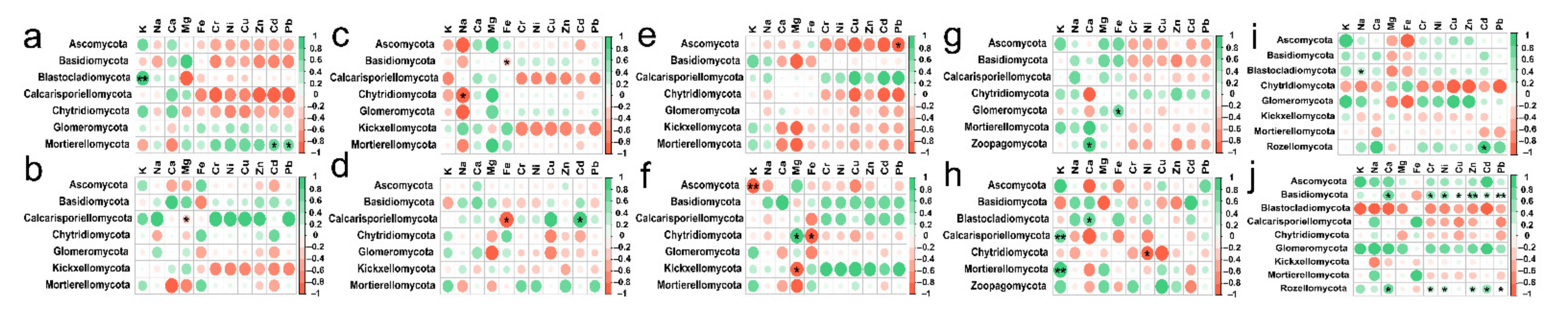
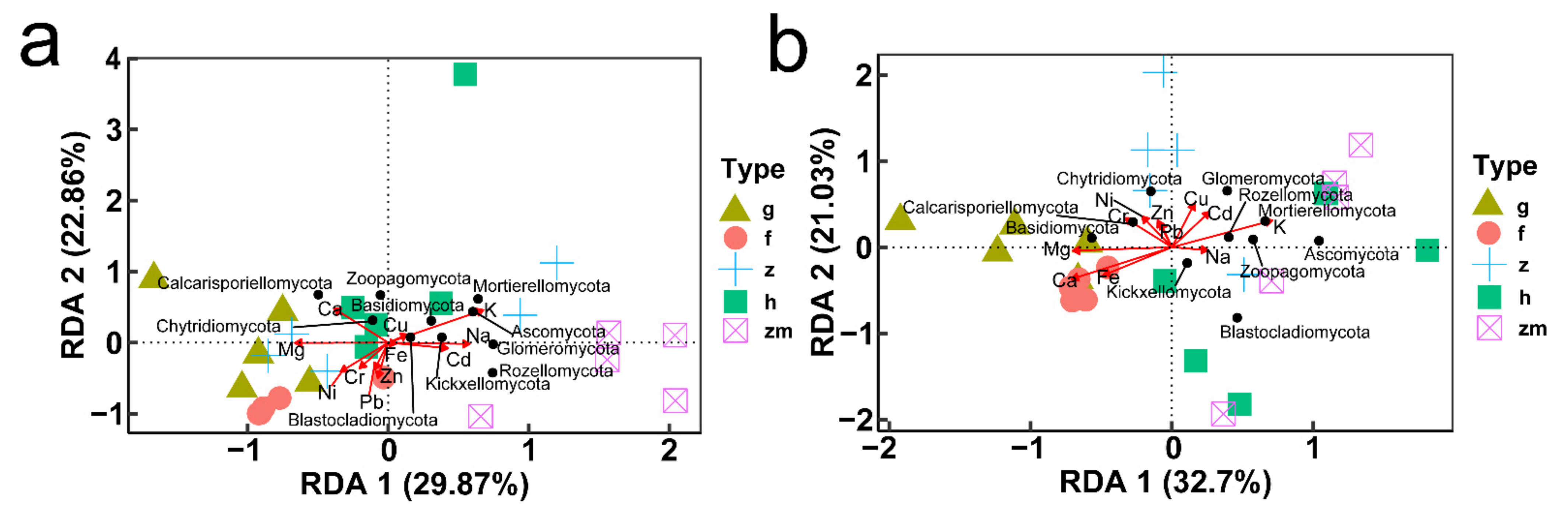
3.7. Correlation of Soil Fungal Communities Composition and Diversity with Content of Soil Mineral Elements
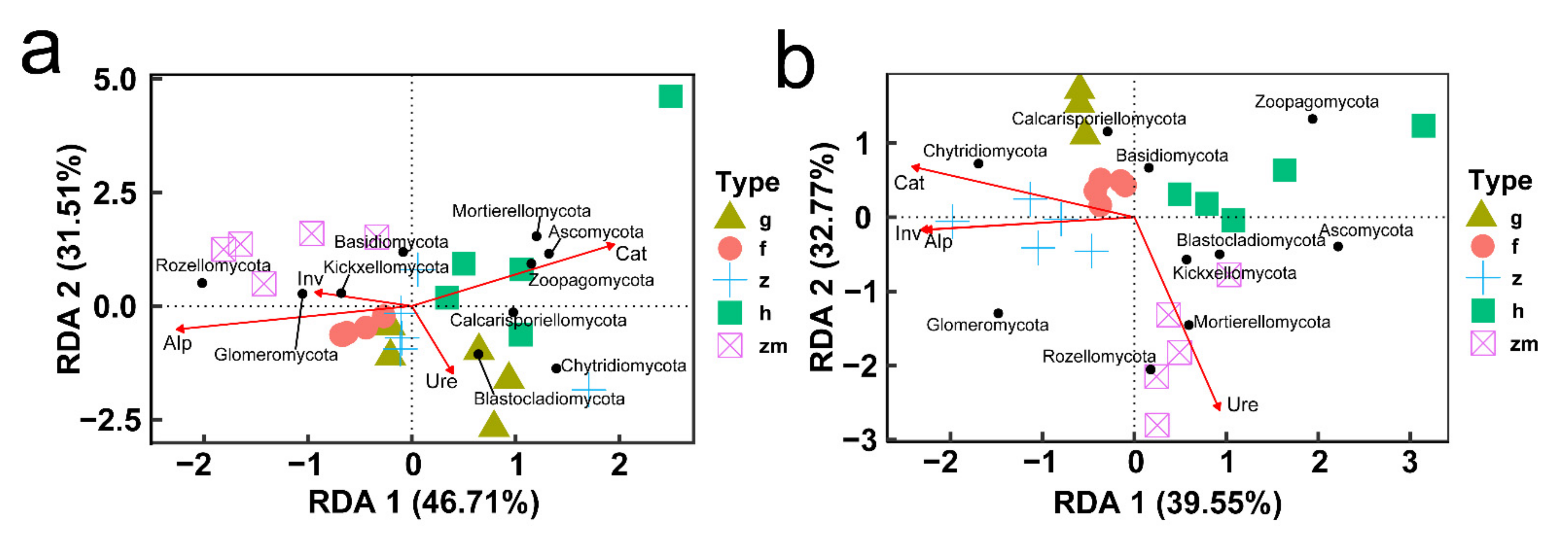
3.8. Correlation of Soil Fungal Communities Composition and Diversity with Soil Enzyme Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sweeting, M.M. Karst in China. In Karst in China Series: Springer Series in Physical Environment; Springer: Berlin/Heidelberg, Germany, 1995; Volume 15. [Google Scholar]
- Green, S.M.; Dungait, J.; Tu, C.; Buss, H.L.; Sanderson, N.; Hawkes, S.J.; Xing, K.; Yue, F.; Hussey, V.L.; Peng, J. Soil functions and ecosystem services research in the Chinese karst Critical Zone. Chem. Geol. 2019, 527, 119107. [Google Scholar] [CrossRef]
- Yang, X.Q.; Hu, B.Q. Quality Characteristics of Soils in Karst Rocky-Desertified Areas With Ecosystem Under Restoration Succession-A Case Study of Chengjiang Subwatershed,Du′an County, Guangxi. J. Ecol. Rural Environ. 2009, 25, 1–5. [Google Scholar]
- Richardson, M.; Kumar, P. Critical Zone services as environmental assessment criteria in intensively managed landscapes. Earth’s Future 2017, 5, 617–632. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Aber, J.D.; Howarth, R.W.; Likens, G.E.; Matson, P.A.; Schindler, D.W.; Schlesinger, W.H.; Tilman, D.G. Human alteration of the global nitrogen cycle: Sources and consequences. Ecol. Appl. 1997, 7, 737–750. [Google Scholar] [CrossRef]
- Xiao, S.; Wei, Z.; Ye, Y.; Jie, Z.; Wang, K. Soil aggregate mediates the impacts of land uses on organic carbon, total nitrogen, and microbial activity in a Karst ecosystem. Sci. Rep. 2017, 7, 41402. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Su, Y.; He, X.; Wei, Y.; Wei, W.; Wu, J. Soil bacterial community composition and diversity respond to cultivation in Karst ecosystems. World J. Microbiol. Biotechnol. 2012, 28, 205–213. [Google Scholar] [CrossRef]
- He, S.; Guo, L.; Niu, M.; Miao, F.; Jiao, S.; Hu, T.; Long, M. Ecological diversity and co-occurrence patterns of bacterial community through soil profile in response to long-term switchgrass cultivation. Sci. Rep. 2017, 7, 3608. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.F.; Xie, S.Y.; Feng, H.F.; Yuan, W.H.; Wang, C.X. Characteristic Study of Soil Microbe under Different Land-use Types in Chongqing Karst Region. J. Environ. Sci. Manag. 2010, 35, 150–154. [Google Scholar]
- Makoi, J.; Ndakidemi, P.A. Selected soil enzymes: Examples of their potential roles in the ecosystem. Afr. J. Biotechnol. 2018, 7, 181–191. [Google Scholar]
- Aon, M.A.; Colaneri, A.C. Temporal and spatial evolution of enzymatic activities and physico-chemical properties in an agricultural soil. Appl. Soil Ecol. 2011, 18, 255–270. [Google Scholar] [CrossRef]
- Wang, X.; Fan, J.; Xing, Y.; Xu, G.; Wang, H.; Jian, D.; Wang, Y.; Zhang, F.; Li, P.; Li, Z. The Effects of Mulch and Nitrogen Fertilizer on the Soil Environment of Crop Plants. Adv. Agron. 2019, 153, 121–173. [Google Scholar]
- Wu, J.; Wang, H.; Li, G.; Ma, W.; Xu, G. Vegetation degradation impacts soil nutrients and enzyme activities in wet meadow on the Qinghai-Tibet Plateau. Sci. Rep. 2020, 10, 21271. [Google Scholar] [CrossRef] [PubMed]
- Dick, R.P.; Kandeler, E. Enzymes in soils. Encycl. Soils Environ. 2005, 448–456. [Google Scholar]
- Jílková, V.; Jandová, K.; Kukla, J. Responses of microbial activity to carbon, nitrogen, and phosphorus additions in forest mineral soils differing in organic carbon content. Biol. Fertil. Soils 2021, 57, 513–521. [Google Scholar] [CrossRef]
- Maithani, S.; Pal, M.; Maity, A.; Pradhan, M. Isotope selective activation: A new insight into the catalytic activity of urease. RSC Adv. 2017, 7, 31372–31376. [Google Scholar] [CrossRef]
- Khadem, A.; Raiesi, F. Response of soil alkaline phosphatase to biochar amendments: Changes in kinetic and thermodynamic characteristics. Geoderma 2019, 337, 44–54. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, S.; Li, X.; Wang, J.; Ding, Q.; Wang, H.; Tian, C.; Yao, M.; An, J.; Huang, Y. Changes of soil prokaryotic communities after clear-cutting in a karst forest: Evidences for cutting-based disturbance promoting deterministic processes. FEMS Microbiol. Ecol. 2016, 92, fiw026. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Wrigh, J.P.; Cadotte, M.W.; Carroll, I.T.; Hector, S. Impacts of plant diversity on biomass production increase through time because of species complementarity. Proc. Nat. Acad. Sci. USA 2007, 104, 18123–18128. [Google Scholar] [CrossRef]
- Narottam, S.; Mollah, M.Z.I.; Alam, M.F.; Rahman, S.M. Seasonal investigation of heavy metals in marine fishes captured from the Bay of Bengal and the implications for human health risk assessment. Food Control 2016, 70, 110–118. [Google Scholar]
- Liu, R.; Zhang, Z.; Shen, J.; Wang, Z. Community characteristics of bryophyte in Karst caves and its effect on heavy metal pollution: A case study of Zhijin Cave, Guizhou Province. Biodivers. Sci. 2018, 26, 1277–1288. [Google Scholar] [CrossRef]
- Ping, C.; Ruan, Y.; Wang, S.; Liu, X.; Lian, B. Effects of organic mineral fertiliser on heavy metal migration and potential carbon sink in soils in a karst region. Chin. J. Geochem. 2017, 36, 539–543. [Google Scholar]
- Jiao, W.; Ouyang, W.; Hao, F.; Liu, B.; Wang, F. Geochemical variability of heavy metals in soil after land use conversions in Northeast China and its environmental applications. Environ. Sci. Processes Impacts 2014, 16, 924–931. [Google Scholar] [CrossRef]
- Chrastný, V.; Komárek, M.; Procházka, J.; Pechar, L.; Vaněk, A.; Penížek, V.; Farka, J. 50 years of different landscape management influencing retention of metals in soils. J. Geochem. Explor. 2012, 115, 59–68. [Google Scholar] [CrossRef]
- Qi, D.; Wieneke, X.; Zhou, X.; Jiang, X.; Xue, P. Succession of plant community composition and leaf functional traits in responding to karst rocky desertification in the Wushan County in Chongqing, China. Community Ecol. 2017, 18, 157–168. [Google Scholar] [CrossRef][Green Version]
- Li, Y.; Han, C.; Sun, S.; Zhao, C. Effects of Tree Species and Soil Enzyme Activities on Soil Nutrients in Dryland Plantations. Forests 2021, 12, 1153. [Google Scholar] [CrossRef]
- Zhong, Y.; Yan, W.; Wang, R.; Wang, W.; Shangguan, Z.P. Decreased occurrence of carbon cycle functions in microbial communities along with long-term secondary succession. Soil Biol. Biochem. 2018, 123, 207–217. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 2013, 4, 914–919. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microb. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.; Bahram, M.; Bates, S.; Bruns, T.; Bengtsson-Palme, J.; Callaghan, T. Towards a unified paradigm for sequence-based identification of Fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.P.; Wang, J.F.; Nan, Z.B. Epichloë gansuensis endophyte-infection alters soil enzymes activity and soil nutrients at different growth stages of Achnatherum inebrians. Plant Soil 2020, 455, 227–240. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, G.B.; Xue, S.; Lin, Z. Rhizosphere soil microbial activity under different vegetation types on the Loess Plateau, China. Geoderma 2011, 161, 115–125. [Google Scholar] [CrossRef]
- Macdonald, C.A.; Thomas, N.; Robinson, L.; Tate, K.R.; Ross, D.J.; Dando, J.; Singh, B.K. Biochemical and molecular responses of the soil microbial community after afforestation of pastures with Pinus radiata. Soil Biol. Biochem. 2009, 41, 1642–1651. [Google Scholar] [CrossRef]
- Liao, Q.; Nan, Z.; Wang, S.; Huang, H.; Ding, H. Spatial variability and abundance evaluation of available microelements in the middle reaches of Heihe River. J. Arid. Land Resour. Environ. 2012, 13, 187. [Google Scholar]
- Chen, C.; Yang, F.; Liu, H.; Yao, H.; Song, G. Effects and evaluation of soil trace elements after grassland converted into cropland in Guizhou karst area. Trans. Chin. Soc. Agric. Eng. 2013, 29, 230–237. [Google Scholar]
- Li, J.; Richter, D.D.; Mendoza, A.; Heine, P. Effects of land-use history on soil spatial heterogeneity of macro- and trace elements in the Southern Piedmont USA. Geoderma 2010, 156, 60–73. [Google Scholar] [CrossRef]
- Song, M.; Zou, D.; Hu, D.U.; Peng, W.; Zeng, F.; Tan, Q.; Fan, F. Characteristics of soil microbial populations in depressions between karst hills under different land use patterns. Chin. J. Appl. Ecol. 2013, 24, 2471–2478. [Google Scholar]
- Feyisa, D.; Kissi, E.; Kebebew, Z. Rethinking Eucalyptus globulus Labill. Based Land Use Systems in Smallholder Farmers Livelihoods: A Case of Kolobo Watershed, West Shewa, Ethiopia. Nephron Clin. Pract. 2018, 37, 57–68. [Google Scholar] [CrossRef]
- Yu, Y.H.; Yang, D.; Zhong, X. Characteristics of Soil Affinity Elements of Typical Land Use Types in the Rocky Desertification Area of Central Guizhou. Earth Environ. 2019, 47, 429–435. [Google Scholar]
- Yang, Y.; Hu, D.; Song, T.; Peng, W.; Zeng, F.; Wang, K.; Lu, S.Y.; Fan, F.; Lu, C.Y. Characteristics of soil fertility in different ecosystems in depressions between karst hills. Acta Ecol. Sin. 2013, 33, 7455–7466. [Google Scholar] [CrossRef]
- Chen, T.; Wei, X.; Guan, G.; Li, Z.Y. Impact of Different Land Use Types on Soil Calcium in Northern Guangdong. Trop. Geogr. 2014, 15, 61–68. [Google Scholar]
- Chen, J.R.; Cao, J.H.; Liang, Y.; Yang, H. Relationship of the humus components and the calcium form with the development of limestone soil. Carsol. Sin. 2012, 31, 7–11. [Google Scholar]
- Xie, L.P.; Wang, S.J.; Xiao, D.A. Ca Covariant Relation in Plant-soil System in a Small Karst Catshment. Earth Environ. 2007, 1, 26–32. [Google Scholar]
- Fraterrigo, J.M.; Turner, M.G.; Pearson, S.M.; Dixon, P. Effects of Past Land Use on Spatial Heterogeneity of Soil Nutrients in Southern Appalachian Forests. Ecol. Monogr. 2005, 75, 215–230. [Google Scholar] [CrossRef]
- Han, M.R.; Song, T.Q.; Peng, W.X.; Huang, G.Q.; Shi, W.W. Compositional characteristics and roles of soil mineral substances in depressions between hills in karst region China. J. Appl. Ecol. 2012, 23, 685–693. [Google Scholar]
- Liu, X.L.; He, Y.Q.; Zhang, H.L.; Schroder, J.K.; Li, C.L.; Zhou, J.; Zhang, Z.Y. Impact of Land Use and Soil Fertility on Distributions of Soil Aggregate Fractions and Some Nutrients. Pedosphere 2010, 20, 666–673. [Google Scholar] [CrossRef]
- Negasa, D.J. Effects of Land Use Types on Selected Soil Properties in Central Highlands of Ethiopia. Appl. Environ. Soil Sci. 2020, 2020, 7026929. [Google Scholar] [CrossRef]
- Liu, S.; Fu, J.P.; Cai, X.; Zhou, J.; Dang, Z.; Zhu, R. Effect of Heavy Metals Pollution on Ecological Characteristics of Soil Microbes: A Review. Ecol. Environ. Sci. 2018, 27, 1173–1178. [Google Scholar]
- Du, J.Y.; Tu, C.L.; Sheng, M.Y.; Cui, L.F.; Chen, Z.Y.; Zhang, L.K. Effect of Land Use Change on Microbial Community Structure in Central Guizhou Province. J. Sichuan Agric. Univ. 2018, 36, 350–356. [Google Scholar]
- Wilcke, W.; Krauss, M.; Kobza, J. Concentrations and forms of heavy metals in Slovak soils. J. Plant Nutr. Soil Sci. 2010, 168, 676–686. [Google Scholar] [CrossRef]
- Li, Q.; Hu, Q.; Zhang, C.; Jin, Z. Effects of Pb, Cd, Zn, and Cu on Soil Enzyme Activity and Soil Properties Related to Agricultural Land-Use Practices in Karst Area Contaminated by Pb-Zn Tailings. Pol. J. Environ. Stud. 2018, 27, 2623–2632. [Google Scholar] [CrossRef]
- Jia, Y.N.; Yuan, D.X. Effects of Land Use Changes on Trace Elements of Karst Soil in Shuicheng Basin. J. Soil Sci. 2007, 6, 1174–1177. [Google Scholar]
- Thinh, N.V.; Akinori, O.; Hoang, N.T.; Anh, N.D.; Yen, T.T.; Kiyoshi, K. Arsenic and Heavy Metal Contamination in Soils under Different Land Use in an Estuary in Northern Vietnam. Int. J. Environ. Res. Public Health 2016, 13, 1091. [Google Scholar]
- Ouyang, W.; Shan, Y.; Hao, F.; Lin, C. Differences in soil organic carbon dynamics in paddy fields and drylands in northeast China using the CENTURY model. Agric. Ecosyst. Environ. 2014, 194, 38–47. [Google Scholar] [CrossRef]
- Zheng, R.; Zhao, J.; Zhou, X.; Chao, M.A.; Wang, L.; Gao, X. Land Use Effects on the Distribution and Speciation of Heavy Metals and Arsenic in Coastal Soils on Chongming Island in the Yangtze River Estuary, China. Psedosphere 2016, 56, 74–84. [Google Scholar] [CrossRef]
- Sun, C.L.; Wang, Y.W.; Wang, C.J.; Li, Q.J.; Wu, Z.H.; Yuan, D.S.; Zhang, J.L. Effects of land use conversion on soil extracellular enzyme activity and its stoichiometric characteristics in karst mountainous. Acta Ecol. Sin. 2021, 41, 4140–4149. [Google Scholar]
- Gocke, M.I.; Huguet, A.; Derenne, S.; Kolb, S.; Dippold, M.A.; Wiesenberg, G. Disentangling interactions between microbial communities and roots in deep subsoil. Sci. Total Environ. 2017, 575, 135–145. [Google Scholar] [CrossRef]
- Stone, M.M.; Deforest, J.L.; Plante, A.F. Changes in extracellular enzyme activity and microbial community structure with soil depth at the Luquillo Critical Zone Observatory. Soil Biol. Biochem. 2014, 75, 237–247. [Google Scholar] [CrossRef]
- Gelsomino, A.; Azzellino, A. Multivariate analysis of soils: Microbial biomass, metabolic activity, and bacterial-community structure and their relationships with soil depth and type. J. Plant Nutr. Soil Sci. 2011, 174, 381–394. [Google Scholar] [CrossRef]
- Hao, C.; Pan, L.; Li, W.; Yang, L.; Li, D. Determinants of soil extracellular enzyme activity in a karst region, southwest China. Eur. J. Soil Biol. 2017, 80, 69–76. [Google Scholar]
- Wallenius, K.; Rita, H.; Mikkonen, A.; Lappi, K.; Lindstr, M.K.; Hartikainen, H.; Raateland, A.; Niemi, R.M. Effects of land use on the level, variation and spatial structure of soil enzyme activities and bacterial communities. Soil Biol. Biochem. 2011, 43, 1464–1473. [Google Scholar] [CrossRef]
- Keeler, B.L.; Hobbie, S.E.; Kellogg, L.E. Effects of Long-Term Nitrogen Addition on Microbial Enzyme Activity in Eight Forested and Grassland Sites: Implications for Litter and Soil Organic Matter Decomposition. Ecosystems 2009, 12, 1–15. [Google Scholar] [CrossRef]
- Liu, L.; Chen, H.; Li, D.; Liang, S. Changes of soil hydrolytic and oxidized enzyme activities under the process of vegetation restoration in a karst area, southwest china. Acta Sci. Circumstantiae 2017, 37, 3528–3534. [Google Scholar]
- Schlatter, D.C.; Kendall, K.; Bryan, C.; Huggins, D.R.; Timothy, P. Fungal community composition and diversity vary with soil depth and landscape position in a no-till wheat-based cropping system. FEMS Microbiol. Ecol. 2018, 7, fiy098. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zheng, W.; Zhang, S.; Gao, W.; Fan, Y. Soil Microbial Community Variation with Time and Soil Depth in Eurasian Steppe (Inner Mongolia, China). Ann. Microbiol. 2021, 71, 21. [Google Scholar] [CrossRef]
- Ko, D.; Yoo, G.; Yun, S.T.; Jun, S.C.; Chung, H. Bacterial and fungal community composition across the soil depth profiles in a fallow field. J. Ecol. Environ. 2017, 41, 34. [Google Scholar] [CrossRef]
- Qin, H.; Li, C.X.; Ren, Q. Effects of different land use patterns on soil bacterial and fungal biodiversity in the hydro-fluctuation zone of the Three Gorges Reservoir region. Acta Ecol. Sin. 2017, 37, 3494–3504. [Google Scholar]
- Yang, Y.; Dou, Y.; Huang, Y.; An, S. Links between Soil Fungal Diversity and Plant and Soil Properties on the Loess Plateau. Front. Micro. 2017, 8, 2198. [Google Scholar] [CrossRef]
- Cheng, Y.Y.; Jin, Z.J.; Wang, X.T.; Jia, Y.H.; Zhou, J.B. Effect of Land-use on Soil Fungal Community Structure and Associated Functional Group in Huixian Karst Wetland. China Environ. Sci. 2020, 41, 4294–4304. [Google Scholar]
- Wang, G.; Liu, Y.; Cui, M.; Zhou, Z.; Zhou, J. Effects of secondary succession on soil fungal and bacterial compositions and diversities in a karst area. Plant Soil 2021. [Google Scholar] [CrossRef]
- Sui, X.; Zhang, R.T.; Xu, N. Fungal community structure of different degeneration Deyeuxia angustifoliawetlands in Sanjiang Plain. Environ. Sci. 2016, 37, 3598–3605. [Google Scholar]
- Geisseler, D.; Scow, K.M. Long-term effects of mineral fertilizers on soil microorganisms-a review. Soil Biol. Biochem. 2014, 75, 54–63. [Google Scholar] [CrossRef]
- Mylène, H.; Patricia, L.; Julien, G.; Zahar, H. Plant host habitat and root exudates shape fungal diversity. Mycorrhiza 2018, 28, 451–463. [Google Scholar]
- Peng, D.; Xuan, Y.; Le, H.; Liu, J.; Zhong, Z. Effects of stand age and soil properties on soil bacterial and fungal community composition in Chinese pine plantations on the Loess Plateau. PLoS ONE 2017, 12, e0186501. [Google Scholar]
- Haug, I.; Lempe, J.; Homeier, J.; Wei, M.; Setaro, S.; Oberwinkler, F.; Kottke, I. Graffenrieda emarginata (Melastomataceae) forms mycorrhizas with Glomeromycota and with a member of the Hymenoscyphus ericae aggregate in the organic soil of a neotropical mountain rain forest. Can. J. Bot. 2004, 82, 340–356. [Google Scholar] [CrossRef]
- Chodak, M.; Gołębiewski, M.; Morawska-Płoskonka, J.; Kuduk, K.; Niklińska, M. Diversity of microorganisms from forest soils differently polluted with heavy metals. Appl. Soil Ecol. 2013, 64, 7–14. [Google Scholar] [CrossRef]
- Xue, C.; Penton, C.R.; Zhu, C.; Chen, H.; Duan, Y.; Peng, C.; Guo, S.; Ling, N.; Shen, Q. Alterations in soil fungal community composition and network assemblage structure by different long-term fertilization regimes are correlated to the soil ionome. Biol. Fertil. Soils 2018, 54, 95–106. [Google Scholar] [CrossRef]
- Pan, X.; Zhang, S.; Zhong, Q.; Gong, G.; Wang, G.; Guo, X.; Xu, X. Effects of soil chemical properties and fractions of Pb, Cd, and Zn on bacterial and fungal communities. Sci. Total Environ. 2020, 715, 136901–136904. [Google Scholar] [CrossRef]
- Benedicte, B.; María, U.J.; Zimmerman, J.K.; Thompson, J.; Jonathan, W.L. Long-lasting effects of land use history on soil fungal communities in second-growth tropical rain forests. Ecol. Appl. 2016, 26, 1881–1895. [Google Scholar]
- Huang, M.; Fu, H.; Kong, X.; Ma, L.; Liu, C.; Fang, Y.; Zhang, Z.; Song, F.; Yang, F. Effects of Fertilization Methods on Chemical Properties, Enzyme Activity, and Fungal Community Structure of Black Soil in Northeast China. Diversity 2020, 12, 476. [Google Scholar] [CrossRef]
- Zhang, Y.; Cao, H.; Zhao, P.; Wei, X.; Shi, M. Vegetation Restoration Alters Fungal Community Composition and Functional Groups in a Desert Ecosystem. Front. Environ. Sci. 2021, 9, 589068. [Google Scholar] [CrossRef]
- Ai, C.; Zhang, S.; Zhang, X.; Guo, D.; Zhou, W.; Huang, S. Distinct responses of soil bacterial and fungal communities to changes in fertilization regime and crop rotation. Geodera 2018, 319, 156–166. [Google Scholar] [CrossRef]
- Wang, J.; Yuan, Y.; Zhang, M.; Dai, X.; He, H.; Li, H.; Li, Y. Impact of degradation and restoration on soil fungi and extracellular enzyme activity in alpine rangelands on the Tibetan Plateau. Arch. Agron. Soil Sci. 2021, 14, 1917–1929. [Google Scholar] [CrossRef]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, J.; Hou, W.; Liu, J.; Malik, K.; Kong, X.; Wang, L.; Chen, X.; Tang, M.; Zhu, R.; Cheng, C.; et al. Effects of Different Land Use Types and Soil Depths on Soil Mineral Elements, Soil Enzyme Activity, and Fungal Community in Karst Area of Southwest China. Int. J. Environ. Res. Public Health 2022, 19, 3120. https://doi.org/10.3390/ijerph19053120
Gong J, Hou W, Liu J, Malik K, Kong X, Wang L, Chen X, Tang M, Zhu R, Cheng C, et al. Effects of Different Land Use Types and Soil Depths on Soil Mineral Elements, Soil Enzyme Activity, and Fungal Community in Karst Area of Southwest China. International Journal of Environmental Research and Public Health. 2022; 19(5):3120. https://doi.org/10.3390/ijerph19053120
Chicago/Turabian StyleGong, Jiyi, Wenpeng Hou, Jie Liu, Kamran Malik, Xin Kong, Li Wang, Xianlei Chen, Ming Tang, Ruiqing Zhu, Chen Cheng, and et al. 2022. "Effects of Different Land Use Types and Soil Depths on Soil Mineral Elements, Soil Enzyme Activity, and Fungal Community in Karst Area of Southwest China" International Journal of Environmental Research and Public Health 19, no. 5: 3120. https://doi.org/10.3390/ijerph19053120
APA StyleGong, J., Hou, W., Liu, J., Malik, K., Kong, X., Wang, L., Chen, X., Tang, M., Zhu, R., Cheng, C., Liu, Y., Wang, J., & Yi, Y. (2022). Effects of Different Land Use Types and Soil Depths on Soil Mineral Elements, Soil Enzyme Activity, and Fungal Community in Karst Area of Southwest China. International Journal of Environmental Research and Public Health, 19(5), 3120. https://doi.org/10.3390/ijerph19053120