
Effects of Polyurethane Small-Sized Microplastics in the Chironomid, Chironomus riparius: Responses at Organismal and Sub-Organismal Levels


 ,
,  ,
,  and
and 
Abstract
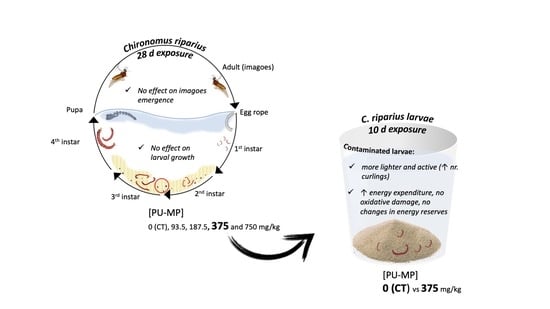
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chironomid Species | Polymer Type, Size, and Concentrations Tested | Main results | |
|---|---|---|---|
| Chironomus riparius | Polyethylene (PE, 32 to 64 μm; [MP] up to 20 g/kg) | Reduced larval growth and delayed imagoes emergence | [6] |
| Induced oxidative damage and reduced aerobic energy production | [9] | ||
| Increased basal phenoloxidase activity | [20] | ||
| Polyethylene terephthalate (PET, 14 μm; [MP] up to 5 × 105 items/kg) | Emergence, weight, or head capsule lengths were not affected | [14] | |
| Polystyrene and tire rubber (PS and TR, 38 and 82 μm, respectively; [MP] up to 10 mg/L) | Caused cellular stress | [8] | |
| Some alterations in gene expression | |||
| Polyvinyl chloride (PVC, <50 μm; [MP] up to 20 g/kg) | Reduced the emergence and weight | [12] | |
| PET, PS, PVC, and polyamide (PA) (10–400 μm as mixture; 8–80 g/m2 sediment) | Increased body mass, larval body length, mouthparts deformities, and female imagoes wind deformities | [11] | |
| Chironomus tepperi | PE (1 to 126 μm, 500 items/L) | Affected survival, growth, and emergence | [7] |
2. Materials and Methods
2.1. Culture Conditions of Chironomus riparius
2.2. Polyurethane Microplastics Used in the Experiment
2.3. A 28-Day Exposure and Evaluated Endpoints
2.4. A 10-Day Exposure and Evaluated Endpoints
2.5. Total Body Length Measurement
2.6. PU-MP Presence in the Larval Digestive Tract
2.7. Larvae Behaviour (Locomotion) Assessment
2.8. Biochemical Parameters Assessment: Oxidative Damage (via LPO), Energy Reserves (Lipids, Proteins, and Carbohydrates), and Energy Consumption (via ETS)
2.9. Statistical Analysis
3. Results and Discussion
3.1. Effects on C. riparius Life History Traits after 28-d Exposure
3.2. Presence of PU-MPs in Chironomus riparius Larval Digestive Tract, after 10 Days of Exposure
3.3. Effects on Larvae Behaviour and Homeostasis after 10-d Exposure
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maltby, L.; Brown, R.; Wilkinson, H. Applying Ecosystem Services Principles to the Derivation of Freshwater Environmental Quality Standards. Front. Environ. Sci. 2022, 10, 1160. [Google Scholar] [CrossRef]
- Hurley, R.; Woodward, J.; Rothwell, J.J. Microplastic Contamination of River Beds Significantly Reduced by Catchment-Wide Flooding. Nat. Geosci. 2018, 11, 251–257. [Google Scholar] [CrossRef]
- Kallenbach, E.M.F.; Rødland, E.S.; Buenaventura, N.T.; Hurley, R. Microplastics in Terrestrial and Freshwater Environments. In Microplastic in the Environment: Pattern and Process; Bank, M.S., Ed.; Springer: Cham, Switzerland, 2022; pp. 87–130. [Google Scholar] [CrossRef]
- Li, C.; Busquets, R.; Campos, L.C. Assessment of Microplastics in Freshwater Systems: A Review. Sci. Total Environ. 2020, 707, 135578. [Google Scholar] [CrossRef]
- Karima, Z.; Karima, Z. Chironomidae: Biology, Ecology and Systematics. In Wonders Diptera—Charact. Divers. Significance World’s Ecosyst; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Silva, C.J.M.; Silva, A.L.P.; Gravato, C.; Pestana, J.L.T. Ingestion of Small-Sized and Irregularly Shaped Polyethylene Microplastics Affect Chironomus riparius Life-History Traits. Sci. Total Environ. 2019, 672, 862–868. [Google Scholar] [CrossRef]
- Ziajahromi, S.; Kumar, A.; Neale, P.A.; Leusch, F.D.L. Environmentally Relevant Concentrations of Polyethylene Microplastics Negatively Impact the Survival, Growth and Emergence of Sediment-Dwelling Invertebrates. Environ. Pollut. 2018, 236, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Navarro, V.; Muñiz-González, A.B.; Sorvari, J.; Martínez-Guitarte, J.L. Altered Gene Expression in Chironomus riparius (Insecta) in Response to Tire Rubber and Polystyrene Microplastics. Environ. Pollut. 2021, 285, 117462. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.J.M.; Patrício Silva, A.L.; Campos, D.; Machado, A.L.; Pestana, J.L.T.; Gravato, C. Oxidative Damage and Decreased Aerobic Energy Production Due to Ingestion of Polyethylene Microplastics by Chironomus riparius (Diptera) Larvae. J. Hazard. Mater. 2021, 402, 123775. [Google Scholar] [CrossRef] [PubMed]
- Yıldız, D.; Yalçın, G.; Jovanović, B.; Boukal, D.S.; Vebrová, L.; Riha, D.; Stanković, J.; Savić-Zdraković, D.; Metin, M.; Akyürek, Y.N.; et al. Effects of a Microplastic Mixture Differ across Trophic Levels and Taxa in a Freshwater Food Web: In Situ Mesocosm Experiment. Sci. Total Environ. 2022, 836, 155407. [Google Scholar] [CrossRef]
- Stanković, J.; Milošević, D.; Savić-Zdraković, D.; Yalçın, G.; Yildiz, D.; Beklioğlu, M.; Jovanović, B. Exposure to a Microplastic Mixture Is Altering the Life Traits and Is Causing Deformities in the Non-Biting Midge Chironomus riparius Meigen (1804). Environ. Pollut. 2020, 262, 114248. [Google Scholar] [CrossRef]
- Scherer, C.; Brennholt, N.; Reifferscheid, G.; Wagner, M. Feeding Type and Development Drive the Ingestion of Microplastics by Freshwater Invertebrates. Sci. Rep. 2017, 7, 17006. [Google Scholar] [CrossRef]
- Silva, C.J.M.; Machado, A.L.; Campos, D.; Rodrigues, A.C.M.; Patrício Silva, A.L.; Soares, A.M.V.M.; Pestana, J.L.T. Microplastics in Freshwater Sediments: Effects on Benthic Invertebrate Communities and Ecosystem Functioning Assessed in Artificial Streams. Sci. Total Environ. 2022, 804, 150118. [Google Scholar] [CrossRef] [PubMed]
- Setyorini, L.; Michler-Kozma, D.; Sures, B.; Gabel, F. Transfer and Effects of PET Microfibers in Chironomus riparius. Sci. Total Environ. 2021, 757, 143735. [Google Scholar] [CrossRef]
- Khosrovyan, A.; Kahru, A. Evaluation of the Hazard of Irregularly-Shaped Co-Polyamide Microplastics on the Freshwater Non-Biting Midge Chironomus riparius through Its Life Cycle. Chemosphere 2020, 244, 125487. [Google Scholar] [CrossRef]
- Armitage, P.D.; Cranston, P.S.; Pinder, L.C.V. The Chironomidae: Biology and Ecology of Non-Biting Midges; Springer B.V.: Dor-drecht, The Netherlands, 1995. [Google Scholar] [CrossRef]
- Rennie, A.R. Thermoplastics and Thermosets. Mech. Prop. Test. Polym. 1999, 3, 248. [Google Scholar] [CrossRef]
- Esterhuizen, M.; Buchenhorst, L.; Kim, Y.J.; Pflugmacher, S. In Vivo Oxidative Stress Responses of the Freshwater Basket Clam Corbicula javanicus to Microplastic Fibres and Particles. Chemosphere 2022, 296, 134037. [Google Scholar] [CrossRef]
- Zimmermann, L.; Dierkes, G.; Ternes, T.A.; Völker, C.; Wagner, M. Benchmarking the in Vitro Toxicity and Chemical Composition of Plastic Consumer Products. Environ. Sci. Technol. 2019, 53, 11467–11477. [Google Scholar] [CrossRef]
- Silva, C.J.M.; Beleza, S.; Campos, D.; Soares, A.M.V.M.; Patrício Silva, A.L.; Pestana, J.L.T.; Gravato, C. Immune Response Triggered by the Ingestion of Polyethylene Microplastics in the Dipteran Larvae Chironomus riparius. J. Hazard. Mater. 2021, 414, 125401. [Google Scholar] [CrossRef] [PubMed]
- ASTM. Standard Guide for Conducting Acute Toxicity Tests on Test Materials with Fishes, Macroinvertebrates, and Amphibians; Philadelphia. 1996. Available online: https://www.astm.org/Standards/E729.htm (accessed on 17 June 2022).
- OECD. Test No. 233: Sediment-Water Chironomid Life-Cycle Toxicity Test Using Spiked Water or Spiked Sediment. 2010. Available online: https://doi.org/10.1787/20745761 (accessed on 14 August 2022).
- Das, A.; Mahanwar, P. A Brief Discussion on Advances in Polyurethane Applications. Adv. Ind. Eng. Polym. Res. 2020, 3, 93–101. [Google Scholar] [CrossRef]
- Conkle, J.L.; Báez Del Valle, C.D.; Turner, J.W. Are We Underestimating Microplastic Contamination in Aquatic Environments? Environ. Manage. 2018, 61, 1–8. [Google Scholar] [CrossRef] [PubMed]
- European Parliament; Council of the European Union. Legislation for the Protection of Animals Used for Scientific Purposes. Available online: https://ec.europa.eu/environment/chemicals/lab_animals/legislation_en.htm (accessed on 2 November 2022).
- Silva, C.J.M.; Patrício Silva, A.L.; Campos, D.; Soares, A.M.V.M.; Pestana, J.L.T.; Gravato, C. Lumbriculus variegatus (Oligochaeta) Exposed to Polyethylene Microplastics: Biochemical, Physiological and Reproductive Responses. Ecotoxicol. Environ. Saf. 2021, 207, 111375. [Google Scholar] [CrossRef]
- Campos, D.; Gravato, C.; Fedorova, G.; Burkina, V.; Soares, A.M.V.M.; Pestana, J.L.T. Ecotoxicity of Two Organic UV-Filters to the Freshwater Caddisfly Sericostoma vittatum. Environ. Pollut. 2017, 228, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Bird, R.P.; Draper, H.H. Comparative Studies on Different Methods of Malonaldehyde Determination. Methods Enzymol. 1984, 105, 299–305. [Google Scholar] [CrossRef]
- De Coen, W.M.; Janssen, C.R. The Use of Biomarkers in Daphnia magna Toxicity Testing. IV. Cellular Energy Allocation: A New Methodology to Assess the Energy Budget of Toxicant-Stressed Daphnia Populations. J. Aquat. Ecosyst. Stress Recover. 1997, 6, 43–55. [Google Scholar] [CrossRef]
- Rodrigues, A.C.M.; Gravato, C.; Quintaneiro, C.; Barata, C.; Soares, A.M.V.M.; Pestana, J.L.T. Sub-Lethal Toxicity of Environmentally Relevant Concentrations of Esfenvalerate to Chironomus riparius. Environ. Pollut. 2015, 207, 273–279. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Savić-Zdravković, D.; Jovanović, B.; Đurđević, A.; Stojković-Piperac, M.; Savić, A.; Vidmar, J.; Milošević, D. An Environmentally Relevant Concentration of Titanium Dioxide (TiO2) Nanoparticles Induces Morphological Changes in the Mouthparts of Chironomus Tentans. Chemosphere 2018, 211, 489–499. [Google Scholar] [CrossRef]
- Drago, C.; Pawlak, J.; Weithoff, G. Biogenic Aggregation of Small Microplastics Alters Their Ingestion by a Common Freshwater Micro-Invertebrate. Front. Environ. Sci. 2020, 8, 264. [Google Scholar] [CrossRef]
- Su, L.; Deng, H.; Li, B.; Chen, Q.; Pettigrove, V.; Wu, C.; Shi, H. The Occurrence of Microplastic in Specific Organs in Commercially Caught Fishes from Coast and Estuary Area of East China. J. Hazard. Mater. 2019, 365, 716–724. [Google Scholar] [CrossRef]
- Prata, J.C.; Silva, A.L.P.; da Costa, J.P.; Dias-Pereira, P.; Carvalho, A.; Fernandes, A.J.S.; Mendes Da Costa, F.; Duarte, A.C.; Rocha-Santos, T. Microplastics in Internal Tissues of Companion Animals from Urban Environments. Animimal 2022, 12, 1979. [Google Scholar] [CrossRef]
- Sokolova, I.M.; Frederich, M.; Bagwe, R.; Lannig, G.; Sukhotin, A.A. Energy Homeostasis as an Integrative Tool for Assessing Limits of Environmental Stress Tolerance in Aquatic Invertebrates. Mar. Environ. Res. 2012, 79, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, A.S.; Sarmento, R.A.; Gravato, C.; Rodrigues, A.C.M.; Campos, D.; Simão, F.C.P.; Soares, A.M.V.M. Strategies of Cellular Energy Allocation to Cope with Paraquat-Induced Oxidative Stress: Chironomids vs Planarians and the Importance of Using Different Species. Sci. Total Environ. 2020, 741, 140443. [Google Scholar] [CrossRef] [PubMed]
- Hölker, F.; Stief, P. Adaptive Behaviour of Chironomid Larvae (Chironomus riparius) in Response to Chemical Stimuli from Predators and Resource Density. Behav. Ecol. Sociobiol. 2005, 58, 256–263. [Google Scholar] [CrossRef]
- Ford, A.T.; Ågerstrand, M.; Brooks, B.W.; Allen, J.; Bertram, M.G.; Brodin, T.; Dang, Z.; Duquesne, S.; Sahm, R.; Hoffmann, F.; et al. The Role of Behavioral Ecotoxicology in Environmental Protection. Environ. Sci. Technol. 2021, 55, 5620–5628. [Google Scholar] [CrossRef]






| Parameters | PU-MPs (mg/kg) | ||||
|---|---|---|---|---|---|
| 0 | 93.5 | 187.5 | 375 | 750 | |
| Emergence ratio (ER) ± SEM | 84 ± 7 | 92 ± 5 | 84 ± 7 | 100 ± 0 | 92 ± 5 |
| Mortality (M) ± SEM | 16 ± 7 | 8 ± 5 | 16 ± 7 | 0 ± 0 | 8 ± 5 |
| Emerged individuals/day (ED) ± SEM | 2.3 ± 0.9 | 3 ± 1 | 2.3 ± 0.9 | 2.8 ± 0.9 | 2.6 ± 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, S.A.M.; Rodrigues, A.C.M.; Rocha-Santos, T.; Silva, A.L.P.; Gravato, C. Effects of Polyurethane Small-Sized Microplastics in the Chironomid, Chironomus riparius: Responses at Organismal and Sub-Organismal Levels. Int. J. Environ. Res. Public Health 2022, 19, 15610. https://doi.org/10.3390/ijerph192315610
Silva SAM, Rodrigues ACM, Rocha-Santos T, Silva ALP, Gravato C. Effects of Polyurethane Small-Sized Microplastics in the Chironomid, Chironomus riparius: Responses at Organismal and Sub-Organismal Levels. International Journal of Environmental Research and Public Health. 2022; 19(23):15610. https://doi.org/10.3390/ijerph192315610
Chicago/Turabian StyleSilva, Sara A. M., Andreia C. M. Rodrigues, Teresa Rocha-Santos, Ana L. Patrício Silva, and Carlos Gravato. 2022. "Effects of Polyurethane Small-Sized Microplastics in the Chironomid, Chironomus riparius: Responses at Organismal and Sub-Organismal Levels" International Journal of Environmental Research and Public Health 19, no. 23: 15610. https://doi.org/10.3390/ijerph192315610
APA StyleSilva, S. A. M., Rodrigues, A. C. M., Rocha-Santos, T., Silva, A. L. P., & Gravato, C. (2022). Effects of Polyurethane Small-Sized Microplastics in the Chironomid, Chironomus riparius: Responses at Organismal and Sub-Organismal Levels. International Journal of Environmental Research and Public Health, 19(23), 15610. https://doi.org/10.3390/ijerph192315610