Ionophore Toxin Maduramicin Produces Haff Disease-Like Rhabdomyolysis in a Mouse Model



Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Animals
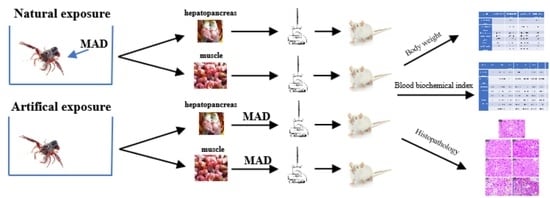
2.3. Crayfish Exposure to Maduramicin
2.4. Crayfish Sample Collection and Preparation
2.5. Mice Gavage Test
2.6. Mice Sample Collection
2.7. Biochemical Analysis
2.8. Histopathology Examination
2.9. Data Analysis
3. Results
3.1. General Observation
3.2. Effects of Crayfish Tissues with Maduramicin on the Body Weight of Mice
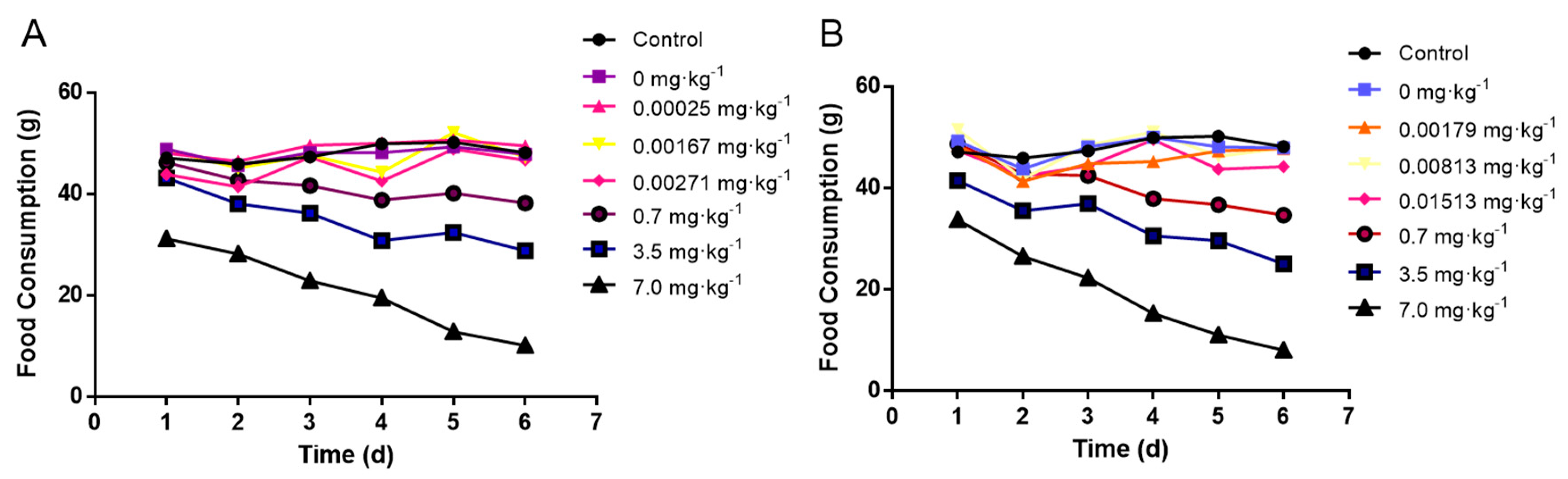
3.3. Effects of Crayfish Tissues with Maduramicin on Feed Consumption of Mice
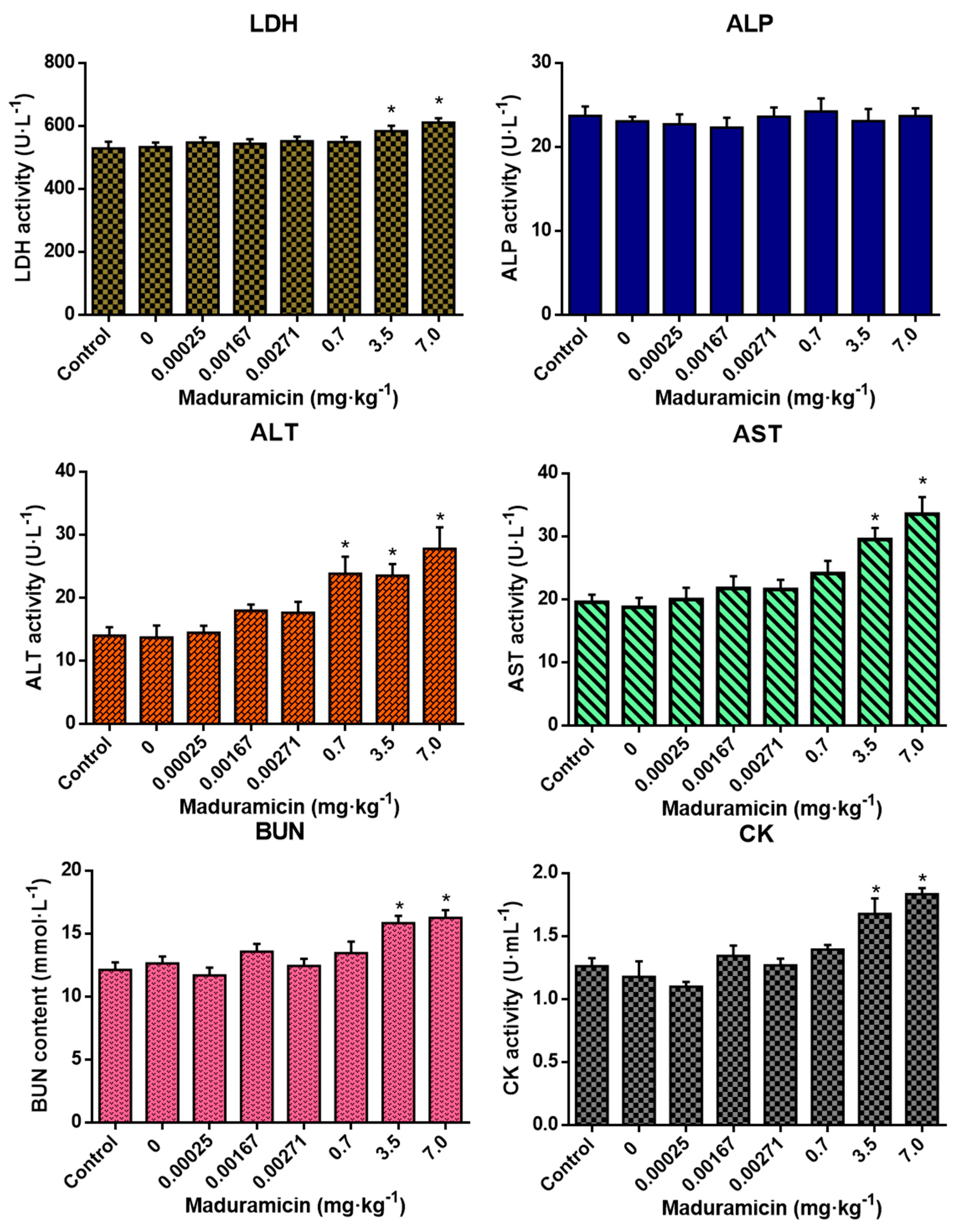
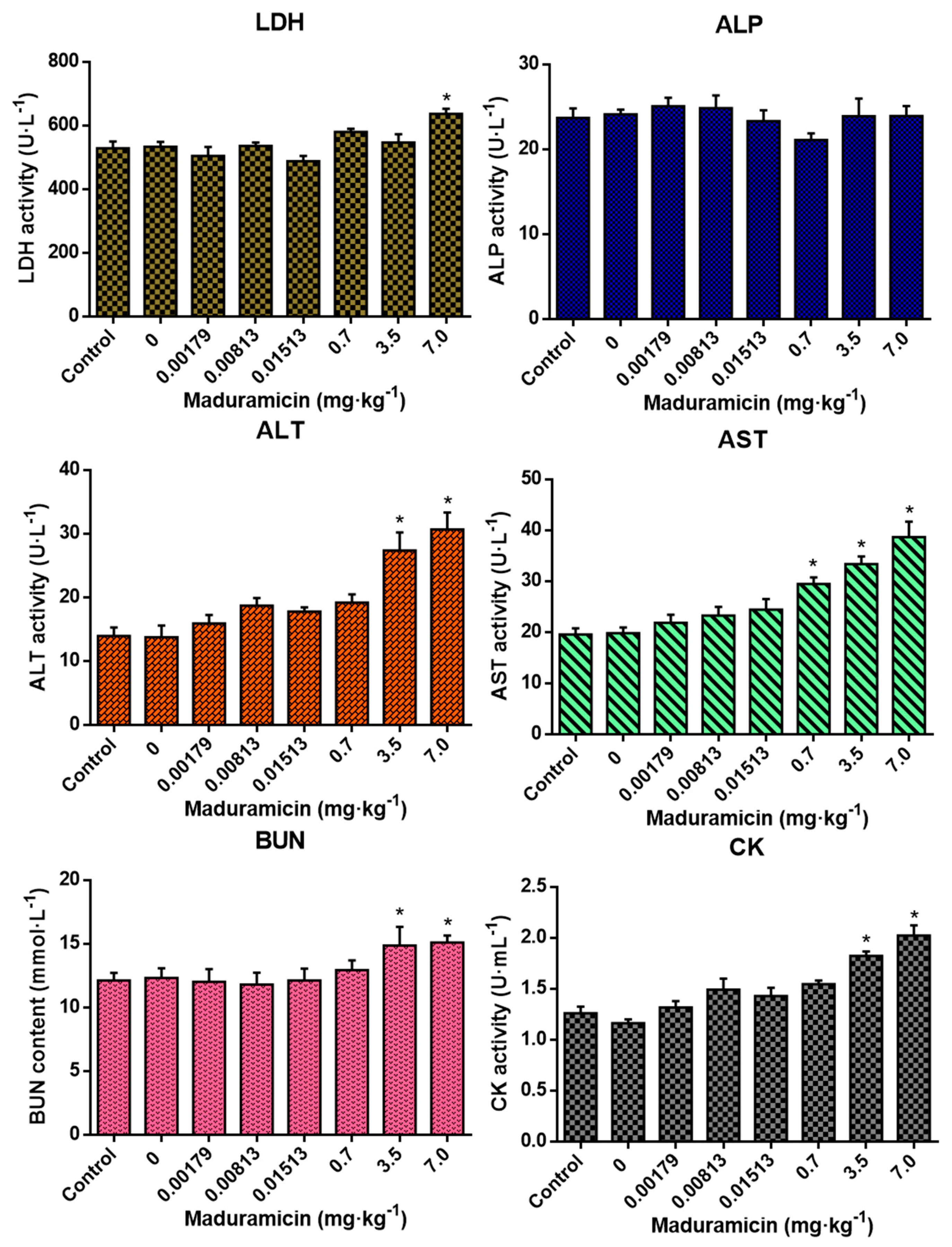
3.4. Effects of Crayfish Muscle with Maduramicin on Blood Biochemistry of Mice
3.5. Effect of Crayfish Hepatopancreas with Maduramicin on Blood Biochemistry of Mice
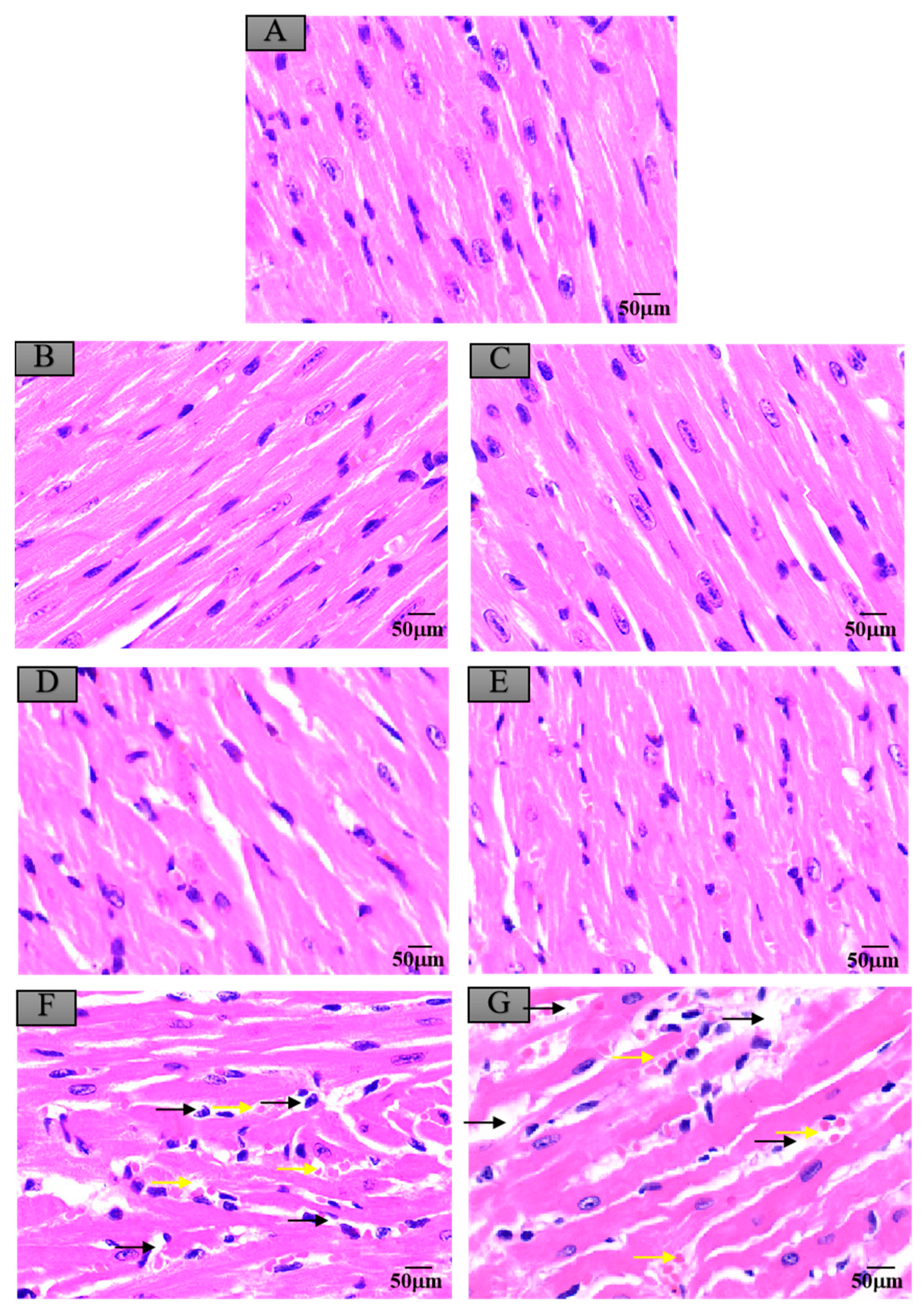
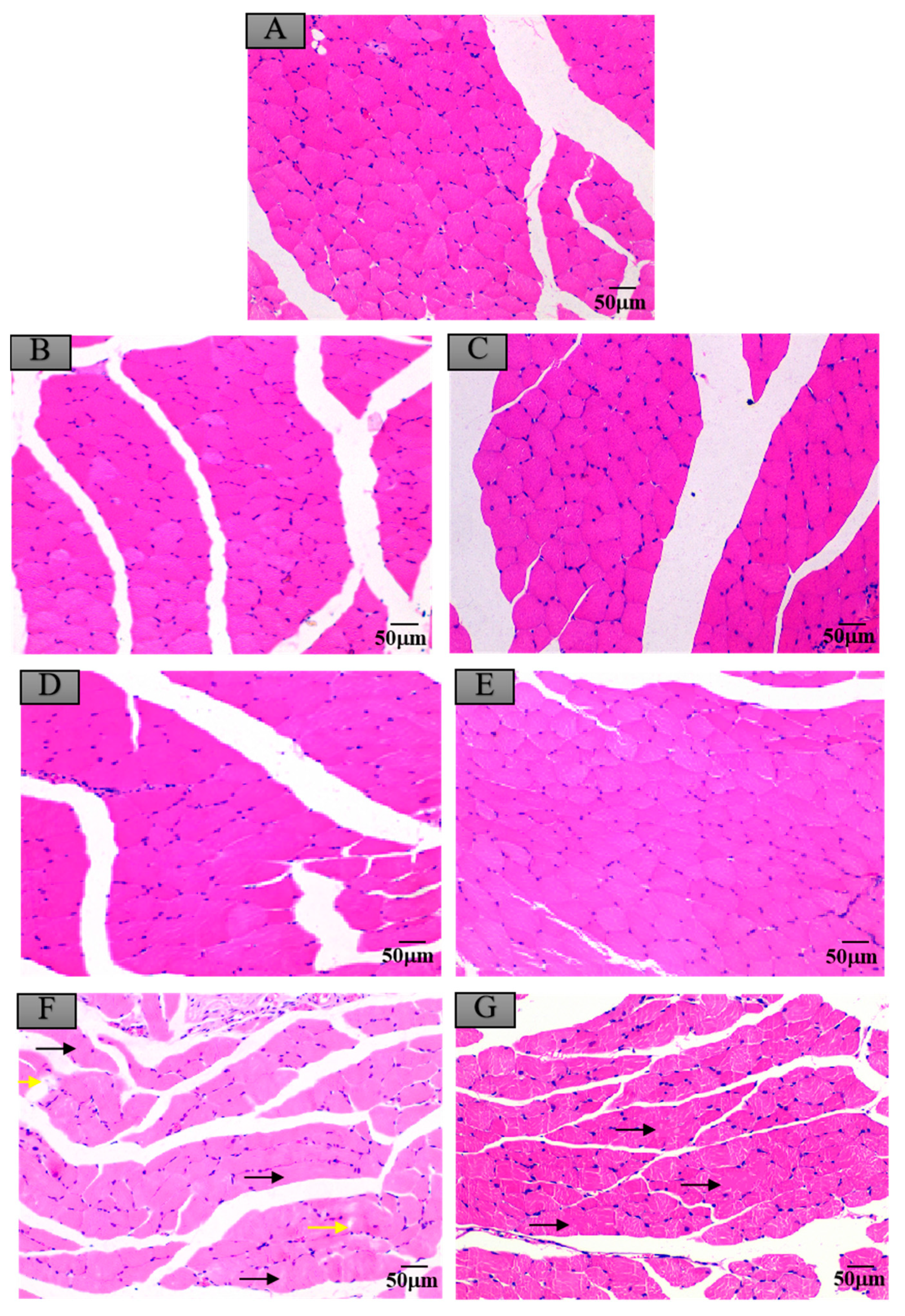
3.6. Histopathological Changes in Mice Exposed to Crayfish Muscle with Maduramicin
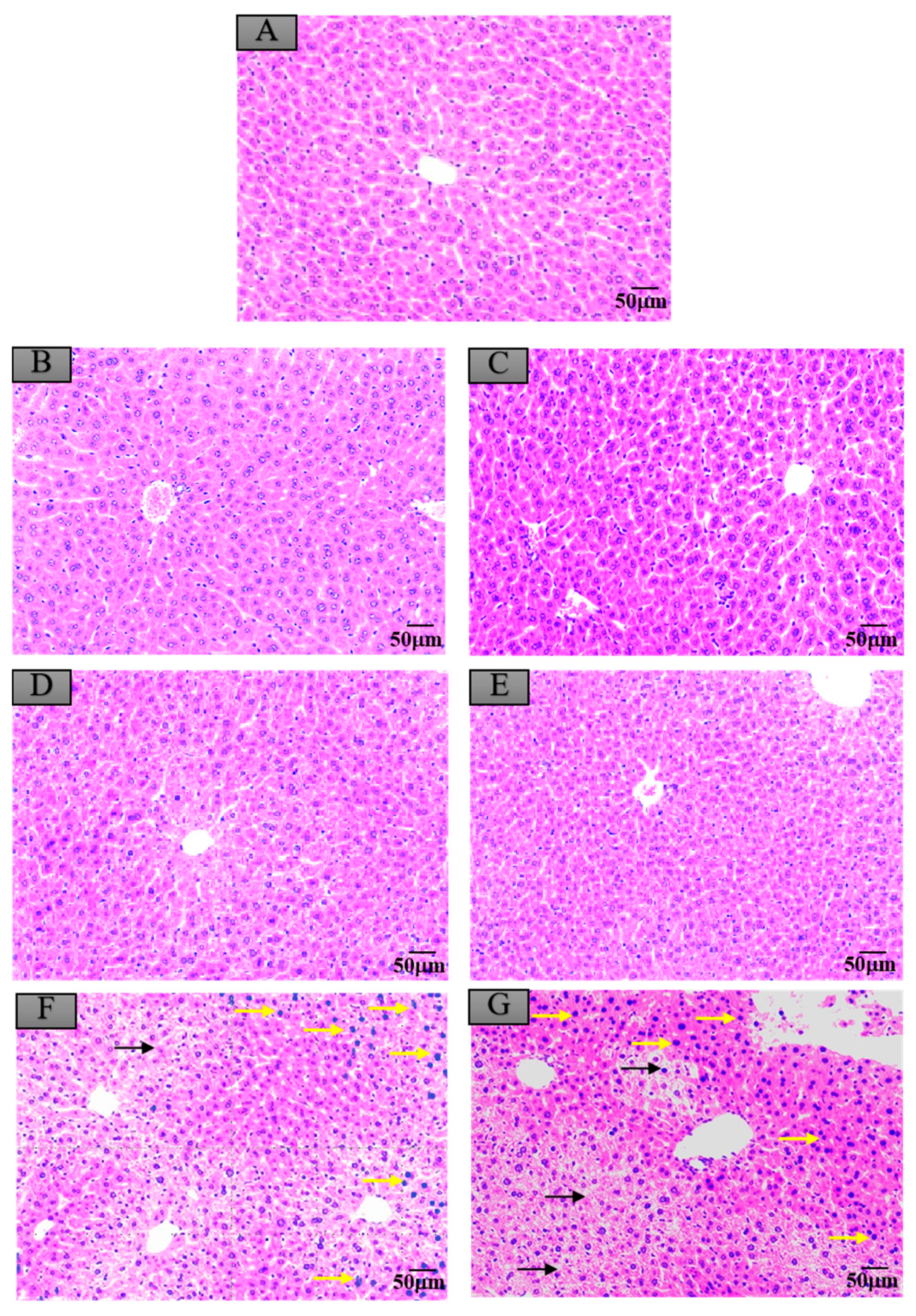
3.7. Histopathological Changes in Mice Exposed to Crayfish Hepatopancreas with Maduramicin
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Buchholz, U.; Mouzin, E.; Dickey, R.; Moolenaar, R.; Sass, N.; Mascola, L. Haff disease: From the baltic sea to the U.S. Shore. Emerg. Infect. Dis. 2000, 6, 192–195. [Google Scholar] [CrossRef] [PubMed]
- Diaz, J.H. Global incidence of rhabdomyolysis after cooked seafood consumption (haff disease). Clin. Toxicol. 2015, 53, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Tolesani Junior, O.; Roderjan, C.N.; do Carmo Neto, E.; Ponte, M.M.; Seabra, M.C.; Knibel, M.F. Haff disease associated with the ingestion of the freshwater fish mylossoma duriventre (pacu-manteiga). Rev. Bras. Ter. Intensiv. 2013, 25, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Pei, P.; Li, X.Y.; Lu, S.S.; Liu, Z.; Wang, R.; Lu, X.C.; Lu, K. The emergence, epidemiology, and etiology of haff disease. Biomed. Environ. Sci. 2019, 32, 769–778. [Google Scholar] [PubMed]
- Berlin, R. Haff disease in sweden. Acta Med. Scand. 1948, 129, 560–572. [Google Scholar] [CrossRef] [PubMed]
- Basciarina, A.A.; Kurockina, Z.Z. Haff disease or lake juksov disease (acute alimentary myositis in the karelian-finnish soviet state). Ann. D’igiene 1950, 60, 127–131. [Google Scholar] [PubMed]
- Herman, L.L.; Bies, C. Haff disease: Rhabdomyolysis after eating buffalo fish. West. J. Emerg. Med. 2014, 15, 664–666. [Google Scholar] [CrossRef]
- Diaz, J.H. Rhabdomyolysis after cooked seafood consumption (haff disease) in the united states vs china. Ochsner J. 2015, 15, 170–175. [Google Scholar]
- Dos Santos, M.C.; de Albuquerque, B.C.; Pinto, R.C.; Aguiar, G.P.; Lescano, A.G.; Santos, J.H.; Alecrim, M. Outbreak of haff disease in the brazilian amazon. Pan Am. J. Public Health 2009, 26, 469–470. [Google Scholar] [CrossRef]
- Yuan, Y.; Chen, Q.T. Clinical analysis of six cases with Haff disease after eating crayfish. Natl. Med. J. China 2001, 81, 1530–1531. (In Chinese) [Google Scholar]
- Huang, X.; Li, Y.; Huang, Q.; Liang, J.; Liang, C.; Chen, B.; Lu, L.; Deng, X.; Chen, Z.; Zhang, Y.; et al. A past haff disease outbreak associated with eating freshwater pomfret in south china. BMC Public Health 2013, 13, 447. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yuan, B.J.; Xie, G.X.; Zhen, S.Q.; Zhou, Y.J.; Shao, B.; Zhang, J.; Ji, H.; Wu, Y.N. Outbreak of haff disease caused by consumption of crayfish (procambarus clarkii), nanjing, jiangsu province, china. Food Control 2016, 59, 690–694. [Google Scholar] [CrossRef]
- Feng, G.; Luo, Q.; Zhuang, P.; Guo, E.; Yao, Y.; Gao, Z. Haff disease complicated by multiple organ failure after crayfish consumption: A case study. Rev. Bras. Ter. Intensiva 2014, 26, 407–409. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Xie, G.; Li, X.; Jiang, Y.; Jin, D.; Zhou, Y.; Dai, Y.; Zhen, S.; Sun, G. Outbreak of haff disease caused by consumption of crayfish (procambarus clarkii) in nanjing, china. Clin. Toxicol. 2019, 57, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Noack, S.; Chapman, H.D.; Selzer, P.M. Anticoccidial drugs of the livestock industry. Parasitol. Res. 2019, 118, 2009–2026. [Google Scholar] [CrossRef]
- Fourie, N.; Bastianello, S.S.; Prozesky, L.; Nel, P.W.; Kellerman, T.S. Cardiomyopathy of ruminants induced by the litter of poultry fed on rations containing the ionophore antibiotic, maduramicin. I. Epidemiology, clinical signs and clinical pathology. Onderstepoort J. Vet. 1991, 58, 291–296. [Google Scholar]
- Bastianello, S.S.; Fourie, N.; Prozesky, L.; Nel, P.W.; Kellermann, T.S. Cardiomyopathy of ruminants induced by the litter of poultry fed on rations containing the ionophore antibiotic, maduramicin. II. Macropathology and histopathology. Onderstepoort J. Vet. 1995, 62, 5–18. [Google Scholar]
- Sharma, N.; Bhalla, A.; Varma, S.; Jain, S.; Singh, S. Toxicity of maduramicin. Emerg. Med. J. 2005, 22, 880–882. [Google Scholar] [CrossRef]
- Shimshoni, J.A.; Britzi, M.; Pozzi, P.S.; Edery, N.; Berkowitz, A.; Bouznach, A.; Cuneah, O.; Soback, S.; Bellaiche, M.; Younis, A.; et al. Acute maduramicin toxicosis in pregnant gilts. Food Chem. Toxicol. 2014, 68, 283–289. [Google Scholar] [CrossRef]
- Dorne, J.L.; Fernandez-Cruz, M.L.; Bertelsen, U.; Renshaw, D.W.; Peltonen, K.; Anadon, A.; Feil, A.; Sanders, P.; Wester, P.; Fink-Gremmels, J. Risk assessment of coccidostatics during feed cross-contamination: Animal and human health aspects. Toxicol. Appl. Pharm. 2013, 270, 196–208. [Google Scholar] [CrossRef]
- Gao, X.; Zheng, Y.; Peng, L.; Ruan, X.; Ji, H.; Qiu, Y.; Liu, X.; Teng, P.; Guo, D.; Jiang, S. Maduramicin induces apoptosis in chicken myocardial cells via intrinsic and extrinsic pathways. Toxicol. Vitr. 2018, 50, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Peng, L.; Ruan, X.; Chen, X.; Ji, H.; Ma, J.; Ni, H.; Jiang, S.; Guo, D. Transcriptome profile analysis reveals cardiotoxicity of maduramicin in primary chicken myocardial cells. Arch. Toxicol. 2018, 92, 1267–1281. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.J.; Zhou, H.J.; Gu, W. Clinical characteristics of patients with haff disease after eating crayfish. World J. Emerg. Med. 2019, 10, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Blain, M.; Garrard, A.; Poppenga, R.; Chen, B.; Valento, M.; Halliday Gittinger, M. Survival after severe rhabdomyolysis following monensin ingestion. J. Med. Toxicol. 2017, 13, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Kouyoumdjian, J.A.; Morita, M.P.; Sato, A.K.; Pissolatti, A.F. Fatal rhabdomyolysis after acute sodium monensin (rumensin) toxicity: Case report. Arq. Neuro-Psiquiat. 2001, 59, 596–598. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fang, J.; Wu, S.; Ma, K.; Li, H.; Yan, X.; Dong, F. Identification and quantification of salinomycin in intoxicated human plasma by liquid chromatography-electrospray tandem mass spectrometry. Anal. Bioanal. Chem. 2010, 398, 955–961. [Google Scholar] [CrossRef]
- Oehme, F.W.; Pickrell, J.A. An analysis of the chronic oral toxicity of polyether ionophore antibiotics in animals. Vet. Hum. Toxicol. 1999, 41, 251–257. [Google Scholar]
- Hamscher, G.; Bachour, G. Veterinary drugs in the environment: Current knowledge and challenges for the future. J. Agric. Food Chem. 2018, 66, 751–752. [Google Scholar] [CrossRef]
- Hu, Y.N.; Cheng, H.F. Health risk from veterinary antimicrobial use in china’s food animal production and its reduction. Environ. Pollut. 2016, 219, 993–997. [Google Scholar] [CrossRef]
- Iglesias, A.; Nebot, C.; Vazquez, B.I.; Miranda, J.M.; Abuin, C.M.F.; Cepeda, A. Detection of veterinary drug residues in surface waters collected nearby farming areas in galicia, north of spain. Environ. Sci. Pollut. Res. 2014, 21, 2367–2377. [Google Scholar] [CrossRef]
- Kim, S.C.; Carlson, K. Occurrence of ionophore antibiotics in water and sediments of a mixed-landscape watershed. Water Res. 2006, 40, 2549–2560. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.; Carlson, K.H. Occurrence of beta-lactam and polyether ionophore antibiotics in lagoon water and animal manure. Sci. Total Environ. 2018, 640–641, 1346–1353. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.; Krogh, K.A.; Bjorklund, E.; Brandt, A.; Halling-Sorensen, B. Environmental risk assessment of ionophores. Trac-Trend. Anal. Chem. 2009, 28, 534–542. [Google Scholar] [CrossRef]
- Zhang, B.; Yang, G.; Yu, X.; Mao, H.; Xing, C.; Liu, J. Haff disease after eating crayfish in east china. Intern. Med. 2012, 51, 487–489. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chan, T.Y. The emergence and epidemiology of haff disease in china. Toxins 2016, 8, 359. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Peng, L.; Gong, N.; Xue, C.; Wang, W.; Jiang, J. A retrospective analysis of crayfish-related rhabdomyolysis (haff disease). Emerg. Med. Int. 2019, 2019, 4209745. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Xu, M.J.; Li, W.W.; Han, H.H.; Liu, J.K.; Fu, P.; Xu, L.Z.; Ouyang, Y.Y.; You, X.Y.; Chen, J.; et al. Retrospective case analysis of crayfish-transmitted haff disease in china during 2016-2017. Food Control 2019, 104, 181–186. [Google Scholar] [CrossRef]
- Shinzato, T.; Furusu, A.; Nishino, T.; Abe, K.; Kanda, T.; Maeda, T.; Kohno, S. Cowfish (umisuzume, lactoria diaphana) poisoning with rhabdomyolysis. Intern. Med. 2008, 47, 853–856. [Google Scholar] [CrossRef]
- Chen, X.F.; Lin, J.W.; Pan, T.M.; Cao, M.J.; Shi, C.L.; Cai, Q.F.; Liu, G.M. Investigation of the hazardous substance causing crayfish-induced rhabdomyolysis via a mouse model, a hemolysis assay, and a cytotoxicity assay. Fish. Sci. 2015, 81, 551–558. [Google Scholar] [CrossRef][Green Version]
- Yopasa-Arenas, A.; Fostier, A.H. Exposure of brazilian soil and groundwater to pollution by coccidiostats and antimicrobial agents used as growth promoters. Sci. Total Environ. 2018, 644, 112–121. [Google Scholar] [CrossRef]
- Alonso, L.L.; Demetrio, P.M.; Capparelli, A.L.; Marino, D.J.G. Behavior of ionophore antibiotics in aquatic environments in argentina: The distribution on different scales in water courses and the role of wetlands in depuration. Environ. Int. 2019, 133, 105144. [Google Scholar] [CrossRef] [PubMed]
- Bak, S.A.; Björklund, E. Occurrence of ionophores in the danish environment. Antibiotics 2014, 3, 564–571. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances Used in Animal Feed (FEEDAP). Scientific opinion on safety and efficacy of cygro® 10g (maduramicin ammonium α) for chickens for fattening. EFSA J. 2011, 9, 1952. [Google Scholar] [CrossRef]
- He, F.; Ni, J.; Huang, J.A.; Liu, Y.; Wu, C.; Wang, J. Clinical features of haff disease and myositis after the consumption of boiled brackish water crayfish: A retrospective study of 96 cases at a single centre. Intern. Emerg. Med. 2018, 13, 1265–1271. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Oda, S.; Matsubara, A.; Akai, S.; Yokoi, T. Establishment and characterization of a mouse model of rhabdomyolysis by coadministration of statin and fibrate. Toxicol. Lett. 2019, 307, 49–58. [Google Scholar] [CrossRef]
- Matsubara, A.; Oda, S.; Akai, S.; Tsuneyama, K.; Yokoi, T. Establishment of a drug-induced rhabdomyolysis mouse model by co-administration of ciprofloxacin and atorvastatin. Toxicol. Lett. 2018, 291, 184–193. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Dose (mg kg−1) | Initial Weight (g) | Final Weight (g) | Body Weight Gain (g) |
|---|---|---|---|---|
| Blank control group | 0 | 24.46 ± 2.35 | 30.24 ± 4.73 | 5.78 |
| Muscle control group | 0 | 24.97 ± 1.62 | 30.29 ± 2.58 | 5.32 |
| Natural exposure group | 0.00025 | 24.35 ± 1.20 | 30.73 ± 3.16 | 6.38 |
| 0.00167 | 24.39 ± 1.82 | 28.62 ± 2.03 | 4.23 | |
| 0.00271 | 24.15 ± 2.20 | 28.47 ± 3.59 | 4.32 | |
| Artificial exposure group | 0.7 | 25.27 ± 1.36 | 28.11 ± 3.02 * | 2.84 |
| 3.5 | 24.42 ± 1.18 | 24.45 ± 2.59 * | 0.03 | |
| 7.0 | 24.54 ± 1.56 | 21.40 ± 3.84 * | −3.14 |
| Group | Dose (mg kg−1) | Initial Weight (g) | Final Weight (g) | Body Weight Gain (g) |
|---|---|---|---|---|
| Blank control group | 0 | 24.46 ± 2.35 | 30.24 ± 4.73 | 5.78 |
| Hepatopancreas control group | 0 | 23.82 ± 1.12 | 29.10 ± 2.85 | 5.28 |
| Natural exposure group | 0.00179 | 24.70 ± 1.02 | 30.54 ± 3.52 | 5.84 |
| 0.00813 | 24.14 ± 1.44 | 28.50 ± 3.23 | 4.36 | |
| 0.01513 | 23.79 ± 0.55 | 27.89 ± 3.29 | 4.10 | |
| Artificial exposure group | 0.7 | 24.69 ± 1.49 | 26.62 ± 3.18 * | 1.93 |
| 3.5 | 24.38 ± 1.58 | 23.32 ± 3.52 * | −1.06 | |
| 7.0 | 24.87 ± 1.18 | 21.13 ± 1.04 * | −3.74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Song, X.; Zuo, R.; Yang, D.; Ji, C.; Ji, H.; Peng, L.; Qiu, Y.; Guo, D.; Jiang, S. Ionophore Toxin Maduramicin Produces Haff Disease-Like Rhabdomyolysis in a Mouse Model. Int. J. Environ. Res. Public Health 2020, 17, 7882. https://doi.org/10.3390/ijerph17217882
Gao X, Song X, Zuo R, Yang D, Ji C, Ji H, Peng L, Qiu Y, Guo D, Jiang S. Ionophore Toxin Maduramicin Produces Haff Disease-Like Rhabdomyolysis in a Mouse Model. International Journal of Environmental Research and Public Health. 2020; 17(21):7882. https://doi.org/10.3390/ijerph17217882
Chicago/Turabian StyleGao, Xiuge, Xinhao Song, Runan Zuo, Dan Yang, Chunlei Ji, Hui Ji, Lin Peng, Yawei Qiu, Dawei Guo, and Shanxiang Jiang. 2020. "Ionophore Toxin Maduramicin Produces Haff Disease-Like Rhabdomyolysis in a Mouse Model" International Journal of Environmental Research and Public Health 17, no. 21: 7882. https://doi.org/10.3390/ijerph17217882
APA StyleGao, X., Song, X., Zuo, R., Yang, D., Ji, C., Ji, H., Peng, L., Qiu, Y., Guo, D., & Jiang, S. (2020). Ionophore Toxin Maduramicin Produces Haff Disease-Like Rhabdomyolysis in a Mouse Model. International Journal of Environmental Research and Public Health, 17(21), 7882. https://doi.org/10.3390/ijerph17217882