Ciguatoxins: Cyclic Polyether Modulators of Voltage-gated Iion Channel Function

Abstract
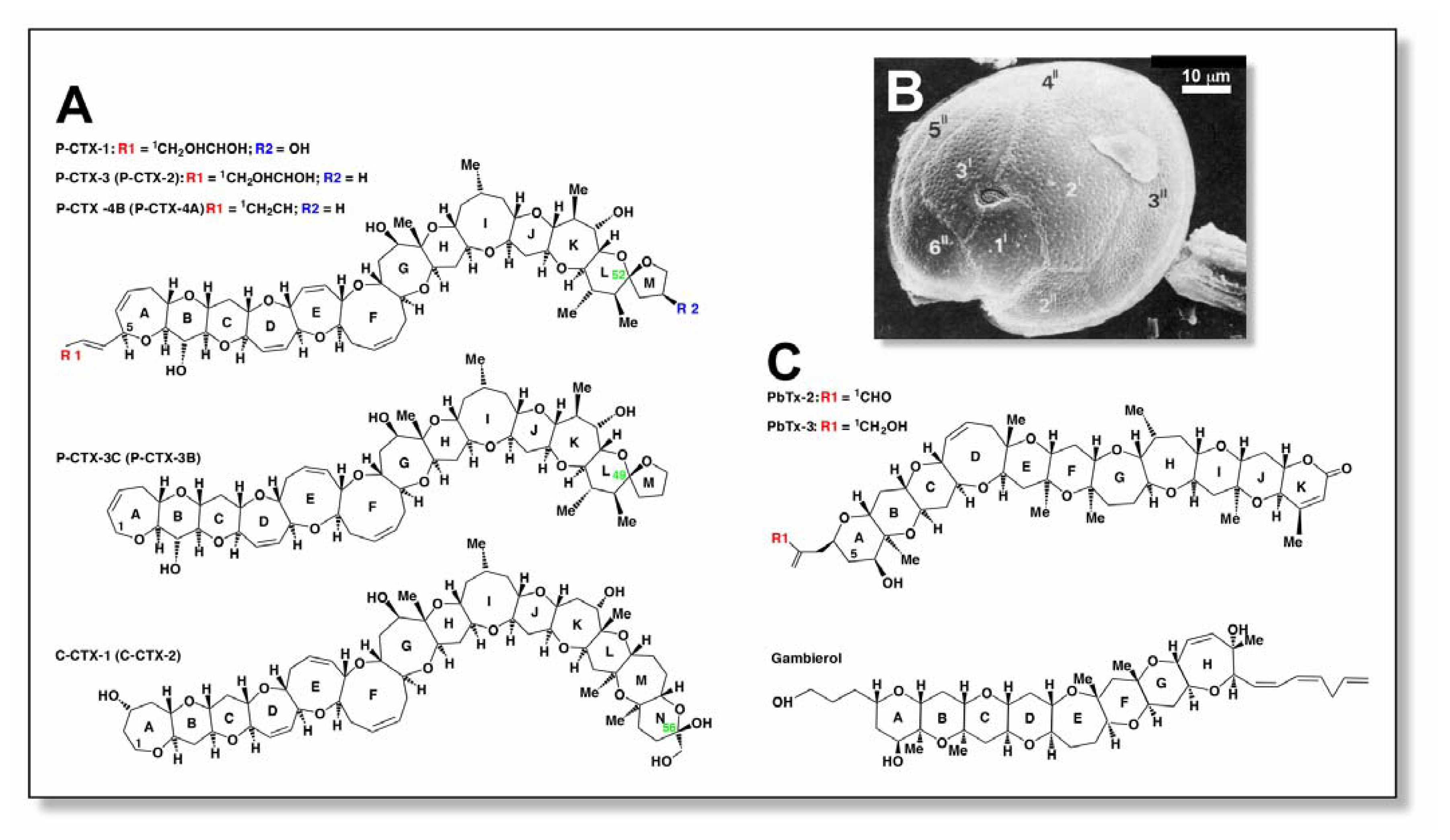
:1. Ciguatoxins: Sources and structures
2. Ciguatera poisoning in humans
3. Pharmacology of Ciguatoxins
3.1. Cardiovascular effects of ciguatoxins
3.2 Gastrointestinal effects of ciguatoxins
3.3. Effects of ciguatoxin on other smooth muscle
3.4. Action of ciguatoxins on ganglionic and neuromuscular transmission
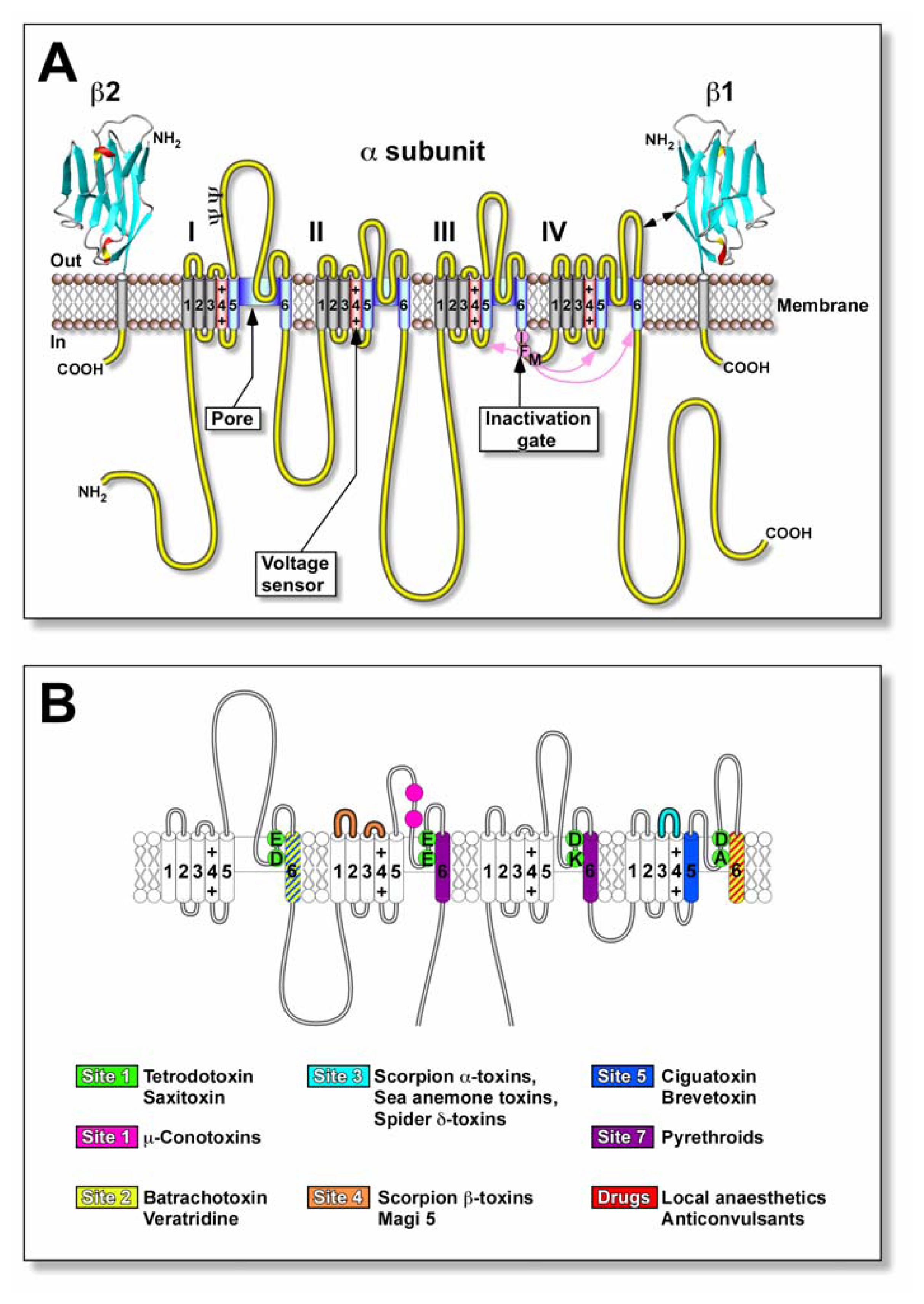
4. Actions of ciguatoxins on voltage-gated ion channels
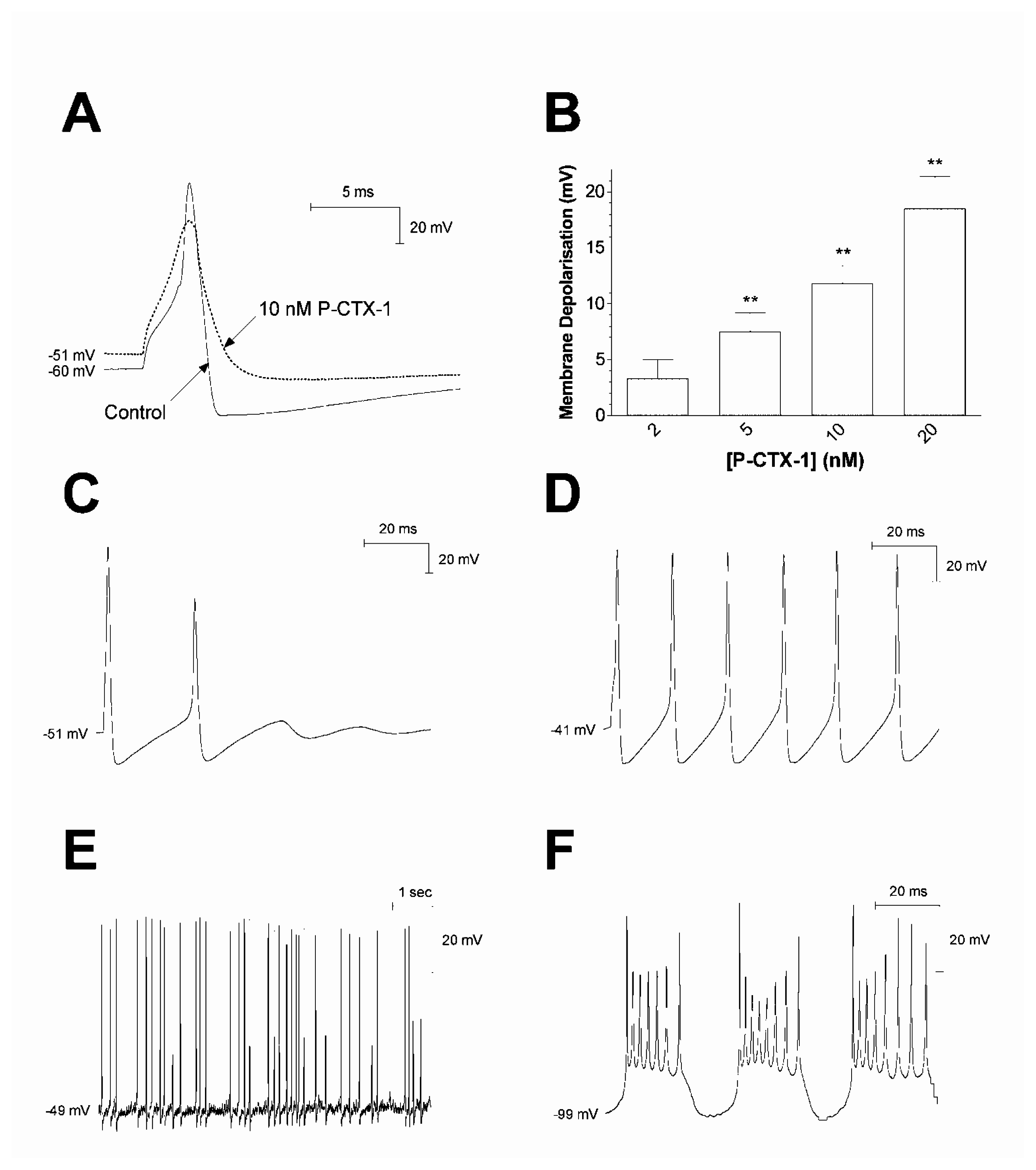
4.1. Ciguatoxin enhances neuronal excitability
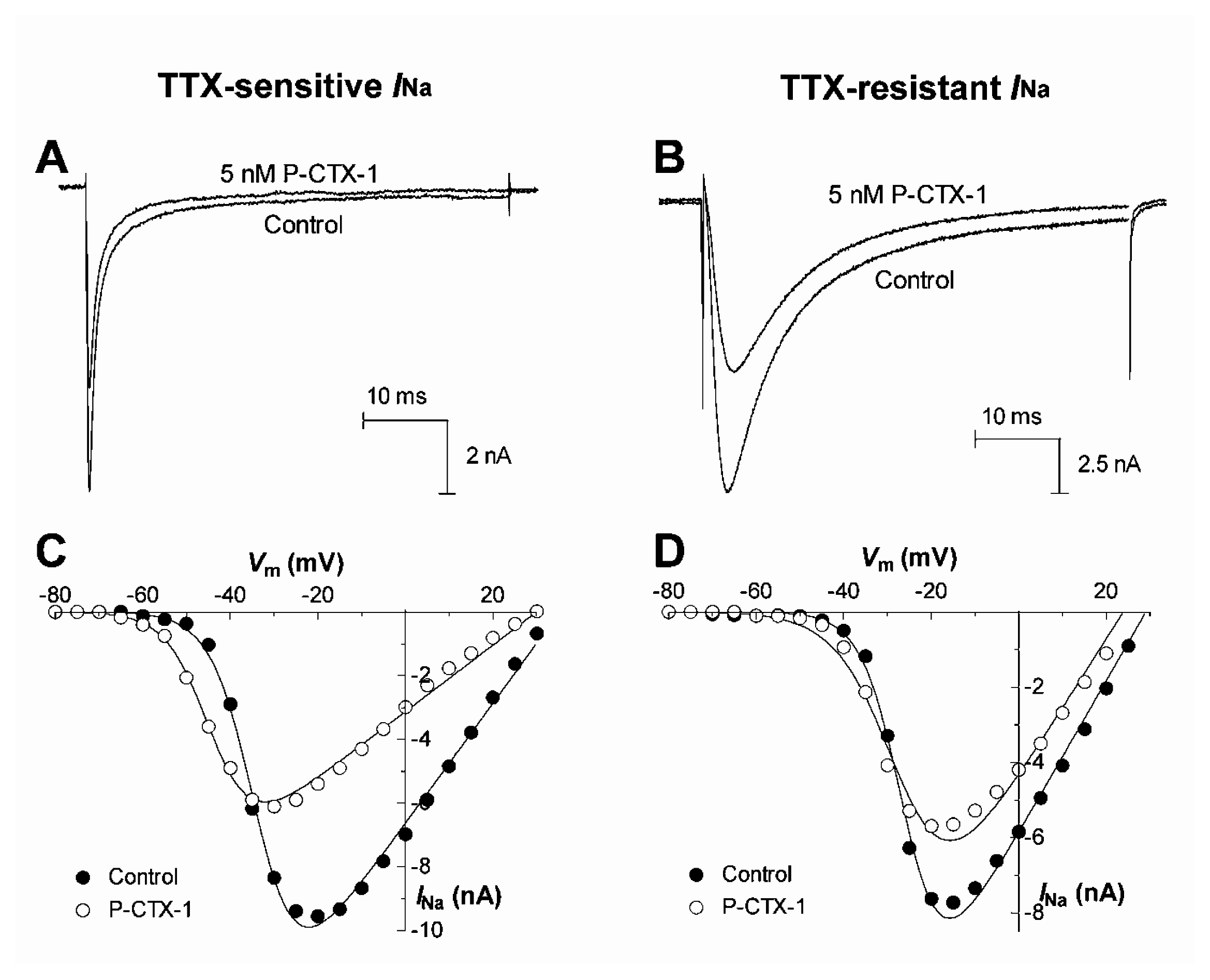
4.2. Mode(s) of activation of Nav channels by ciguatoxin
4.3. Ciguatoxin competes for neurotoxin receptor site 5 on Nav channels
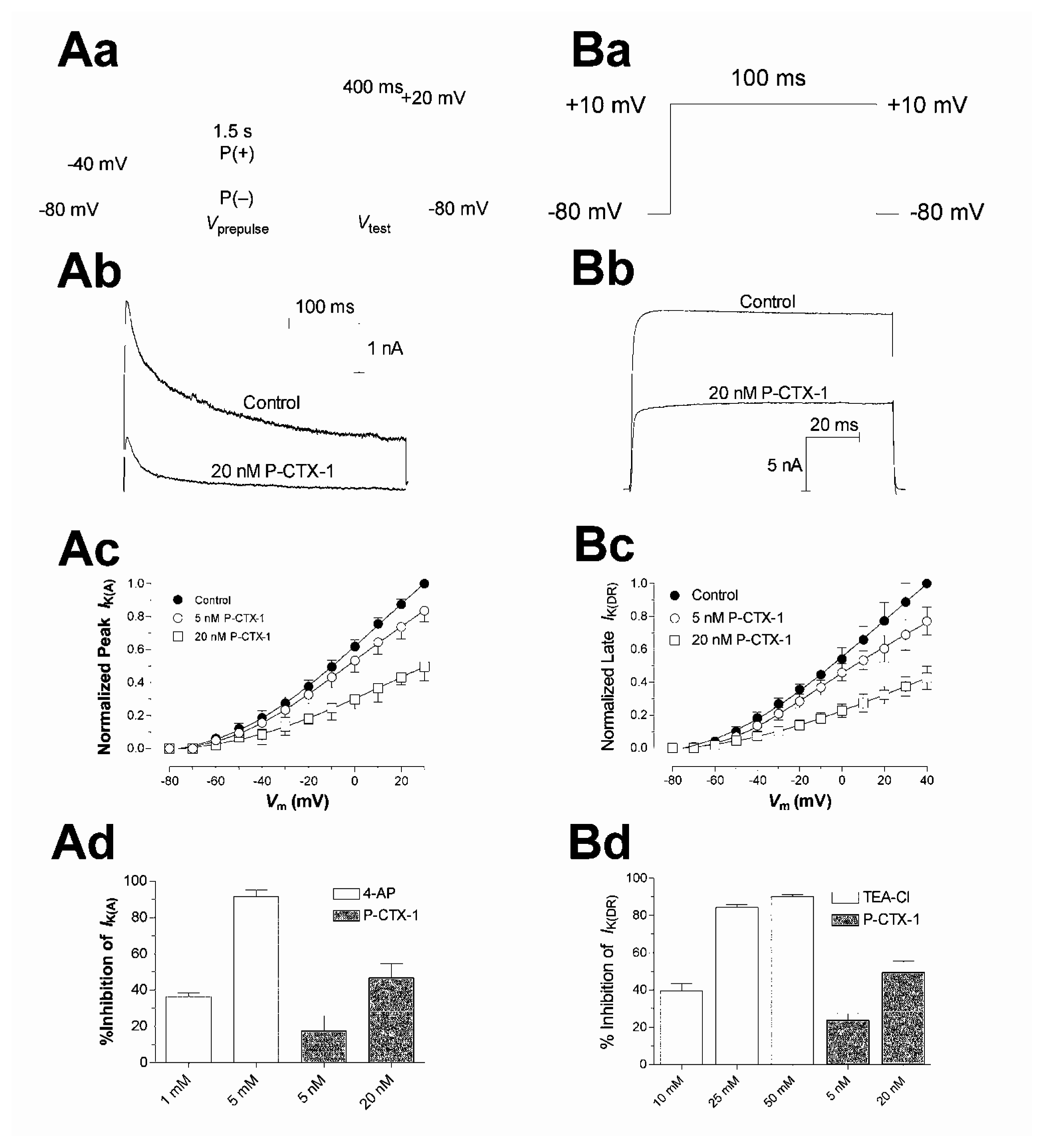
5. Actions of ciguatoxin on voltage-gated potassium channels
6. Other Na+-dependent actions of ciguatoxin
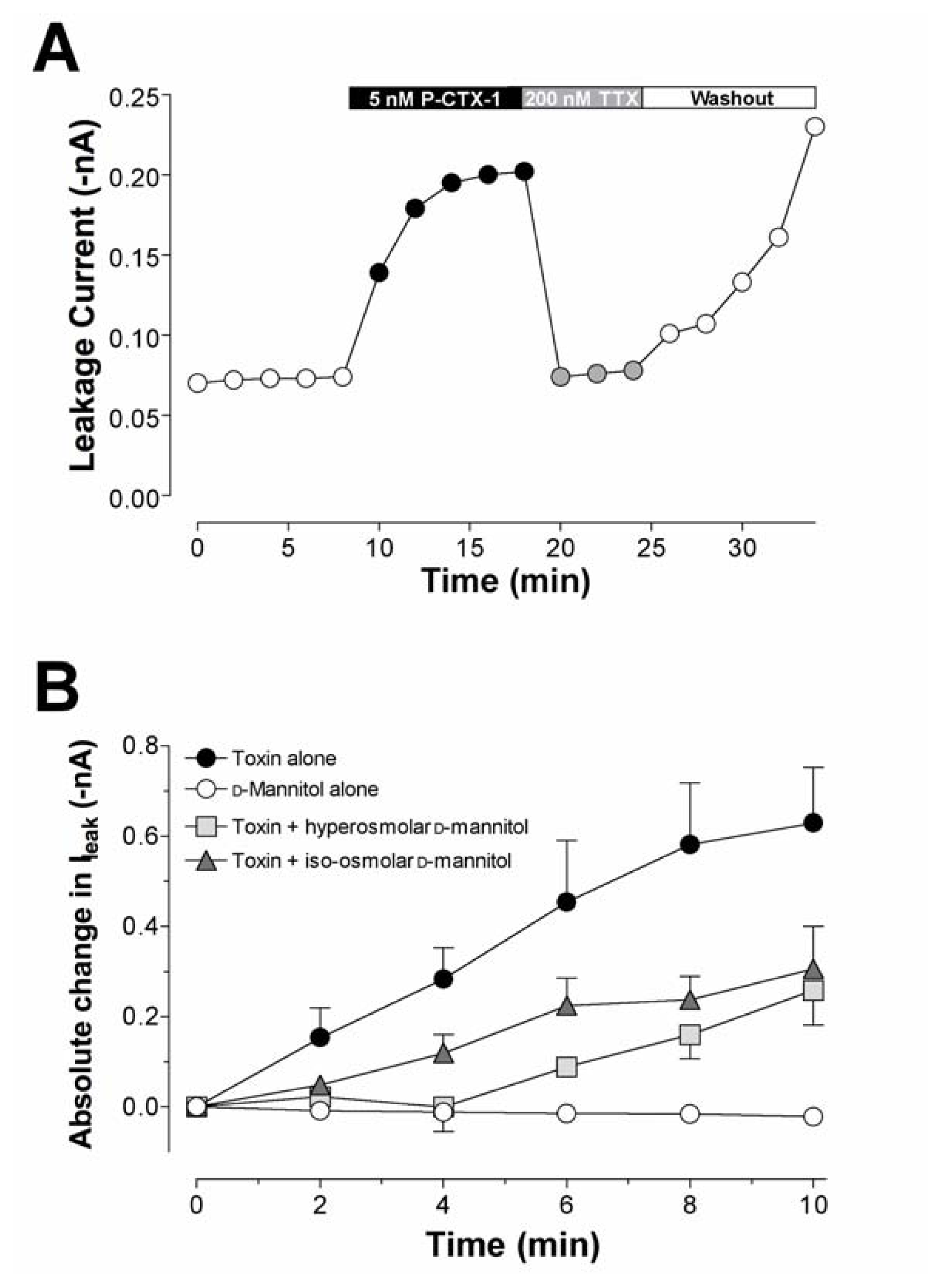
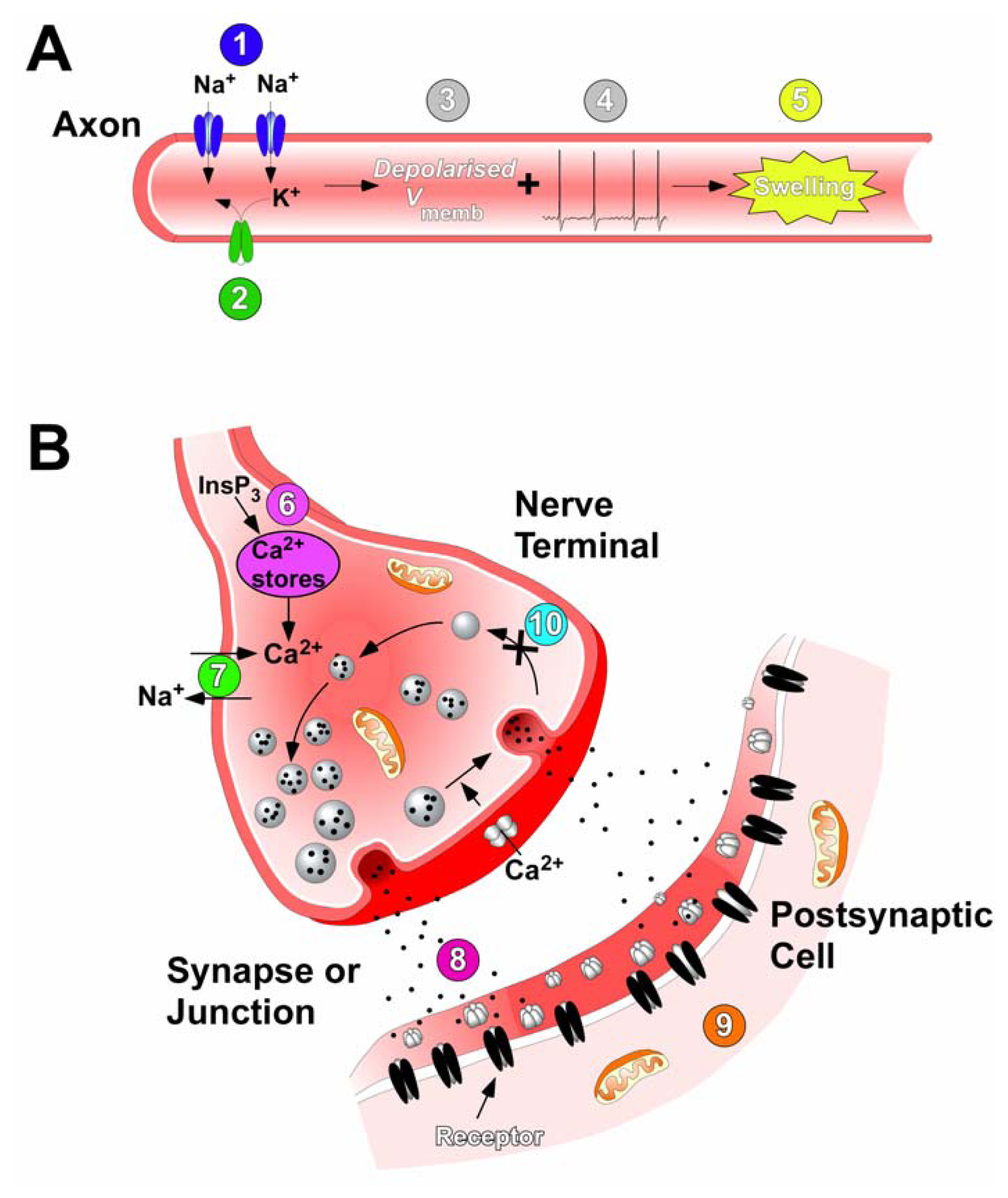
6.1. Ciguatoxin induced swelling of cells
6.2. Action of ciguatoxin on intracellular Ca2+ stores and the Na+ – Ca2+ exchange mechanism
7. Detection of ciguatoxins
8. Treatment of ciguatera
9. Conclusions
Acknowledgements
Abbreviations
| Ch | acetylcholine |
| 4-AP | 4-aminopyridine |
| [Ca2+]o | extracellular concentration of Ca2+ |
| Cav channel | voltage-gated Ca2+ channel |
| CTX | ciguatoxin |
| C-CTX | Caribbean Sea ciguatoxin |
| DRG | dorsal root ganglion |
| EJP | excitatory junction potential |
| EPP | endplate potential |
| I-CTX | Indian Ocean ciguatoxin |
| InsIP3 | inositol 1,4,5-triphosphate |
| KA channel | transient ‘A-type’ K+ channel |
| [K+]o | extracellular concentration of K+ |
| KDR channel | delayed-rectifier K+ channel |
| Kv channel | voltage-gated potassium channel |
| LD50 | median lethal dose |
| MEPP | miniature endplate potential |
| mAChR | muscarinic acetylcholine receptor |
| [Na+]i | intracellular concentration of Na+ |
| Nav channel | voltage-gated sodium channel |
| nAChR | nicotinic ACh receptor |
| PbTx | brevetoxin |
| P-CTX | Pacific Ocean ciguatoxin |
| STX | saxitoxin |
| TEA | tetraethylammonium |
| TTX | tetrodotoxin |
- Samples Availability: Available from Dr Richard J. Lewis.
References
- Miyahara, J. T.; Akau, C. K.; Yasumoto, T. Effects of ciguatoxin and maitotoxin on the isolated guinea pig atria. Res. Commun. Chem. Pathol. Pharmacol 1979, 25, 177–180. [Google Scholar]
- Adachi, R.; Fukuyo, Y. The thecal structure of a marine toxic dinoflagellate Gambierdiscus toxicus collected in a ciguatera endemic area. Bull. Jap. Soc. Scientific Fisheries 1979, 45, 67–71. [Google Scholar]
- Lewis, R. J.; Vernoux, J. P.; Brereton, I. M. Structure of Caribbean ciguatoxin isolated from Caranx latus. J. Am. Chem. Soc 1998, 120, 5914–5920. [Google Scholar]
- Murata, M.; Legrand, A.-M.; Ishibashi, Y.; Yasumoto, T. Structures and configurations of ciguatoxin from the moray eel Gymnothorax javanicus and its likely precursor from the dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc 1990, 112, 4380–4386. [Google Scholar]
- Lewis, R. J.; Sellin, M.; Poli, M. A.; Norton, R. S.; MacLeod, J. K.; Sheil, M. M. Purification and characterization of ciguatoxins from moray eel (Lycodontis javanicus, Muraenidae). Toxicon 1991, 29, 1115–1127. [Google Scholar]
- Lewis, R. J.; Sellin, M. Multiple ciguatoxins in the flesh of fish. Toxicon 1992, 30, 915–919. [Google Scholar]
- Lewis, R. J. The changing face of ciguatera. Toxicon 2001, 39, 97–106. [Google Scholar]
- Hamilton, B.; Hurbungs, M.; Jones, A.; Lewis, R. J. Multiple ciguatoxins present in Indian Ocean reef fish. Toxicon 2002, 40, 1347–1353. [Google Scholar]
- Hamilton, B.; Hurbungs, M.; Vernoux, J. P.; Jones, A.; Lewis, R. J. Isolation and characterisation of Indian Ocean ciguatoxin. Toxicon 2002, 40, 685–693. [Google Scholar]
- Pottier, I.; Hamilton, B.; Jones, A.; Lewis, R. J.; Vernoux, J. P. Identification of slow and fast-acting toxins in a highly ciguatoxic barracuda (Sphyraena barracuda) by HPLC/MS and radiolabelled ligand binding. Toxicon 2003, 42, 663–672. [Google Scholar]
- Vernoux, J. P.; Lewis, R. J. Isolation and characterisation of Caribbean ciguatoxins from the horse-eye jack (Caranx latus). Toxicon 1997, 35, 889–900. [Google Scholar]
- Murata, M.; Legrand, A.-M.; Ishibashi, Y.; Yasumoto, T. Structures and configurations of ciguatoxin and its congener. J. Am. Chem. Soc 1989, 111, 8929–8931. [Google Scholar]
- Satake, M.; Murata, M.; Yasumoto, T. The structure of CTX3c, a ciguatoxin congener isolated from cultured Gambierdiscus toxicus. Tetrahedron Lett 1993, 34, 1975–1978. [Google Scholar]
- Satake, M.; Ishibashi, Y.; Legrand, A.-M.; Yasumoto, T. Isolation and structure of ciguatoxin-4A, a new ciguatoxin precursor, from cultures of dinoflagellate Gambierdiscus toxicus and parrotfish Scarus gibbus. Biosci. Biotechnol. Biochem 1996, 60, 2103–2105. [Google Scholar]
- Lewis, R. J.; Norton, R. S.; Brereton, I. M.; Eccles, C. D. Ciguatoxin-2 is a diastereomer of ciguatoxin-3. Toxicon 1993, 31, 637–643. [Google Scholar]
- Satake, M.; Fukui, M.; Legrand, A.-M.; Cruchet, P.; Yasumoto, T. Isolation and structures of new ciguatoxin analogs, 2,3-dihydroxyCTX3C and 51-hydroxyCTX3C, accumulated in tropical reef fish. Tetrahedron Lett 1998, 39, 1197–1198. [Google Scholar]
- Holmes, M. J.; Lewis, R. J.; Poli, M. A.; Gillespie, N. C. Strain dependent production of ciguatoxin precursors (gambiertoxins) by Gambierdiscus toxicus (Dinophyceae) in culture. Toxicon 1991, 29, 761–775. [Google Scholar]
- Legrand, A.-M.; Fukui, M.; Cruchet, P.; Ishibashi, Y.; Yasumoto, T. Characterization of ciguatoxins from different fish species and wild Gambierdiscus toxicus. Proceedings of the Third International Conference on Ciguatera Fish Poisoning, Quebec; 1992; pp. 25–32. [Google Scholar]
- Lewis, R. J.; Sellin, M. Recovery of ciguatoxin from fish flesh. Toxicon 1993, 31, 1333–1336. [Google Scholar]
- Lewis, R. J.; Jones, A. Characterization of ciguatoxins and ciguatoxin congeners present in ciguateric fish by gradient reverse-phase high-performance liquid chromatography/mass spectrometry. Toxicon 1997, 35, 159–168. [Google Scholar]
- Legrand, A.-M.; Litaudon, M.; Genthon, J. N.; Bagnis, R.; Yasumoto, T. Isolation and some properties of ciguatoxin. J. Appl. Phycol 1989, 1, 183–188. [Google Scholar]
- Lewis, R. J.; Holmes, M. J. Origin and transfer of toxins involved in ciguatera. Comp. Biochem. Physiol. C 1993, 106, 615–628. [Google Scholar]
- Yasumoto, T.; Murata, M. Marine toxins. Chem. Rev 1993, 93, 1897–1909. [Google Scholar]
- Faust, M. Observation of sand-dwelling toxic dinoflagellates (Dinophyceae) from widely differing sites, including two new species. J. Phycol 1995, 31, 996–1003. [Google Scholar]
- Holmes, M. J. Gambierdiscus yasumotoi sp. nov. (Dinophyceae), a toxic dinoflagellate from southeastern Asia. J. Phycol 1998, 34, 661–668. [Google Scholar]
- Chinain, M.; Faust, M.; Pauillac, S. Morphology and molecular analyses of three new toxic species of Gambierdiscus (Dinophyceae): G. pacificus sp. nov., G. australes sp. nov., and G. polynesiensis sp. nov. J. Phycol 1999, 35, 1282–1296. [Google Scholar]
- Inoue, M.; Hirama, M.; Satake, M.; Sugiyama, K.; Yasumoto, T. Inhibition of brevetoxin binding to the voltage-gated sodium channel by gambierol and gambieric acid-A. Toxicon 2003, 41, 469–474. [Google Scholar]
- Satake, M.; Murata, M.; Yasumoto, T. Gambierol: a new toxic polyether compound isolated from the marine dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc 1993, 115, 361–362. [Google Scholar]
- Morohashi, A.; Satake, M.; Yasumoto, T. The absolute configuration of gambierol, a toxic marine polyether from the dinoflagellate, Gambierdiscus toxicus. Tetrahedron Lett 1998, 39, 97–100. [Google Scholar]
- Satake, M.; Ishibashi, Y.; Legrand, A.-M.; Yasumoto, T. Isolation and structure of ciguatoxin- 4A, a new ciguatoxin precursor, from cultures of dinoflagellate Gambierdiscus toxicus and parrotfish Scarus gibbus. Biosci Biotechnol Biochem 1996, 60, 2103–2105. [Google Scholar]
- Lin, Y.-Y.; Risk, M. A.; Ray, S. M.; Van Engen, D.; Clardy, J.; Golik, J.; James, J. C.; Nakanishi, K. Isolation and structure of brevetoxin B from the “red tide” dinoflagellate Ptychodiscus brevis (Gymnodinium breve). J. Am. Chem. Soc 1981, 103, 6773–3775. [Google Scholar]
- Chou, H. N.; Shimizu, Y. A new polyether toxin from Gymnodinium breve Davis. Tetrahedron Lett 1982, 23, 5521–5524. [Google Scholar]
- Dalton, C. B.; Gregory, J.; Kirk, M. D.; Stafford, R. J.; Givney, R.; Kraa, E.; Gould, D. Foodborne disease outbreaks in Australia, 1995 to 2000. Commun. Dis. Intell 2004, 28, 211–224. [Google Scholar]
- Fleming, L. E.; Katz, D.; Bean, J. A.; Hammond, R. Epidemiology of seafood poisoning. In Seafood and Environmental Toxins; Hui, Y. H., Kits, D. D., Eds.; Marcel Dekker: New York, 2001; pp. 287–310. [Google Scholar]
- Higerd, T. Ciguatera food poisoning: a circumtropical fisheries problem. In Natural Toxins and Human Pathogens in the Marine Environment; Colwell, R., Ed.; Maryland Sea Grant Publication, 1983; pp. 1–7. [Google Scholar]
- Lewis, R. J.; Ruff, T. A. Ciguatera: Ecological, clinical, and socioeconomic perspectives. Crit. Rev. Env. Sci. Technol 1993, 23, 137–156. [Google Scholar]
- Lange, W. R. Ciguatera fish poisoning. Am. Fam. Physician 1994, 50, 579–584. [Google Scholar]
- Lewis, R. J. Socioeconomic impacts and management ciguatera in the pacific. Bull. Soc. Path. Ex 1992, 85, 427–434. [Google Scholar]
- Morris, P. D.; Campbell, D. S.; Freeman, J. I. Ciguatera fish poisoning: an outbreak associated with fish caught from North Carolina coastal waters. South. Med. J 1990, 83, 379–382. [Google Scholar]
- DeFusco, D. J.; O'Dowd, P.; Hokama, Y.; Ott, B. R. Coma due to ciguatera poisoning in Rhode Island. Am. J. Med 1993, 95, 240–243. [Google Scholar]
- Halstead, B. W. Poisonous and Venomous Marine Animals of the World; United States Government Printing Office: Washington D.C, 1967. [Google Scholar]
- Banner, A. H. Ciguatera: a disease from coral reef fish. In Biology and Geology of Coral Reefs; Jones, O.A., Endean, R., Eds.; Academic Press: London, 1976; Volume 3, pp. 177–213. [Google Scholar]
- Gillespie, N. C.; Lewis, R. J.; Pearn, J. H.; Bourke, A. T.; Holmes, M. J.; Bourke, J. B.; Shields, W. J. Ciguatera in Australia. Occurrence, clinical features, pathophysiology and management. Med. J. Aust 1986, 145, 584–590. [Google Scholar]
- Glaziou, P.; Chinain, M.; Legrand, A.-M. Clinical toxicology of ciguatera poisoning. In Handbook of Clinical Toxicology of Animal Venoms and Poisons; Meier, J., White, J., Eds.; CRC Press: Boca Raton, 1995; pp. 59–74. [Google Scholar]
- Lewis, R. J.; Molgó, J.; Adams, D. J. Ciguatera toxins: Pharmacology of toxins involved in ciguatera and related fish poisonings. In Seafood and Freshwater Toxins. Pharmacology, Physiology and Detection; Botana, L. M., Ed.; Marcel Dekker, Inc: New York, 2000; pp. 419–447. [Google Scholar]
- Bagnis, R.; Kuberski, T.; Laugier, S. Clinical observations on 3,009 cases of ciguatera (fish poisoning) in the South Pacific. Am. J. Trop. Med. Hyg 1979, 28, 1067–1073. [Google Scholar]
- Lawrence, D. N.; Enriquez, M. B.; Lumish, R. M.; Maceo, A. Ciguatera fish poisoning in Miami. JAMA, J. Am. Med. Assoc 1980, 244, 254–258. [Google Scholar]
- Swift, A. E.; Swift, T. R. Ciguatera. J. Toxicol.-Clin. Toxicol 1993, 31, 1–29. [Google Scholar]
- Glaziou, P.; Legrand, A.-M. The epidemiology of ciguatera fish poisoning. Toxicon 1994, 32, 863–873. [Google Scholar]
- Quod, J. P.; Turquet, J. Ciguatera in Reunion Island (SW Indian Ocean): epidemiology and clinical patterns. Toxicon 1996, 34, 779–785. [Google Scholar]
- Pearn, J. H. Neurology of ciguatera. J. Neurol. Neurosurg. Psychiatry 2001, 70, 4–8. [Google Scholar]
- Arena, P.; Levin, B.; Fleming, L. E.; Friedman, M. A.; Blythe, D. G. A pilot study of the cognitive and psychological correlates of chronic ciguatera poisoning. Harmful Algae 2004, 3, 51–60. [Google Scholar]
- Lehane, L.; Lewis, R. J. Ciguatera: recent advances but the risk remains. Int. J. Food Microbiol 2000, 61, 91–125. [Google Scholar]
- Wong, C. K.; Hung, P.; Lee, K. L.; Kam, K. M. Study of an outbreak of ciguatera fish poisoning in Hong Kong. Toxicon 2005, 46, 563–571. [Google Scholar]
- Withers, N. W. Ciguatera fish toxins and poisoning. In Handbook of Natural Toxins and Venoms; Tu, A. T., Ed.; Marcel Dekker: New York, 1988; Volume 3, pp. 31–61. [Google Scholar]
- Backer, L. C.; Dickey, R. W.; Poli, M. A.; Davis, C.; Luber, G.; Friedman, M. A.; Fleming, L. E.; Patel, M.; Blythe, D. G. Development of a biological marker for exposure to ciguatera toxins. 11th International Conference on Harmful Algae Blooms, Cape Town, South Africa; 2004. [Google Scholar]
- Bagnis, R. A.; Legrand, A.-M. Clinical features on 12,890 cases of ciguatera (fish poisoning) in French Polynesia. In Progress in Venom and Toxin Research; Gopalakrishnakone, P., Tan, C. K., Eds.; Faculty of Science, National University of Singapore: Singapore, 1987; pp. 372–384. [Google Scholar]
- Narayan, Y. Fish poisoning in Fiji. Fiji Med. J 1980, 8, 567. [Google Scholar]
- Poon-King, C. M.; Chen, A.; Poon-King, T. Ciguatera fish poisoning in industrial ship crewmembers: a retrospective study in a seaport general practice in Trinidad and Tobago. West Indian Med. J 2004, 53, 220–226. [Google Scholar]
- Johnson, R.; Jong, E. C. Ciguatera: Caribbean and Indo-Pacific fish poisoning. West. J. Med 1983, 138, 872–874. [Google Scholar]
- Holt, R. J.; Miro, G.; Del Valle, A. An analysis of poison control center reports of ciguatera toxicity in Puerto Rico for one year. J. Toxicol.-Clin. Toxicol 1984, 22, 177–185. [Google Scholar]
- Dawson, J. Fish poisoning in American Samoa. Hawaii Med. J 1977, 36, 239–243. [Google Scholar]
- Schnorf, H.; Taurarii, M.; Cundy, T. Ciguatera fish poisoning: a double-blind randomized trial of mannitol therapy. Neurology 2002, 58, 873–880. [Google Scholar]
- Goonetilleke, A.; Harris, J. B. Envenomation and consumption of poisonous seafood. J. Neurol. Neurosurg. Psychiatry 2002, 73, 103–109. [Google Scholar]
- Bagnis, R. Clinical aspects of ciguatera (fish poisoning) in French Polynesia. Hawaii Med. J 1968, 28, 25–28. [Google Scholar]
- Cameron, J.; Capra, M. F. The basis of the paradoxical disturbance of temperature perception in ciguatera poisoning. J. Toxicol.-Clin. Toxicol 1993, 31, 571–579. [Google Scholar]
- Peng, Y. G.; Taylor, T. B.; Finch, R. E.; Moeller, P. D.; Ramsdell, J. S. Neuroexcitatory actions of ciguatoxin on brain regions associated with thermoregulation. Neuroreport 1995, 6, 305–309. [Google Scholar]
- Lewis, N. D. Ciguatera in the Pacific: incidence and implication for marine resources development. In Seafood Toxins; Ragelis, E. P., Ed.; American Chemical Society: Washington D.C, 1984; pp. 289–306. [Google Scholar]
- Lewis, N. D. Ciguatera, health and human adaptation in the Pacific. PhD Thesis, University of California, Berkeley, 1981. [Google Scholar]
- Pearce, R.; Hines, C., Jr; Burns, T. W.; Clark, C. E. Ciguatera (fish poisoning). Southern Medical Journal 1983, 76, 560–561. [Google Scholar]
- Miller, J. A.; Agnew, W. S.; Levinson, S. R. Principal glycopeptide of the tetrodotoxin/saxitoxin binding protein from Electrophorus electricus: isolation and partial chemical and physical characterization. Biochemistry 1983, 22, 462–470. [Google Scholar]
- Ayyar, D. R.; Mullaly, W. J. Ciguatera: clinical and electrophysiologic observations. Neurology 1977, 26, 354. [Google Scholar]
- Cameron, J.; Flowers, A. E.; Capra, M. F. Effects of ciguatoxin on nerve excitability in rats (Part I). J. Neurol. Sci 1991, 101, 87–92. [Google Scholar]
- Cameron, J.; Flowers, A. E.; Capra, M. F. Electrophysiological studies on ciguatera poisoning in man (Part II). J. Neurol. Sci 1991, 101, 93–97. [Google Scholar]
- Parkin, P. J.; Le Quesne, P. M. Effect of a synthetic pyrethroid deltamethrin on excitability changes following a nerve impulse. J. Neurol. Neurosurg. Psychiatry 1982, 45, 337–342. [Google Scholar]
- Withers, N. W. Ciguatera fish poisoning. Annu. Rev. Med 1982, 33, 97–111. [Google Scholar]
- Chan, T. Y.; Wang, A. Y. Life-threatening bradycardia and hypotension in a patient with ciguatera fish poisoning. Trans. R. Soc. Trop. Med. Hyg 1993, 87, 71. [Google Scholar]
- Gillespie, N. C. Ciguatera poisoning. In Toxic Plants and Animals: A Guide for Australia; Covacevich, J., Davie, P., Pearn, J., Eds.; Queensland Museum; Brisbane, 1987; pp. 160–169. [Google Scholar]
- Lewis, R. J. Ciguatera in south-eastern Queensland. In Toxic Plants and Animals: a Guide for Australia; Covacevich, J., Davie, P., Pearn, J., Eds.; Queensland Museum; Brisbane, 1987; pp. 181–187. [Google Scholar]
- Russell, F. E.; Egen, N. B. Ciguateric fishes, ciguatoxin (CTX) and ciguatera poisoning. J. Toxicol 1991, 10, 36–62. [Google Scholar]
- Barton, E. D.; Tanner, P.; Turchen, S. G.; Tunget, C. L.; Manoguerra, A.; Clark, R. F. Ciguatera fish poisoning. A southern California epidemic. West. J. Med 1995, 163, 31–35. [Google Scholar]
- Bagnis, R.; Chanteau, S.; Chungue, E.; Hurtel, J. M.; Yasumoto, T.; Inoue, A. Origins of ciguatera fish poisoning: a new dinoflagellate, Gambierdiscus toxicus Adachi and Fukuyo, definitively involved as a causal agent. Toxicon 1980, 18, 199–208. [Google Scholar]
- Helfrich, P. Fish poisoning in Hawaii. Hawaii Med. J 1963, 22, 361–372. [Google Scholar]
- Sawyer, P.; Jollow, D.; Scheuer, P. J.; York, R.; McMillan, J.; Withers, N. W.; Fundenburg, H.; Higerd, T. The effect of ciguatera associated toxins in mice. Am. Chem. Soc. Symp. on Seafood Toxins 1984, 262, 322–329. [Google Scholar]
- Kew, C. K.; Ming, L. K.; Hang, W. M. The mechanism of respiratory failure in ciguatera poisoning. J. Pathol 1969, 97, 89–92. [Google Scholar]
- Legrand, A.-M.; Lotte, C.; Bagnis, R. Respiratory and cardiovascular effects of ciguatoxin in cats; antagonistic action of hexamethonium, atropine, propranolol, phentolamine, yohimbine, prazosin, verapamil, calcium and lidocaine. Proceedings of the 5th International Coral Reef Congress, Tahiti; 1985; pp. 463–466. [Google Scholar]
- Legrand, A.-M.; Galonnier, M.; Bagnis, R. Studies on the mode of action of ciguateric toxins. Toxicon 1982, 20, 311–315. [Google Scholar]
- Terao, K.; Ito, E.; Oarada, M.; Ishibashi, Y.; Legrand, A.-M.; Yasumoto, T. Light and electron microscopic studies of pathologic changes induced in mice by ciguatoxin poisoning. Toxicon 1991, 29, 633–643. [Google Scholar]
- Terao, K.; Ito, E.; Yasumoto, T. Light and electron microscopic studies of the murine heart after repeated administrations of ciguatoxin or ciguatoxin-4c. Nat. Toxins 1992, 1, 19–26. [Google Scholar]
- Legrand, A.-M.; Bagnis, R. Effects of ciguatoxin and maitotoxin on isolated rat atria and rabbit duodenum. Toxicon 1984, 22, 471–475. [Google Scholar]
- Lewis, R. J.; Endean, R. Direct and indirect effects of ciguatoxin on guinea-pig atria and papillary muscles. Naunyn-Schmiedeberg's Arch. Pharmacol 1986, 334, 313–322. [Google Scholar]
- Lewis, R. J.; Hoy, A. W.; McGiffin, D. C. Action of ciguatoxin on human atrial trabeculae. Toxicon 1992, 30, 907–914. [Google Scholar]
- Seino, A.; Kobayashi, M.; Momose, K.; Yasumoto, T.; Ohizumi, Y. The mode of inotropic action of ciguatoxin on guinea-pig cardiac muscle. Brit. J. Pharmacol 1988, 95, 876–882. [Google Scholar]
- Lewis, R. J. Negative inotropic and arrhythmic effects of high doses of ciguatoxin on guinea-pig atria and papillary muscles. Toxicon 1988, 26, 639–649. [Google Scholar]
- Marquais, M.; Vernoux, J. P.; Molgó, J.; Sauviat, M. P.; Lewis, R. J. Isolation and electrophysiological characterisation of a new ciguatoxin extracted from Caribbean fish. In Harmful Microalgae; Reguera, B., Blanco, J., Fernández, M. L., Wyatt, R. J., Eds.; Santiago De Compostella: Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO: Santiago, 1998; pp. 476–477. [Google Scholar]
- Sauviat, M. P.; Marquais, M.; Vernoux, J. P. Muscarinic effects of the Caribbean ciguatoxin C-CTX-1 on frog atrial heart muscle. Toxicon 2002, 40, 1155–1163. [Google Scholar]
- Lewis, R. J.; Endean, R. Mode of action of ciguatoxin from the Spanish Mackerel, Scomberomorus commersoni, on the guinea-pig ileum and vas deferens. J. Pharmacol. Exp. Ther 1984, 228, 756–760. [Google Scholar]
- Lewis, R. J.; Hoy, A. W. Comparative action of three major ciguatoxins on guinea-pig atria and ilea. Toxicon 1993, 31, 437–446. [Google Scholar]
- Ohizumi, Y.; Shibata, S.; Tachibana, K. Mode of the excitatory and inhibitory actions of ciguatoxin in the guinea-pig vas deferens. J. Pharmacol. Exp. Ther 1981, 217, 475–480. [Google Scholar]
- Ohizumi, Y.; Ishida, Y.; Shibata, S. Mode of the ciguatoxin-induced supersensitivity in the guinea-pig vas deferens. J. Pharmacol. Exp. Ther 1982, 221, 748–752. [Google Scholar]
- Brock, J. A.; McLachlan, E. M.; Jobling, P.; Lewis, R. J. Electrical activity in rat tail artery during asynchronous activation of postganglionic nerve terminals by ciguatoxin-1. Brit. J. Pharmacol 1995, 116, 2213–2220. [Google Scholar]
- Bidard, J. N.; Vijverberg, H. P.; Frelin, C.; Chungue, E.; Legrand, A.-M.; Bagnis, R.; Lazdunski, M. Ciguatoxin is a novel type of Na+ channel toxin. J. Biol. Chem 1984, 259, 8353–8357. [Google Scholar]
- Molgó, J.; Comella, J. X.; Legrand, A.-M. Ciguatoxin enhances quantal transmitter release from frog motor nerve terminals. Brit. J. Pharmacol 1990, 99, 695–700. [Google Scholar]
- Molgó, J.; Gaudry-Talarmain, Y. M.; Legrand, A.-M.; Moulian, N. Ciguatoxin extracted from poisonous moray eels Gymnothorax javanicus triggers acetylcholine release from Torpedo cholinergic synaptosomes via reversed Na+-Ca2+ exchange. Neurosci Lett 1993, 160, 65–68. [Google Scholar]
- Gaudry-Talarmain, Y. M.; Molgó, J.; Meunier, F. A.; Moulian, N.; Legrand, A.-M. Reversed mode Na+-Ca2+ exchange activated by ciguatoxin (CTX-1b) enhances acetylcholine release from Torpedo cholinergic synaptosomes. Ann. N. Y. Acad. Sci 1996, 779, 404–406. [Google Scholar]
- Molgó, J.; Comella, J. X.; Shimahara, T.; Legrand, A.-M. Tetrodotoxin-sensitive ciguatoxin effects on quantal release, synaptic vesicle depletion, and calcium mobilization. Ann. N. Y. Acad. Sci 1991, 635, 485–488. [Google Scholar]
- Hamblin, P. A.; McLachlan, E. M.; Lewis, R. J. Sub-nanomolar concentrations of ciguatoxin-1 excite preganglionic terminals in guinea pig sympathetic ganglia. Naunyn-Schmiedeberg's Arch. Pharmacol 1995, 352, 236–246. [Google Scholar]
- Benoit, E.; Legrand, A.-M.; Dubois, J. M. Effects of ciguatoxin on current and voltage clamped frog myelinated nerve fibre. Toxicon 1986, 24, 357–364. [Google Scholar]
- Birinyi-Strachan, L. C.; Gunning, S. J.; Lewis, R. J.; Nicholson, G. M. Block of voltage-gated potassium channels by Pacific ciguatoxin-1 contributes to increased neuronal excitability in rat sensory neurons. Toxicol. Appl. Pharmacol 2005, 204, 175–186. [Google Scholar]
- Benoit, E.; Legrand, A.-M. Gambiertoxin-induced modifications of the membrane potential of myelinated nerve fibres. Mem. Queensland Mus 1994, 34, 461–464. [Google Scholar]
- Shapiro, L.; Doyle, J. P.; Hensley, P.; Colman, D. R.; Hendrickson, W. A. Crystal structure of the extracellular domain from P0, the major structural protein of peripheral nerve myelin. Neuron 1996, 17, 435–449. [Google Scholar]
- Cestèle, S.; Catterall, W. A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar]
- Nicholson, G. M.; Little, M. J. Spider neurotoxins targeting voltage-gated sodium channels. Toxin Rev 2005, 25, 315–345. [Google Scholar]
- Strachan, L. C.; Lewis, R. J.; Nicholson, G. M. Differential actions of Pacific ciguatoxin-1 on sodium channel subtypes in mammalian sensory neurons. J. Pharmacol. Exp. Ther 1999, 288, 379–388. [Google Scholar]
- Hogg, R. C.; Lewis, R. J.; Adams, D. J. Ciguatoxin-induced oscillations in membrane potential and action potential firing in rat parasympathetic neurons. Eur. J. Neurosci 2002, 16, 242–248. [Google Scholar]
- Hogg, R. C.; Lewis, R. J.; Adams, D. J. Ciguatoxin (CTX-1) modulates single tetrodotoxin-sensitive sodium channels in rat parasympathetic neurones. Neurosci. Lett 1998, 252, 103–106. [Google Scholar]
- Sangameswaran, L.; Delgado, S. G.; Fish, L. M.; Koch, B. D.; Jakeman, L. B.; Stewart, G. R.; Sze, P.; Hunter, J. C.; Eglen, R. M.; Herman, R. C. Structure and function of a novel voltage-gated, tetrodotoxin-resistant sodium channel specific to sensory neurons. J. Biol. Chem 1996, 271, 5953–5956. [Google Scholar]
- Dib-Hajj, S. D.; Tyrrell, L.; Black, J. A.; Waxman, S. G. NaN, a novel voltage-gated Na channel, is expressed preferentially in peripheral sensory neurons and down-regulated after axotomy. Proc. Natl. Acad. Sci. USA 1998, 95, 8963–8968. [Google Scholar]
- Beckh, S. Differential expression of sodium channel mRNAs in rat peripheral nervous system and innervated tissues. FEBS Lett 1990, 262, 317–322. [Google Scholar]
- Caldwell, J. H.; Schaller, K. L.; Lasher, R. S.; Peles, E.; Levinson, S. R. Sodium channel Nav1.6 is localized at nodes of Ranvier, dendrites, and synapses. Proc. Natl. Acad. Sci. USA 2000, 97, 5616–5620. [Google Scholar]
- Toledo-Aral, J. J.; Moss, B. L.; He, Z. J.; Koszowski, A. G.; Whisenand, T.; Levinson, S. R.; Wolf, J. J.; Silos-Santiago, I.; Halegoua, S.; Mandel, G. Identification of PN1, a predominant voltage-dependent sodium channel expressed principally in peripheral neurons. Proc. Natl. Acad. Sci. USA 1997, 94, 1527–1532. [Google Scholar]
- Felts, P. A.; Yokoyama, S.; Dib-Hajj, S.; Black, J. A.; Waxman, S. G. Sodium channel α-subunit mRNAs I, II, III, NaG, Na6 and hNE (PN1): different expression patterns in developing rat nervous system. Mol. Brain Res 1997, 45, 71–82. [Google Scholar]
- Yamaoka, K.; Inoue, M.; Miyahara, H.; Miyazaki, K.; Hirama, M. A quantitative and comparative study of the effects of a synthetic ciguatoxin CTX3C on the kinetic properties of voltage-dependent sodium channels. Brit. J. Pharmacol 2004, 142, 879–889. [Google Scholar]
- Benoit, E.; Legrand, A.-M. Purified ciguatoxin-induced modifications in excitability of myelinated nerve fibre. Bull. Soc. Pathol. Exot 1992, 85, 497–499. [Google Scholar]
- Benoit, E.; Juzans, P.; Legrand, A.-M.; Molgó, J. Nodal swelling produced by ciguatoxin-induced selective activation of sodium channels in myelinated nerve fibers. Neuroscience 1996, 71, 1121–1131. [Google Scholar]
- Birinyi-Strachan, L. C.; Davies, M. J.; Lewis, R. J.; Nicholson, G. M. Neuroprotectant effects of iso-osmolar D-mannitol to prevent Pacific ciguatoxin-1 induced alterations in neuronal excitability: a comparison with other osmotic agents and free radical scavengers. Neuropharmacology 2005, 49, 669–686. [Google Scholar]
- Rizzo, M. A.; Kocsis, J. D.; Waxman, S. G. Mechanisms of paresthesiae, dysesthesiae, and hyperesthesiae: role of Na+ channel heterogeneity. Eur. Neurol 1996, 36, 3–12. [Google Scholar]
- Wall, P. D.; Devor, M. Sensory afferent impulses originate from dorsal root ganglia as well as from the periphery in normal and nerve injured rats. Pain 1983, 17, 321–339. [Google Scholar]
- Gordon, D. Sodium channels as targets for neurotoxins: mode of action and interaction of neurotoxins with receptor sites on sodium channels. In Toxins and Signal Transduction; Lazarowici, P., Gutman, Y., Eds.; Harwood Press: Amsterdam, 1997; Vol Cellular And Molecular Mechanisms of Toxin Action Series; pp. 119–149. [Google Scholar]
- Gordon, D. A new approach to insect-pest control-combination of neurotoxins interacting with voltage sensitive sodium channels to increase selectivity and specificity. Invert. Neurosci 1997, 3, 103–116. [Google Scholar]
- Tan, J.; Liu, Z.; Wang, R.; Huang, Z. Y.; Chen, A. C.; Gurevitz, M.; Dong, K. Identification of amino acid residues in the insect sodium channel critical for pyrethroid binding. Mol. Pharmacol 2005, 67, 513–522. [Google Scholar]
- Lombet, A.; Bidard, J.-N.; Lazdunski, M. Ciguatoxin and brevetoxins share a common receptor site on the neuronal voltage-dependent Na+ channel. FEBS Lett 1987, 219, 355–359. [Google Scholar]
- Pauillac, S.; Bléhaut, J.; Cruchet, P.; Lotte, C.; Legrand, A.-M. Recent advances in detection of ciguatoxins in French Polynesia. In Harmful Algal Blooms; Lassus, P., Arzul, G., Erard, E., Gentien, P., Marcaillou, C., Eds.; Lavoisier, Intercept Ltd: France, 1995; pp. 801–808. [Google Scholar]
- Dechraoui, M. Y.; Naar, J.; Pauillac, S.; Legrand, A.-M. Ciguatoxins and brevetoxins, neurotoxic polyether compounds active on sodium channels. Toxicon 1999, 37, 125–143. [Google Scholar]
- Poli, M. A.; Lewis, R. J.; Dickey, R. W.; Musser, S. M.; Buckner, C. A.; Carpenter, L. G. Identification of Caribbean ciguatoxins as the cause of an outbreak of fish poisoning among U.S. soldiers in Haiti. Toxicon 1997, 35, 733–741. [Google Scholar]
- Bottein Dechraoui, M. Y.; Ramsdell, J. S. Type B brevetoxins show tissue selectivity for voltage-gated sodium channels: comparison of brain, skeletal muscle and cardiac sodium channels. Toxicon 2003, 41, 919–927. [Google Scholar]
- Poli, M. A.; Mende, T. J.; Baden, D. G. Brevetoxins, unique activators of voltage-sensitive sodium channels, bind to specific sites in rat brain synaptosomes. Mol. Pharmacol 1986, 30, 129–135. [Google Scholar]
- Sharkey, R. G.; Jover, E.; Couraud, F.; Baden, D. G.; Catterall, W. A. Allosteric modulation of neurotoxin binding to voltage-sensitive sodium channels by Ptychodiscus brevis toxin 2. Mol. Pharmacol 1987, 31, 273–278. [Google Scholar]
- Cestèle, S.; Sampieri, F.; Rochat, H.; Gordon, D. Tetrodotoxin reverses brevetoxin allosteric inhibition of scorpion α-toxin binding on rat brain sodium channels. J. Biol. Chem 1996, 271, 18329–18332. [Google Scholar]
- Trainer, V. L.; Baden, D. G.; Catterall, W. A. Identification of peptide components of the brevetoxin receptor site of rat brain sodium channels. J. Biol. Chem 1994, 269, 19904–19909. [Google Scholar]
- Trainer, V. L.; Thomsen, W. J.; Catterall, W. A.; Baden, D. G. Photoaffinity labelling of the brevetoxin receptor on sodium channels in rat brain synaptosomes. Mol. Pharmacol. 1991, 40, 988–994. [Google Scholar]
- Hidalgo, J.; Liberona, J. L.; Molgó, J.; Jaimovich, E. Pacific ciguatoxin-1b effect over Na+ and K+ currents, inositol 1,4,5-triphosphate content and intracellular Ca2+ signals in cultured rat myotubes. Brit. J. Pharmacol 2002, 137, 1055–1062. [Google Scholar]
- Safronov, B. V.; Bischoff, U.; Vogel, W. Single voltage-gated K+ channels and their functions in small dorsal root ganglion neurones of rat. J. Physiol 1996, 493, 393–408. [Google Scholar]
- Hille, B. Ion channels of excitable membranes; Sinauer Associates, Inc: Sunderland; p. 2001.
- Baker, M.; Bostock, H.; Grafe, P. Accommodation in rat myelinated axons depends on two pharmacologically distinct types of potassium channels. J. Physiol 1985, 369, 102P. [Google Scholar]
- Amir, R.; Liu, C. N.; Kocsis, J. D.; Devor, M. Oscillatory mechanism in primary sensory neurones. Brain 2002, 125, 421–435. [Google Scholar]
- Ghiaroni, V.; Sasaki, M.; Fuwa, H.; Rossini, G. P.; Scalera, G.; Yasumoto, T.; Pietra, P.; Bigiani, A. Inhibition of voltage-gated potassium currents by gambierol in mouse taste cells. Toxicol. Sci 2005, 85, 657–665. [Google Scholar]
- Molgó, J.; Meunier, F. A.; Dechraoui, M. Y.; Benoit, E.; Mattei, C.; Legrand, A.-M. Sodium-dependent alterations of synaptic transmission mechanisms by brevetoxins and ciguatoxins. In Harmful Microalgae; Reguera, B., Blanco, J., Fernández, M. L., Wyatt, T., Eds.; Santiago De Compostella: Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO: Santiago, 1998; pp. 594–497. [Google Scholar]
- Molgó, J.; Shimahara, T.; Legrand, A.-M. Ciguatoxin, extracted from poisonous moray eels, causes sodium-dependent calcium mobilization in NG108-15 neuroblastoma x glioma hybrid cells. Neurosci. Lett 1993, 158, 147–150. [Google Scholar]
- Mattei, C.; Dechraoui, M. Y.; Molgó, J.; Meunier, F. A.; Legrand, A.-M.; Benoit, E. Neurotoxins targetting receptor site 5 of voltage-dependent sodium channels increase the nodal volume of myelinated axons. J. Neurosci. Res 1999, 55, 666–673. [Google Scholar]
- Allsop, J. L.; Martini, L.; Lebris, H.; Pollard, J.; Walsh, J.; Hodgkinson, S. Les manifestations neurologiques de la ciguatera. Rev. Neurol. (Paris) 1986, 142, 590–597. [Google Scholar]
- Mattei, C.; Benoit, E.; Juzans, P.; Legrand, A.-M.; Molgó, J. Gambiertoxin (CTX-4B), purified from wild Gambierdiscus toxicus dinoflagellates, induces Na+-dependent swelling of single frog myelinated axons and motor nerve terminals in situ. Neurosci. Lett 1997, 234, 75–78. [Google Scholar]
- Mattei, C.; Molgó, J.; Marquais, M.; Vernoux, J.; Benoit, E. Hyperosmolar d-mannitol reverses the increased membrane excitability and the nodal swelling caused by Caribbean ciguatoxin-1 in single frog myelinated axons. Brain Res 1999, 847, 50–58. [Google Scholar]
- Molgó, J.; Juzans, P.; Legrand, A.-M. Confocal laser scanning microscopy: a new tool for studying the effects of ciguatoxin (CTX-1b) and mannitol at motor nerve terminals of the neuromuscular junction in situ. Mem. Queensland Mus 1994, 34, 577–585. [Google Scholar]
- Benoit, E.; Mattei, C.; Ouanounou, G.; Meunier, F. A.; Suput, D.; Le Gall, F.; Marquais, M.; Dechraoui, M. Y.; Molgó, J. Ionic mechanisms involved in the nodal swelling of myelinated axons caused by marine toxins. Cell. Mol. Biol. Lett 2002, 7, 317–321. [Google Scholar]
- Kostyuk, P.; Verkhratsky, A. Calcium stores in neurons and glia. Neuroscience 1994, 63, 381–404. [Google Scholar]
- Molgó, J.; Shimahara, T.; Gaudry-Talarmain, Y. M.; Comella, J. X.; Legrand, A.-M. Ciguatoxin-induced changes in acetylcholine release and in cytosolic calcium levels. Bull. Soc. Pathol. Exot 1992, 85, 486–488. [Google Scholar]
- Yano, K.; Higashida, H.; Inoue, R.; Nozawa, Y. Bradykinin-induced rapid breakdown of phosphatidylinositol 4,5-bisphosphate in neuroblastoma X glioma hybrid NG108-15 cells. J. Biol. Chem 1984, 259, 10201–10207. [Google Scholar]
- Henzi, V.; MacDermott, A. B. Characteristics and function of Ca2+- and inositol 1,4,5-trisphosphate-releasable stores of Ca2+ in neurons. Neuroscience 1992, 46, 251–273. [Google Scholar]
- Gusovsky, F.; Daly, J. W.; Yasumoto, T.; Rojas, E. Differential effects of maitotoxin on ATP secretion and on phosphoinositide breakdown in rat pheochromocytoma cells. FEBS Lett 1988, 233, 139–142. [Google Scholar]
- Gusovsky, F.; Hollingsworth, E. B.; Daly, J. W. Regulation of phosphatidylinositol turnover in brain synaptoneurosomes: stimulatory effects of agents that enhance influx of sodium ions. Proc. Natl. Acad. Sci. USA 1986, 83, 3003–3007. [Google Scholar]
- Carrasco, M. A.; Gaudry-Talarmain, Y. M.; Molgó, J. Ca2+-dependent changes of acetylcholine release and IP3 mass in Torpedo cholinergic synaptosomes. Neurochem. Int 1996, 29, 637–643. [Google Scholar]
- McDonough, P. M.; Goldstein, D.; Brown, J. H. Elevation of cytoplasmic calcium concentration stimulates hydrolysis of phosphatidylinositol bisphosphate in chick heart cells: effect of sodium channel activators. Mol. Pharmacol 1988, 33, 310–315. [Google Scholar]
- Hermoni, M.; Barzilai, A.; Rahamimoff, H. Modulation of the Na+-Ca2+ antiport by its ionic environment: the effect of lithium. Isr. J. Med. Sci 1987, 23, 44–48. [Google Scholar]
- Banner, A. H.; Sasaki, S.; Helfrich, P.; Alender, C. B.; Scheuer, P. J. Bioassay of ciguatera toxin. Nature 1961, 189, 229–230. [Google Scholar]
- Hoffman, P. A.; Granade, H. R.; McMillan, J. P. The mouse ciguatoxin bioassay: a dose-response curve and symptomatology analysis. Toxicon 1983, 21, 363–369. [Google Scholar]
- Labrousse, H.; Matile, L. Toxicological biotest on Diptera larvae to detect ciguatoxins and various other toxic substances. Toxicon 1996, 34, 881–891. [Google Scholar]
- Lewis, R. J. Detection of ciguatoxins and related benthic dinoflagellate toxins: in vivoand in vitromethods. In Manual on Harmful Marine Microalgae; Hallegraph, G. M., Anderson, D. M., Cembella, A. D., Eds.; IOC Manuals and Guides UNESCO: France, 1995; Volume 33, pp. 135–161. [Google Scholar]
- Vernoux, J. P.; Lahlou, N.; Magras, L. P.; Greaux, J. B. Chick feeding test: a simple system to detect ciguatoxin. Acta Trop 1985, 42, 235–240. [Google Scholar]
- Chungue, E.; Bagnis, R.; Parc, F. The use of mosquitoes (Aedes aegypti) to detect ciguatoxin in surgeon fishes (Ctenochaetus striatus). Toxicon 1984, 22, 161–164. [Google Scholar]
- Lewis, R. J.; Hoy, A. W.; Sellin, M. Ciguatera and mannitol: in vivo and in vitro assessment in mice. Toxicon 1993, 31, 1039–1050. [Google Scholar]
- Park, D. L. Evolution of methods for assessing ciguatera toxins in fish. Rev. Environ. Contam. Toxicol 1994, 136, 1–20. [Google Scholar]
- Manger, R. L.; Leja, L. S.; Lee, S. Y.; Hungerford, J. M.; Hokama, Y.; Dickey, R. W.; Granade, H. R.; Lewis, R.; Yasumoto, T.; Wekell, M. M. Detection of sodium channel toxins: directed cytotoxicity assays of purified ciguatoxins, brevetoxins, saxitoxins, and seafood extracts. J. AOAC Int 1995, 78, 521–527. [Google Scholar]
- Manger, R. L.; Leja, L. S.; Lee, S. Y.; Hungerford, J. M.; Wekell, M. M. Tetrazolium-based cell bioassay for neurotoxins active on voltage-sensitive sodium channels: semiautomated assay for saxitoxins, brevetoxins, and ciguatoxins. Anal. Biochem 1993, 214, 190–194. [Google Scholar]
- Louzao, M. C.; Vieytes, M. R.; Yasumoto, T.; Botana, L. M. Detection of sodium channel activators by a rapid fluorimetric microplate assay. Chem. Res. Toxicol 2004, 17, 572–578. [Google Scholar]
- Fairey, E. R.; Edmunds, J. S.; Ramsdell, J. S. A cell-based assay for brevetoxins, saxitoxins, and ciguatoxins using a stably expressed c-fos-luciferase reporter gene. Anal. Biochem 1997, 251, 129–132. [Google Scholar]
- Lewis, R. J.; Jones, A.; Vernoux, J. P. HPLC/tandem electrospray mass spectrometry for the determination of Sub-ppb levels of Pacific and Caribbean ciguatoxins in crude extracts of fish. Anal. Chem 1999, 71, 247–250. [Google Scholar]
- Oguri, H.; Hirama, M.; Tsumuraya, T.; Fujii, I.; Maruyama, M.; Uehara, H.; Nagumo, Y. Synthesis-based approach toward direct sandwich immunoassay for ciguatoxin CTX3C. J. Am. Chem. Soc 2003, 125, 7608–7612. [Google Scholar]
- Kimura, L. H.; Hokama, Y.; Abad, M. A.; Oyama, M.; Miyahara, J. T. Comparison of three different assays for the assessment of ciguatoxin in fish tissues: radioimmunoassay, mouse bioassay and in vitro guinea pig atrium assay. Toxicon 1982, 20, 907–912. [Google Scholar]
- Ganal, C. A.; Asahina, A. Y.; Hokama, Y.; Miyahara, J. T. Characterization of marine toxin(s) in Myripristis sp. by immunological, mouse toxicity, and guinea pig assays. J. Clin. Lab. Anal 1993, 7, 41–45. [Google Scholar]
- Hokama, Y.; Banner, A. H.; Boylan, D. B. A radioimmunoassay for the detection of ciguatoxin. Toxicon 1977, 15, 317–325. [Google Scholar]
- Hokama, Y.; Abad, M. A.; Kimura, L. H. A rapid enzyme-immunoassay for the detection of ciguatoxin in contaminated fish tissues. Toxicon 1983, 21, 817–824. [Google Scholar]
- Hokama, Y. A rapid, simplified enzyme immunoassay stick test for the detection of ciguatoxin and related polyethers from fish tissues. Toxicon 1985, 23, 939–946. [Google Scholar]
- Hokama, Y.; Asahina, A. Y.; Hong, T. W.; Shang, E. S.; Miyahara, J. T. Evaluation of the stick enzyme immunoassay in Caranx sp. and Seriola dumerili associated with ciguatera. J. Clin. Lab. Anal 1990, 4, 363–366. [Google Scholar]
- Hokama, Y. Simplified solid-phase immunobead assay for detection of ciguatoxin and related polyethers. J. Clin. Lab. Anal 1990, 4, 213–217. [Google Scholar]
- Hokama, Y.; Asahina, A. Y.; Shang, E. S.; Hong, T. W.; Shirai, J. L. Evaluation of the Hawaiian reef fishes with the solid phase immunobead assay. J. Clin. Lab. Anal 1993, 7, 26–30. [Google Scholar]
- Hokama, Y. Immunological studies using monoclonal antibodies for detection of low dalton marine toxins. Food Addit. Contam 1993, 10, 83–95. [Google Scholar]
- Hokama, Y. Recent methods for detection of seafood toxins: recent immunological methods for ciguatoxin and related polyethers. Food Addit. Contam 1993, 10, 71–82. [Google Scholar]
- Bottein Dechraoui, M. Y.; Wang, Z.; Turquet, J.; Chinain, M.; Darius, T.; Cruchet, P.; Radwan, F. F.; Dickey, R. W.; Ramsdell, J. S. Biomonitoring of ciguatoxin exposure in mice using blood collection cards. Toxicon 2005, 46, 243–251. [Google Scholar]
- Matta, J.; Navas, J.; Milad, M.; Manger, R.; Hupka, A.; Frazer, T. A pilot study for the detection of acute ciguatera intoxication in human blood. J. Toxicol.-Clin. Toxicol 2002, 40, 49–57. [Google Scholar]
- Hokama, Y.; Takenaka, W. E.; Nishimura, K. L.; Ebesu, J. S.; Bourke, R.; Sullivan, P. K. A simple membrane immunobead assay for detecting ciguatoxin and related polyethers from human ciguatera intoxication and natural reef fishes. J. AOAC Int 1998, 81, 727–735. [Google Scholar]
- Hokama, Y.; Nishimura, K.; Takenaka, W.; Ebesu, J. S. Simplified solid-phase membrane immunobead assay (MIA) with monoclonal anti-ciguatoxin antibody (MAb-CTX) for detection of ciguatoxin and related polyether toxins. J. Nat. Toxins 1998, 7, 1–21. [Google Scholar]
- Halstead, B. W. Poisonous and Venomous Marine Animals of the World; Darwin Press: Princeton, NJ, 1978. [Google Scholar]
- Davis, R. T.; Villar, L. A. Symptomatic improvement with amitriptyline in ciguatera fish poisoning. N. Engl. J. Med 1986, 315, 65. [Google Scholar]
- Calvert, G. M.; Hryhorczuk, D. O.; Leikin, J. B. Treatment of ciguatera fish poisoning with amitriptyline and nifedipine. J. Toxicol.-Clin. Toxicol 1987, 25, 423–428. [Google Scholar]
- Lange, W. R.; Snyder, F. R.; Fudala, P. J. Travel and ciguatera fish poisoning. Arch. Intern. Med 1992, 152, 2049–2053. [Google Scholar]
- Berlin, R. M.; King, S. L.; Blythe, D. G. Symptomatic improvement of chronic fatigue with fluoxetine in ciguatera fish poisoning. Med. J. Aust 1992, 157, 567. [Google Scholar]
- Nicholson, G. M.; Blanche, T.; Mansfield, K.; Tran, Y. Differential blockade of neuronal voltage-gated Na+ and K+ channels by antidepressant drugs. Eur. J. Pharmacol 2002, 452, 35–48. [Google Scholar]
- Perez, C. M.; Vasquez, P. A.; Perret, C. F. Treatment of ciguatera poisoning with gabapentin. N. Engl. J. Med 2001, 344, 692–693. [Google Scholar]
- Lange, W. R.; Kreider, S. D. A pilot study of the potential benefit of tocainide in the management of ciguatera toxicity. 1st Conference on International Travel Medicine, Zurich; 1988. [Google Scholar]
- Lange, W. R.; Kreider, S. D.; Hattwick, M.; Hobbs, J. Potential benefit of tocainide in the treatment of ciguatera: report of three cases. Am. J. Med 1988, 84, 1087–1088. [Google Scholar]
- Cameron, J.; Flowers, A. E.; Capra, M. F. Modification of the peripheral nerve disturbance in ciguatera poisoning in rats with lidocaine. Muscle & Nerve 1993, 16, 782–786. [Google Scholar]
- Hoy, A. W.; Lewis, R. J. The effect of potential therapeutics on ciguatoxin-inhibition of the rat phrenic nerve. In Recent Advances in Toxinology Research; Gopalakrishnakone, P., Tan, C. K., Eds.; Faculty of Science, National University of Singapore: Singapore, 1992; Volume 2, pp. 509–519. [Google Scholar]
- Russell, F. E. Ciguatera poisoning: a report of 35 cases. Toxicon 1975, 13, 383–385. [Google Scholar]
- Bourdy, G.; Cabalion, P.; Amade, P.; Laurent, D. Traditional remedies used in the western Pacific for the treatment of ciguatera poisoning. J. Ethnopharmacol 1992, 36, 163–174. [Google Scholar]
- Amade, P.; Laurent, D. Screening of traditional remedies used in ciguatera fish poisoning treatment. Rec. Adv. Toxinol. Res 1992, 2, 503–508. [Google Scholar]
- Benoit, E.; Laurent, D.; Mattei, C.; Legrand, A.-M.; Molgó, J. Reversal of Pacific-ciguatoxin-1B effects on myelinated axons by agents used in ciguatera treatment. Cymbium 2000, 24 Suppl, 33–40. [Google Scholar]
- Garrec, R. B.; Benoit, E.; Sauviat, M. P.; Lewis, R. J.; Molgó, J.; Laurent, D. Ability of some plant extracts, traditionally used to treat ciguatera fish poisoning, to prevent the in vitro neurotoxicity produced by sodium channel activators. Toxicon 2005, 46, 625–634. [Google Scholar]
- Palafox, N. A.; Jain, L. G.; Pinano, A. Z.; Gulick, T. M.; Williams, R. K.; Schatz, I. J. Successful treatment of ciguatera fish poisoning with intravenous mannitol. JAMA 1988, 259, 2740–2742. [Google Scholar]
- Pearn, J. H.; Lewis, R. J.; Ruff, T.; Tait, M.; Quinn, J.; Murtha, W.; King, G.; Mallett, A.; Gillespie, N. C. Ciguatera and mannitol: experience with a new treatment regimen. Med. J. Aust 1989, 151, 77–80. [Google Scholar]
- Blythe, D. G.; De Sylva, D. P.; Fleming, L. E.; Ayyar, R. A.; Baden, D. G.; Shrank, K. Clinical experience with i.v. mannitol in the treatment of ciguatera. Bull. Soc. Pathol. Exot 1992, 85, 425–426. [Google Scholar]
- Fenner, P. J.; Lewis, R. J.; Williamson, J. A.; Williams, M. L. A Queensland family with ciguatera after eating coral trout. Med. J. Aust 1997, 166, 473–475. [Google Scholar]
- Bagnis, R.; Spiegel, A.; Boutin, J. P.; Burucoa, C.; Nguyen, L.; Cartel, J. L.; Capdevielle, P.; Imbert, P.; Prigent, D.; Gras, C.; et al. (Evaluation of the efficacy of mannitol in the treatment of ciguatera in French Polynesia). Med. Trop. (Mars.) 1992, 52, 67–73.
- De Haro, L.; Hayek-Lanthois, M.; Joossen, F.; Affaton, M. F.; Jouglard, J. (Mass ciguatera poisoning after eating barracuda in Mexico: prognostic and therapeutic implications). Med. Trop. (Mars.) 1997, 57, 55–58.
- Stewart, M. P. Ciguatera fish poisoning: treatment with intravenous mannitol. Trop. Doct 1991, 21, 54–55. [Google Scholar]
- Lewis, R. J.; Endean, R. Occurrence of a ciguatoxin-like substance in the Spanish mackerel (Scomberomorus commersoni). Toxicon 1983, 21, 19–24. [Google Scholar]
- Magovern, G. J., Jr; Bolling, S. F.; Casale, A. S.; Bulkley, B. H.; Gardner, T. J. The mechanism of mannitol in reducing ischemic injury: hyperosmolarity or hydroxyl scavenger? Circulation 1984, 70, I91–95. [Google Scholar]
- Desesso, J. M.; Scialli, A. R.; Goeringer, G. C. d-Mannitol, a specific hydroxyl free radical scavenger, reduces the developmental toxicity of hydroxyurea in rabbits. Teratology 1994, 49, 248–259. [Google Scholar]
- Isbister, G. K.; Kiernan, M. C. Neurotoxic marine poisoning. Lancet Neurol 2005, 4, 219–228. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ciguatoxin | Alternative Name | Sourcea | Mass (Da) | bLD50 (mg/kg, i.p.) | Reference |
|---|---|---|---|---|---|
| P-CTX-1 | CTX, CTX-1B | Carnivore | 1110 | 0.25 | [3,4] |
| P-CTX-2 | 52-epi P-CTX-3 | Carnivore | 1094 | 2.3 | [15] |
| P-CTX-3 | Carnivore | 1094 | 0.9 | [5] | |
| P-CTX-3C | G. toxicus | 1044 | ND | [13] | |
| 2,3-dihydroxyP-CTX-3C | CTX-2A1 | Carnivore | 1056 | 1.8 | [16] |
| 51-hydroxyP-CTX-3C | Carnivore | 1038 | 0.27 | [16] | |
| CTX-4Ac | 52-epi GT-4B | G. toxicus, herbivore | 1060 | 2 | [14] |
| CTX-4Bc | GT-4B | G. toxicus, herbivore | 1060 | 4 | [4] |
| C-CTX-1 | Carnivore | 1140 | 3.6 | [3,11] | |
| C-CTX-2 | 56-epi C-CTX-1 | Carnivore | 1140 | 1 | [3,11] |
| I-CTX-1 | Carnivore | 1140 | ~0.5 | [8] | |
| I-CTX-2 | Carnivore | 1140 | ~0.5 | [8] |
| Sign or Symptom | Percentage of patients with clinical finding | ||
|---|---|---|---|
| Queensland Australia a | Fiji b | French Polynesia New Caledonia c | |
| Neurological | |||
| Paresthesias of the: | |||
| - hands | 73 | - | - |
| - feet | 64 | - | - |
| - mouth | 67 | 52 | 89 |
| Dysesthesia (temperature reversal syndrome) | 77 | 55 | 88 |
| Arthralgia | 79 | 69 | 86 |
| Myalgia | 85 | - | 82 |
| Dental pain | 42 | - | 25 |
| Headache | 64 | - | 59 |
| Weakness | 90 | - | 60 |
| Vertigo | 45 | 38 | 42 |
| Pruritis | 77 | 35 | 45 |
| Chills | 52 | 42 | 59 |
| Paralysis of limbs | 28 | - | 11 |
| Shaking | 31 | – | 27 |
| Neck stiffness | 26 | – | 24 |
| Eye soreness | 43 | – | 22 |
| Sweating | 45 | 34 | 37 |
| Pain on urination | 23 | – | 10 |
| Skin rash | 27 | 2 | 21 |
| Salivation | 12 | 10 | 19 |
| Shortness of breath | 28 | 9 | 16 |
| Gastrointestinal | |||
| Diarrhoea | 65 | 51 | 71 |
| Nausea | 57 | - | 43 |
| Abdominal pain | 54 | 59 | 47 |
| Vomiting | 36 | 30 | 38 |
| Cardiovascular | |||
| Hypotension | - | 1 | 12 |
© 2006 by MDPI Reproduction is permitted for noncommercial purposes.
Share and Cite
Nicholson, G.M.; Lewis, R.J. Ciguatoxins: Cyclic Polyether Modulators of Voltage-gated Iion Channel Function. Mar. Drugs 2006, 4, 82-118. https://doi.org/10.3390/md403082
Nicholson GM, Lewis RJ. Ciguatoxins: Cyclic Polyether Modulators of Voltage-gated Iion Channel Function. Marine Drugs. 2006; 4(3):82-118. https://doi.org/10.3390/md403082
Chicago/Turabian StyleNicholson, Graham M., and Richard J. Lewis. 2006. "Ciguatoxins: Cyclic Polyether Modulators of Voltage-gated Iion Channel Function" Marine Drugs 4, no. 3: 82-118. https://doi.org/10.3390/md403082
APA StyleNicholson, G. M., & Lewis, R. J. (2006). Ciguatoxins: Cyclic Polyether Modulators of Voltage-gated Iion Channel Function. Marine Drugs, 4(3), 82-118. https://doi.org/10.3390/md403082