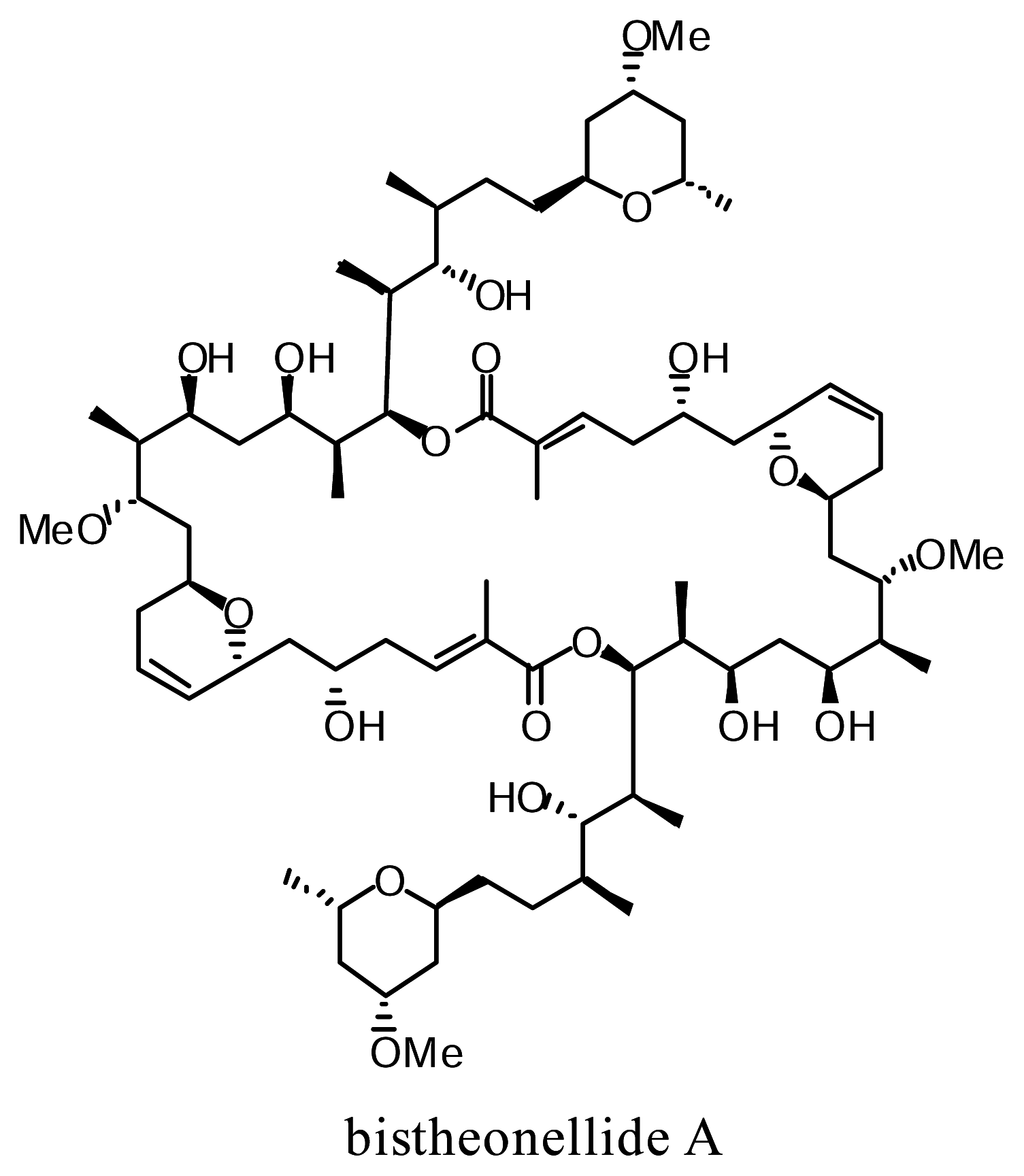
Effects of bistheonellide A, an Actin-polymerization Inhibitor, on Chinese Hamster V79 Cells and on IL-8 Production in PMA-stimulated HL-60 Cells

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell lines and culture conditions
2.3. Relative Plating Efficiency
2.4. Detection of human IL-8 by ELISA
2.5. Determination of cell proliferation
2.6. Determination of cytotoxicity
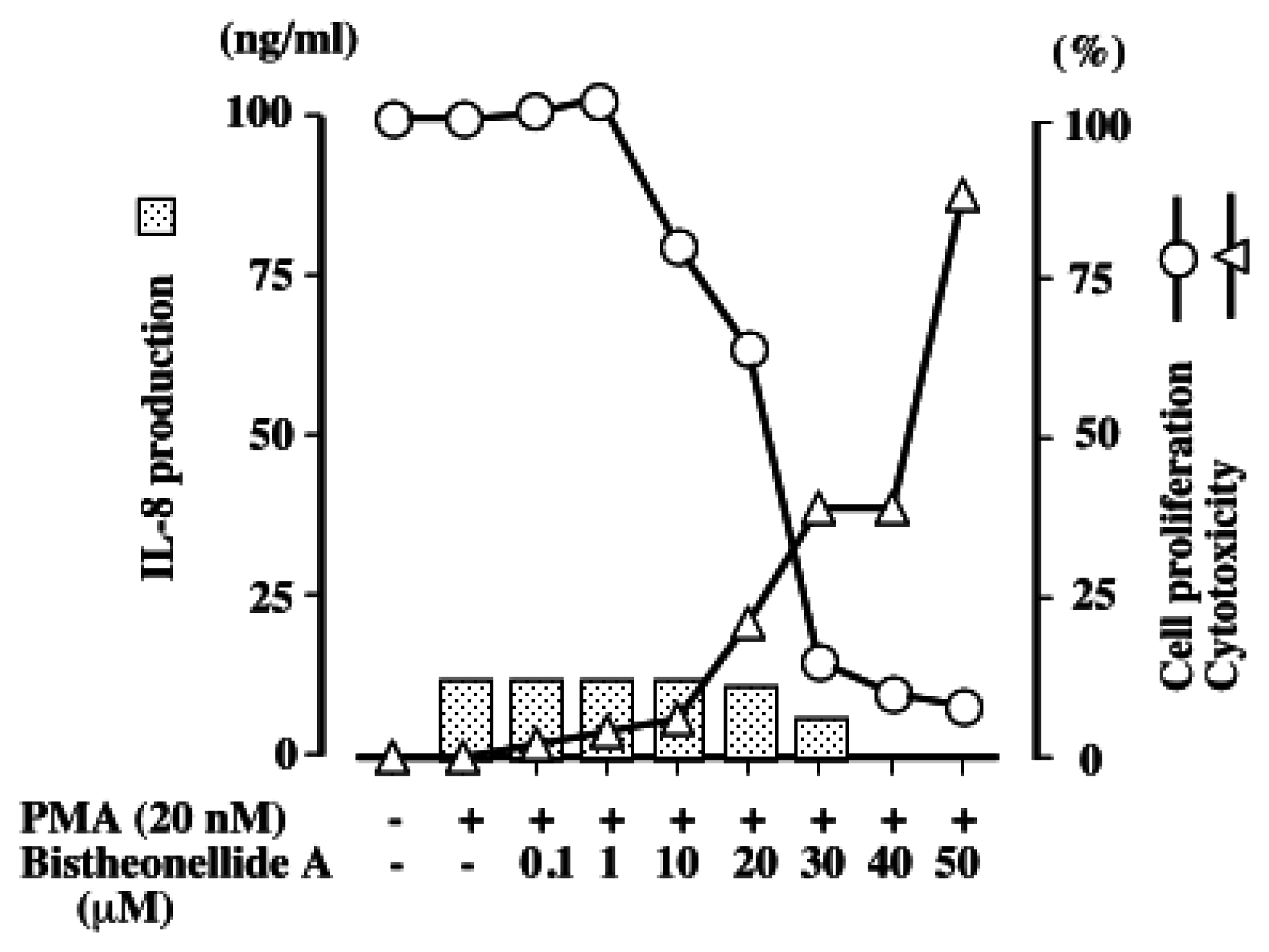
3. Results and Discussion
Acknowledgements
References
- Graves, D. T.; Jiang, Y. Molecular cloning and functional analysis of the promoter of the human squalene synthase gene. Crit. Rev. Oral Biol. Med 1995, 6, 109–118. [Google Scholar]
- Di Celle, P. F.; Carbone, A.; Marchis, D.; Zhou, D.; Sozzani, S.; Zupo, S.; Pini, M.; Mantovani, A.; Foa, R. Cytokine gene expression in B-cell chronic lymphocytic leukemia: evidence of constitutive interleukin-8 (IL-8) mRNA expression and secretion of biologically active IL-8 protein. Blood 1994, 84, 220–228. [Google Scholar]
- Green, A. R.; Green, V. L.; White, M. C.; Speirs, V. Expression of Cytokine messenger RNA in normal and neoplastic human breast tissue: identification of interleukin-8 as a potential regulatory factor in breast tumours. Int. J. Cancer 1997, 72, 937–941. [Google Scholar]
- Konig, B.; Steinbach, F.; Janocha, B.; Drynda, A.; Stumm, M.; Philipp, C.; Allhoff, E. P.; Konig, W. The differential expression of proinflammatory cytokines IL-6, IL-8 and TNF-alpha in renal cell carcinoma. Anticancer Res 1999, 19, 1519–1524. [Google Scholar]
- Galffy, G.; Mohammed, K. A.; Dowling, P. A.; Nasreen, N.; Ward, M. J.; Antony, V. B. Interleukin 8: an autocrine growth factor for malignant mesothelioma. Cancer Res 1999, 59, 367–371. [Google Scholar]
- Kato, Y.; Fusetani, N.; Matsunaga, S.; Hashimoto, K.; Sakai, R.; Higa, T.; Kashman, Y. Antitumor macrolides isolated from a marine sponge Theonella sp.: structure revision of misakinolide A. Tetrahedron Lett 1987, 28, 6225–6228. [Google Scholar]
- Watabe, S.; Wada, S.; Saito, S.; Matsunaga, S.; Fusetani, N.; Ozaki, H.; Karaki, H. Cellular changes of rat embryonic fibroblasts by an actin-polymerization inhibitor, bistheonellide A, from a marine sponge. Cell Struct. Funct 1996, 21, 199–212. [Google Scholar]
- Saito, S.; Karaki, H. A family of novel actin-inhibiting marine toxins. Clin. Exp. Pharmcol. Physiol 1996, 23, 743–746. [Google Scholar]
- Sato, Y.; Sakakibara, Y.; Oda, T.; Aizu-Yokota, E.; Ichinoseki, I. Effects of estradiol and ethynylestradiol on microtubule distribution in Chinese hamster V79 cells. Chem. Pharm. Bull 1992, 40, 182–184. [Google Scholar]
- Kasahara, T.; Oda, T.; Hatake, K.; Akiyama, M.; Mukaida, N.; Matsushima, K. Interleukin-8 and monocyte chemotactic protein-1 production by a human glioblastoma cell line, T98G in coculture with monocytes: involvement of monocyte-derived interleukin-1alpha. Eur. Cytokine Netw 1998, 9, 47–55. [Google Scholar]
- Carmichael, J.; DeGraff, W. G.; Gazdar, A. F.; Minna, J. D.; Mitchell, J. B. Evaluation of a tetrazolium-based semiautomated colorimetric assay: assessment of chemosensitivity testing. Cancer Res 1987, 47, 939–942. [Google Scholar]
- Shimizu, S.; Nomoto, M.; Naito, S.; Yamamoto, T.; Momose, K. Stimulation of nitric oxide synthase during oxidative endothelial cell injury. Biochem. Pharmacol. 1998, 55, 77–83. [Google Scholar]
- Saito, S. Y.; Watanabe, S.; Ozaki, H.; Kobayashi, M.; Suzuki, T.; Kobayashi, H.; Fusetani, N.; Karaki, H. Actin-depolymerizing effect of dimeric macrolides, bistheonellide A and swinholide A. J. Biochem 1998, 123, 571–578. [Google Scholar]
- Kasahara, T.; Mukaida, N.; Yamashita, K.; Yagisawa, H.; Akahoshi, T.; Matsushima, K. IL-1 and TNF-α induction of IL-8 and monocyte chemotactie and activating factor (MCAF) mRNA espression in a human astrocytoma cell line. Immunology 1991, 74, 60–67. [Google Scholar]
- Brasier, A. R.; Jamaluddin, R. M.; Casola, A.; Duan, W.; Shen, Q.; Garofalo, R. P. A promotor recruitment mechanism for tumor necrosis factor-a-induced interleukin-8 transcription in type II pulmonary epithelial cells. J. Biol. Chem 1996, 273, 3551–3561. [Google Scholar]
- Meier, R. W.; Niklaus, G.; Dewald, B.; Fey, M. F.; Tobler, A. Inhibition of the arachidonic acid pathway prevents induction of IL-8 mRNA by phorbol ester and changes the release of IL-8 from HL-60 cells: differential inhibition of induced expression of IL-8, TNF-alpha, IL-1 alpha, and IL-1 beta. J. Cell Physiol 1995, 165, 62–70. [Google Scholar]
- Tomoda, H.; Namatame, I.; Si, S.; Kawaguchi, K.; Masuma, R.; Namikoshi, M.; Omura, S. Phenochalasins, inhibitors of lipid droplet formation in mouse macrophages, produced by Phomopsis sp. FT-0211. J. Antibiot 1999, 52, 851–856. [Google Scholar]
- Etil, J.; Heise, T.; Dersch, P. Cell invasion and IL-8 production pathways initiated by YadA of Yersinia pseudotuberculosis require common signalling molecules (FAK, c-Src, Ras) and distinct cell factors. Cellular Micribiol 2005, 7, 63–77. [Google Scholar]
- Sample availability: not available.


© 2006 by MDPI Reproduction is permitted for noncommercial purposes.
Share and Cite
Oda, T.; Xu, J.; Fujita, A.; Mochizuki, M.; Namikoshi, M. Effects of bistheonellide A, an Actin-polymerization Inhibitor, on Chinese Hamster V79 Cells and on IL-8 Production in PMA-stimulated HL-60 Cells. Mar. Drugs 2006, 4, 22-27. https://doi.org/10.3390/md401022
Oda T, Xu J, Fujita A, Mochizuki M, Namikoshi M. Effects of bistheonellide A, an Actin-polymerization Inhibitor, on Chinese Hamster V79 Cells and on IL-8 Production in PMA-stimulated HL-60 Cells. Marine Drugs. 2006; 4(1):22-27. https://doi.org/10.3390/md401022
Chicago/Turabian StyleOda, Taiko, Jinzhong Xu, Ayako Fujita, Masataka Mochizuki, and Michio Namikoshi. 2006. "Effects of bistheonellide A, an Actin-polymerization Inhibitor, on Chinese Hamster V79 Cells and on IL-8 Production in PMA-stimulated HL-60 Cells" Marine Drugs 4, no. 1: 22-27. https://doi.org/10.3390/md401022
APA StyleOda, T., Xu, J., Fujita, A., Mochizuki, M., & Namikoshi, M. (2006). Effects of bistheonellide A, an Actin-polymerization Inhibitor, on Chinese Hamster V79 Cells and on IL-8 Production in PMA-stimulated HL-60 Cells. Marine Drugs, 4(1), 22-27. https://doi.org/10.3390/md401022