
Chemical Compositions and Experimental and Computational Modeling of the Anticancer Effects of Cnidocyte Venoms of Jellyfish Cassiopea andromeda and Catostylus mosaicus on Human Adenocarcinoma A549 Cells

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Results
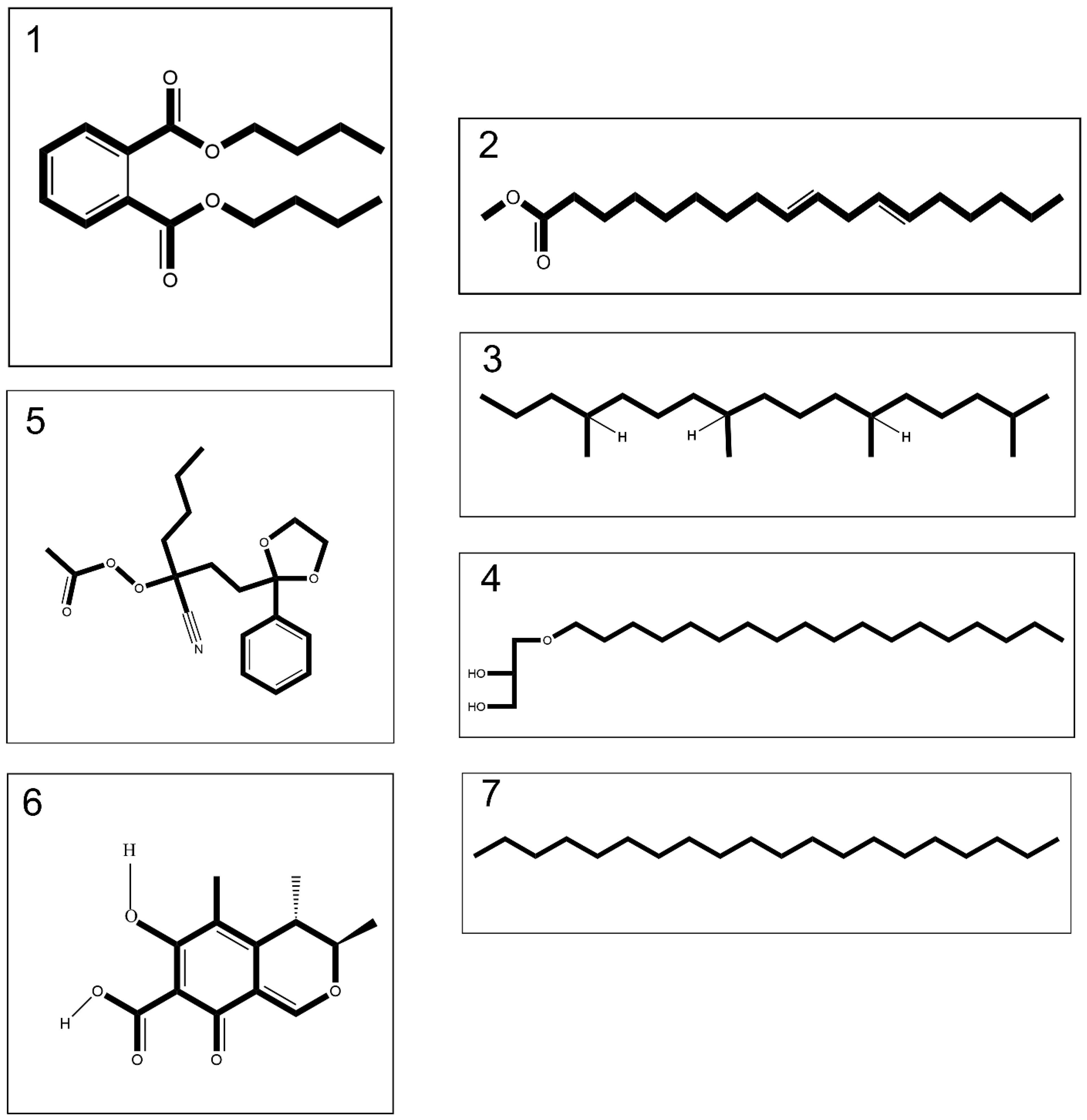
2.1. GC-MS Findings of Venoms of C. andromeda and C. mosaicus
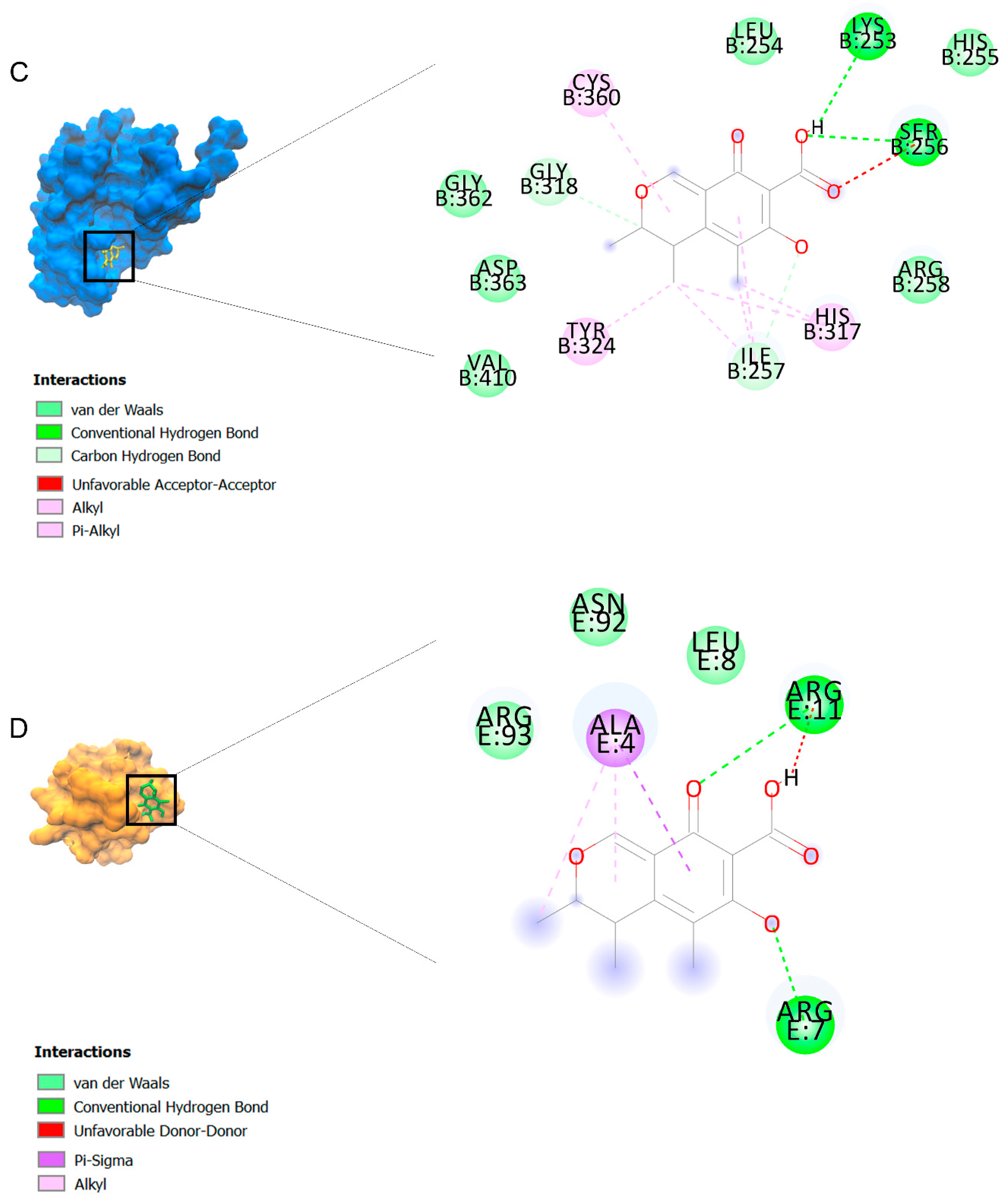
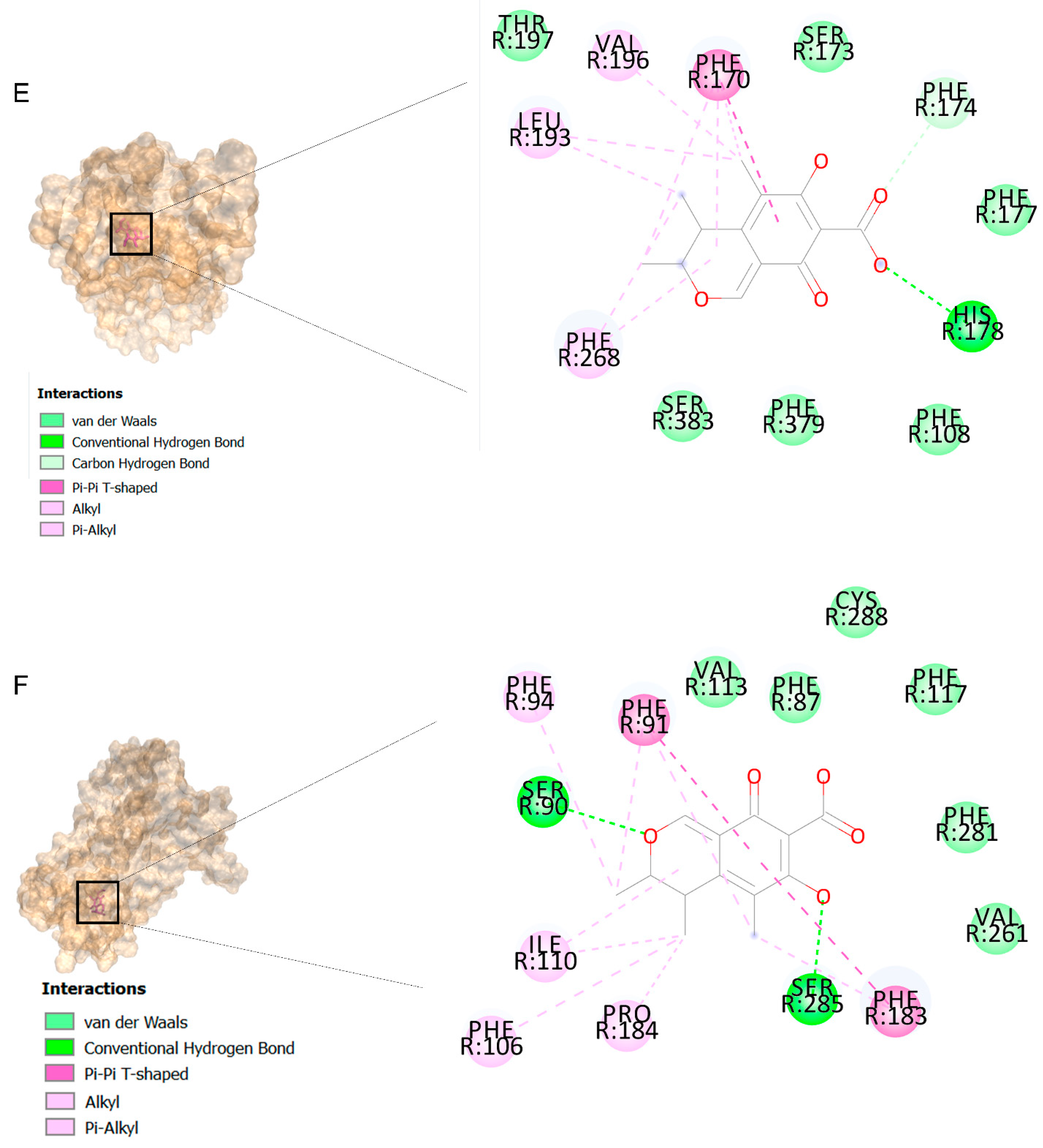
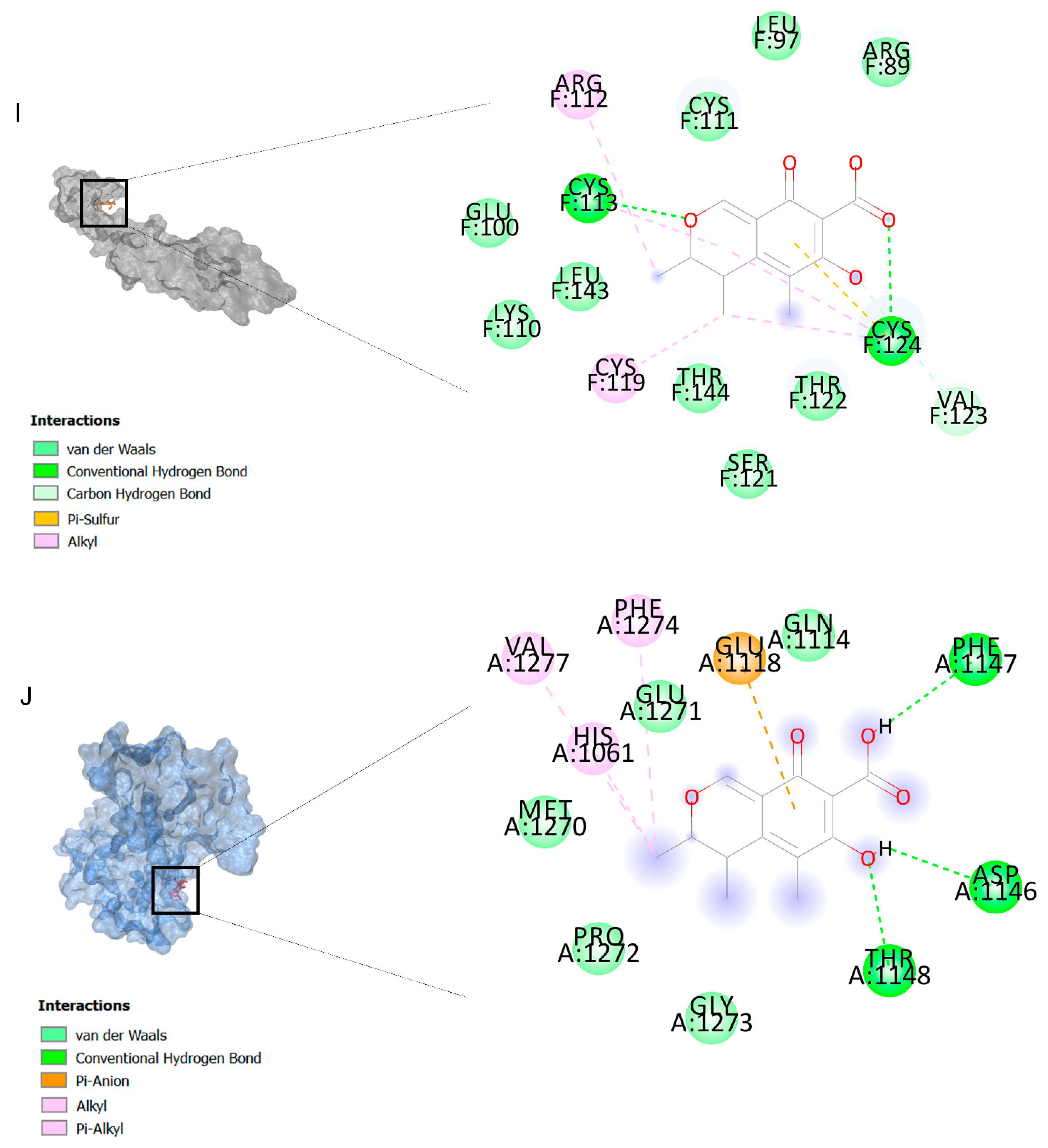
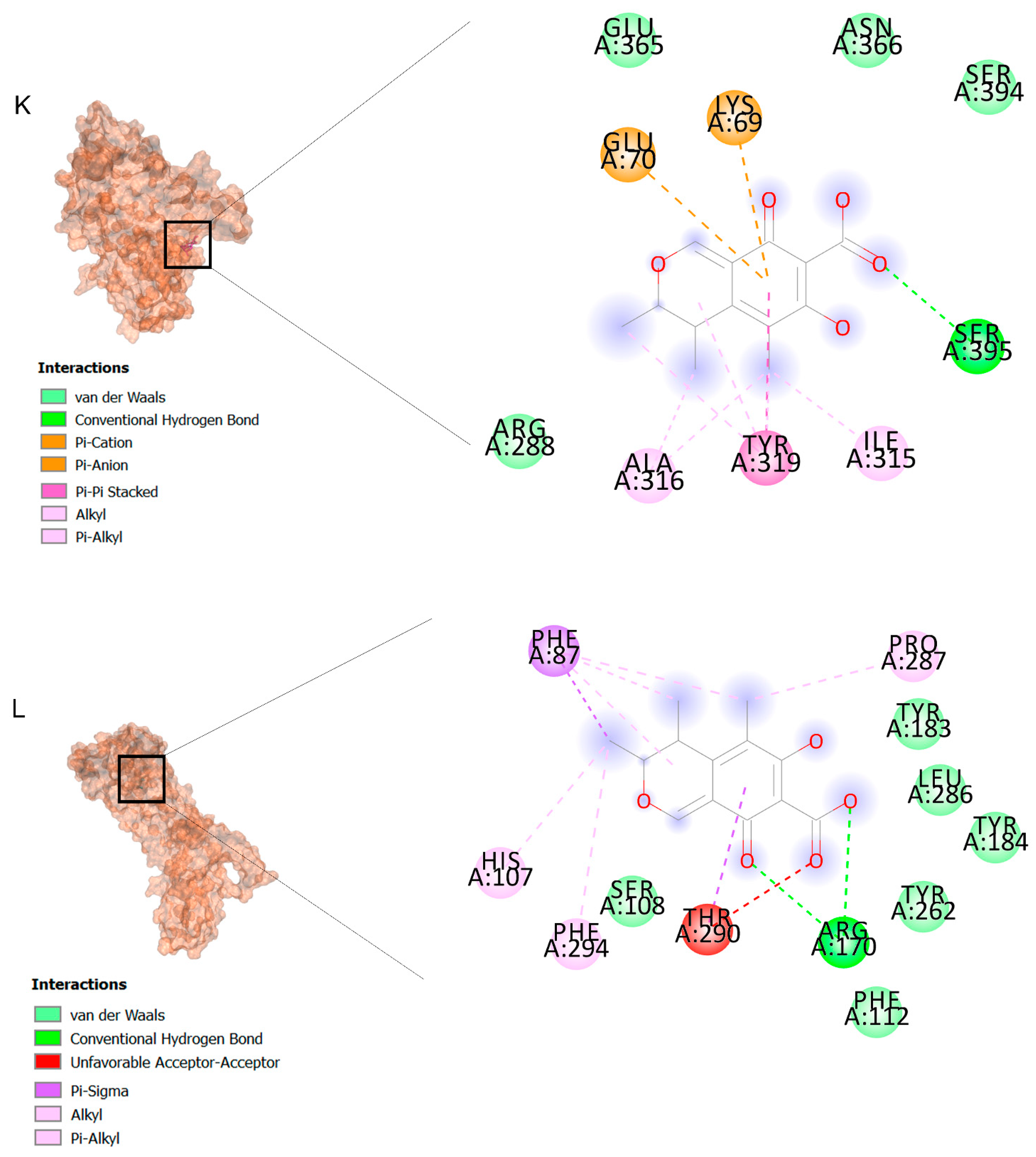
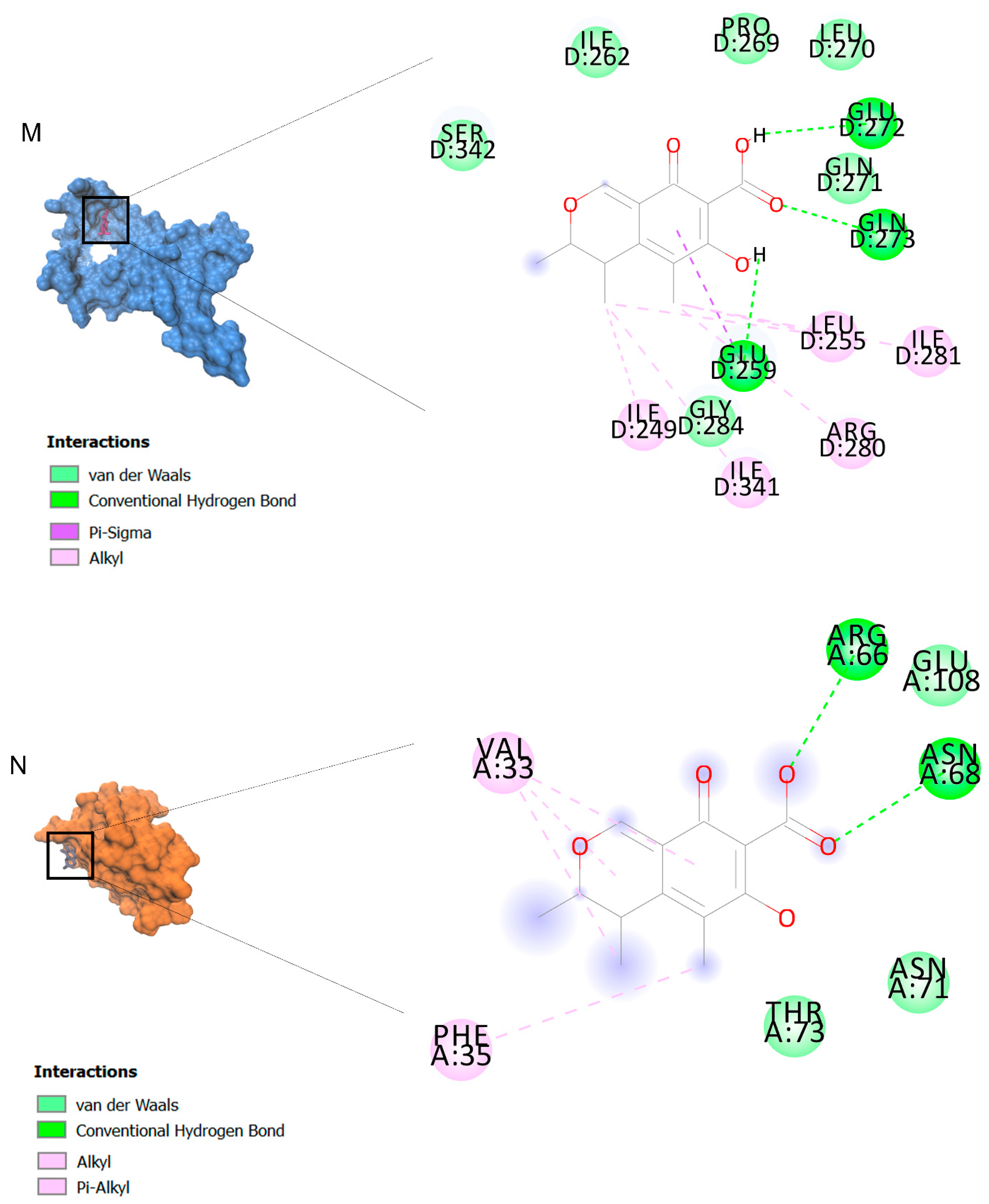
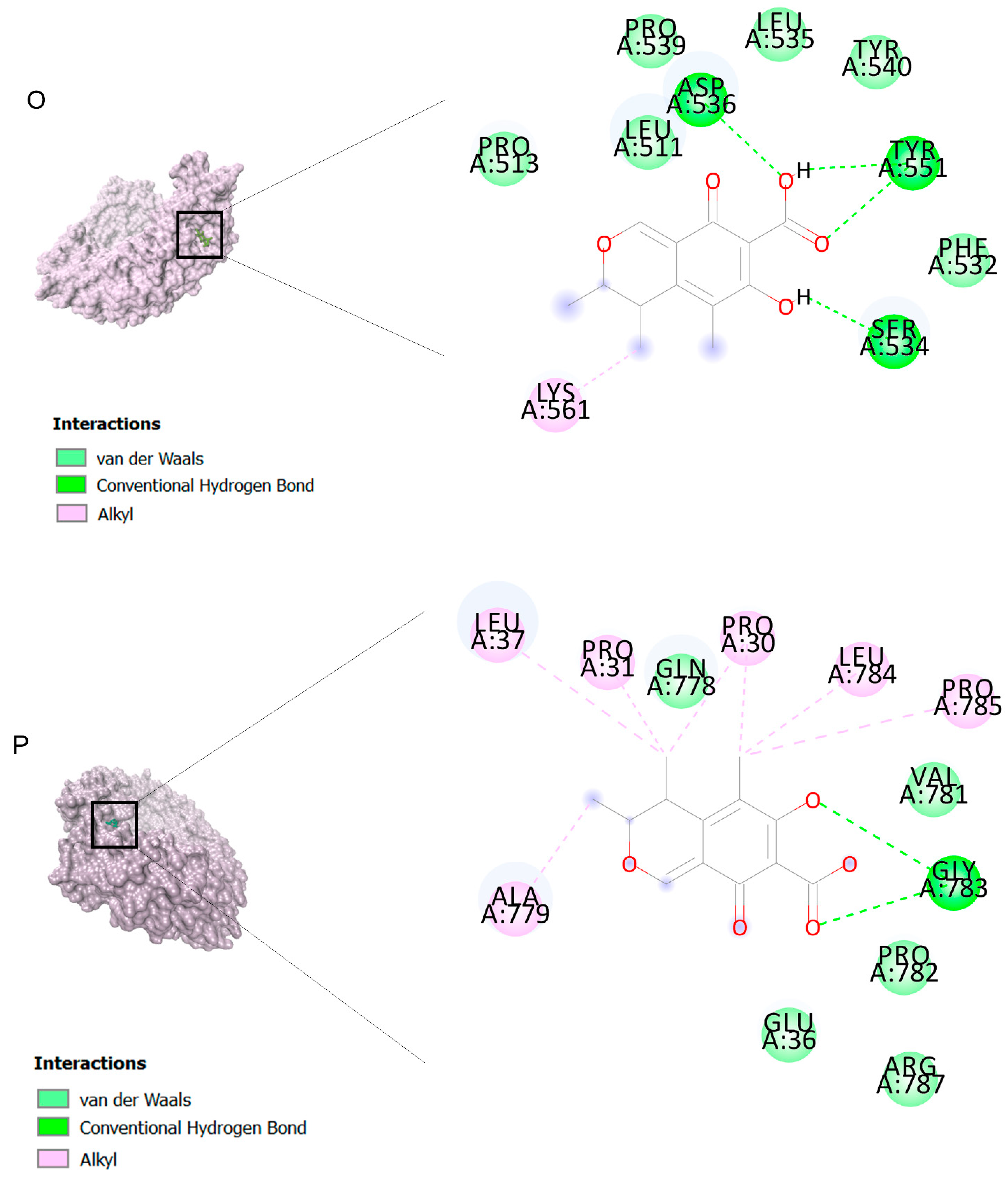
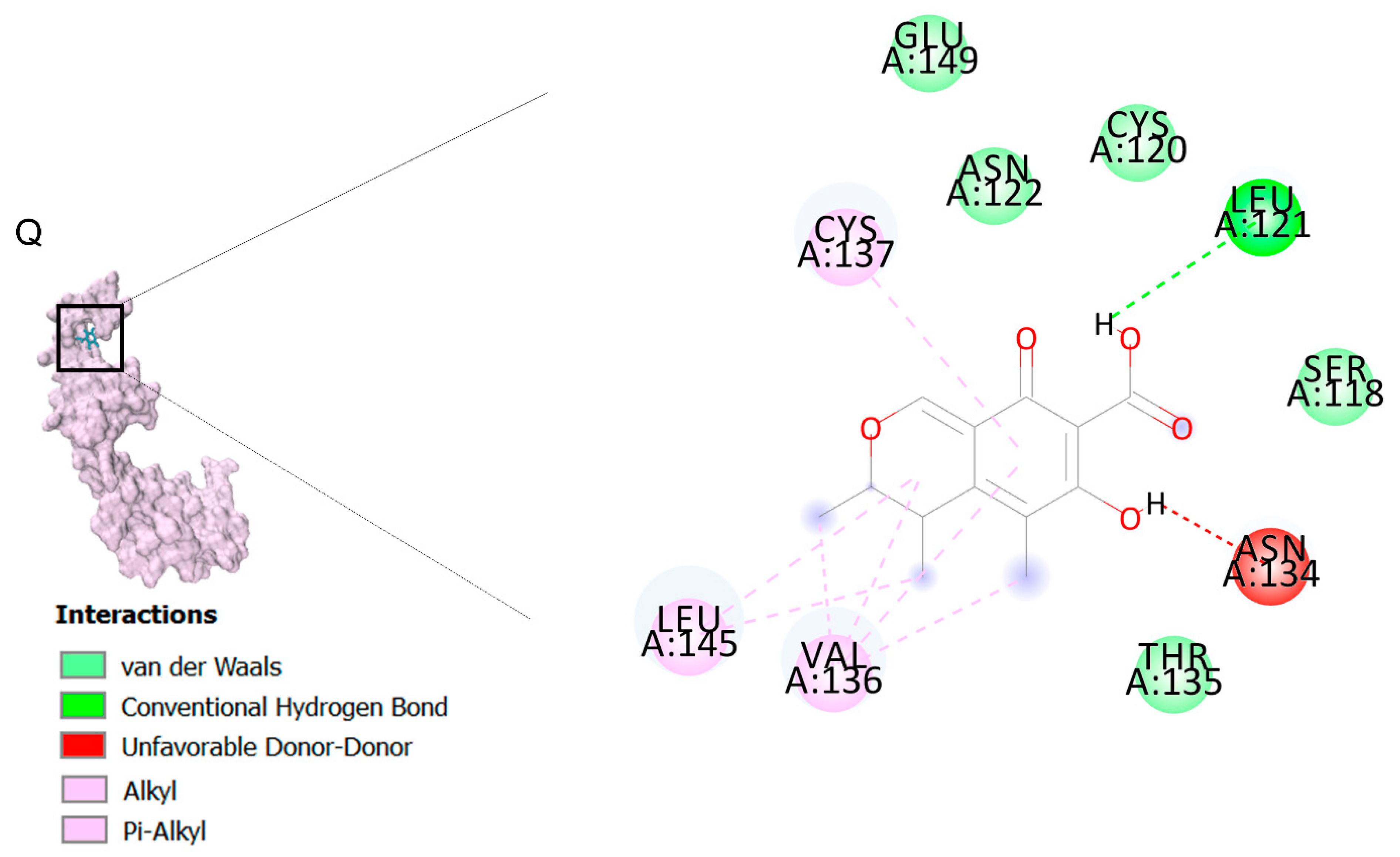
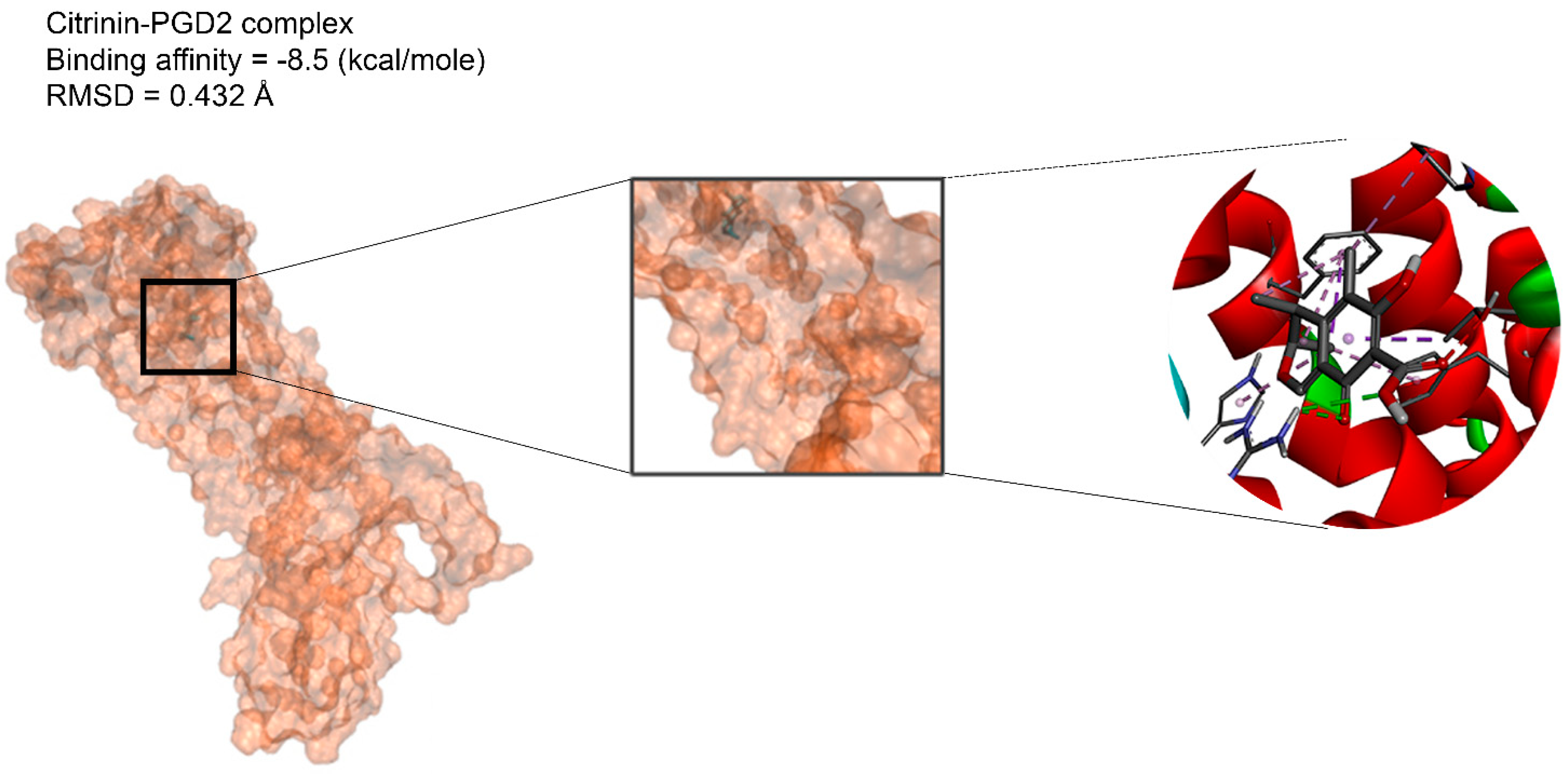
2.2. Docking Findings
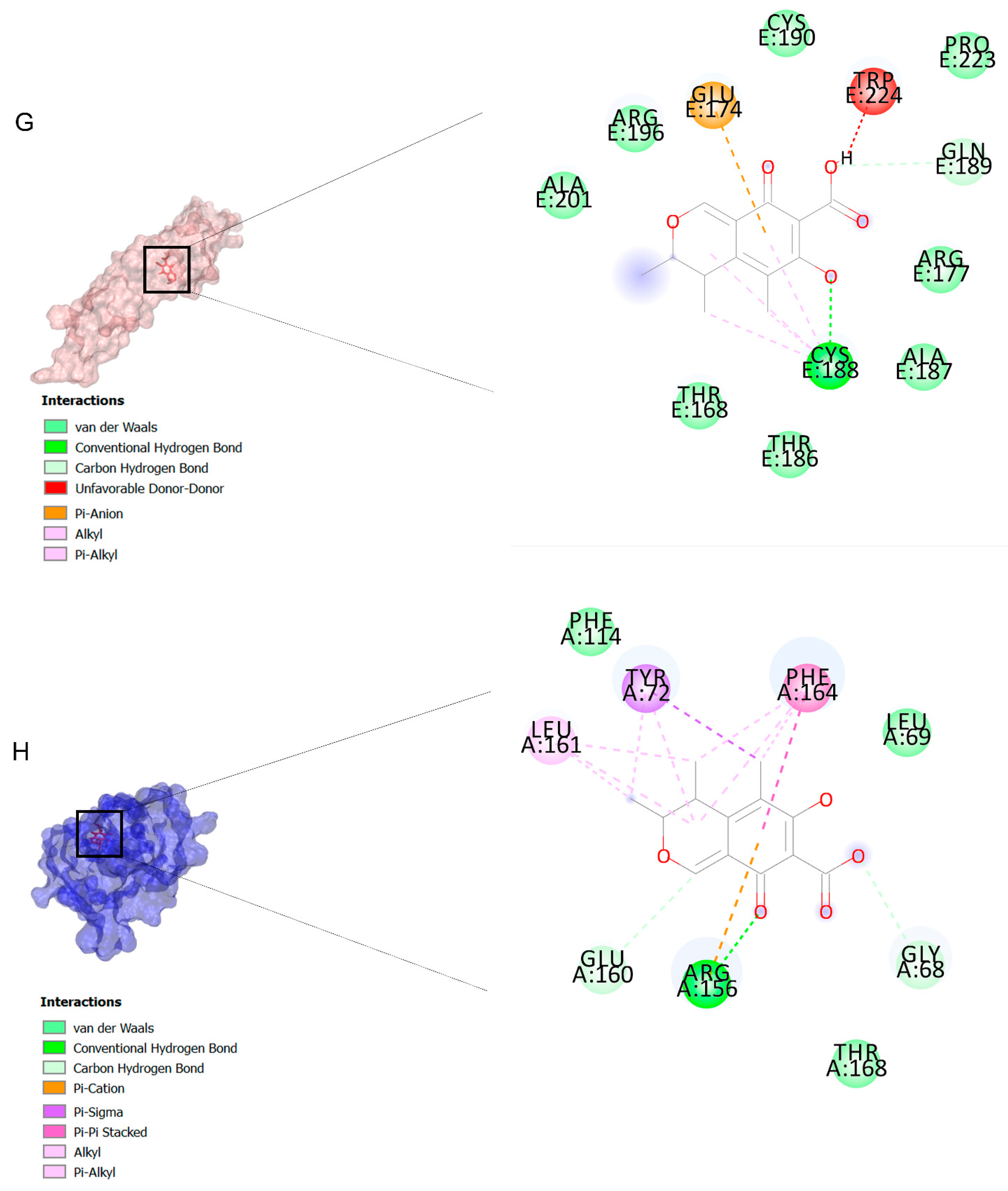
2.3. Molecular Dynamic (MD) Findings
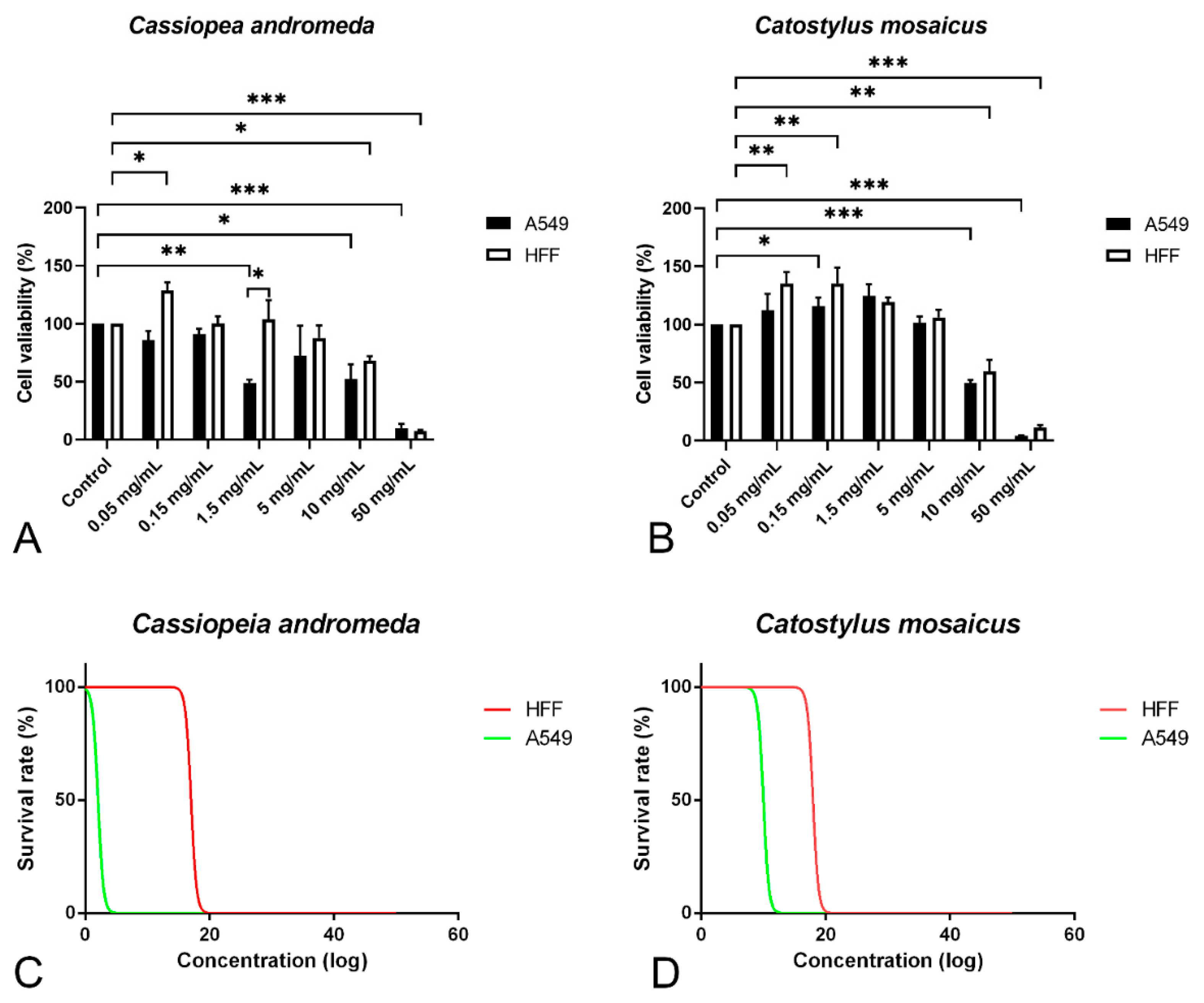
2.4. MTT Assay Showed Anti-Proliferative Effects of Venoms of C. andromeda and C. mosaicus on A549 Cells
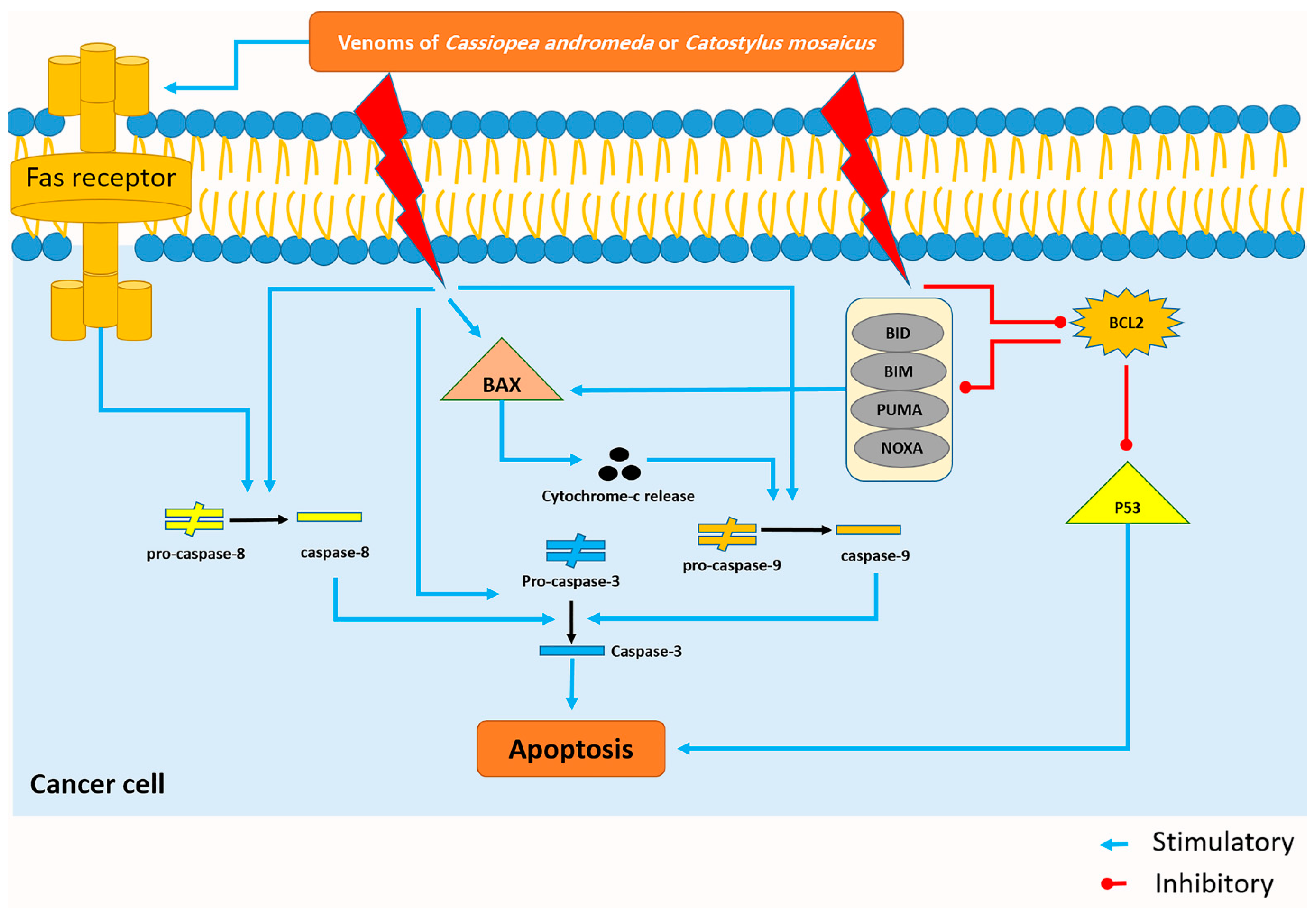
2.5. Western Blot Assay Findings
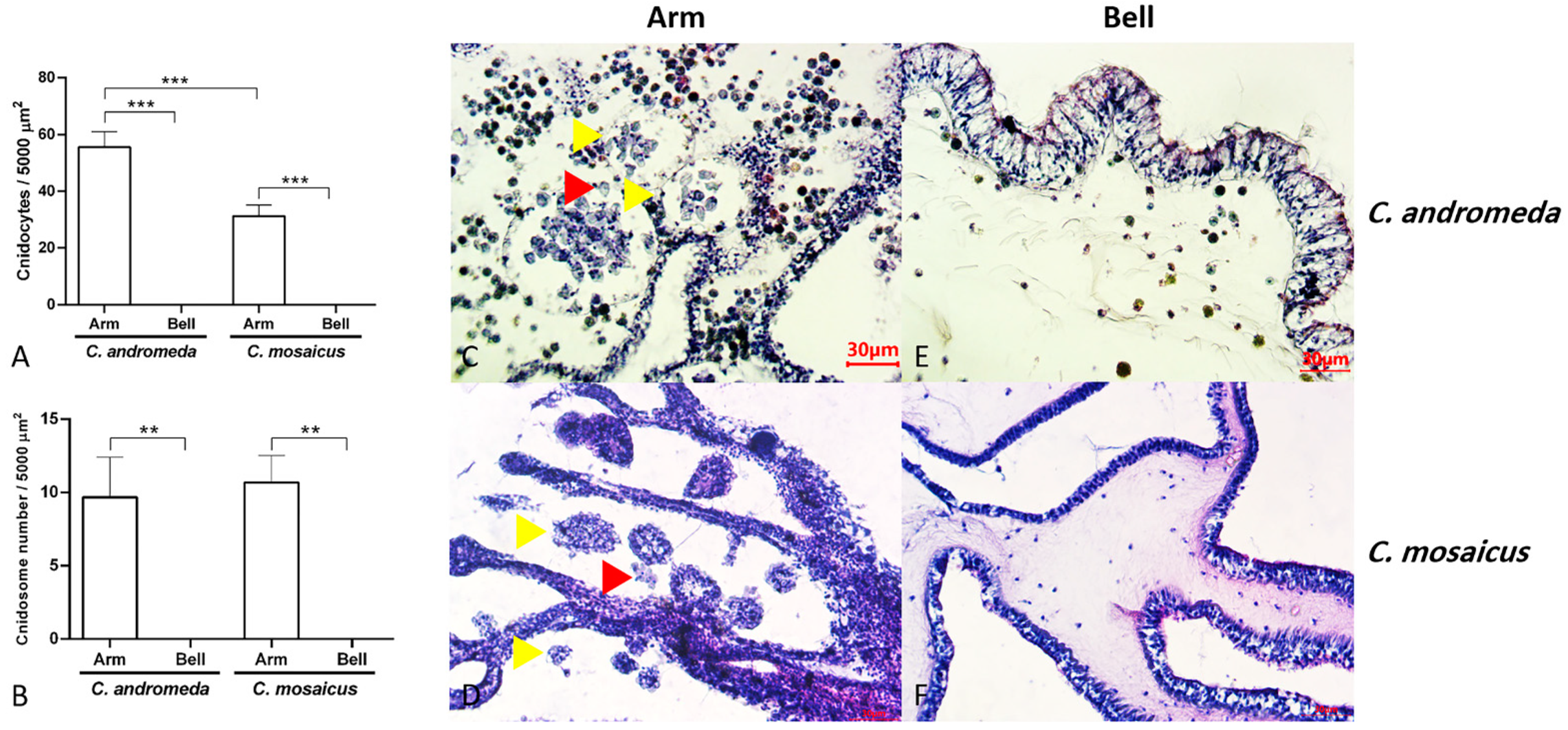
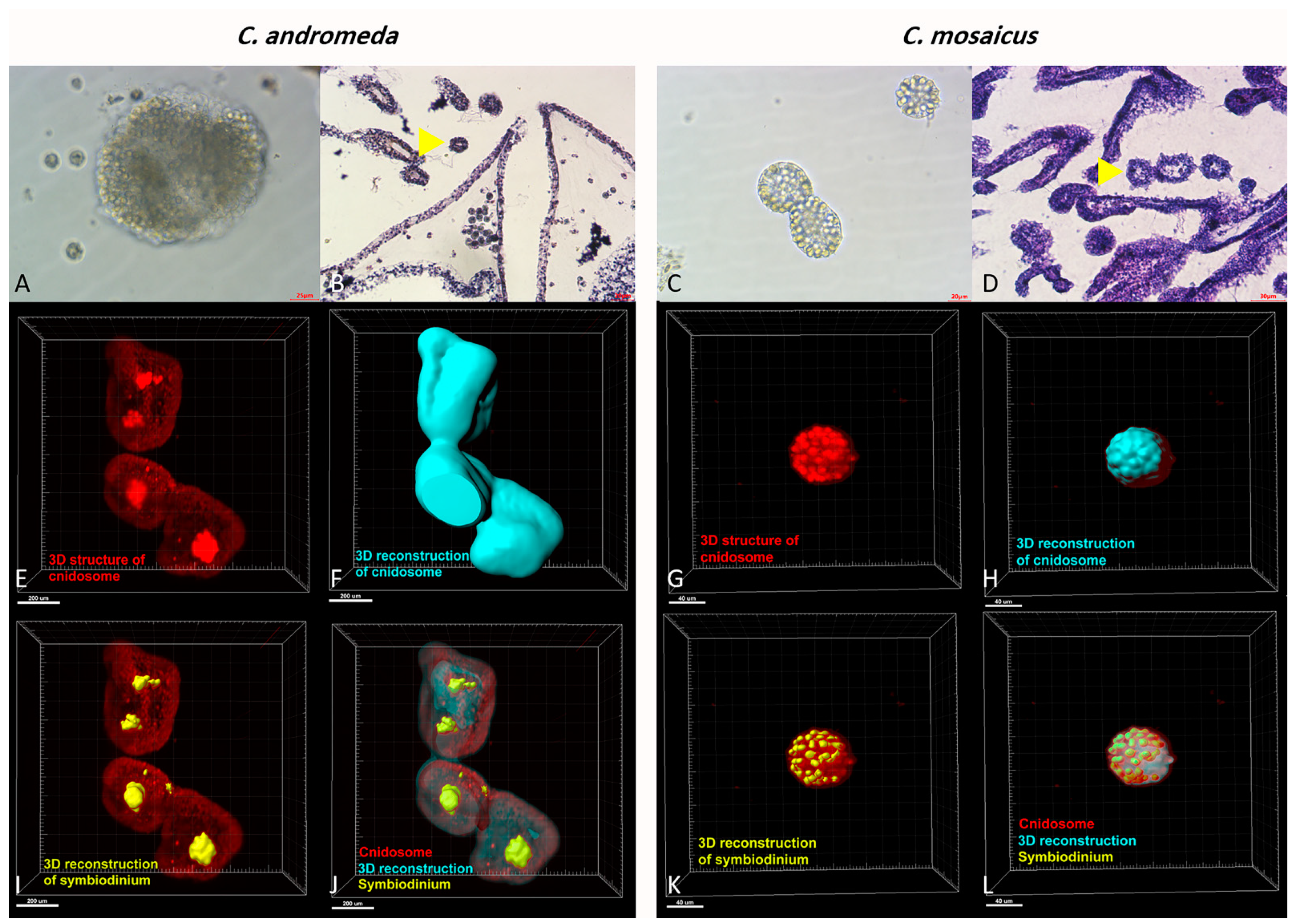
2.6. Cnidocytes and Cnidosomes: Histology and Histomorphology
3. Discussion
3.1. Venom of C. andromeda and Venom of C. mosaicus Have Anti-Cancer Effects on A549 Cells
3.2. The Venom of C. andromeda and C. mosaicus Induced Apoptosis in A549 Cells
3.3. Molecular Docking and MD Simulation Findings Showed Two Molecules with Anti-Cancer Effect
3.4. C. andromeda and C. mosaicus Venoms Can Be Isolated from Cnidocytes and Cnidosomes
4. Materials and Methods
4.1. Jellyfish Collection and Adaptation
4.2. Histological Distribution of Cnidocytes and Cnidosomes in Jellyfish Body
4.3. Isolation of Cnidocytes and Cnidosomes in Jellyfish Body
4.4. Light Sheet Imaging of Jellyfish Cnidosomes
4.5. Imaris Reconstruction of Light Sheet Images of Jellyfish Cnidosomes
4.6. Venom Isolation from Cnidocytes of Jellyfish
4.7. A549 Cell Culture and Treatment with Venoms of C. andromeda and C. mosaicus
4.8. GC-MS Analysis of Venoms of C. andromeda and C. mosaicus
4.9. Western Blot Analysis of Apoptosis Pathways in A549 Cells after Exposure to Venoms of C. andromeda and C. mosaicus
4.10. Molecular Interactions and Docking Studies of A549 Apoptotic Pathways and Effective Molecules of Jellyfish’ Venoms
4.11. MD Simulation Studies for Effective Molecules of Jellyfish’ Venoms
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, D.; Yang, W.; Zhang, Y.; Yang, J.Y.; Guan, R.; Xu, D.; Yang, M.Q. Genomic analyses based on pulmonary adenocarcinoma in situ reveal early lung cancer signature. BMC Med. Genom. 2018, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Zappa, C.; Mousa, S.A. Non-small cell lung cancer: Current treatment and future advances. Transl. Lung Cancer Res. 2016, 5, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.E.; Meehan, J.; Sullivan, P.; Marland, J.R.K.; Greenhalgh, S.N.; Gregson, R.; Clutton, R.E.; Ward, C.; Cousens, C.; Griffiths, D.J.; et al. Ovine pulmonary adenocarcinoma: A unique model to improve lung cancer research. Front. Oncol. 2019, 9, 335. [Google Scholar] [CrossRef] [PubMed]
- Ali Mohammed Hammamy, R.; Farooqui, K.; Ghadban, W. Sclerotic bone metastasis in pulmonary adenocarcinoma. Case Rep. Med. 2018, 2018, 5. [Google Scholar] [CrossRef]
- Zhang, B.; Fan, J.; Zhang, X.; Shen, W.; Cao, Z.; Yang, P.; Xu, Z.; Ju, D. Targeting asparagine and autophagy for pulmonary adenocarcinoma therapy. Appl. Microbiol. Biotechnol. 2016, 100, 9145–9161. [Google Scholar] [CrossRef]
- Yuan, M.; Huang, L.L.; Chen, J.H.; Wu, J.; Xu, Q. The emerging treatment landscape of targeted therapy in non-small-cell lung cancer. Signal Transduct. Target Ther. 2019, 4, 61. [Google Scholar] [CrossRef]
- Dyshlovoy, S.A.; Honecker, F. Marine compounds and cancer: Updates 2020. Mar. Drugs 2020, 18, 643. [Google Scholar] [CrossRef]
- Pereira, R.B.; Evdokimov, N.M.; Lefranc, F.; Valentao, P.; Kornienko, A.; Pereira, D.M.; Andrade, P.B.; Gomes, N.G.M. Marine-derived anticancer agents: Clinical benefits, innovative mechanisms, and new targets. Mar. Drugs 2019, 17, 329. [Google Scholar] [CrossRef]
- D'Ambra, I.; Lauritano, C. A review of toxins from cnidaria. Mar. Drugs 2020, 18, 507. [Google Scholar] [CrossRef]
- Bosch, T.C.G.; Klimovich, A.; Domazet-Loso, T.; Grunder, S.; Holstein, T.W.; Jekely, G.; Miller, D.J.; Murillo-Rincon, A.P.; Rentzsch, F.; Richards, G.S.; et al. Back to the basics: Cnidarians start to fire. Trends Neurosci. 2017, 40, 92–105. [Google Scholar] [CrossRef]
- Morabito, R.; Costa, R.; Rizzo, V.; Remigante, A.; Nofziger, C.; La Spada, G.; Marino, A.; Paulmichl, M.; Dossena, S. Crude venom from nematocysts of Pelagia noctiluca (Cnidaria: Scyphozoa) elicits a sodium conductance in the plasma membrane of mammalian cells. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Jayathilake, J.; Gunathilake, K. Cnidarian toxins: Recent evidences for potential therapeutic uses. Eur. Zool. J. 2020, 87, 708–713. [Google Scholar] [CrossRef]
- Ha, S.-H.; Jin, F.; Kwak, C.-H.; Abekura, F.; Park, J.-Y.; Park, N.G.; Chang, Y.-C.; Lee, Y.-C.; Chung, T.-W.; Ha, K.-T. Jellyfish extract induces apoptotic cell death through the p38 pathway and cell cycle arrest in chronic myelogenous leukemia K562 cells. PeerJ 2017, 5, 18. [Google Scholar] [CrossRef]
- Nisa, S.A.; Vinu, D.; Krupakar, P.; Govindaraju, K.; Sharma, D.; Vivek, R. Jellyfish venom proteins and their pharmacological potentials: A review. Int. J. Biol. Macromol. 2021, 176, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Bae, S.K.; Kim, M.; Pyo, M.J.; Kim, M.; Yang, S.; Won, C.K.; Yoon, W.D.; Han, C.H.; Kang, C.; et al. Anticancer effect of Nemopilema nomurai jellyfish venom on HepG2 cells and a tumor xenograft animal model. Evid. Based Complement. Altern. Med. 2017, 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Maduraiveeran, H.; Raja, K.; Chinnasamy, A. Antiproliferative and antioxidant properties of nematocysts crude venom from jellyfish Acromitus flagellatus against human cancer cell lines. Saudi J. Biol. Sci. 2021, 28, 1954–1961. [Google Scholar] [CrossRef] [PubMed]
- Holland, B.S.; Dawson, M.N.; Crow, G.L.; Hofmann, D.K. Global phylogeography of Cassiopea (Scyphozoa: Rhizostomeae): Molecular evidence for cryptic species and multiple invasions of the Hawaiian Islands. Mar. Biol. 2004, 145, 1119–1128. [Google Scholar] [CrossRef]
- Mirshamsi, M.R.; Omranipour, R.; Vazirizadeh, A.; Fakhri, A.; Zangeneh, F.; Mohebbi, G.H.; Seyedian, R.; Pourahmad, J. Persian Gulf jellyfish (Cassiopea andromeda) venom fractions induce selective injury and cytochrome c release in mitochondria obtained from breast adenocarcinoma patients. Asian Pac. J. Cancer Prev. 2017, 18, 277–286. [Google Scholar] [CrossRef]
- Dehghani, R.; Obeidi, N.; Mohebbi, G.; Amrooni, A.; Derakhshan, Z. Effect of Cassiopea andromeda venom on P15INK4b, P21 WAF1/CIP1, P53, DNA methyltransferase 1, and Bcl-2 genes expression, apoptosis induction, and cell growth inhibition in acute promyelocytic leukemia NB4 cell line. Iran. J. Pediatr. Hematol. Oncol. 2021, 11, 96–104. [Google Scholar] [CrossRef]
- Rastian, Z.; Putz, S.; Wang, Y.; Kumar, S.; Fleissner, F.; Weidner, T.; Parekh, S.H. Type I collagen from jellyfish Catostylus mosaicus for biomaterial applications. ACS Biomater. Sci. Eng. 2018, 4, 2115–2125. [Google Scholar] [CrossRef]
- Pitt, K.A.; Kingsford, M.J. Temporal variation in the virgin biomass of the edible jellyfish, Catostylus mosaicus (Scyphozoa, Rhizostomeae). Fish. Res. 2003, 63, 303–313. [Google Scholar] [CrossRef]
- Sujanitha, V.; Sivansuthan, S.; Luckshman, W.V.; Gnaneswaran, R.; Jeyakanth, T.; Gunarathna, U. The clinical manifestations, outcome and identification of jellyfish stings in Jaffna, Sri Lanka. Trop. Doct. 2017, 47, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Azila, N.; Othman, I. Haemolytic and phospholipase A activities of the tentacle extract of Catostylus mosaicus. Biochem. Int. 1990, 20, 291–299. [Google Scholar]
- Mohebbi, G.; Nabipour, I.; Vazirizadeh, A.; Vatanpour, H.; Farrokhnia, M.; Maryamabadi, A.; Bargahi, A. Acetylcholinesterase inhibitory activity of a neurosteroidal alkaloid from the upside-down jellyfish Cassiopea andromeda venom. Rev. Bras. Farmacogn. 2018, 28, 568–574. [Google Scholar] [CrossRef]
- Wiltshire, C.J.; Sutherland, S.K.; Fenner, P.J.; Young, A.R. Optimization and preliminary characterization of venom isolated from 3 medically important jellyfish: The box (Chironex fleckeri), Irukandji (Carukia barnesi), and blubber (Catostylus mosaicus) jellyfish. Wilderness Environ. Med. 2000, 11, 241–250. [Google Scholar] [CrossRef]
- Leone, A.; Lecci, R.M.; Durante, M.; Piraino, S. Extract from the zooxanthellate jellyfish Cotylorhiza tuberculata modulates gap junction intercellular communication in human cell cultures. Mar. Drugs 2013, 11, 1728–1762. [Google Scholar] [CrossRef] [PubMed]
- Khalil, E.A.; Swelim, H.; El-Tantawi, H.; Bakr, A.F.; Abdellatif, A. Characterization, cytotoxicity and antioxidant activity of sea urchins (Diadema savignyi) and jellyfish (Aurelia aurita) extracts. Egypt. J. Aquat. Res. 2022. [Google Scholar] [CrossRef]
- Nabipour, I.; Mohebbi, G.; Vatanpour, H.; Vazirizadeh, A. Hematological parameters on the effect of the jellyfish venom Cassiopea andromeda in animal models. Data Br. 2017, 11, 517–521. [Google Scholar] [CrossRef]
- Azila, N.; Siao, F.K.; Othman, I. Haemolytic, oedema and haemorrhage inducing activities of tentacular extract of the blubber jellyfish (Catostylus mosaicus). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 1991, 99, 153–156. [Google Scholar] [CrossRef]
- Balamurugan, E.; Reddy, B.V.; Menon, V.P. Antitumor and antioxidant role of Chrysaora quinquecirrha (sea nettle) nematocyst venom peptide against ehrlich ascites carcinoma in swiss albino mice. Mol. Cell. Biochem. 2010, 338, 69–76. [Google Scholar] [CrossRef]
- Ramezanpour, M.; Burke da Silva, K.; Sanderson, B. Differential susceptibilities of human lung, breast and skin cancer cell lines to killing by five sea anemone venoms. J. Venom. Anim. Toxins Incl. Trop. Dis. 2012, 18, 157–163. [Google Scholar] [CrossRef]
- Maharajasri, V.; Devamalar, D. Characterization of antibacterial, anticancer properties and bioactive compounds of methanolic leaf extract of catharanthus roseus. Int. J. Humanit. Arts Med. Sci. 2015, 1, 35–42. [Google Scholar]
- de Oliveira Filho, J.W.G.; Islam, M.T.; Ali, E.S.; Uddin, S.J.; Santos, J.V.O.; de Alencar, M.; Junior, A.L.G.; Paz, M.; de Brito, M.; JMC, E.S.; et al. A comprehensive review on biological properties of citrinin. Food Chem. Toxicol. 2017, 110, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Porter, A.G.; Janicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Kuida, K. Caspase-9. Int. J. Biochem. Cell Biol. 2000, 32, 121–124. [Google Scholar] [CrossRef]
- Roy, R.N. Bioactive natural derivatives of phthalate ester. Crit. Rev. Biotechnol. 2020, 40, 913–929. [Google Scholar] [CrossRef]
- Reza, A.; Haque, M.A.; Sarker, J.; Nasrin, M.S.; Rahman, M.M.; Tareq, A.M.; Khan, Z.; Rashid, M.; Sadik, M.G.; Tsukahara, T.; et al. Antiproliferative and antioxidant potentials of bioactive edible vegetable fraction of Achyranthes ferruginea Roxb. in cancer cell line. Food Sci. Nutr. 2021, 9, 3777–3805. [Google Scholar] [CrossRef]
- Tutino, V.; Caruso, M.G.; De Nunzio, V.; Lorusso, D.; Veronese, N.; Gigante, I.; Notarnicola, M.; Giannelli, G. Down-regulation of cannabinoid type 1 (CB1) receptor and its downstream signaling pathways in metastatic colorectal cancer. Cancers 2019, 11, 708. [Google Scholar] [CrossRef]
- Refolo, M.G.; D’Alessandro, R.; Malerba, N.; Laezza, C.; Bifulco, M.; Messa, C.; Caruso, M.G.; Notarnicola, M.; Tutino, V. Anti proliferative and pro apoptotic effects of flavonoid quercetin are mediated by CB1 receptor in human colon cancer cell lines. J. Cell. Physiol. 2015, 230, 2973–2980. [Google Scholar] [CrossRef]
- Akwu, N.A.; Naidoo, Y.; Channangihalli, S.T.; Singh, M.; Nundkumar, N.; Lin, J. The essential oils of Grewia Lasiocarpa E. Mey. Ex Harv.: Chemical composition, in vitro biological activity and cytotoxic effect on Hela cells. An. Acad. Bras. Cienc. 2021, 93, 93. [Google Scholar] [CrossRef]
- Hussein, H.A.; Mohamad, H.; Ghazaly, M.M.; Laith, A.; Abdullah, M.A. Anticancer and antioxidant activities of Nannochloropsis oculata and Chlorella sp. extracts in co-application with silver nanoparticle. J. King Saud Univ. Sci. 2020, 32, 3486–3494. [Google Scholar] [CrossRef]
- Yusuf, M.; Atthamid, N.F.U.; Indriati, S.; Saleh, R.; Latief, M.; Rifai, A. Optimization ultrasonic assisted extraction (UAE) of bioactive compound and antibacterial potential from sea urchin (Diadema setosum). Curr. Res. Nutr. Food Sci. 2020, 8, 556–569. [Google Scholar] [CrossRef]
- Kazemi, M. Phenolic profile, antioxidant capacity and anti-inflammatory activity of Anethum graveolens L. essential oil. Nat. Prod. Res. 2015, 29, 551–553. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Kang, J.; Fu, C. The independence of and associations among apoptosis, autophagy, and necrosis. Signal Transduct. Target. Ther. 2018, 3, 1–11. [Google Scholar] [CrossRef]
- Phung, H.M.; Lee, H.; Lee, S.; Jang, D.; Kim, C.-E.; Kang, K.S.; Seo, C.-S.; Choi, Y.-K. Analysis and anticancer effects of active compounds from Spatholobi caulis in human breast cancer cells. Processes 2020, 8, 1193. [Google Scholar] [CrossRef]
- Walensky, L.D. Targeting BAX to drug death directly. Nat. Chem. Biol. 2019, 15, 657–665. [Google Scholar] [CrossRef]
- Seervi, M.; Rani, A.; Sharma, A.K.; Santhosh Kumar, T.R. ROS mediated ER stress induces Bax-Bak dependent and independent apoptosis in response to Thioridazine. Biomed. Pharmacother. 2018, 106, 200–209. [Google Scholar] [CrossRef]
- Orning, P.; Lien, E. Multiple roles of caspase-8 in cell death, inflammation, and innate immunity. J. Leukoc. Biol. 2021, 109, 121–141. [Google Scholar] [CrossRef]
- Naumova, N.; Sachl, R. Regulation of cell death by mitochondrial transport systems of calcium and Bcl-2 proteins. Membranes 2020, 10, 299. [Google Scholar] [CrossRef]
- Warren, C.F.A.; Wong-Brown, M.W.; Bowden, N.A. BCL-2 family isoforms in apoptosis and cancer. Cell Death Dis. 2019, 10, 177. [Google Scholar] [CrossRef]
- Uehara, I.; Tanaka, N. Role of p53 in the regulation of the inflammatory tumor microenvironment and tumor suppression. Cancers 2018, 10, 219. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Qi, L.; Li, L.; Li, Y. The caspase-3/GSDME signal pathway as a switch between apoptosis and pyroptosis in cancer. Cell Death Discov. 2020, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- McGlorthan, L.; Paucarmayta, A.; Casablanca, Y.; Maxwell, G.L.; Syed, V. Progesterone induces apoptosis by activation of caspase-8 and calcitriol via activation of caspase-9 pathways in ovarian and endometrial cancer cells in vitro. Apoptosis 2021, 26, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.S.; Yang, E.J.; Lee, S.G.; Youn, U.J.; Han, S.J.; Kim, I.C.; Kim, S. Bioactivities of ethanol extract from the Antarctic freshwater microalga, Chloromonas sp. Int. J. Med. Sci. 2017, 14, 560–569. [Google Scholar] [CrossRef]
- Aubrey, B.J.; Kelly, G.L.; Janic, A.; Herold, M.J.; Strasser, A. How does p53 induce apoptosis and how does this relate to p53-mediated tumour suppression? Cell Death Differ. 2018, 25, 104–113. [Google Scholar] [CrossRef]
- Roy, S.; Sil, A.; Chakraborty, T. Potentiating apoptosis and modulation of p53, Bcl2, and Bax by a novel chrysin ruthenium complex for effective chemotherapeutic efficacy against breast cancer. J. Cell Physiol. 2019, 234, 4888–4909. [Google Scholar] [CrossRef]
- Jara-Gutierrez, A.; Baladron, V. The role of prostaglandins in different types of cancer. Cells 2021, 10, 1487. [Google Scholar] [CrossRef]
- Ramírez, D.; Caballero, J. Is it reliable to use common molecular docking methods for comparing the binding affinities of enantiomer pairs for their protein target? Int. J. Mol. Sci. 2016, 17, 525. [Google Scholar] [CrossRef]
- Zhang, H.; Ahima, J.; Yang, Q.; Zhao, L.; Zhang, X.; Zheng, X. A review on citrinin: Its occurrence, risk implications, analytical techniques, biosynthesis, physiochemical properties and control. Food Res. Int. 2021, 141, 16. [Google Scholar] [CrossRef]
- Gayathri, L.; Karthikeyan, B.S.; Rajalakshmi, M.; Dhanasekaran, D.; Li, A.P.; Akbarsha, M.A. Metabolism-dependent cytotoxicity of citrinin and ochratoxin A alone and in combination as assessed adopting integrated discrete multiple organ co-culture (IdMOC). Toxicol. In Vitro 2018, 46, 166–177. [Google Scholar] [CrossRef]
- Hashiesh, H.M.; Sharma, C.; Goyal, S.N.; Sadek, B.; Jha, N.K.; Kaabi, J.A.; Ojha, S. A focused review on CB2 receptor-selective pharmacological properties and therapeutic potential of beta-caryophyllene, a dietary cannabinoid. Biomed. Pharmacother. 2021, 140, 24. [Google Scholar] [CrossRef] [PubMed]
- Wojtukiewicz, M.Z.; Hempel, D.; Sierko, E.; Tucker, S.C.; Honn, K.V. Endothelial protein C receptor (EPCR), protease activated receptor-1 (PAR-1) and their interplay in cancer growth and metastatic dissemination. Cancers 2019, 11, 51. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Filho, J.W.G.; Dos Santos, T.d.J.A.; Tí, R.M.; Dos Reis, A.C.; Hameed, A.; de Oliveira Santos, J.V.; Afzal, M.I.; De Menezes, A.-A.P.M.; De Alencar, M.V.O.B.; Silva, D.H.S. Antitumor effects of citrinin in an animal model of sarcoma 180 via cytogenetic mechanisms. Cell. Mol. Biol. 2020, 66, 120–126. [Google Scholar] [CrossRef]
- Kadri, H.S.; Minocheherhomji, F.P. ADMET analysis of phyto-components of Syzygium cumini seeds and Allium cepa peels. Future J. Pharm. Sci. 2020, 6, 1–9. [Google Scholar] [CrossRef]
- Americus, B.; Lotan, T.; Bartholomew, J.L.; Atkinson, S.D. A comparison of the structure and function of nematocysts in free-living and parasitic cnidarians (Myxozoa). Int. J. Parasitol. 2020, 50, 763–769. [Google Scholar] [CrossRef] [PubMed]
- Lewis Ames, C. Medusa: A review of an ancient cnidarian body form. Results Probl. Cell Differ. 2018, 65, 105–136. [Google Scholar] [CrossRef] [PubMed]
- Ames, C.L.; Klompen, A.M.L.; Badhiwala, K.; Muffett, K.; Reft, A.J.; Kumar, M.; Janssen, J.D.; Schultzhaus, J.N.; Field, L.D.; Muroski, M.E.; et al. Cassiosomes are stinging-cell structures in the mucus of the upside-down jellyfish Cassiopea xamachana. Commun. Biol. 2020, 3, 67. [Google Scholar] [CrossRef]
- Miranda, L.S.; Collins, A.G.; Hirano, Y.M.; Mills, C.E.; Marques, A.C. Comparative internal anatomy of Staurozoa (Cnidaria), with functional and evolutionary inferences. PeerJ 2016, 4, 83. [Google Scholar] [CrossRef]
- Choudhary, I.; Lee, H.; Pyo, M.J.; Heo, Y.; Chae, J.; Yum, S.S.; Kang, C.; Kim, E. Proteomic investigation to identify anticancer targets of Nemopilema nomurai jellyfish venom in human hepatocarcinoma HepG2 Cells. Toxins 2018, 10, 194. [Google Scholar] [CrossRef]
- Arandian, A.; Mokari, M.; Zibaii, M.I.; Latifi, H. Optimization of Airy beam profile to improve light-sheet microscopy illumination. In Optical Technologies for Biology and Medicine; SPIE: Bellingham, WA, USA, 2022; pp. 141–148. [Google Scholar]
- Seymour, J.; Carrette, T.; Cullen, P.; Little, M.; Mulcahy, R.F.; Pereira, P.L. The use of pressure immobilization bandages in the first aid management of cubozoan envenomings. Toxicon 2002, 40, 1503–1505. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Sari, G.; Ozdal, T.; Capanoglu, E. Guidelines for cell viability assays. Food Front. 2020, 1, 332–349. [Google Scholar] [CrossRef]
- Zhang, J.-y.; Tao, L.-y.; Liang, Y.-j.; Chen, L.-m.; Mi, Y.-j.; Zheng, L.-s.; Wang, F.; She, Z.-g.; Lin, Y.-c.; To, K.K.W. Anthracenedione derivatives as anticancer agents isolated from secondary metabolites of the mangrove endophytic fungi. Mar. Drugs 2010, 8, 1469–1481. [Google Scholar] [CrossRef]
- Sathyaprabha, G.; Panneerselvam, A.; Kumaravel, S. Bioactive compounds identification of Pleurotus platypus and Pleurotus eous by GC-MS. Adv. Appl. Sci. Res. 2011, 2, 51–54. [Google Scholar]
- Bekele, D.; Tekie, H.; Asfaw, Z.; Petros, B. Bioactive chemical constituents from the leaf of Oreosyce africana Hook. f (Cucurbitaceae) with mosquitocidal activities against adult Anopheles arabiensis, the principal malaria vector in Ethiopia. J. fertil. pestic. 2016, 7. [Google Scholar] [CrossRef]
- Malik, S.K.; Ahmed, M.; Khan, F. Identification of novel anticancer terpenoids from Prosopis juliflora (Sw) DC (Leguminosae) pods. Trop. J. Pharm. Res. 2018, 17, 661–668. [Google Scholar] [CrossRef]
- Cui, Y.; Zhang, X.; Yin, K.; Qi, X.; Zhang, Y.; Zhang, J.; Li, S.; Lin, H. Dibutyl phthalate-induced oxidative stress, inflammation and apoptosis in grass carp hepatocytes and the therapeutic use of taxifolin. Sci. Total Environ. 2021, 764, 10. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Fu, C.; Zhou, F.; Luo, X.; Li, J.; Zhao, J.; He, J.; Li, X.; Li, J. Chemical composition, antioxidant and antitumor activities of sub-fractions of wild and cultivated Pleurotus ferulae ethanol extracts. PeerJ 2018, 6, e6097. [Google Scholar] [CrossRef] [PubMed]
- Sutanto, H.; Ainny, L.; Susanto, B.H.; Nasikin, M. Reaction product of pyrogallol with methyl linoleate and its antioxidant potential for biodiesel. In Proceedings of the IOP Conference Series: Materials Science and Engineering; 2018; p. 012019. [Google Scholar]
- Batalha, M.M.C.; Goulart, H.F.; Santana, A.E.G.; Barbosa, L.A.O.; Nascimento, T.G.; da Silva, M.K.H.; Dornelas, C.B.; Grillo, L.A.M. Chemical composition and antimicrobial activity of cuticular and internal lipids of the insect Rhynchophorus palmarum. Arch. Insect. Biochem. Physiol. 2020, 105, e21723. [Google Scholar] [CrossRef] [PubMed]
- El-anssary, A.A.; Raoof, G.F.A.; Saleh, D.O.; El-Masry, H.M. Bioactivities, physicochemical parameters and GC/MS profiling of the fixed oil of Cucumis melo L seeds: A focus on anti-inflammatory, immunomodulatory, and antimicrobial activities. J. HerbMed Pharmacol. 2021, 10, 476–485. [Google Scholar] [CrossRef]
- Marie Elvire, N.; Hortense, G.; Emmanuel, N.; Rose, N.; Francine, M. Evaluation of the in vitro antibacterial activity of Azadirachta indica used for the treat-ment of alveolitis. Int. J. Biopharm. Sci. 2020, 2, 119. [Google Scholar] [CrossRef]
- Dadwal, V.; Agrawal, H.; Sonkhla, K.; Joshi, R.; Gupta, M. Characterization of phenolics, amino acids, fatty acids and antioxidant activity in pulp and seeds of high altitude Himalayan crab apple fruits (Malus baccata). J. Food Sci. Technol. 2018, 55, 2160–2169. [Google Scholar] [CrossRef] [PubMed]
- Khiralla, A.; Spina, R.; Varbanov, M.; Philippot, S.; Lemiere, P.; Slezack-Deschaumes, S.; Andre, P.; Mohamed, I.; Yagi, S.M.; Laurain-Mattar, D. Evaluation of antiviral, antibacterial and antiproliferative activities of the endophytic fungus Curvularia papendorfii, and isolation of a new polyhydroxyacid. Microorganisms 2020, 8, 1353. [Google Scholar] [CrossRef] [PubMed]
- El Fakir, L.; Bouothmany, K.; Alotaibi, A.; Bourhia, M.; Ullah, R.; Zahoor, S.; El Mzibri, M.; Gmouh, S.; Alaoui, T.; Zaid, A. Antioxidant and understanding the anticancer properties in human prostate and breast cancer cell lines of chemically characterized methanol extract from Berberis hispanica Boiss. & Reut. Appl. Sci. 2021, 11, 3510. [Google Scholar] [CrossRef]
- Adenola, O.; Adeleke, S. Determination of bioactive compounds and antimicrobial capabilities of purified Nymphaea lotus Linn. (Nymphaeaceae) extract to multidrug resistant enteric bacteria. J. Altern. Complement. Med. 2021, 13, 39–53. [Google Scholar] [CrossRef]
- Burford, R.G.; Gowdey, C.W. Anti-inflammatory activity of alkoxyglycerols in rats. Arch. Int. Pharmacodyn. Ther. 1968, 173, 56–70. [Google Scholar] [PubMed]
- Ge, G.F.; Yu, C.H.; Yu, B.; Shen, Z.H.; Zhang, D.L.; Wu, Q.F. Antitumor effects and chemical compositions of Eupolyphaga sinensis walker ethanol extract. J. Ethnopharmacol. 2012, 141, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Dwivedi, P.D.; Dhawan, A.; Das, M.; Ansari, K.M. Citrinin-generated reactive oxygen species cause cell cycle arrest leading to apoptosis via the intrinsic mitochondrial pathway in mouse skin. Toxicol. Sci. 2011, 122, 557–566. [Google Scholar] [CrossRef]
- Heider, E.M.; Harper, J.K.; Grant, D.M.; Hoffman, A.; Dugan, F.; Tomer, D.P.; O'Neill, K.L. Exploring unusual antioxidant activity in a benzoic acid derivative: A proposed mechanism for citrinin. Tetrahedron 2006, 62, 1199–1208. [Google Scholar] [CrossRef]
- Sharath Babu, G.R.; Anand, T.; Ilaiyaraja, N.; Khanum, F.; Gopalan, N. Pelargonidin modulates Keap1/Nrf2 pathway gene expression and ameliorates citrinin-induced oxidative stress in HepG2 cells. Front. Pharmacol. 2017, 8, 868. [Google Scholar] [CrossRef]
- de Sousa Oliveira, K.; Queiroz, P.R.M.; Fensterseifer, I.C.M.; Migliolo, L.; Oliveira, A.L.; Franco, O.L. Purified citritin in combination with vancomycin inhibits VRE in vitro and in vivo. Microbiology 2017, 163, 1525–1531. [Google Scholar] [CrossRef]
- Shams ul Hassan, S.; Ishaq, M.; Zhang, W.-d.; Jin, H.-Z. An overview of the mechanisms of marine fungi-derived anti-inflammatory and anti-tumor agents and their novel role in drug targeting. Curr. Pharm. Des. 2021, 27, 2605–2614. [Google Scholar] [CrossRef] [PubMed]
- Vats, S.; Gupta, T. Evaluation of bioactive compounds and antioxidant potential of hydroethanolic extract of Moringa oleifera Lam. from Rajasthan, India. Physiol. Mol. Biol. Plants 2017, 23, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Saleh, E.I.M.M.; Bhattacharyya, P.; Van Staden, J. Chemical composition and cytotoxic activity of the essential oil and oleoresins of in vitro micropropagated Ansellia africana Lindl: A vulnerable medicinal orchid of Africa. Molecules 2021, 26, 4556. [Google Scholar] [CrossRef] [PubMed]
- Elmosallamy, A.; Ibraheem, E.; Sarhan, M.; Hussein, S.A.A. Chemical and biological activities of Deverra triradiata Hochst. ex. Boiss. Aerial parts from St. Catherine, southern Sinai, Egypt. Egypt. J. Chem. 2021, 64, 1387–1394. [Google Scholar] [CrossRef]
- Okechukwu, P. Evaluation of anti-inflammatory, analgesic, antipyretic effect of eicosane, pentadecane, octacosane, and heneicosane. Asian J. Pharm. Clin. Res. 2020, 13, 29–35. [Google Scholar] [CrossRef]
- Chathuranga, K.; Weerawardhana, A.; Dodantenna, N.; Ranathunga, L.; Cho, W.K.; Ma, J.Y.; Lee, J.S. Inhibitory effect of Sargassum fusiforme and its components on replication of respiratory syncytial virus in vitro and in vivo. Viruses 2021, 13, 548. [Google Scholar] [CrossRef]
- Zhao, C.; Gao, W.; Chen, T. Synergistic induction of apoptosis in A549 cells by dihydroartemisinin and gemcitabine. Apoptosis 2014, 19, 668–681. [Google Scholar] [CrossRef]
- Lu, J.; Chen, J.; Kang, Y.; Wu, J.; Shi, H.; Fu, Y.; Jiao, L.; Dong, C.; Li, X.; Jin, Y. Jinfukang induces cellular apoptosis through activation of Fas and DR4 in A549 cells. Oncol. Lett. 2018, 16, 4343–4352. [Google Scholar] [CrossRef]
- Dong, A.; Kong, M.; Ma, Z.; Qian, J.; Cheng, H.; Xu, X. Knockdown of insulin-like growth factor 1 receptor enhances chemosensitivity to cisplatin in human lung adenocarcinoma A549 cells. Acta Biochim. Biophys. Sin. 2008, 40, 497–504. [Google Scholar] [CrossRef]
- Mao, J.T.; Nie, W.-X.; Tsu, I.; Jin, Y.-S.; Rao, J.Y.; Lu, Q.-Y.; Zhang, Z.-F.; Go, V.L.W.; Serio, K.J. White tea extract induces apoptosis in non–small cell lung cancer cells: The role of peroxisome proliferator-activated receptor-γ and 15-lipoxygenaseswhite tea induces apoptosis in human lung cancer cells. Cancer Prev. Res. 2010, 3, 1132–1140. [Google Scholar] [CrossRef]
- Lin, M.; Tang, S.; Zhang, C.; Chen, H.; Huang, W.; Liu, Y.; Zhang, J. Euphorbia factor L2 induces apoptosis in A549 cells through the mitochondrial pathway. Acta Pharm. Sin. B 2017, 7, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Moghadamtousi, S.Z.; Kadir, H.A.; Paydar, M.; Rouhollahi, E.; Karimian, H. Annona muricata leaves induced apoptosis in A549 cells through mitochondrial-mediated pathway and involvement of NF-κB. BMC Complement. Atern. Med. 2014, 14, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Preet, A.; Qamri, Z.; Nasser, M.W.; Prasad, A.; Shilo, K.; Zou, X.; Groopman, J.E.; Ganju, R.K. Cannabinoid receptors, CB1 and CB2, as novel targets for inhibition of non–small cell lung cancer growth and metastasis cannabinoid receptors in NSCLC treatment. Cancer Prev. Res. 2011, 4, 65–75. [Google Scholar] [CrossRef]
- Hsu, H.-Y.; Lin, T.-Y.; Lu, M.-K.; Leng, P.-J.; Tsao, S.-M.; Wu, Y.-C. Fucoidan induces Toll-like receptor 4-regulated reactive oxygen species and promotes endoplasmic reticulum stress-mediated apoptosis in lung cancer. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Qiao, T.; Li, X.; Zhuang, X.; Chen, W.; Chen, X.; Zhang, Q. Toll-like receptor 9 activation by CpG oligodeoxynucleotide 7909 enhances the radiosensitivity of A549 lung cancer cells via the p53 signaling pathway. Oncol. Lett. 2018, 15, 5271–5279. [Google Scholar] [CrossRef] [PubMed]
- Antón, I.; Molina, E.; Luis-Ravelo, D.; Zandueta, C.; Valencia, K.; Ormazabal, C.; Martínez-Canarias, S.; Perurena, N.; Pajares, M.J.; Agorreta, J. Receptor of activated protein C promotes metastasis and correlates with clinical outcome in lung adenocarcinoma. Am. J. Respir. Crit. Care Med. 2012, 186, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Li, T.-J.; Huang, Y.-H.; Chen, X.; Zhou, Z.; Luo, S.-W.; Feng, D.-D.; Han, J.-Z.; Luo, Z.-Q. Metabotropic glutamate receptor 8 activation promotes the apoptosis of lung carcinoma A549 cells in vitro. Sheng Li Xue Bao 2015, 67, 513–520. [Google Scholar]
- Wang, J.J.; Mak, O.T. Induction of apoptosis in non-small cell lung carcinoma A549 cells by PGD2 metabolite, 15d-PGJ2. Cell Biol. Int. 2011, 35, 1089–1096. [Google Scholar] [CrossRef]
- Tarfiei, G.A.; Shadboorestan, A.; Montazeri, H.; Rahmanian, N.; Tavosi, G.; Ghahremani, M.H. GDF15 induced apoptosis and cytotoxicity in A549 cells depends on TGFBR2 expression. Cell Biochem. Funct. 2019, 37, 320–330. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Jellyfish | No. | Compounds | Formula | MW | RT | Abundance Ratio (%) * | |
|---|---|---|---|---|---|---|---|
| CA | CM | ||||||
| Cassiopea andromeda/Catostylus mosaicus | 1 | Dibutyl phthalate | C16H22O4 | 278.34 | 18.41 | 3.153 | 0.746 |
| 2 | 9,12-Octadecadienoic acid, methyl ester | C19H34O2 | 294.5 | 21.86 | 4.852 | 5.552 | |
| 3 | Heptadecane,2,6,10,14-Tetramethyl- | C21H44 | 296.6 | 24.4 | 0.775 | 5.221 | |
| 4 | Batilol | C21H44O3 | 344.6 | 30.14 | 0.506 | 1.397 | |
| Cassiopea andromeda | 5 | Ethaneperoxoic acid, 1-cyano-1-[2-(2-phenyl-1,3-dioxolan-2-yl)ethyl]pentyl ester | C19H25NO5 | 347.4 | 15.94 | 1.098 | NA |
| Catostylus mosaicus | 6 | Citrinin | C13H14O5 | 250.25 | 17.33 | NA | 4.402 |
| 7 | Eicosane | C20H42 | 282.5 | 19.29 | NA | 1.269 | |
| Ligands | Receptors | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Caspase-3 | Caspase-7 | Caspase-9 | CB1 | CB2 | DR4 | EPCR | Fas Receptor | IGF1R | mGluRs | PPAR-γ | TGFBR2 | TLR4 | TLR9 | TNFR1 | PGD2 | Caspase-8 | |
| 9,12-Octadecadienoic acid, methyl ester | −5.2 | −5.3 | −4.6 | −6.7 | −7.3 | −3.5 | −7.6 | −3.8 | −5.6 | −4.9 | −5.3 | −4.9 | −4.4 | −5.5 | −3.9 | −7 | −5.5 |
| Batilol | −5.4 | −5.6 | −3.7 | −6.3 | −4.3 | −3.2 | −7.3 | −3.9 | −4.6 | −5.2 | −5.6 | −4.2 | −4.4 | −4.8 | −3.9 | −6.6 | −5.6 |
| Citrinin | −6.8 | −6.4 | −5.3 | −7.8 | −7.8 | −6 | −8.1 | −6.5 | −6.4 | −5.9 | −7.6 | −5.6 | −6.6 | −6.8 | −5.8 | −8.5 | −6.1 |
| Dibutyl phthalate | −5.5 | −5.5 | −4.3 | −7.6 | −5.1 | −4.4 | −7.7 | −5.1 | −5.8 | −5.5 | −6.2 | −4.8 | −5.1 | −5.2 | −4.6 | −7.8 | −6 |
| Eicosane | −4.6 | −4.4 | −3.5 | −6.8 | −4.4 | −3 | −7.4 | −3.2 | −4.6 | −3.7 | −5.2 | −4.4 | −4.3 | −4.3 | −3.3 | −6.6 | −4.8 |
| Ethaneperoxoic acid, 1-cyano-1-[2-(2-phenyl-1,3-dioxolan-2-yl)ethyl]pentyl ester | −6.7 | −6.6 | −4.4 | −6.9 | −8.1 | −5.6 | −8.1 | −5.4 | −6.4 | −6.2 | −7.5 | −5.3 | −5.8 | −6.5 | −5.4 | −8.4 | −6.8 |
| Heptadecane,2,6,10,14-Tetramethyl- | −4.5 | −5.1 | −3.7 | −7.3 | −4.9 | −3.4 | −8 | −4 | −5.6 | −4.3 | −6.1 | −4.7 | −4.6 | −5.7 | −4.1 | −7.1 | −5.5 |
| Receptors | Avg | SD | Min | Max |
|---|---|---|---|---|
| caspase-3 | 0.530 | 0.077 | 0.422 | 0.638 |
| caspase-7 | 0.621 | 0.102 | 0.453 | 0.768 |
| caspase-8 | 0.633 | 0.058 | 0.551 | 0.718 |
| caspase-9 | 0.821 | 0.081 | 0.702 | 0.938 |
| CB1 | 0.464 | 0.062 | 0.367 | 0.549 |
| CB2 | 0.461 | 0.070 | 0.356 | 0.558 |
| DR4 | 0.685 | 0.061 | 0.604 | 0.769 |
| EPCR | 0.451 | 0.062 | 0.364 | 0.535 |
| Fas Receptor | 0.601 | 0.104 | 0.464 | 0.744 |
| IGF1R | 0.628 | 0.062 | 0.524 | 0.738 |
| mGluRs | 0.739 | 0.070 | 0.636 | 0.838 |
| PGD2 | 0.432 | 0.060 | 0.312 | 0.538 |
| PPAR-γ | 0.482 | 0.062 | 0.388 | 0.572 |
| TGFBR2 | 0.795 | 0.079 | 0.679 | 0.914 |
| TLR4 | 0.555 | 0.070 | 0.454 | 0.654 |
| TLR9 | 0.540 | 0.60 | 0.454 | 0.647 |
| TNFR1 | 0.729 | 0.90 | 0.614 | 0.838 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zare, A.; Afshar, A.; Khoradmehr, A.; Baghban, N.; Mohebbi, G.; Barmak, A.; Daneshi, A.; Bargahi, A.; Nabipour, I.; Almasi-Turk, S.; et al. Chemical Compositions and Experimental and Computational Modeling of the Anticancer Effects of Cnidocyte Venoms of Jellyfish Cassiopea andromeda and Catostylus mosaicus on Human Adenocarcinoma A549 Cells. Mar. Drugs 2023, 21, 168. https://doi.org/10.3390/md21030168
Zare A, Afshar A, Khoradmehr A, Baghban N, Mohebbi G, Barmak A, Daneshi A, Bargahi A, Nabipour I, Almasi-Turk S, et al. Chemical Compositions and Experimental and Computational Modeling of the Anticancer Effects of Cnidocyte Venoms of Jellyfish Cassiopea andromeda and Catostylus mosaicus on Human Adenocarcinoma A549 Cells. Marine Drugs. 2023; 21(3):168. https://doi.org/10.3390/md21030168
Chicago/Turabian StyleZare, Afshin, Alireza Afshar, Arezoo Khoradmehr, Neda Baghban, Gholamhossein Mohebbi, Alireza Barmak, Adel Daneshi, Afshar Bargahi, Iraj Nabipour, Sahar Almasi-Turk, and et al. 2023. "Chemical Compositions and Experimental and Computational Modeling of the Anticancer Effects of Cnidocyte Venoms of Jellyfish Cassiopea andromeda and Catostylus mosaicus on Human Adenocarcinoma A549 Cells" Marine Drugs 21, no. 3: 168. https://doi.org/10.3390/md21030168
APA StyleZare, A., Afshar, A., Khoradmehr, A., Baghban, N., Mohebbi, G., Barmak, A., Daneshi, A., Bargahi, A., Nabipour, I., Almasi-Turk, S., Arandian, A., Zibaii, M. I., Latifi, H., & Tamadon, A. (2023). Chemical Compositions and Experimental and Computational Modeling of the Anticancer Effects of Cnidocyte Venoms of Jellyfish Cassiopea andromeda and Catostylus mosaicus on Human Adenocarcinoma A549 Cells. Marine Drugs, 21(3), 168. https://doi.org/10.3390/md21030168