
Functional Characterization, Antimicrobial Effects, and Potential Antibacterial Mechanisms of NpHM4, a Derived Peptide of Nautilus pompilius Hemocyanin

 ,
, 
Abstract
1. Introduction
2. Results
2.1. In Silico Predicted Peptides with Antimicrobial Activity
2.2. Antibacterial Activity of the Six Predicted Antimicrobial Peptides (NpHN1–NpHN6)
2.3. Sequence Analysis of Mutant AMPs Derived from NpHN5
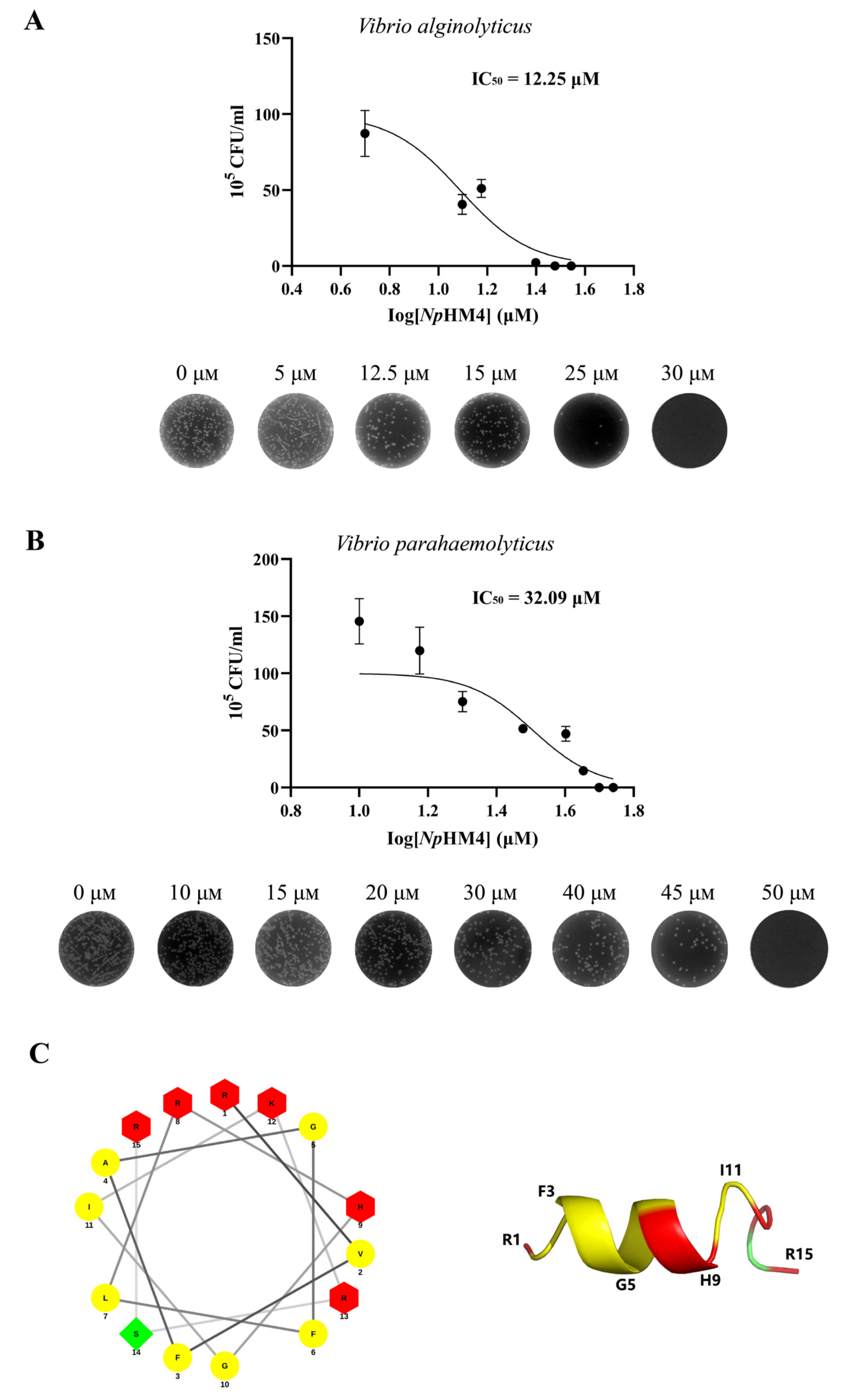
2.4. Bactericidal Activity of Seven Mutant AMPs
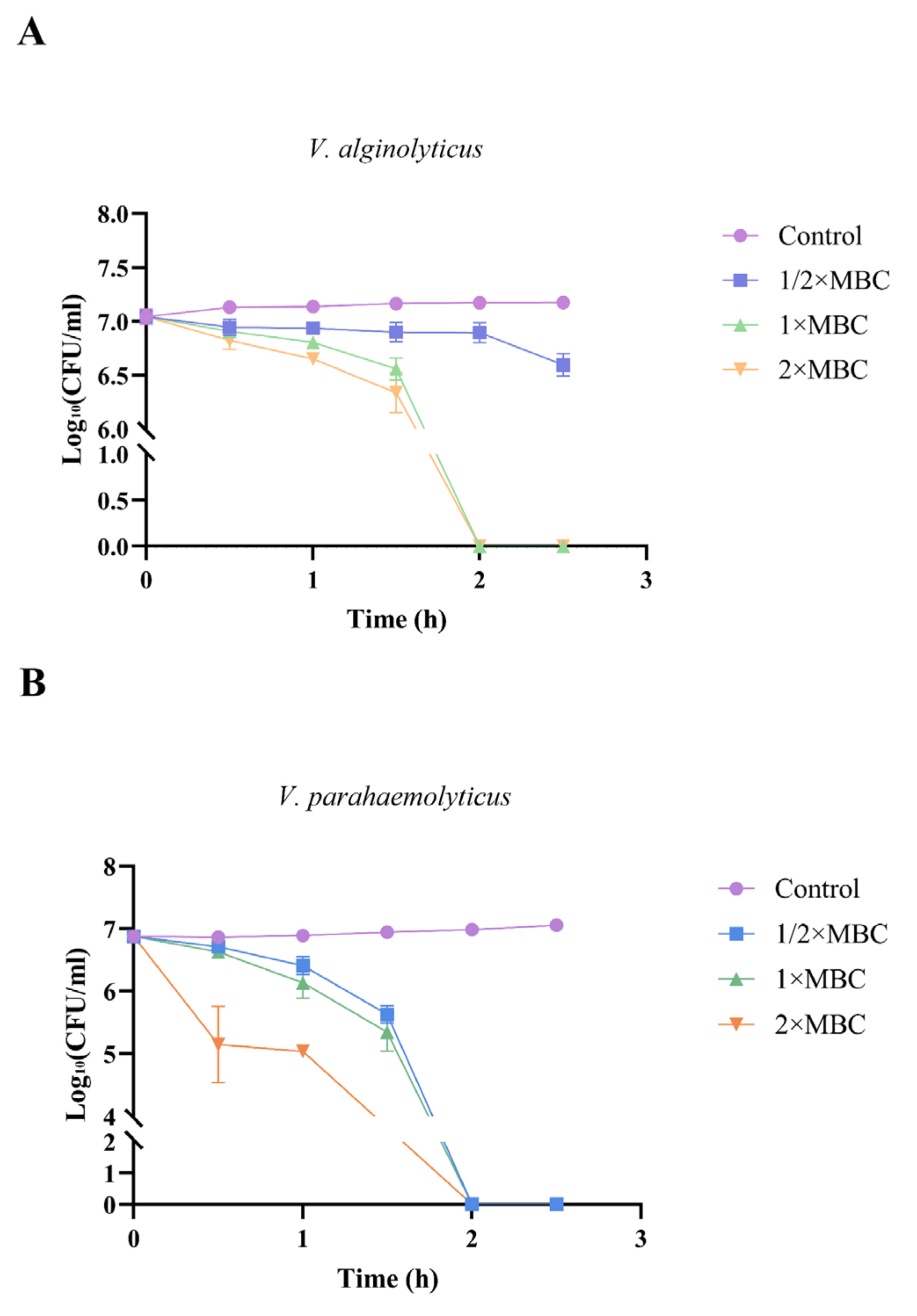
2.5. Time-Course Bactericidal Activity of NpHM4 towards V. alginolyticus and V. parahaemolyticus
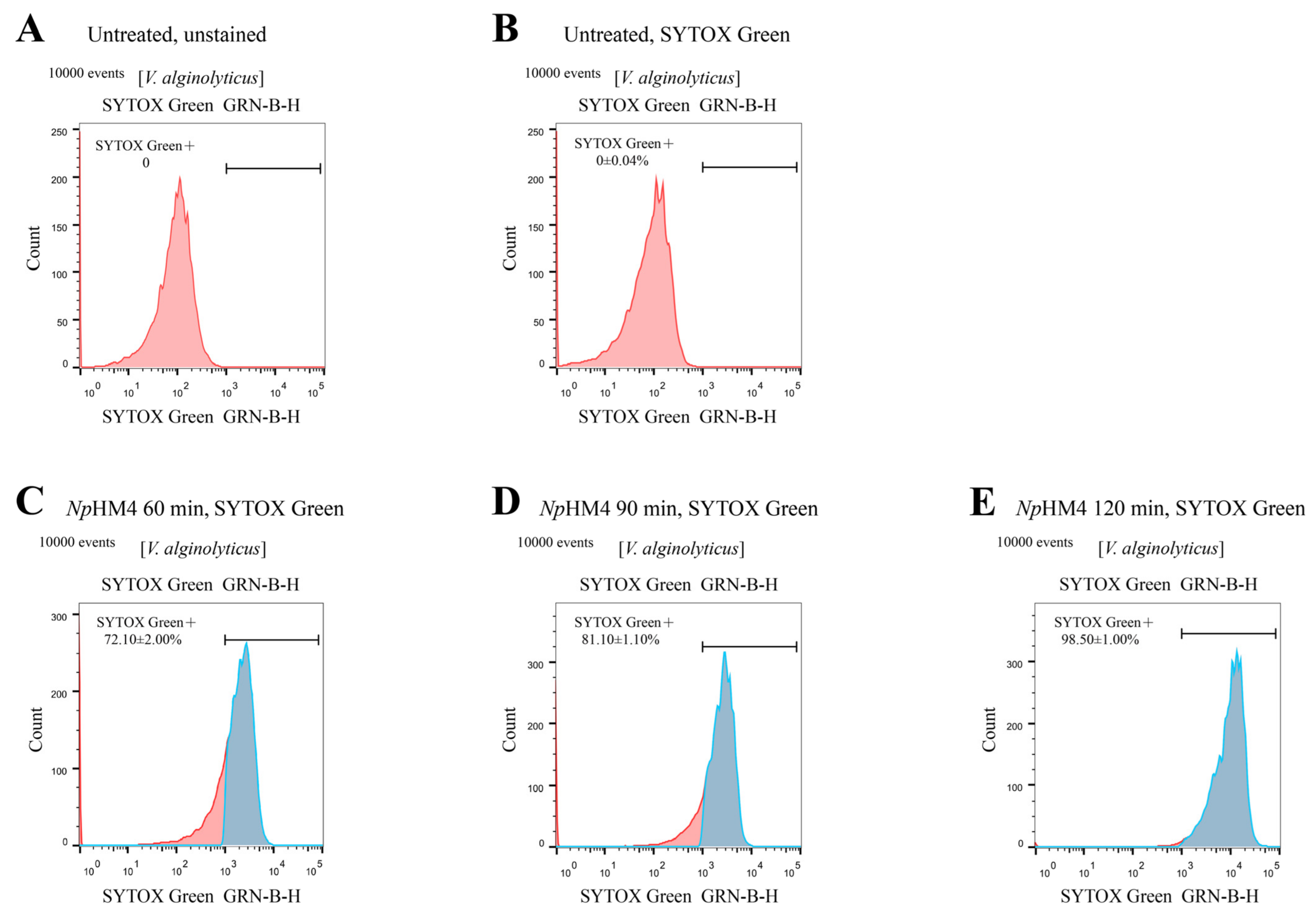
2.6. Membrane-Penetrating Activity of NpHM4 toward V. alginolyticus
2.7. Effect of Peptide NpHM4 on V. alginolyticus Ultrastructure via Scanning Electron Microscopy
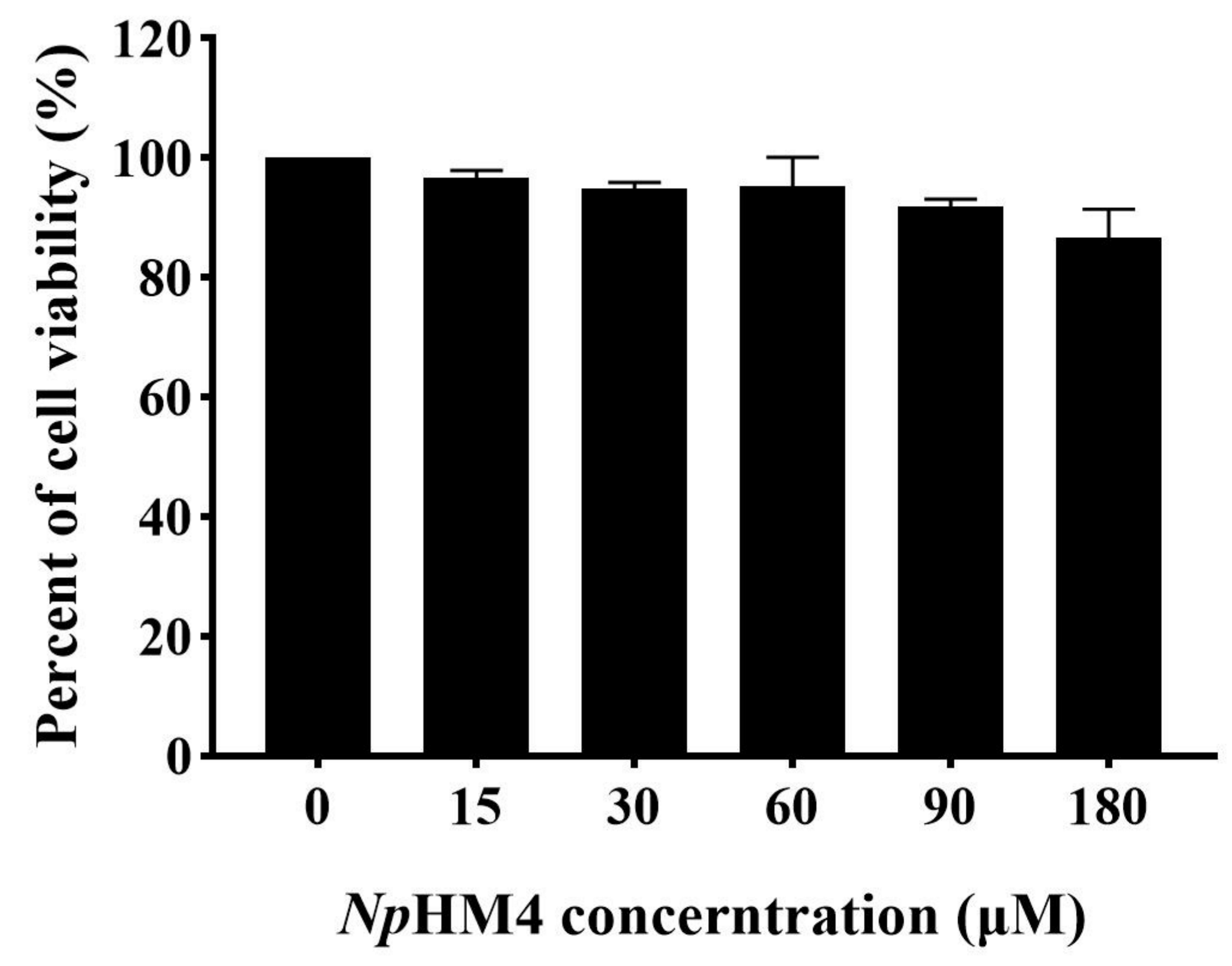
2.8. Cytotoxicity of NpHM4 to HEK293 Cells
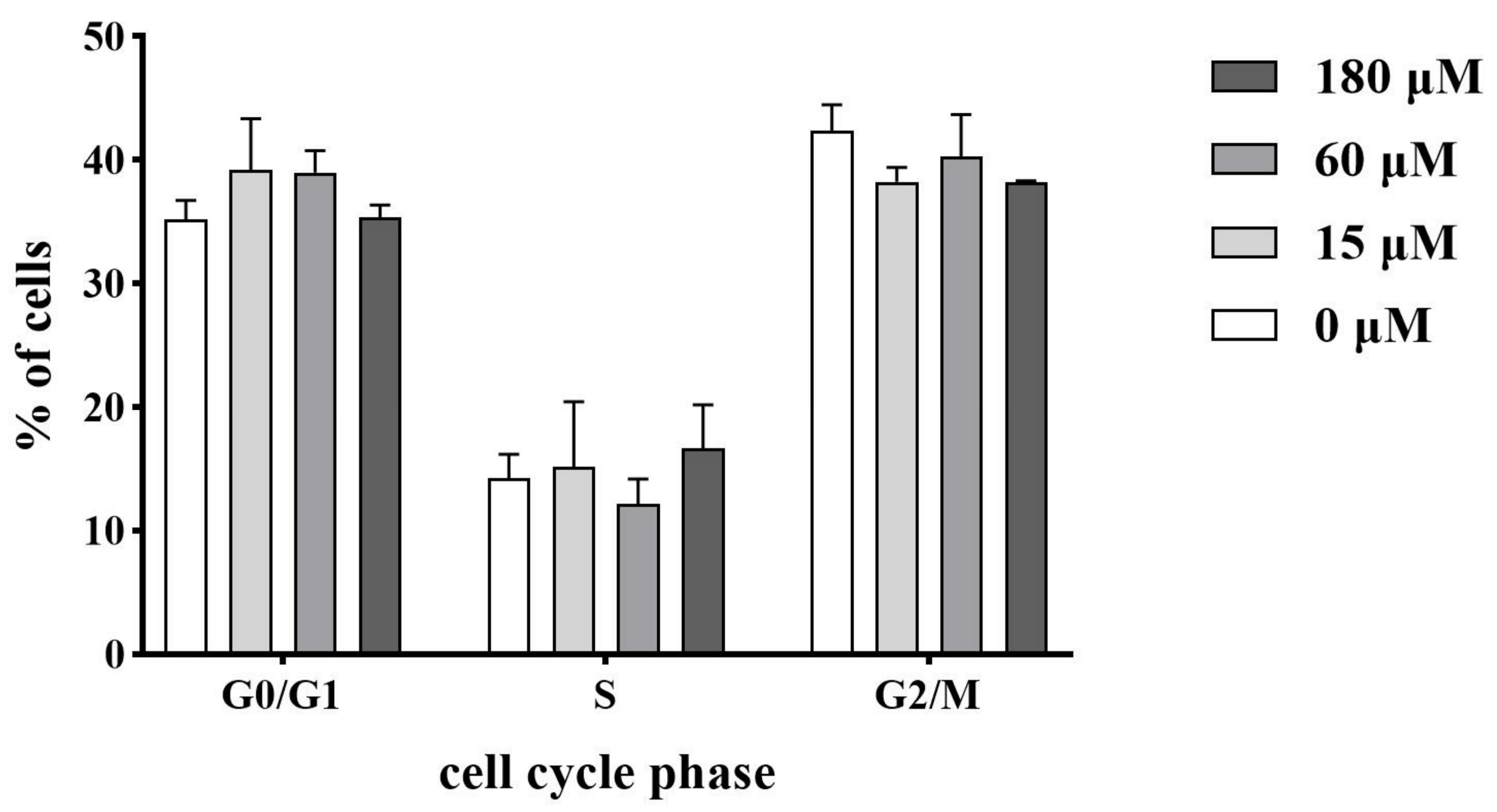
2.9. Influence on the Cell Cycle Progression
3. Discussion
4. Materials and Methods
4.1. Prediction of AMPs from N. pompilius Hemocyanin
4.2. Mutant Peptide Design on the Basis of NpHN5
4.3. Peptide Synthesis
4.4. Antimicrobial Assay
4.5. Time-Course Bactericidal Activity of NpHM4
4.6. Membrane Permeability
4.7. Analysis of Cellular Morphology via Scanning Electron Microscopy (SEM)
4.8. Cytotoxicity Assay
4.9. Cell Cycle Analysis via Flow Cytometry
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Rossolini, G.M.; Arena, F.; Pecile, P.; Pollini, S. Update on the antibiotic resistance crisis. Curr. Opin. Pharmacol. 2014, 18, 56–60. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance. Available online: https://www.who.int/publications/i/item/9789241564748 (accessed on 19 July 2021).
- Zhang, L.-J.; Gallo, R.L. Antimicrobial peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Huang, J.; Chen, Y. Alpha-helical cationic antimicrobial peptides: Relationships of structure and function. Protein Cell 2010, 1, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Waghu, F.H.; Joseph, S.; Ghawali, S.; Martis, E.A.; Madan, T.; Venkatesh, K.V.; Idicula-Thomas, S. Designing antibacterial peptides with enhanced killing kinetics. Front. Microbiol. 2018, 9, 325. [Google Scholar] [CrossRef]
- Murugan, R.N.; Jacob, B.; Ahn, M.; Hwang, E.; Sohn, H.; Park, H.N.; Lee, E.; Seo, J.H.; Cheong, C.; Nam, K.Y.; et al. De novo design and synthesis of ultra-short peptidomimetic antibiotics having dual antimicrobial and anti-inflammatory activities. PLoS ONE 2013, 8, e80025. [Google Scholar] [CrossRef]
- Li, J.; Koh, J.-J.; Liu, S.; Lakshminarayanan, R.; Verma, C.S.; Beuerman, R.W. Membrane active antimicrobial peptides: Translating mechanistic insights to design. Front. Neurosci. 2017, 11, 73. [Google Scholar] [CrossRef]
- Gianazza, E.; Eberini, I.; Palazzolo, L.; Miller, I. Hemolymph proteins: An overview across marine arthropods and molluscs. J. Proteom. 2021, 245, 104294. [Google Scholar] [CrossRef]
- Dolashka, P.; Velkova, L.; Shishkov, S.; Kostova, K.; Dolashki, A.; Dimitrov, I.; Atanasov, B.; Devreese, B.; Voelter, W.; van Beeumen, J. Glycan structures and antiviral effect of the structural subunit rvh2 of rapana hemocyanin. Carbohydr. Res. 2010, 345, 2361–2367. [Google Scholar] [CrossRef]
- Qin, Z.; Babu, V.S.; Wan, Q.; Muhammad, A.; Li, J.; Lan, J.; Lin, L. Antibacterial activity of hemocyanin from red swamp crayfish (Procambarus clarkii). Fish Shellfish Immunol. 2018, 75, 391–399. [Google Scholar] [CrossRef]
- Coates, C.J.; Nairn, J. Diverse immune functions of hemocyanins. Dev. Comp. Immunol. 2014, 45, 43–55. [Google Scholar] [CrossRef]
- Monteiro, M.L.; Lima, D.B.; Menezes, R.R.P.P.B.d.; Sampaio, T.L.; Silva, B.P.; Serra Nunes, J.V.; Cavalcanti, M.M.; Morlighem, J.-E.; Martins, A.M.C. Antichagasic effect of hemocyanin derived from antimicrobial peptides of Penaeus monodon shrimp. Exp. Parasitol. 2020, 215, 107930. [Google Scholar] [CrossRef]
- Zhuang, J.; Coates, C.J.; Zhu, H.; Zhu, P.; Wu, Z.; Xie, L. Identification of candidate antimicrobial peptides derived from abalone hemocyanin. Dev. Comp. Immunol. 2015, 49, 96–102. [Google Scholar] [CrossRef]
- Fisher, W.S.; DiNuzzo, A.R. Agglutination of bacteria and erythrocytes by serum from six species of marine molluscs. J. Invertebr. Pathol. 1991, 57, 380–394. [Google Scholar] [CrossRef]
- McFall-Ngai, M. Care for the community. Nature 2007, 445, 153. [Google Scholar] [CrossRef]
- Gestal, C.; Castellanos-Martínez, S. Understanding the cephalopod immune system based on functional and molecular evidence. Fish Shellfish Immunol. 2015, 46, 120–130. [Google Scholar] [CrossRef]
- Ford, L.A. Host defense mechanisms of cephalopods. Annu. Rev. Fish Dis. 1992, 2, 25–41. [Google Scholar] [CrossRef]
- Houyvet, B.; Zanuttini, B.; Corre, E.; Le Corguillé, G.; Henry, J.; Zatylny-Gaudin, C. Design of antimicrobial peptides from a cuttlefish database. Amino Acids 2018, 50, 1573–1582. [Google Scholar] [CrossRef]
- Nikapitiya, C.; Dananjaya, S.H.S.; Chandrarathna, H.P.S.U.; de Zoysa, M.; Whang, I. Octominin: A novel synthetic anticandidal peptide derived from defense protein of Octopus minor. Mar. Drugs 2020, 18, 56. [Google Scholar] [CrossRef]
- Kröger, B.; Vinther, J.; Fuchs, D. Cephalopod origin and evolution: A congruent picture emerging from fossils, development and molecules: Extant cephalopods are younger than previously realised and were under major selection to become agile, shell-less predators. Bioessays 2011, 33, 602–613. [Google Scholar] [CrossRef]
- Vandepas, L.E.; Dooley, F.D.; Barord, G.J.; Swalla, B.J.; Ward, P.D. A revisited phylogeography of Nautilus pompilius. Ecol. Evol. 2016, 6, 4924–4935. [Google Scholar] [CrossRef]
- Zhang, Y.; Mao, F.; Mu, H.; Huang, M.; Bao, Y.; Wang, L.; Wong, N.-K.; Xiao, S.; Dai, H.; Xiang, Z.; et al. The genome of Nautilus pompilius illuminates eye evolution and biomineralization. Nat. Ecol. Evol. 2021, 5, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Gatsogiannis, C.; Moeller, A.; Depoix, F.; Meissner, U.; Markl, J. Nautilus pompilius hemocyanin: 9 Å cryo-em structure and molecular model reveal the subunit pathway and the interfaces between the 70 functional units. J. Mol. Biol. 2007, 374, 465–486. [Google Scholar] [CrossRef] [PubMed]
- Klubthawee, N.; Adisakwattana, P.; Hanpithakpong, W.; Somsri, S.; Aunpad, R. A novel, rationally designed, hybrid antimicrobial peptide, inspired by cathelicidin and aurein, exhibits membrane-active mechanisms against Pseudomonas aeruginosa. Sci. Rep. 2020, 10, 9117. [Google Scholar] [CrossRef] [PubMed]
- Ong, Z.Y.; Wiradharma, N.; Yang, Y.Y. Strategies employed in the design and optimization of synthetic antimicrobial peptide amphiphiles with enhanced therapeutic potentials. Adv. Drug Deliv. Rev. 2014, 78, 28–45. [Google Scholar] [CrossRef]
- Zeth, K.; Sancho-Vaello, E. The human antimicrobial peptides dermcidin and ll-37 show novel distinct pathways in membrane interactions. Front. Chem. 2017, 5, 86. [Google Scholar] [CrossRef]
- Takahashi, D.; Shukla, S.K.; Prakash, O.; Zhang, G. Structural determinants of host defense peptides for antimicrobial activity and target cell selectivity. Biochimie 2010, 92, 1236–1241. [Google Scholar] [CrossRef]
- Yang, R.; Zhang, G.; Zhang, F.; Li, Z.; Huang, C. Membrane permeabilization design of antimicrobial peptides based on Chikungunya virus fusion domain scaffold and its antibacterial activity against gram-positive Streptococcus pneumoniae in respiratory infection. Biochimie 2018, 146, 139–147. [Google Scholar] [CrossRef]
- Loth, K.; Vergnes, A.; Barreto, C.; Voisin, S.N.; Meudal, H.; Da Silva, J.; Bressan, A.; Belmadi, N.; Bachère, E.; Aucagne, V.; et al. The ancestral n-terminal domain of big defensins drives bacterially triggered assembly into antimicrobial nanonets. mBio 2019, 10, e01821-19. [Google Scholar] [CrossRef]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Le, C.-F.; Yusof, M.Y.M.; Hassan, H.; Sekaran, S.D. In vitro properties of designed antimicrobial peptides that exhibit potent antipneumococcal activity and produces synergism in combination with penicillin. Sci. Rep. 2015, 5, 9761. [Google Scholar] [CrossRef]
- Koehbach, J.; Craik, D.J. The vast structural diversity of antimicrobial peptides. Trends Pharmacol. Sci. 2019, 40, 517–528. [Google Scholar] [CrossRef]
- Decker, H.; Rimke, T. Tarantula hemocyanin shows phenoloxidase activity. J. Biol. Chem. 1998, 273, 25889–25892. [Google Scholar] [CrossRef]
- Zanjani, N.T.; Sairi, F.; Marshall, G.; Saksena, M.M.; Valtchev, P.; Gomes, V.G.; Cunningham, A.L.; Dehghani, F. Formulation of abalone hemocyanin with high antiviral activity and stability. Eur. J. Pharm. Sci. 2014, 53, 77–85. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, F.; Chen, C.; Zheng, Z.; Aweya, J.J.; Zhang, Y. Glycosylation of hemocyanin in Litopenaeus vannamei is an antibacterial response feature. Immunol. Lett. 2017, 192, 42–47. [Google Scholar] [CrossRef]
- Wen, Y.; Zhan, S.; Huang, H.; Zhong, M.; Chen, J.; You, C.; Wang, F.; Zhang, Y. Identification and characterization of an 18.4 kda antimicrobial truncation from shrimp Litopenaeus vannamei hemocyanin upon vibrio parahaemolyticus infection. Fish Shellfish Immunol. 2016, 56, 450–458. [Google Scholar] [CrossRef]
- Liscano, Y.; Salamanca, C.H.; Vargas, L.; Cantor, S.; Laverde-Rojas, V.; Oñate-Garzón, J. Increases in hydrophilicity and charge on the polar face of alyteserin 1c helix change its selectivity towards gram-positive bacteria. Antibiotics 2019, 8, 238. [Google Scholar] [CrossRef]
- Manzo, G.; Scorciapino, M.A.; Wadhwani, P.; Bürck, J.; Montaldo, N.P.; Pintus, M.; Sanna, R.; Casu, M.; Giuliani, A.; Pirri, G.; et al. Enhanced amphiphilic profile of a short β-stranded peptide improves its antimicrobial activity. PLoS ONE 2015, 10, e0116379. [Google Scholar] [CrossRef]
- Tan, P.; Fu, H.; Ma, X. Design, optimization, and nanotechnology of antimicrobial peptides: From exploration to applications. Nano Today 2021, 39, 101229. [Google Scholar] [CrossRef]
- Yin, L.M.; Edwards, M.A.; Li, J.; Yip, C.M.; Deber, C.M. Roles of hydrophobicity and charge distribution of cationic antimicrobial peptides in peptide-membrane interactions. J. Biol. Chem. 2012, 287, 7738–7745. [Google Scholar] [CrossRef]
- Mukherjee, S.; Chowdhury, P.; Gai, F. Infrared study of the effect of hydration on the amide i band and aggregation properties of helical peptides. J. Phys. Chem. B 2007, 111, 4596–4602. [Google Scholar] [CrossRef]
- Hollmann, A.; Martínez, M.; Noguera, M.E.; Augusto, M.T.; Disalvo, A.; Santos, N.C.; Semorile, L.; Maffía, P.C. Role of amphipathicity and hydrophobicity in the balance between hemolysis and peptide–membrane interactions of three related antimicrobial peptides. Colloids Surf. B Biointerfaces 2016, 141, 528–536. [Google Scholar] [CrossRef]
- Lee, T.H.; Hall, K.N.; Aguilar, M.I. Antimicrobial peptide structure and mechanism of action: A focus on the role of membrane structure. Curr. Top. Med. Chem. 2016, 16, 25–39. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial peptides: An emerging category of therapeutic agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef]
- Fjell, C.D.; Hiss, J.A.; Hancock, R.E.W.; Schneider, G. Designing antimicrobial peptides: Form follows function. Nat. Rev. Drug Discov. 2012, 11, 37–51. [Google Scholar] [CrossRef]
- Yang, S.; Huang, H.; Aweya, J.J.; Zheng, Z.; Liu, G.; Zhang, Y. Pvhs9 is a novel in silico predicted antimicrobial peptide derived from hemocyanin of Penaeus vannamei. Aquaculture 2021, 530, 735926. [Google Scholar] [CrossRef]
- Sun, Y.; Dong, W.; Sun, L.; Ma, L.; Shang, D. Insights into the membrane interaction mechanism and antibacterial properties of chensinin-1b. Biomaterials 2015, 37, 299–311. [Google Scholar] [CrossRef]
- Yang, S.; Li, J.; Aweya, J.J.; He, S.; Deng, S.; Weng, W.; Zhang, Y.; Liu, G.-M. Antimicrobial activity of pvh4a, a peptide derived from histone h4 of Penaeus vannamei. Aquaculture 2022, 549, 737807. [Google Scholar] [CrossRef]
- Kim, H.Y.; Kim, K.-T.; Kim, S.D. Biochemical effects of veterinary antibiotics on proliferation and cell cycle arrest of human hek293 cells. Bull. Environ. Contam. Toxicol. 2012, 89, 234–239. [Google Scholar] [CrossRef]
- Gu, Y.; Chen, X.; Shang, C.; Singh, K.; Barzegar, M.; Mahdavian, E.; Salvatore, B.A.; Jiang, S.; Huang, S. Fusarochromanone induces g1 cell cycle arrest and apoptosis in cos7 and hek293 cells. PLoS ONE 2014, 9, e112641. [Google Scholar] [CrossRef]
- Yang, S.; Huang, H.; Wang, F.; Aweya, J.J.; Zheng, Z.; Zhang, Y. Prediction and characterization of a novel hemocyanin-derived antimicrobial peptide from shrimp Litopenaeus vannamei. Amino Acids 2018, 50, 995–1005. [Google Scholar] [CrossRef]
- Waghu, F.H.; Gopi, L.; Barai, R.S.; Ramteke, P.; Nizami, B.; Idicula-Thomas, S. Camp: Collection of sequences and structures of antimicrobial peptides. Nucleic Acids Res. 2014, 42, D1154–D1158. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Karnik, S.; Barai, R.S.; Jayaraman, V.K.; Idicula-Thomas, S. Camp: A useful resource for research on antimicrobial peptides. Nucleic Acids Res. 2010, 38, D774–D780. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, X.; Wang, Z. Apd3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Tian, H.; Wang, T.; Wang, Z.; Chou, S.; Shan, A.; Cheng, B. Central β-turn increases the cell selectivity of imperfectly amphipathic α-helical peptides. Acta Biomater. 2018, 69, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Fauchere, J.; Pliska, V. Hydrophobic parameters ii of amino acid side-chains from the partitioning of n-acetyl-amino acid amides. Eur. J. Med. Chem. 1983, 18, 369–375. [Google Scholar]
- Zheng, W.; Zhang, C.; Bell, E.W.; Zhang, Y. I-tasser gateway: A protein structure and function prediction server powered by xsede. Future Gener. Comput. Syst. 2019, 99, 73–85. [Google Scholar] [CrossRef]
- Lv, Y.; Wang, J.; Gao, H.; Wang, Z.; Dong, N.; Ma, Q.; Shan, A. Antimicrobial properties and membrane-active mechanism of a potential α-helical antimicrobial derived from cathelicidin pmap-36. PLoS ONE 2014, 9, e86364. [Google Scholar] [CrossRef]
- Jindal, H.M.; Le, C.F.; Mohd Yusof, M.Y.; Velayuthan, R.D.; Lee, V.S.; Zain, S.M.; Isa, D.M.; Sekaran, S.D. Antimicrobial activity of novel synthetic peptides derived from indolicidin and ranalexin against Streptococcus pneumoniae. PLoS ONE 2015, 10, e0128532. [Google Scholar] [CrossRef]
- Mao, F.; Bao, Y.; Wong, N.K.; Huang, M.; Liu, K.; Zhang, X.; Yang, Z.; Yi, W.; Shu, X.; Xiang, Z.; et al. Large-scale plasma peptidomic profiling reveals a novel, nontoxic, Crassostrea hongkongensis-derived antimicrobial peptide against foodborne pathogens. Mar. Drugs 2021, 19, 420. [Google Scholar] [CrossRef]
- Cai, D.; Chen, S.; Wu, B.; Chen, J.; Tao, D.; Li, Z.; Dong, Q.; Zou, Y.; Chen, Y.; Bi, C.; et al. Construction of multifunctional porcine acellular dermal matrix hydrogel blended with vancomycin for hemorrhage control, antibacterial action, and tissue repair in infected trauma wounds. Mater. Today Bio 2021, 12, 100127. [Google Scholar] [CrossRef]
- Alja, Š.; Filipič, M.; Novak, M.; Žegura, B. Double strand breaks and cell-cycle arrest induced by the cyanobacterial toxin cylindrospermopsin in hepg2 cells. Mar. Drugs 2013, 11, 3077–3090. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptides | Gram-Negative Bacteria | |||
|---|---|---|---|---|
| Escherichia coli (DH5α) | Vibrio alginolyticus (A056) | |||
| MIC (μM) | MBC (μM) | MIC (μM) | MBC (μM) | |
| NpHN1 | 250–450 | >450 | 50–260 | >260 |
| NpHN2 | 200–500 | >500 | 100–280 | >280 |
| NpHN3 | 250–450 | >450 | 50–250 | >250 |
| NpHN4 | 200–600 | >600 | 100–300 | >300 |
| NpHN5 | 200–450 | 450 | 50–250 | 250 |
| NpHN6 | 250–500 | >500 | 100–300 | 300 |
| No. | Peptide Sequence | Pho% c | Net Charge | Number of Mutations | Measured MW a | Secondary Structure | uH b |
|---|---|---|---|---|---|---|---|
| NpHM1 | RVFAGFLRHGIKRSA | 46 | 4.25 | 1 | 1715.02 | α-helix | 0.423 |
| NpHM2 | RVFAGFLLHGIKRSR | 46 | 4.25 | 1 | 1757.10 | β-chain | 0.336 |
| NpHM3 | RVRAGFLLHGIKRSA | 46 | 4.25 | 1 | 1681.00 | β-chain | 0.181 |
| NpHM4 | RVFAGFLRHGIKRSR | 40 | 5.25 | 2 | 1800.13 | α-helix | 0.467 |
| NpHM5 | RVRAGFLLHGIKRSR | 40 | 5.25 | 2 | 1766.11 | β-chain | 0.160 |
| NpHM6 | RVRAGFLRHGIKRSA | 40 | 5.25 | 2 | 1724.03 | α-helix | 0.237 |
| NpHM7 | RVRAGFLRHGIKRSR | 33 | 6.25 | 3 | 1809.14 | α-helix | 0.281 |
| Peptides | Gram Negative | Gram Positive | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| E. coli DH5α | V. alginolyticus A056 | Vibrio parahaemolyticus 2013V-1174 | Staphylococcus aureus ATCC 25923 | Bacillus subtilis ATCC 6051 | ||||||
| MIC (μM) | MBC (μM) | MIC (μM) | MBC (μM) | MIC (μM) | MBC (μM) | MIC (μM) | MBC (μM) | MIC (μM) | MBC (μM) | |
| NpHM1 | 15–60 | >60 | 50–150 | >150 | 15–60 | >60 | >60 | >60 | >60 | >60 |
| NpHM2 | 30–60 | >50 | 70–150 | >75 | 30–60 | >60 | >50 | >50 | >50 | >50 |
| NpHM3 | 15–50 | >60 | 30–75 | >150 | 15–60 | >60 | >60 | >60 | >60 | >60 |
| NpHM4 | 50–150 | >150 | 5–25 | 30 | 15–45 | 50 | >50 | >50 | >50 | >50 |
| NpHM5 | >50 | >50 | 15–70 | >70 | 15–70 | >70 | >50 | >50 | >50 | >50 |
| NpHM6 | >50 | >50 | >70 | >70 | 15–60 | >60 | >50 | >50 | >50 | >50 |
| NpHM7 | >150 | >150 | >150 | >150 | >150 | >150 | >150 | >150 | >150 | >150 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, C.; Zheng, X.; Liu, K.; Yuan, W.; Zhang, Y.; Mao, F.; Bao, Y. Functional Characterization, Antimicrobial Effects, and Potential Antibacterial Mechanisms of NpHM4, a Derived Peptide of Nautilus pompilius Hemocyanin. Mar. Drugs 2022, 20, 459. https://doi.org/10.3390/md20070459
Yuan C, Zheng X, Liu K, Yuan W, Zhang Y, Mao F, Bao Y. Functional Characterization, Antimicrobial Effects, and Potential Antibacterial Mechanisms of NpHM4, a Derived Peptide of Nautilus pompilius Hemocyanin. Marine Drugs. 2022; 20(7):459. https://doi.org/10.3390/md20070459
Chicago/Turabian StyleYuan, Chun, Xiaoying Zheng, Kunna Liu, Wenbin Yuan, Yang Zhang, Fan Mao, and Yongbo Bao. 2022. "Functional Characterization, Antimicrobial Effects, and Potential Antibacterial Mechanisms of NpHM4, a Derived Peptide of Nautilus pompilius Hemocyanin" Marine Drugs 20, no. 7: 459. https://doi.org/10.3390/md20070459
APA StyleYuan, C., Zheng, X., Liu, K., Yuan, W., Zhang, Y., Mao, F., & Bao, Y. (2022). Functional Characterization, Antimicrobial Effects, and Potential Antibacterial Mechanisms of NpHM4, a Derived Peptide of Nautilus pompilius Hemocyanin. Marine Drugs, 20(7), 459. https://doi.org/10.3390/md20070459