
Sulfated Polysaccharide Extracted from the Green Algae Codium bernabei: Physicochemical Characterization and Antioxidant, Anticoagulant and Antitumor Activity

 , , ,
, , ,  , , ,
, , , 
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Polysaccharide Characterization
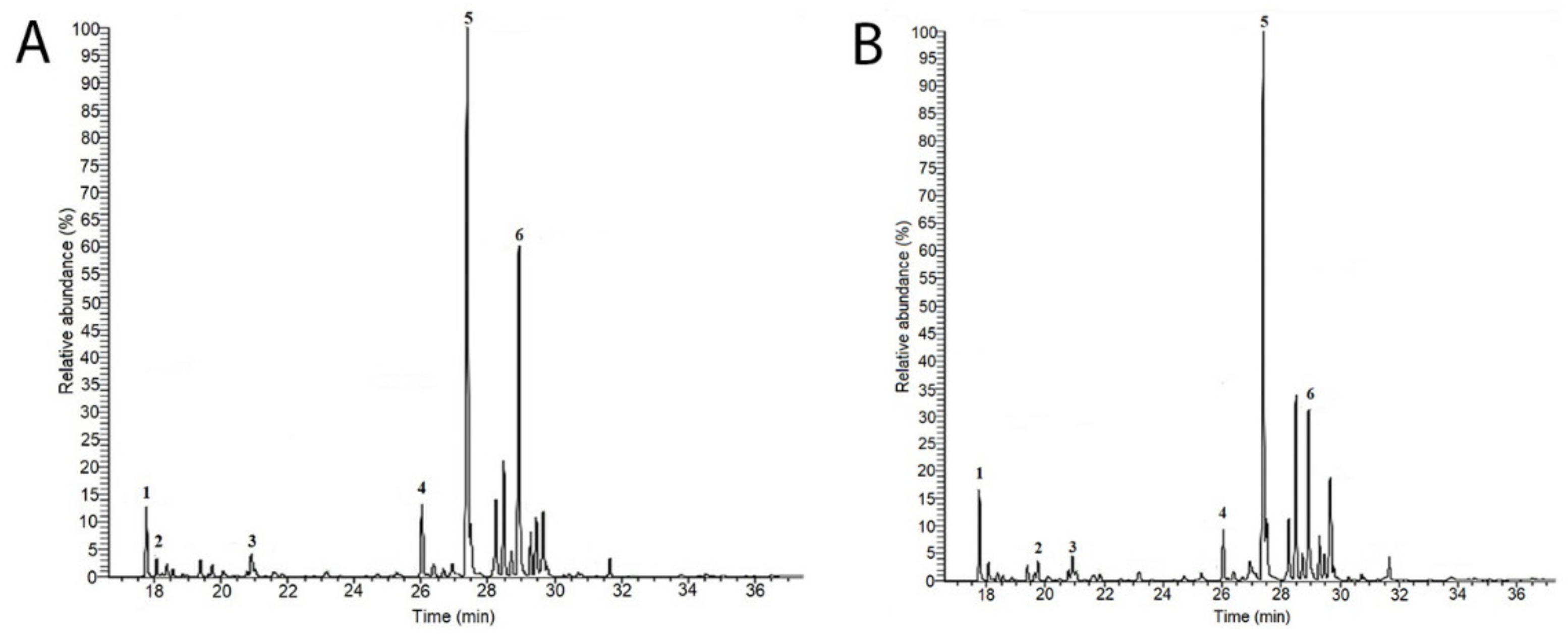
2.1.1. GC-MS Sugar Analysis
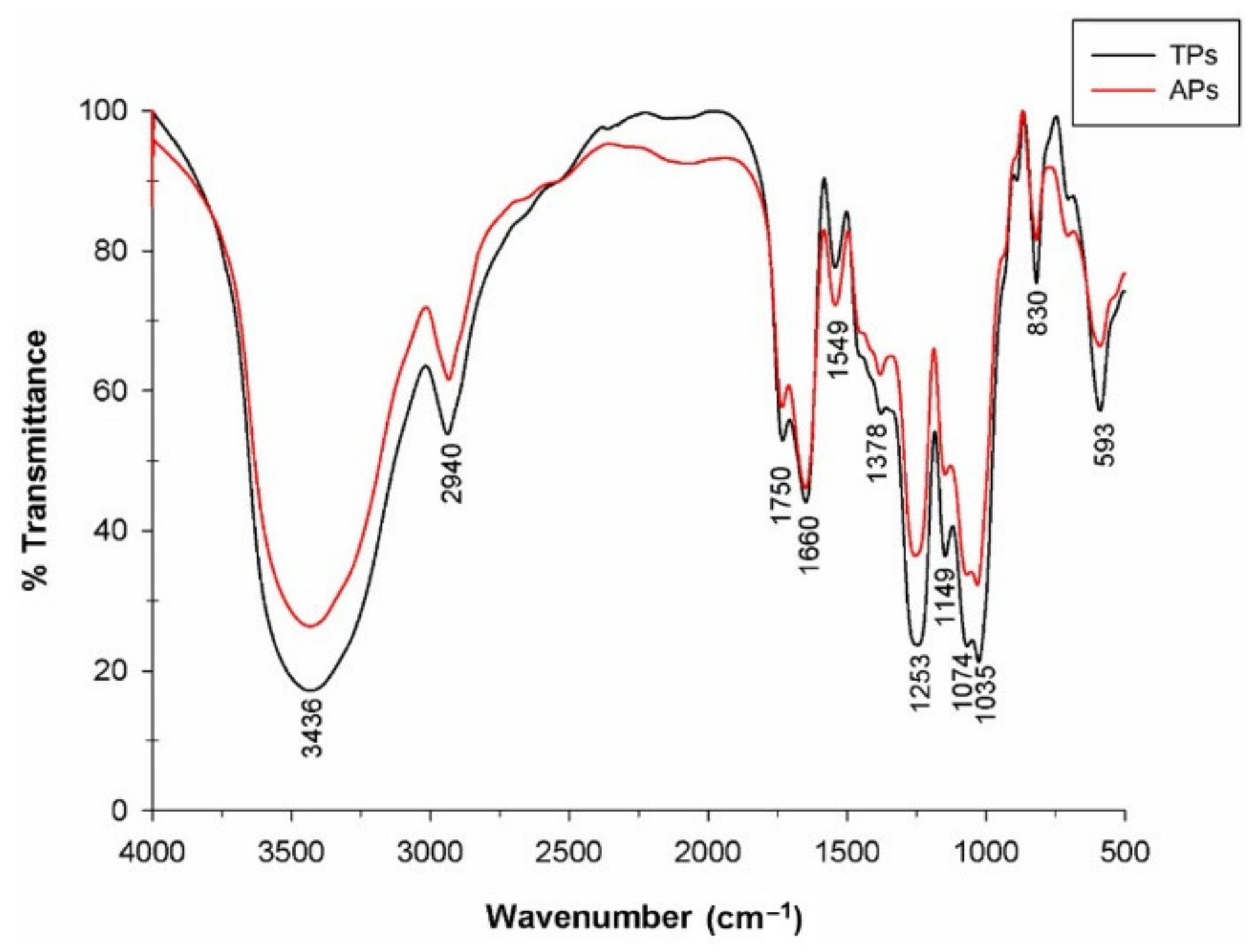
2.1.2. Fourier Transform Infrared Spectroscopy (FTIR)
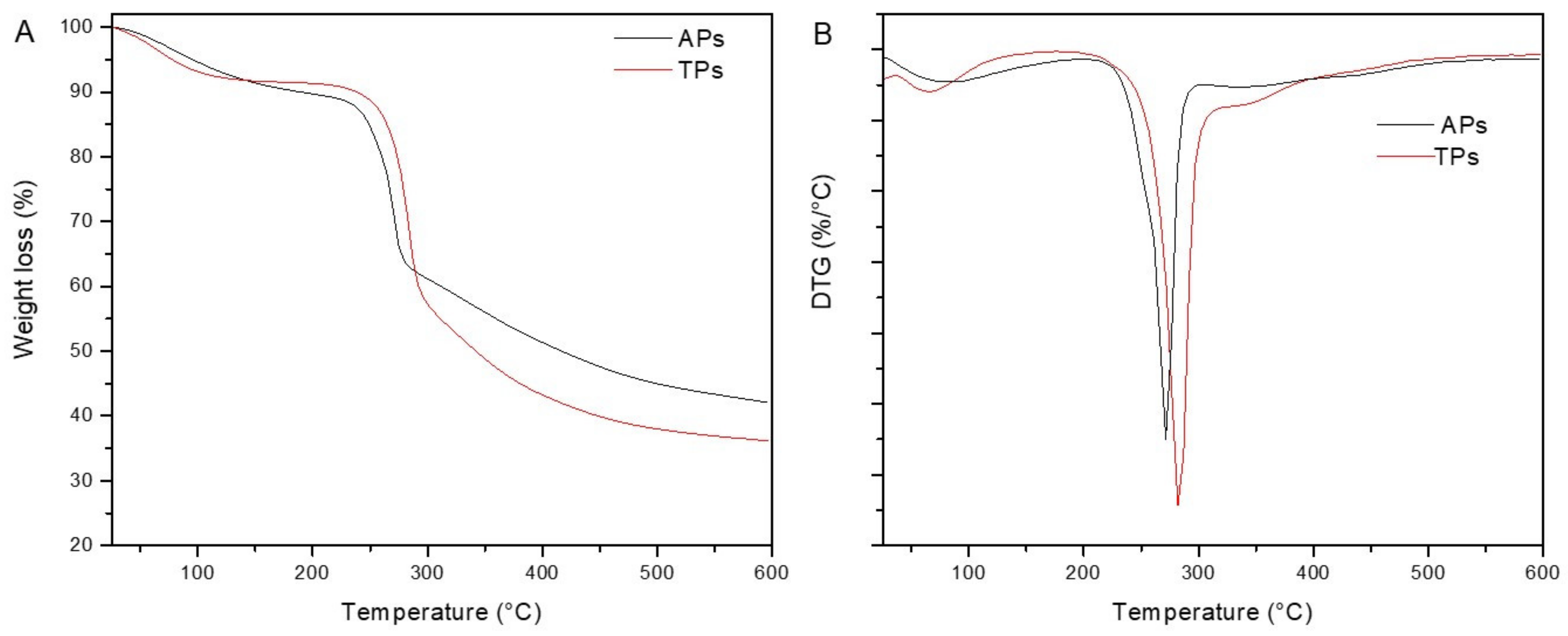
2.1.3. Thermogravimetric Properties (TGA-DSC)
2.1.4. SEM Analysis
2.2. Biological Activity
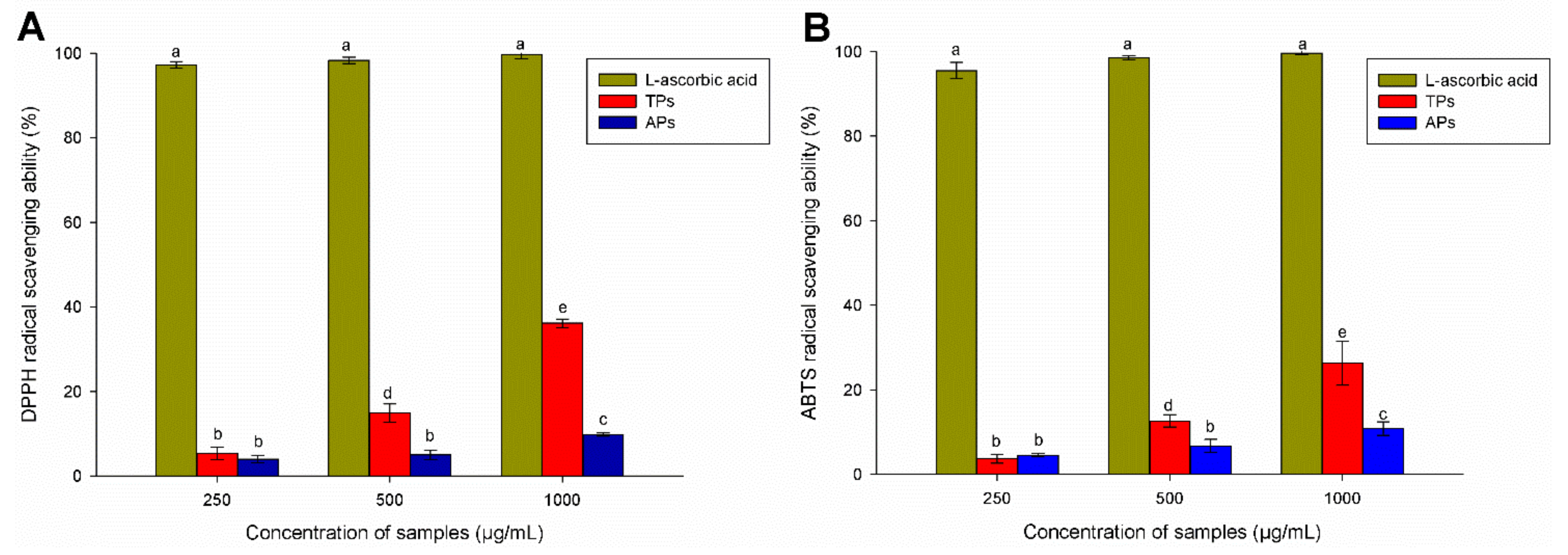
2.2.1. Antioxidant Activity
2.2.2. Anticoagulant Activity
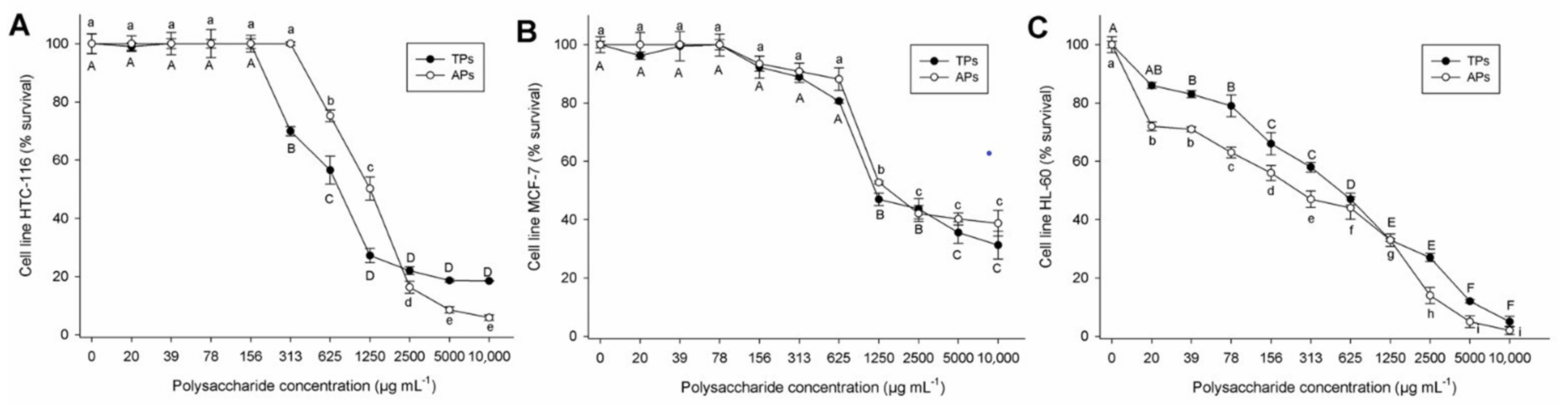
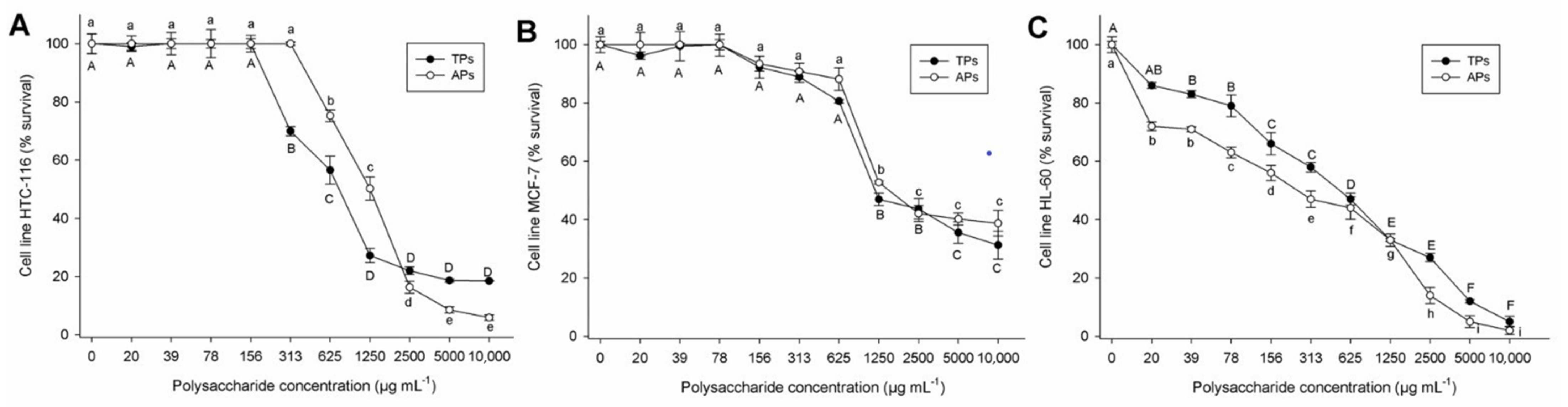
2.2.3. Cell Viability in Tumor Lines
3. Materials and Methods
3.1. Polysaccharide Isolation
3.2. Polysaccharide Characterization
3.2.1. Scanning Electron Microscopy (SEM) Analysis
3.2.2. Elemental Analysis
3.2.3. Fourier Transform Infrared Spectroscopy (FTIR) Analysis
3.2.4. Thermal Analysis
3.2.5. Molecular Weight
3.2.6. Determination of Sulfate Content
3.2.7. Monosaccharide Composition (GC/MS)
Hydrolysis and Derivatization
Gas Chromatography/Mass Spectrometry (GC-MS) Analysis
3.3. Biological Aactivity
3.3.1. Antioxidant Activity
3.3.2. Anticoagulant Activity
3.3.3. Cell Viability in Tumor Lines
3.4. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stiger-Pouvreau, V.; Bourgougnon, N.; Deslandes, E. Carbohydrates from Seaweeds. In Seaweed in Health and Disease Prevention; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 223–274. [Google Scholar]
- Gomez-Zavaglia, A.; Prieto Lage, M.A.; Jimenez-Lopez, C.; Mejuto, J.C.; Simal-Gandara, J. The potential of seaweeds as a source of functional ingredients of prebiotic and antioxidant value. Antioxidants 2019, 8, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salehi, B.; Sharifi-Rad, J.; Seca, A.M.L.; Pinto, D.C.G.A.; Michalak, I.; Trincone, A.; Mishra, A.P.; Nigam, M.; Zam, W.; Martins, N. Current trends on seaweeds: Looking at chemical composition, phytopharmacology, and cosmetic applications. Molecules 2019, 24, 4182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotas, J.; Leandro, A.; Pacheco, D.; Gonçalves, A.M.M.; Pereira, L. A Comprehensive Review of the Nutraceutical and Therapeutic Applications of Red Seaweeds (Rhodophyta). Life 2020, 10, 19. [Google Scholar] [CrossRef] [Green Version]
- Ciancia, M.; Quintana, I.; Vizcarguenaga, M.I.; Kasulin, L.; de Dios, A.; Estevez, J.M.; Cerezo, A.S. Polysaccharides from the green seaweeds Codium fragile and C. vermilara with controversial effects on hemostasis. Int. J. Biol. Macromol. 2007, 41, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Mallabo, M.R.B.; Corpuz, M.J.T.; Salonga, R.B.; Vasquez, R.D. Inhibitory Effect of Sulfated Polysaccharide from Codium edule P.C. Silva Against 2,4-Dinitrofluorobenzene (DNFB)-Induced Allergic Contact Dermatitis on Female BALB/c Mice. Adv. Pharm Bull. 2022, 12, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Je, J.-G.; Huang, C.; Oh, J.-Y.; Fu, X.; Wang, K.; Ahn, G.; Xu, J.; Gao, X.; Jeon, Y.-J. Anti-Inflammatory Effect of Sulfated Polysaccharides Isolated from Codium fragile In Vitro in RAW 264.7 Macrophages and In Vivo in Zebrafish. Mar. Drugs 2022, 20, 391. [Google Scholar] [CrossRef]
- Bedinia, E.; Laezza, A.; Parrilli, M.; Iadonisi, A. A review of chemical methods for the selective sulfation and desulfation of polysaccharides. Carbohydr. Polym. 2017, 174, 1224–1239. [Google Scholar] [CrossRef]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H.S. Chemical structures and bioactivities of sulfated polysaccharides from marine algae. Mar. Drugs. 2011, 9, 196–223. [Google Scholar] [CrossRef] [Green Version]
- Drira, M.; Hentati, F.; Babich, O.; Sukhikh, S.; Larina, V.; Sharifian, S.; Homai, A.; Fendri, I.; Lemos, M.; Félix, C.; et al. Bioactive Carbohydrate Polymers-Between Myth and Reality. Molecules 2021, 26, 7068. [Google Scholar] [CrossRef]
- Pomin, V.H. Review: An overview about the structure-function relationship of marine sulfated homopolysaccharides with regular chemical structures. Biopolymers. 2009, 91, 601–609. [Google Scholar] [CrossRef]
- Hentati, F.; Tounsi, L.; Djomdi, D.; Pierre, G.; Delattre, C.; Ursu, A.V.; Fendri, I.; Abdelkafi, S.; Michaud, P. Bioactive Polysaccharides from Seaweeds. Molecules 2020, 25, 3152. [Google Scholar] [CrossRef] [PubMed]
- Tabarsa, M.; Karnjanapratum, S.; Cho, M.L.; Kim, J.K.; You, S.G. Molecular characteristics and biological activities of anionic macromolecules from Codium fragile. Int. J. Biol. Macromol. 2013, 59, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.S.; Passos, C.P.; Madureira, P.; Vilanova, M.; Coimbra, M.A. Structure-function relationships of immunostimulatory polysaccharides: A review. Carbohydr. Polym. 2015, 132, 378–396. [Google Scholar] [CrossRef] [PubMed]
- Hou Yang, Y.; Lim, J.; Li, C.; Lee, S.; Hong, S. Effects of sulfated polysaccharides isolated from Codium fragile on inflammatory cytokine gene expression and Edwardsiella tarda infection in rockfish, Sebastes schlegelii. Fish Shellfish. Immunol. 2021, 112, 125–134. [Google Scholar] [CrossRef]
- Jurd, K.M.; Rogers, D.J.; Blunden, G.; McLellan, D.S. Anticoagulant properties of sulfated polysaccharide and a proteoglycan from Codium fragile ssp. atlanticum. J. Appl. Phycol. 1995, 7, 339–345. [Google Scholar] [CrossRef]
- Monmai, C.; You, S.; Park, W.J. Immune-enhancing effects of anionic macromolecules extracted from Codium fragile on cyclophosphamide-treated mice. PLoS ONE 2019, 14, e0211570. [Google Scholar] [CrossRef]
- Park, H.-B.; Hwang, J.; Zhang, W.; Go, S.; Kim, J.; Choi, I.; You, S.; Jin, J.-O. Polysaccharide from Codium fragile induces anti-cancer immunity by activating natural killer cells. Mar. Drugs 2020, 18, 626. [Google Scholar] [CrossRef]
- Wang, Y.; An, E.-K.; Kim, S.-J.; You, S.; Jin, J.-O. Intranasal administration of Codium fragile polysaccharide elicits anti-cancer immunity against Lewis lung carcinoma. Int. J. Mol. Sci. 2021, 22, 10608. [Google Scholar] [CrossRef]
- Yan, S.; Pan, C.; Yang, X.; Chen, S.; Qi, B.; Huang, H. Degradation of Codium cylindricum polysaccharides by H2O2-Vc-ultrasonic and H2O2-Fe2+-ultrasonic treatment: Structural characterization and antioxidant activity. Int. J. Biol. Macromol. 2021, 182, 129–135. [Google Scholar] [CrossRef]
- Silva, J.; Martins, A.; Alves, C.; Pinteus, S.; Gaspar, H.; Alfonso, A.; Pedrosa, R. Natural Approaches for Neurological Disorders—The Neuroprotective Potential of Codium tomentosum. Molecules 2020, 25, 5478. [Google Scholar] [CrossRef]
- Oh, S.; Kim, S.; Jung, K.; Pham, T.N.A.; Yang, S.; Ahn, B. Potential Prebiotic and Anti-Obesity Effects of Codium fragile Extract. Appl. Sci. 2022, 12, 959. [Google Scholar] [CrossRef]
- Kolsi, R.B.A.; Jardak, N.; Hajkacem, F.; Chaaben, R.; Jribi, I.; El Feki, A.; Rebai, T.; Jamoussi, K.; Fki, L.; Belghith, H.; et al. Anti-obesity effect and protection of liver-kidney functions by Codium fragile sulphated polysaccharide on high fat diet induced obese rats. Int. J. Biol. Macromol. 2017, 102, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Mori, M.; Matsumoto, H.; Hori, K.; Miyazawa, K. Antiangiogenic properties of a sulfated galactan isolated from a marine alga, Codium cylindricum. J. Appl. Phycol. 2003, 15, 87–90. [Google Scholar] [CrossRef]
- Lee, J.B.; Ohta, Y.; Hayashi, K.; Hayashi, T. Immunostimulating effects of a sulfated galactan from Codium fragile. Carbohydr. Res. 2010, 345, 1452–1454. [Google Scholar] [CrossRef]
- Surayot, U.; You, S. Structural effects of sulfated polysaccharides from Codium fragile on NK cell activation and cytotoxicity. Int. J. Biol. Macromol. 2017, 98, 117–124. [Google Scholar] [CrossRef]
- Pallela, R.; Park, I.K.P.K. Nutraceutical and pharmacological implications of marine carbohydrates. Adv. Food Nutr. Res. 2014, 73, 183–195. [Google Scholar]
- Fernández, P.V.; Estevez, J.M.; Cerezo, A.S.; Ciancia, M. Sulfated β-d-mannan from green seaweed Codium vermilara. Carbohydr. Polym. 2012, 87, 916–919. [Google Scholar] [CrossRef]
- Fernández, P.V.; Ciancia, M.; Miravalles, A.B.; Estevez, J.M. Cell-Wall Polymer Mapping in the Coenocytic Macroalga Codium Vermilara (Bryopsidales, Chlorophyta). J. Phycol. 2010, 46, 456–465. [Google Scholar] [CrossRef]
- Fernández, P.V.; Raffo, M.P.; Alberghina, J.; Ciancia, M. Polysaccharides from the green seaweed Codium decorticatum. Structure and cell wall distribution. Carbohydr. Polym. 2015, 117, 836–844. [Google Scholar] [CrossRef]
- Senthilkumar, D.; Jayanthi, S. Partial characterization and anticancer activities of purified glycoprotein extracted from green seaweed Codium decorticatum. J. Func. Foods 2016, 25, 323–332. [Google Scholar] [CrossRef]
- Aitouguinane, M.; el Alaoui-Talibi, Z.; Rchid, H.; Fendri, I.; Abdelkafi, S.; El-Hadj, M.D.O.; Boual, Z.; Dubessay, P.; Michaud, P.; le Cerf, D.; et al. A Novel Sulfated Glycoprotein Elicitor Extracted from the Moroccan Green Seaweed Codium decorticatum Induces Natural Defenses in Tomato. Appl. Sci. 2022, 12, 3643. [Google Scholar] [CrossRef]
- Robic, A.; Gaillard, C.; Sassi, J.F.; Lerat, Y.; Lahaye, M. Ultrastructure of ulvan: A polysaccharide from green seaweeds. Biopolymers 2009, 91, 652–664. [Google Scholar] [CrossRef] [PubMed]
- Parages, M.; Rico, R.; Abdala, R.; Chabrillón, M.; Sotiroudis, T.; Jiménez, C. Acidic polysaccharides of Arthrospira (Spirulina) platensis induce the synthesis of TNF-α in RAWmacrophages. J. Appl. Phycol. 2012, 24, 1537–1546. [Google Scholar] [CrossRef]
- Bouissil, S.; Alaoui-Talibi, Z.E.; Pierre, G.; Rchid, H.; Michaud, P.; Delattre, C.; El Modafar, C. Fucoidans of Moroccan Brown Seaweed as Elicitors of Natural Defenses in Date Palm Roots. Mar. Drugs 2020, 18, 596. [Google Scholar] [CrossRef]
- Hernández-Garibay, E.; Zertuche-González, J.A.; Pacheco-Ruíz, I. Isolation and chemical characterization of algal polysaccharides from the green seaweed Ulva clathrata (Roth) C. Agardh. J. Appl. Phycol. 2011, 23, 537–542. [Google Scholar] [CrossRef]
- Siddhanta, A.K.; Shanmugam, M.; Mody, K.H.; Goswami, A.M.; Ramavat, B.K. Sulphated polysaccharides of Codium dwarkense Boergs from the west coast of India: Chemical composition and blood anticoagulant activity. Int. J. Biol. Macromol. 1999, 26, 151–154. [Google Scholar] [CrossRef]
- Bilan, M.I.; Vinogradova, E.V.; Shashkov, A.S.; Usov, A.I. Structure of a highly pyruvylated galactan sulfate from the Pacific green alga Codium yezoense (Bryopsidales, Chlorophyta). Carbohydr. Res. 2007, 342, 586–596. [Google Scholar] [CrossRef]
- Li, N.; Mao, W.J.; Yan, M.X.; Liu, X.; Xia, Z.; Wang, S.Y.; Xiao, B.; Chen, C.L.; Zhang, L.F.; Cao, S.J. Structural characterization and anticoagulant activity of a sulfated polysaccharide from the green alga Codium divaricatum. Carbohydr. Polym. 2015, 121, 175–182. [Google Scholar] [CrossRef]
- Zuo, S.; Li, F.; Gu, X.; Wei, Z.; Qiao, L.Q.; Du, C.Y.; Chi, Y.Z.; Liu, R.Z.; Wang, P. Effects of low molecular weight polysaccharides from Ulva prolifera on the tolerance of Triticum aestivum to osmotic stress. Int. J. Biol. Macromol. 2021, 183, 12–22. [Google Scholar] [CrossRef]
- Alipour, H.J.; Rezaei, M.; Shabanpour, B.; Tabarsa, M. Effects of sulfatedpolysaccharides from green alga Ulva intestinalis on physicochemical properties and microstructure of silver carp surimi. Food Hydrocoll. 2017, 74, 87–96. [Google Scholar] [CrossRef]
- Pereira, L.; Amado, A.M.; Critchley, A.T.; van de Velde, F.; Ribeiro-Claro, P.J.A. Identification of selected seaweed polysaccharides (phycocolloids) by vibrational spectroscopy (FTIR-ATR and FTRaman). Food Hydrocoll. 2009, 23, 1903–1909. [Google Scholar] [CrossRef] [Green Version]
- Fernando, I.P.S.; Sanjeewa, K.K.A.; Samarakoon, K.W.; Lee, W.W.; Kim, H.S.; Kim, E.A.; Gunasekara, U.K.D.S.S.; Abeytunga, D.T.U.; Nanayakkara, C.; de Silva, E.D.; et al. FTIR characterization and antioxidant activity of water-soluble crude polysaccharides of Sri Lankan marine algae. Algae 2017, 32, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Alves, A.; Caridade, S.G.; Mano, J.F.; Sousa, R.A.; Reis, R.L. Extraction and physico-chemical characterization of a versatile biodegradable polysaccharide obtained from green algae. Carbohydr. Res. 2010, 345, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Parikh, A.; Madamwar, D. Partial characterization of extracellular polysaccharides from cyanobacteria. Bioresour. Technol. 2006, 97, 1822–1827. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.B.; Wang, H.C.; Liu, Y.W.; Lin, S.H.; Chou, H.N.; Sheen, L.Y. Immunomodulatory activities of polysaccharides from Chlorella pyrenoidosa in a mouse model of Parkinson’s disease. J. Funct. Foods 2014, 11, 103–113. [Google Scholar] [CrossRef]
- Alves, A.; Sousa, R.A.; Reis, R.L. In vitro cytotoxicity assessment of ulvan, a polysaccharide extracted from green algae. Phytother. Res. 2013, 27, 1143–1148. [Google Scholar] [CrossRef]
- Rodríguez-Jasso, R.M.; Mussatto, S.I.; Pastrana, L.; Aguilar, C.N.; Teixeira, J.A. Extraction of sulfated polysaccharides by autohydrolysis of brown seaweed Fucus vesiculosus. J. Appl. Phycol. 2013, 25, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Chen, Y.; Jiang, Z.; Zhong, S.; Chen, W.; Zheng, F.; Shi, G. Purification, partial characterization and bioactivity of sulfated polysaccharides from Grateloupia lívida. Int. J. Biol. Macromol. 2017, 94, 642–652. [Google Scholar] [CrossRef]
- Netanel Liberman, G.; Ochbaum, G.; Mejubovsky-Mikhelis, M.; Bitton, R.; Arad, S.M. Physico-chemical characteristics of the sulfated polysaccharides of the red microalgae Dixoniella grisea and Porphyridium aerugineum. Int. J. Biol. Macromol. 2020, 145, 1171–1179. [Google Scholar] [CrossRef]
- Wu, L.C.; Hsu, H.W.; Chen, Y.C.; Chiu, C.C.; Lin, Y.I.; Ho, J.A. Antioxidant andantiproliferative activities of red pitaya. Food Chem. 2006, 95, 319–327. [Google Scholar] [CrossRef]
- Borazjani, N.J.; Tabarsa, M.; You, S.; Rezaei, M. Improved immunomodulatory and antioxidant properties of unrefined fucoidans from Sargassum angustifolium by hydrolysis. J. Food Sci. Technol. 2017, 54, 4016–4025. [Google Scholar] [CrossRef] [PubMed]
- Foti, M.C. Use and abuse of the DPPH radical. J. Agr. Food. Chem. 2015, 63, 8765–8776. [Google Scholar] [CrossRef]
- Nalinanon, S.; Benjakul, S.; Kishimura, H.; Shahidi, F. Functionalities and antioxidant properties of protein hydrolysates from the muscle of ornate threadfin bream treated with pepsin from skipjack tuna. Food Chem. 2011, 124, 1354–1362. [Google Scholar] [CrossRef]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on mechanisms of in vitro antioxidant activity of polysaccharides. Oxidative Med. Cell. Longev. 2016, 2016, 5692852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obluchinskaya, E.D.; Pozharitskaya, O.N.; Zakharov, D.V.; Flisyuk, E.V.; Terninko, I.I.; Generalova, Y.E.; Smekhova, I.E.; Shikov, A.N. The Biochemical Composition and Antioxidant Properties of Fucus vesiculosus from the Arctic Region. Mar. Drugs 2022, 20, 193. [Google Scholar] [CrossRef]
- Shao, P.; Chen, X.; Sun, P. Improvement of Antioxidant and Moisture-Preserving Activities of Sargassum Horneri Polysaccharide Enzymatic Hydrolyzates. Int. J. Biol. Macromol. 2015, 74, 420–427. [Google Scholar] [CrossRef]
- Su, Y.; Li, L. Structural characterization and antioxidant activity of polysaccharide from four auriculariales. Carbohydr. Polym. 2020, 229, 115407. [Google Scholar] [CrossRef]
- Chagas, F.D.S.; Lima, G.C.; Santos, V.I.N.; Costa, L.E.C.; Sousa, W.M.; Sombra, V.G.; Freitas de Araújo, D.; Nogueira Barros, F.C.; Marinho-Soriano, E.; Pessoa de Andrade Feitosa, J.; et al. Sulfated polysaccharide from the red algae Gelidiella acerosa: Anticoagulant, antiplatelet and antithrombotic effects. Int. J. Biol. Macromol. 2020, 159, 415–421. [Google Scholar] [CrossRef]
- Fernandez, P.V.; Quintana, I.; Cerezo, A.S.; Caramelo, J.J.; Pol-Fachin, L.; Verli, H.; Estevez, J.M.; Ciancia, M. Anticoagulant activity of a unique sulfated pyranosic (1→3)-β-l-arabinan through direct interaction with thrombin. J. Biol. Chem. 2013, 288, 223–233. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, P.V.; Arata, P.X.; Ciancia, M. Polysaccharides from Codium species: Chemical structure and biological activity. Their role as components of the cell wall. Adv. Bot. Res. 2014, 71, 253–278. [Google Scholar]
- Shanmugam, M.; Ramavat, B.K.; Mody, K.H.; Oza, R.M.; Tewari, A. Distribution of heparinoid- active sulphated polysaccharides in some Indian marine green algae. Indian J. Mar. Sci. 2001, 30, 222–227. [Google Scholar]
- Matsubara, K.; Matsuura, Y.; Bacic, A.; Liao, M.L.; Hori, K.; Miyazawa, K. Anticoagulant properties of a sulfated galactan preparation from a marine green alga, Codium cylindricum. Int. J. Biol. Macromol. 2001, 28, 395–399. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Obluchinskaya, E.D.; Shikov, A.N. mechanisms of bioactivities of fucoidan from the brown seaweed Fucus vesiculosus L. of the Barents Sea. Mar. Drugs 2020, 18, 275. [Google Scholar] [CrossRef]
- Adrien, A.; Bonnet, A.; Dufour, D.; Baudouin, S.; Maugard, T.; Bridiau, N. anticoagulant activity of sulfated ulvan isolated from the green macroalga Ulva rigida. Mar. Drugs. 2019, 17, 291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, M.; Yang, Y.; Shao, Z.; Zhang, J.; Feng, C.; Wang, L.; Mao, W. Chemical Structure and Anticoagulant Property of a Novel Sulfated Polysaccharide from the Green Alga Cladophora oligoclada. Mar. Drugs. 2021, 19, 554. [Google Scholar] [CrossRef] [PubMed]
- Al Monla, R.; Dassouki, Z.; Sari-Chmayssem, N.; Mawlawi, H.; Gali-Muhtasib, H. Fucoidan and Alginate from the Brown Algae Colpomenia sinuosa and Their Combination with Vitamin C Trigger Apoptosis in Colon Cancer. Molecules 2022, 27, 358. [Google Scholar] [CrossRef]
- Vishchuk, O.S.; Ermakova, S.P.; Zvyagintseva, T.N. The effect of sulfated (1→3)-α-l-fucan from the brown alga Saccharina cichorioides Miyabe on resveratrol-induced apoptosis in colon carcinoma Cells. Mar. Drugs 2013, 11, 194–212. [Google Scholar] [CrossRef] [Green Version]
- Abdala Díaz, R.T.; Casas Arrojo, V.; Arrojo Agudo, M.A.; Cárdenas, C.; Dobretsov, S.; Figueroa, F.L. Immunomodulatory and Antioxidant Activities of Sulfated Polysaccharides from Laminaria ochroleuca, Porphyra umbilicalis and Gelidium corneum. Mar. Biotechnol. 2019, 21, 577–587. [Google Scholar] [CrossRef]
- Gonzalez, A.V.; Chacana, M.E.; Silva, P.C. Codium bernabei sp. nov. (Bryopsidales Chlorophyta), a coalescing green seaweed from the coast of Chile. Phycologia 2012, 51, 666–671. [Google Scholar] [CrossRef]
- Figueroa, F.A.; Abdala-Díaz, R.T.; Hernández, V.; Pedreros, P.; Aranda, M.; Cabrera-Pardo, J.R.; Pérez, C.; Becerra, J.; Urrutia, R. Invasive diatom Didymosphenia geminata as a source of polysaccharides with antioxidant and immunomodulatory effects on macrophage cell lines. J. Appl. Phycol. 2020, 32, 93–102. [Google Scholar] [CrossRef]
- Lourenço, S.O.; Barbarino, E.; Lavín, P.L.; Lanfer Marquez, U.M.; Aidar, E. Distribution of intracelular nitrogen in marine microalgae: Calculation of new nitrogen to protein conversion factors. Eur. J. Phycol. 2004, 39, 17–32. [Google Scholar] [CrossRef]
- Banerjee, A.; Breig, S.J.M.; Gómez, A.; González-Faune, P.; Sánchez-Arévalo, I.; Sarkar, S.; Bandopadhyay, R.; Vuree, S.; Cornejo, J.; Tapia, J.; et al. Optimization and Characterization of a Novel Exopolysaccharide from Bacillus haynesii CamB6 for Food Applications. Biomolecules 2022, 12, 834. [Google Scholar] [CrossRef] [PubMed]
- Therho, T.T.; Hartiala, K. Method for determination of the sulfate content of glycosamino glycans. Anal. Biochem. 1971, 41, 471–476. [Google Scholar] [CrossRef]
- Wu, G.J.; Shiu, S.M.; Hsieh, M.C.; Tsai, G.J. Anti-inflammatory activity of a sulfated polysaccharide from the brown alga Sargassum cristaefolium. Food Hydrocoll. 2016, 53, 16–23. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Mourao, P.A.S.; Pereira, M.S.; Pavao, M.S.G.; Mulloy, B.; Tollefsen, D.M.; Mowinckel, M.C.; Abildgaard, U. Structure and anticoagulant activity of a fucosylated chon-droitin sulfate from echinoderm. Sulfated fucose branches on the polysaccharideaccount for its high anticoagulant action. J. Biol. Chem. 1996, 271, 23973–23984. [Google Scholar] [CrossRef] [Green Version]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol Meth. 1983, 65, 55–63. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter/Sample | TPs | APs | Codium fragile [5,13] | Codium vermilara [28,29] | Codium decorticatum [30,31,32] |
|---|---|---|---|---|---|
| Yield (wt%) | 2.9 | 2.1 | 6.7 | 6.8 | 3.4 |
| Sugar composition (mol%) | |||||
| Xylose | 2.8 | 3.3 | 3.2 | 2 | 3.0 |
| Glucose | 31.1 | 19.3 | 4.8 | 1.9 | 5.1 |
| Galactose | 51.8 | 61.0 | 61.6 | 49.8 | 36.5 |
| Fucose | 1.2 | 2.1 | tr a | - | 1.6 |
| Mannose | 7.4 | 6.1 | 3.9 | 3.6 | 2.6 |
| Arabinose | 5.7 | 8.2 | 23.1 | 44.7 | 48.8 |
| Degree of sulfation (−SO32−) | 19.37 | 17.2 | 13.0 | 30.4 | 22.5 |
| Protein content (wt%) | 15.6 | 3.5 | 11.3 | 16.2 | 20.7 |
| Mw (kDa) | 17.9 | 14.9 | 11 | 66 | |
| Mn (kDa) | 8.9 | 9.1 | - | - | - |
| Polydispersity Index | 2.0 | 1.6 | - | - | - |
| Sample | Temperature (°C) | Weight Los (%) | Char (%) | ||
|---|---|---|---|---|---|
| Onset | Peak | End | |||
| TPs | 37.1 | 66.3 | 164.3 | 8.2 | 36.2 |
| 196.8 | 282.5 | 402.2 | 52.7 | ||
| APs | 28.6 | 79.1 | 194.1 | 8.1 | 42.4 |
| 208.6 | 271.7 | 393.5 | 37.2 | ||
| Samples | PT (s) * | APTT (s) * |
|---|---|---|
| Normal plasma pool | 10.9 ± 0.2 A | 25.6 ± 0.3 A |
| Normal plasma pool + DMSO | 11.0 ± 0.1 A | 26.4 ± 0.1 A |
| Normal plasma pool + TPs (1000 µg mL−1) | 17.8 ± 1.3 B | >50 |
| Normal plasma pool + TPs (100 µg mL−1) | 10.9 ± 0.2 A | 45.7 ± 2.5 B |
| Normal plasma pool + TPs (10 µg mL−1) | 10.5 ± 0.1 A | 26.9 ± 0.3 A |
| Normal plasma pool + APs (1000 µg mL−1) | 10.4 ± 0.1 A | 35.2 ± 0.1 B |
| Normal plasma pool + APs (100 µg mL−1) | 10.7 ± 0.1 A | 26.8 ± 0.1 A |
| Normal plasma pool + APs (10 µg mL−1) | 10.7 ± 0.1 A | 26.4 ± 0.2 A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figueroa, F.A.; Abdala-Díaz, R.T.; Pérez, C.; Casas-Arrojo, V.; Nesic, A.; Tapia, C.; Durán, C.; Valdes, O.; Parra, C.; Bravo-Arrepol, G.; et al. Sulfated Polysaccharide Extracted from the Green Algae Codium bernabei: Physicochemical Characterization and Antioxidant, Anticoagulant and Antitumor Activity. Mar. Drugs 2022, 20, 458. https://doi.org/10.3390/md20070458
Figueroa FA, Abdala-Díaz RT, Pérez C, Casas-Arrojo V, Nesic A, Tapia C, Durán C, Valdes O, Parra C, Bravo-Arrepol G, et al. Sulfated Polysaccharide Extracted from the Green Algae Codium bernabei: Physicochemical Characterization and Antioxidant, Anticoagulant and Antitumor Activity. Marine Drugs. 2022; 20(7):458. https://doi.org/10.3390/md20070458
Chicago/Turabian StyleFigueroa, Fabian A., Roberto T. Abdala-Díaz, Claudia Pérez, Virginia Casas-Arrojo, Aleksandra Nesic, Cecilia Tapia, Carla Durán, Oscar Valdes, Carolina Parra, Gastón Bravo-Arrepol, and et al. 2022. "Sulfated Polysaccharide Extracted from the Green Algae Codium bernabei: Physicochemical Characterization and Antioxidant, Anticoagulant and Antitumor Activity" Marine Drugs 20, no. 7: 458. https://doi.org/10.3390/md20070458
APA StyleFigueroa, F. A., Abdala-Díaz, R. T., Pérez, C., Casas-Arrojo, V., Nesic, A., Tapia, C., Durán, C., Valdes, O., Parra, C., Bravo-Arrepol, G., Soto, L., Becerra, J., & Cabrera-Barjas, G. (2022). Sulfated Polysaccharide Extracted from the Green Algae Codium bernabei: Physicochemical Characterization and Antioxidant, Anticoagulant and Antitumor Activity. Marine Drugs, 20(7), 458. https://doi.org/10.3390/md20070458