
Anti-Quorum-Sensing Activity of Tryptophan-Containing Cyclic Dipeptides


Abstract
:
1. Introduction
2. Results
2.1. Antimicrobial Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Inhibitory Ratio (%) | |
|---|---|---|
| CV026 | PAO1 | |
| c(WS) | 70% a | 80% a |
| c(ws) | 23% b | 79% a |
| c(Ws) | 50% a | 77% a |
| c(wS) | 67% a | 79% a |
| c(WA) | 39% b | 80% a |
| c(WT) | 11% b | 83% a |
| c(WK) | 27% b | 76% a |
| c(WE) | 59% a | 100% a |
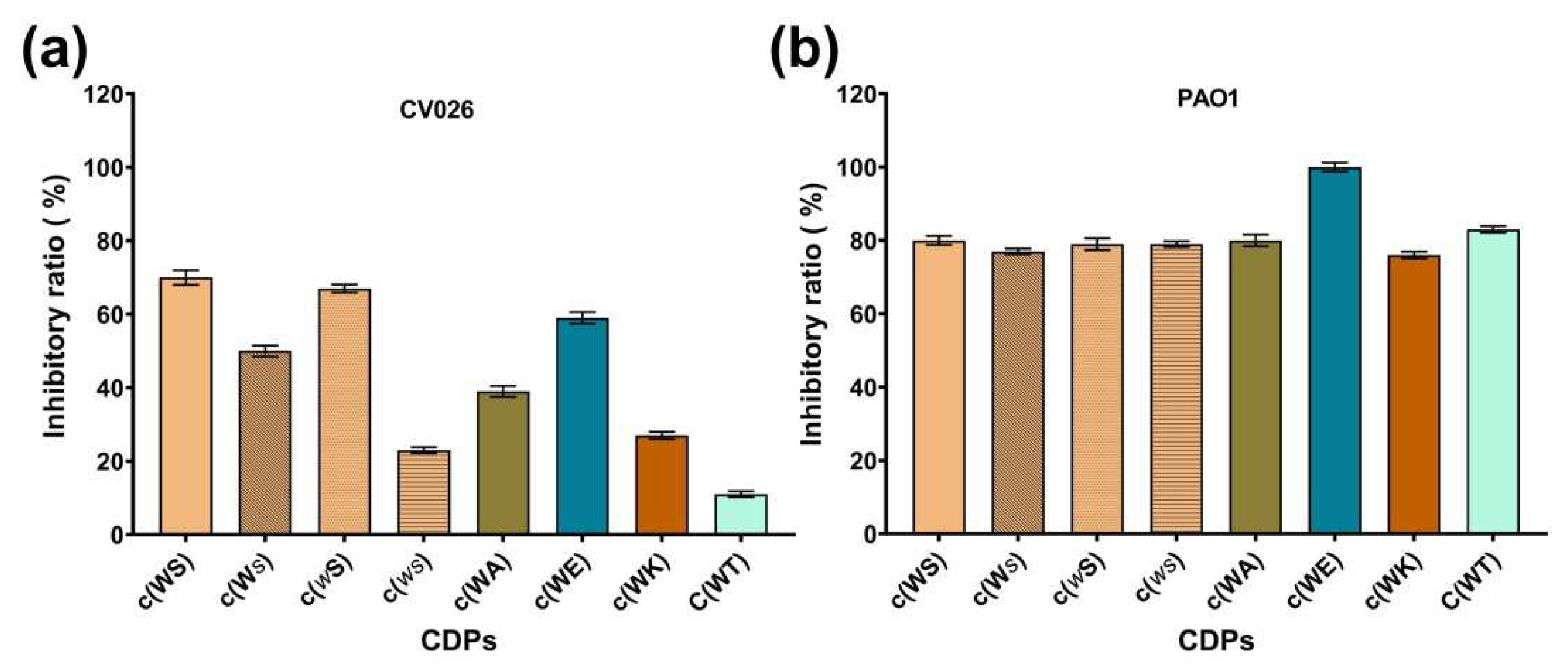
2.2. Anti-QS Ability against C. violaceum CV026

| Molecules | Binding Energy (kcal/mol) | Hydrogen Bonding Interactions |
|---|---|---|
| C6HSL | −6.04 | Asp97, Ser155, Trp84, Tyr80 |
| c(WS) | −7.83 | Tyr88, Ser155 |
| c(ws) | −7.88 | Tyr88, Ser155 |
| c(Ws) | −7.78 | Tyr88, Tyr80 |
| c(wS) | −7.52 | No hydrogen bonds formed |
| c(WA) | −7.56 | Tyr88, Trp84, Ser155 |
| c(WT) | −7.90 | Trp84, Ser155, Asp97, Tyr80 |
| c(WK) | −8.14 | Tyr88, Ser155 |
| c(WE) | −8.09 | Tyr88, Ser155 |
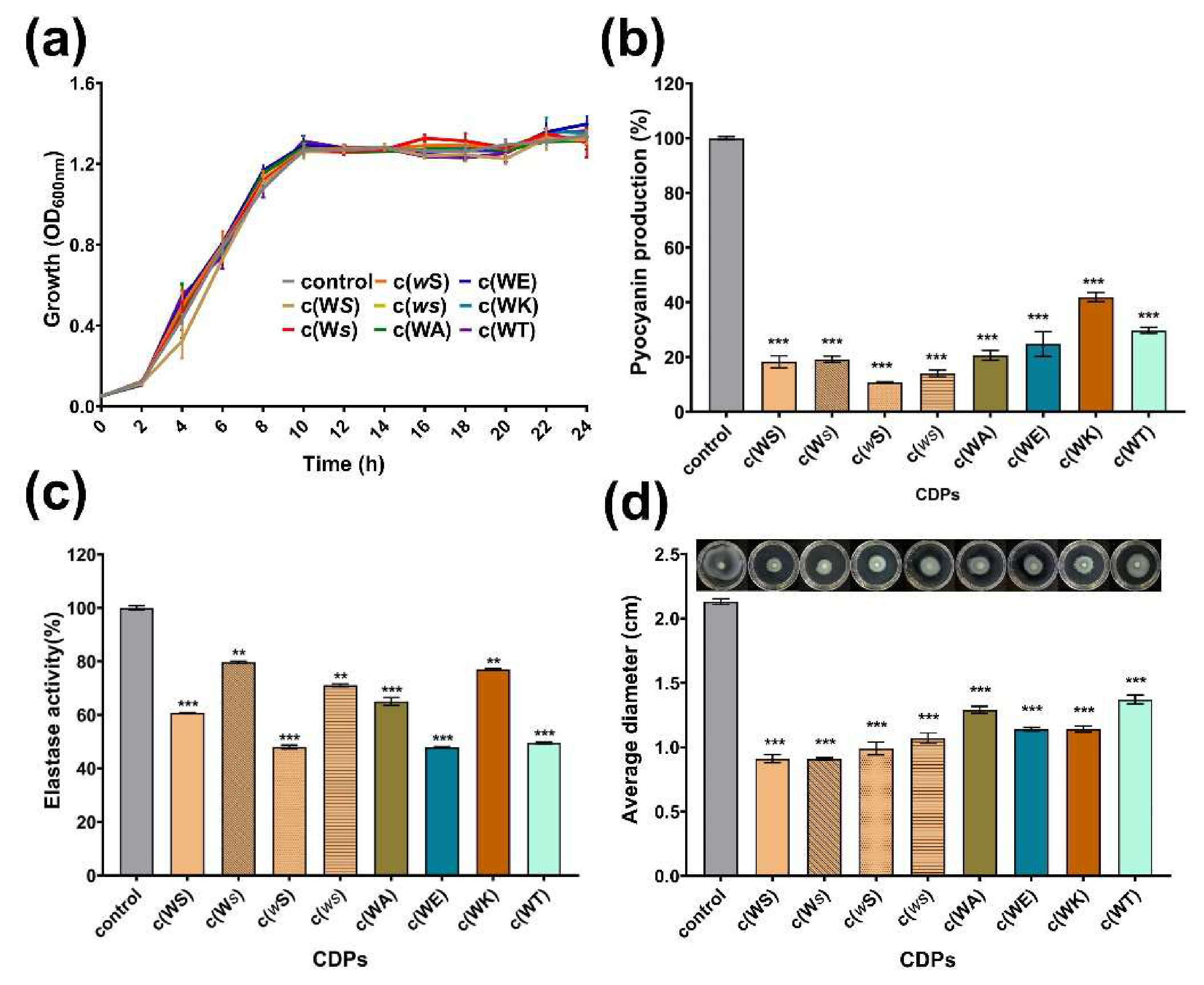
2.3. Inhibition on Production of Virulence Factors of P. aeruginosa PAO1
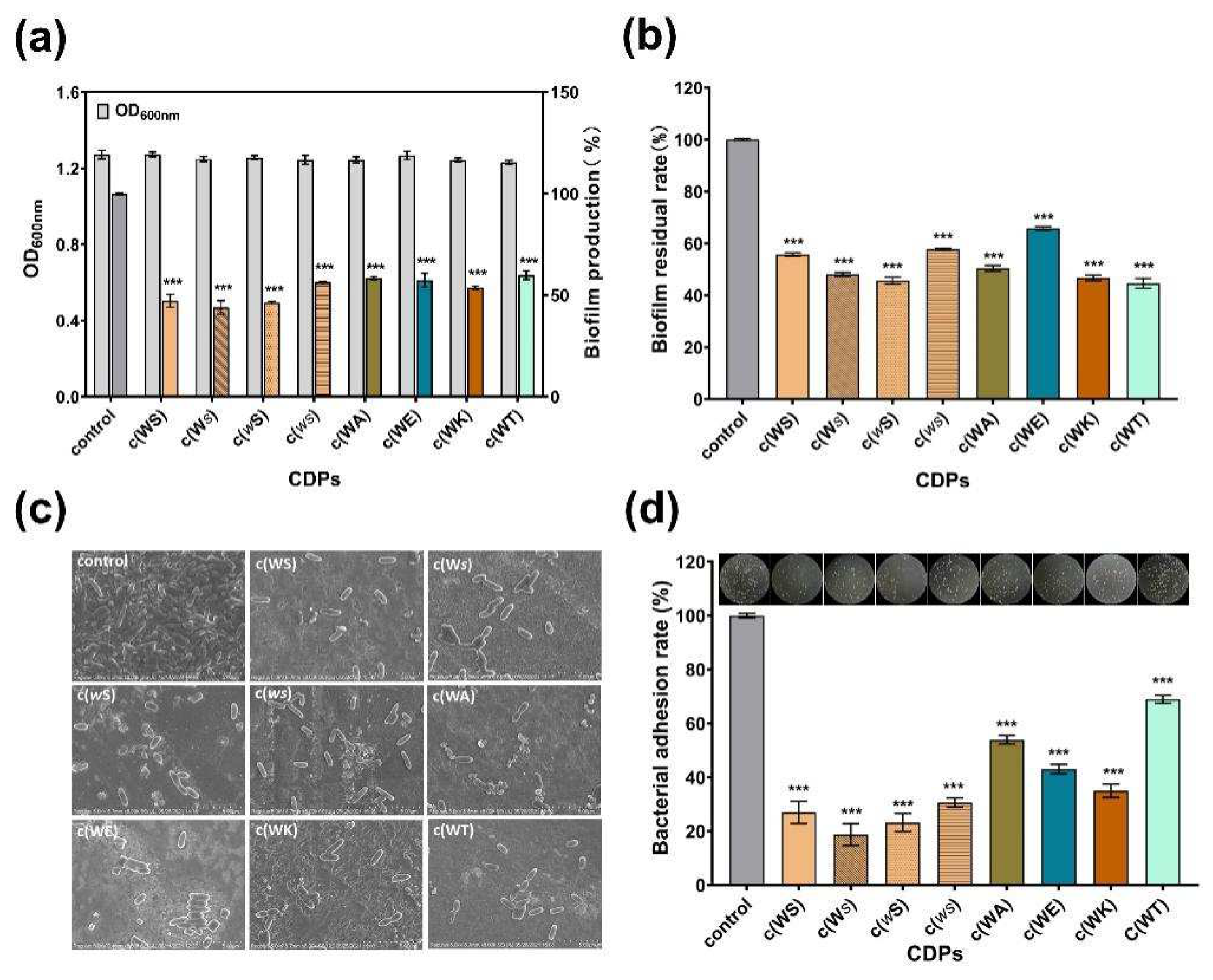
2.4. Inhibition on Biofilm and Adhesion of P. aeruginosa PAO1
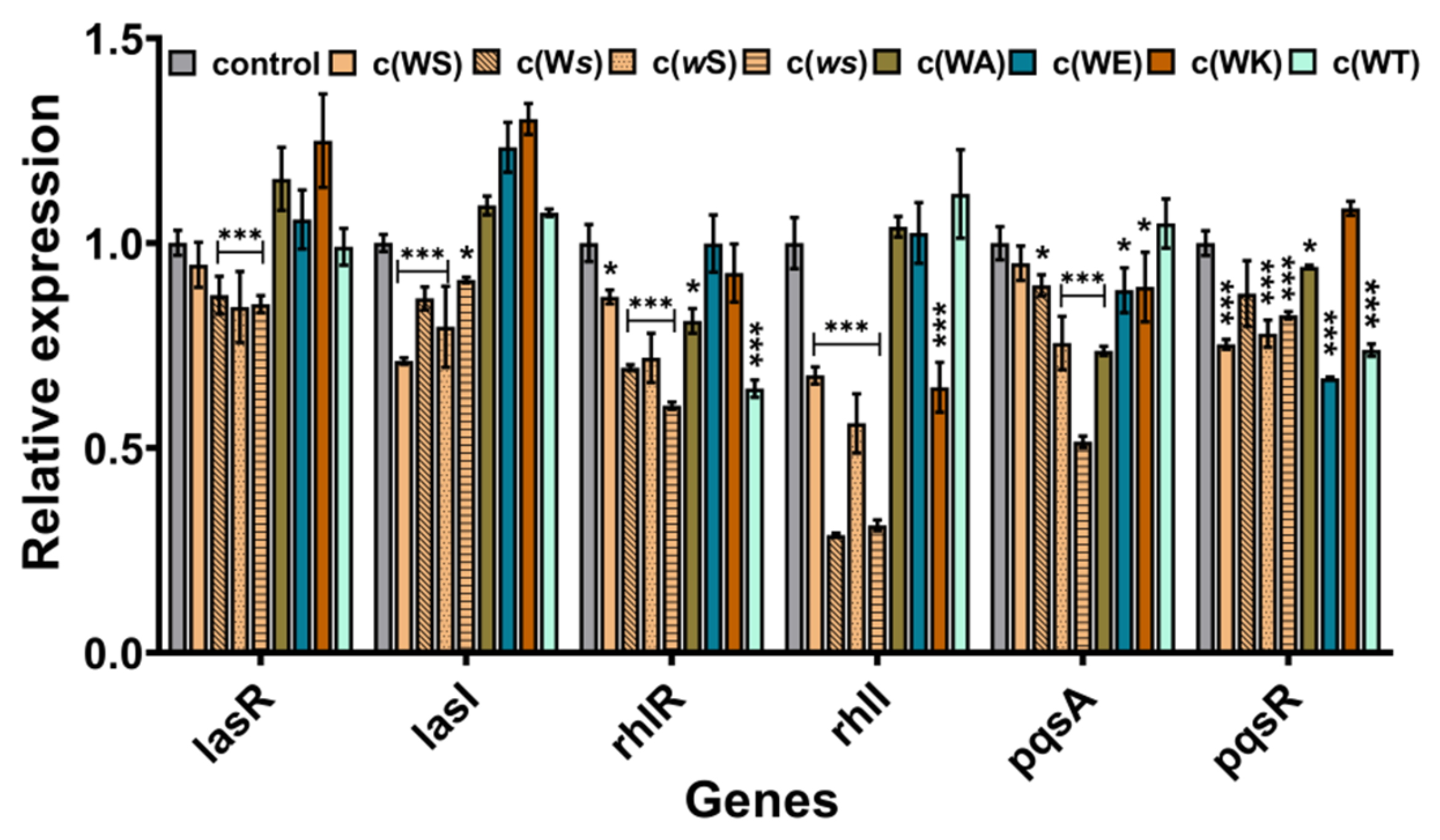
2.5. Inhibition on QS-Regulated Genes of P. aeruginosa PAO1
2.6. Hemolysis and Cytotoxicity

3. Discussion
4. Materials and Methods
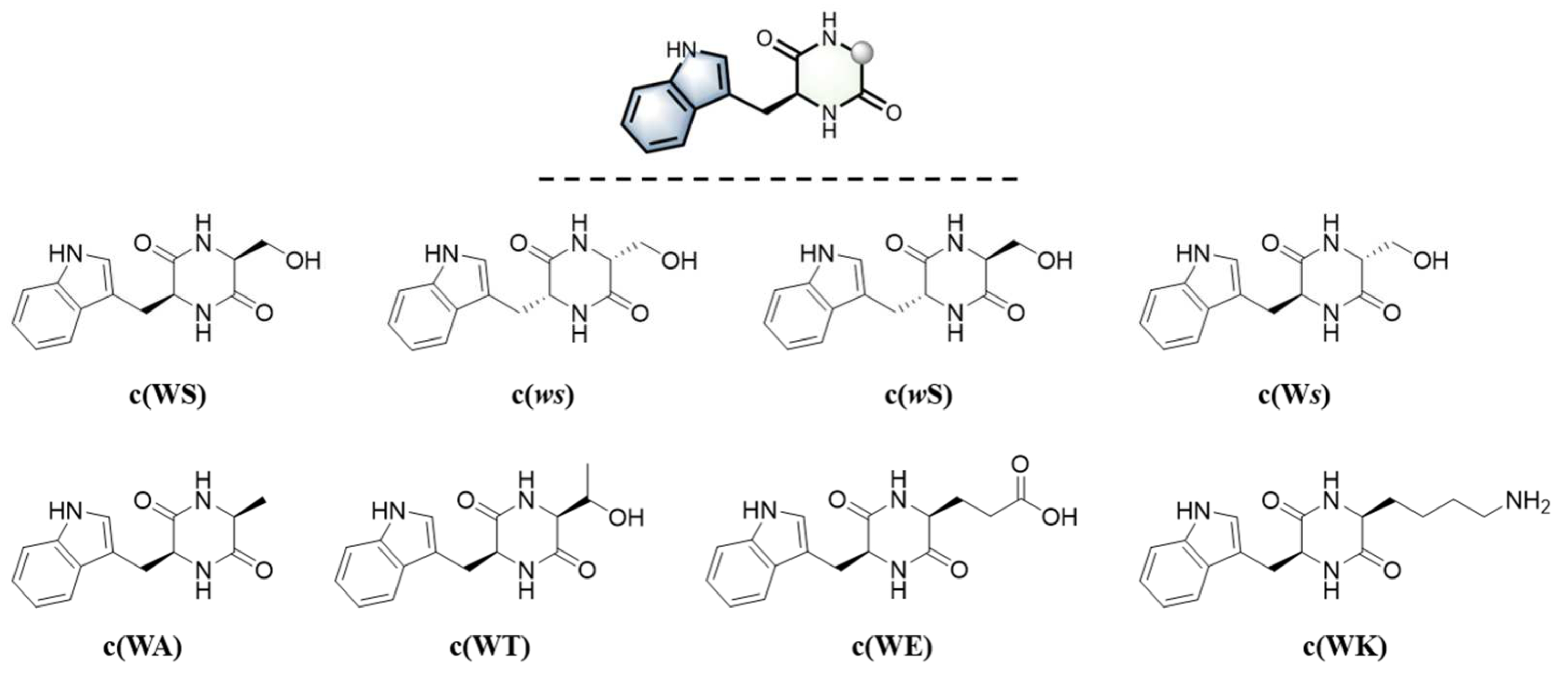
4.1. Materials and Synthetic Methods
4.2. Strains and Growth Medium
4.3. Antimicrobial Assay and Bacterial Growth Measurement
4.4. Violacein Quantification
4.5. Docking
4.6. Pyocyanin Quantification and Elastase Activity Assay
4.7. Swimming Motility
4.8. QS Genes Expression Assay
4.9. Biofilm Assay
4.9.1. Inhibition of Biofilm Formation
4.9.2. Biofilm Dispersion Assay
4.9.3. Observation of Morphology
4.10. Adherence Assay
4.11. Cytotoxicity and Hemolytic Activity
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Waters, C.M.; Bassler, B.L. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef] [PubMed]
- Antunes, L.C.; Ferreira, R.B.; Buckner, M.M.; Finlay, B.B. Quorum sensing in bacterial virulence. Microbiology 2010, 156, 2271–2282. [Google Scholar] [CrossRef] [PubMed]
- Clatworthy, A.E.; Pierson, E.; Hung, D.T. Targeting virulence: A new paradigm for antimicrobial therapy. Nat. Chem. Biol. 2007, 3, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Dickey, S.W.; Cheung, G.Y.C.; Otto, M. Different drugs for bad bugs: Antivirulence strategies in the age of antibiotic resistance. Nat. Rev. Drug Discov. 2017, 16, 457–471. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Bassler, B.L. Surviving as a Community: Antibiotic Tolerance and Persistence in Bacterial Biofilms. Cell Host Microbe 2019, 26, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Gray, B.; Hall, P.; Gresham, H. Targeting agr- and agr-like quorum sensing systems for development of common therapeutics to treat multiple gram-positive bacterial infections. Sensors 2013, 13, 5130–5166. [Google Scholar] [CrossRef] [PubMed]
- Carradori, S.; Di Giacomo, N.; Lobefalo, M.; Luisi, G.; Campestre, C.; Sisto, F. Biofilm and quorum sensing inhibitors: The road so far. Expert Opin. Ther. Pat. 2020, 30, 917–930. [Google Scholar] [CrossRef]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef]
- Zhao, X.; Yu, Z.; Ding, T. Quorum-sensing regulation of antimicrobial resistance in bacteria. Microorganisms 2020, 8, 425. [Google Scholar] [CrossRef]
- Hirsch, E.B.; Tam, V.H. Impact of multidrug-resistant Pseudomonas aeruginosa infection on patient outcomes. Expert Rev. Pharmacoecon. Outcomes Res. 2010, 10, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Fazeli, H.; Akbari, R.; Moghim, S.; Narimani, T.; Arabestani, M.R.; Ghoddousi, A.R. Pseudomonas aeruginosa infections in patients, hospital means, and personnel’s specimens. J. Res. Med. Sci. 2012, 17, 332–337. [Google Scholar] [PubMed]
- De Bentzmann, S.; Plesiat, P. The Pseudomonas aeruginosa opportunistic pathogen and human infections. Environ. Microbiol. 2011, 13, 1655–1665. [Google Scholar] [CrossRef]
- Song, D.; Meng, J.; Cheng, J.; Fan, Z.; Chen, P.; Ruan, H.; Tu, Z.; Kang, N.; Li, N.; Xu, Y.; et al. Pseudomonas aeruginosa quorum-sensing metabolite induces host immune cell death through cell surface lipid domain dissolution. Nat. Microbiol. 2019, 4, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Soukarieh, F.; Williams, P.; Stocks, M.J.; Cámara, M. Pseudomonas aeruginosa quorum sensing systems as drug discovery targets: Current position and future perspectives. J. Med. Chem. 2018, 61, 10385–10402. [Google Scholar] [CrossRef]
- Chang, A.; Sun, S.; Li, L.; Dai, X.; Li, H.; He, Q.; Zhu, H. Tyrosol from marine Fungi, a novel quorum sensing inhibitor against Chromobacterium violaceum and Pseudomonas aeruginosa. Bioorg. Chem. 2019, 91, 103140. [Google Scholar] [CrossRef]
- Chang, A.; He, Q.; Li, L.; Yu, X.; Sun, S.; Zhu, H. Exploring the quorum sensing inhibition of isolated chrysin from Penicillium chrysogenum DXY-1. Bioorg. Chem. 2021, 111, 104894. [Google Scholar] [CrossRef]
- Yu, X.; Li, L.; Sun, S.; Chang, A.; Dai, X.; Li, H.; Wang, Y.; Zhu, H. A cyclic dipeptide from marine fungus Penicillium chrysogenum DXY-1 exhibits anti-quorum sensing activity. ACS Omega 2021, 6, 7693–7700. [Google Scholar] [CrossRef]
- Sun, S.W.; Dai, X.Y.; Sun, J.; Bu, X.G.; Weng, C.H.; Li, H.; Zhu, H. A diketopiperazine factor from Rheinheimera aquimaris QSI02 exhibits anti-quorum sensing activity. Sci. Rep. 2016, 6, 39637. [Google Scholar] [CrossRef]
- Zhu, H.; Sun, S.W.; Li, H.; Chang, A.; Liu, Y.C.; Qian, J.; Shen, Y.L. Significantly improved production of Welan gum by Sphingomonas sp. WG through a novel quorum-sensing-interfering dipeptide cyclo(L-Pro-L-Phe). Int. J. Biol. Macromol. 2019, 126, 118–122. [Google Scholar] [CrossRef]
- Galloway, W.R.; Hodgkinson, J.T.; Bowden, S.D.; Welch, M.; Spring, D.R. Quorum sensing in gram-negative bacteria: Small-molecule modulation of AHL and AI-2 quorum sensing pathways. Chem. Rev. 2011, 111, 28–67. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.; Lin, Q.; Geske, G.D.; Blackwell, H.E. New and unexpected insights into the modulation of LuxR-type quorum sensing by cyclic dipeptides. ACS Chem. Biol. 2009, 4, 1051–1059. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, M.P.; Abraham, W.R. Antimicrobial and biofilm inhibiting diketopiperazines. Curr. Med. Chem. 2012, 19, 3564–3577. [Google Scholar] [CrossRef] [PubMed]
- Simon, G.; Berube, C.; Voyer, N.; Grenier, D. Anti-biofilm and anti-adherence properties of novel cyclic dipeptides against oral pathogens. Bioorg. Med. Chem. 2019, 27, 2323–2331. [Google Scholar] [CrossRef]
- Chen, G.; Swem, L.R.; Swem, D.L.; Stauff, D.L.; O’Loughlin, C.T.; Jeffrey, P.D.; Bassler, B.L.; Hughson, F.M. A strategy for antagonizing quorum sensing. Mol. Cell 2011, 42, 199–209. [Google Scholar] [CrossRef]
- Borthwick, A.D. 2,5-Diketopiperazines: Synthesis, reactions, medicinal chemistry, and bioactive natural products. Chem. Rev. 2012, 112, 3641–3716. [Google Scholar] [CrossRef]
- Prasad, C. Bioactive cyclic dipeptides. Peptides 1995, 16, 151–164. [Google Scholar] [CrossRef]
- Holden, M.T.; Ram Chhabra, S.; de Nys, R.; Stead, P.; Bainton, N.J.; Hill, P.J.; Manefield, M.; Kumar, N.; Labatte, M.; England, D.; et al. Quorum-sensing cross talk: Isolation and chemical characterization of cyclic dipeptides from Pseudomonas aeruginosa and other gram-negative bacteria. Mol. Microbiol. 1999, 33, 1254–1266. [Google Scholar] [CrossRef]
- Degrassi, G.; Aguilar, C.; Bosco, M.; Zahariev, S.; Pongor, S.; Venturi, V. Plant growth-promoting Pseudomonas putida WCS358 produces and secretes four cyclic dipeptides: Cross-talk with quorum sensing bacterial sensors. Curr. Microbiol. 2002, 45, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Park, D.K.; Lee, K.E.; Baek, C.H.; Kim, I.H.; Kwon, J.H.; Lee, W.K.; Lee, K.H.; Kim, B.S.; Choi, S.H.; Kim, K.S. Cyclo(Phe-Pro) modulates the expression of ompU in Vibrio spp. J. Bacteriol. 2006, 188, 2214–2221. [Google Scholar] [CrossRef]
- Li, J.; Wang, W.; Xu, S.X.; Magarvey, N.A.; McCormick, J.K. Lactobacillus reuteri-produced cyclic dipeptides quench agr-mediated expression of toxic shock syndrome toxin-1 in staphylococci. Proc. Natl. Acad. Sci. USA 2011, 108, 3360–3365. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.; Sommer, R.; Hinsberger, S.; Lu, C.; Hartmann, R.W.; Empting, M.; Titz, A. Novel strategies for the treatment of Pseudomonas aeruginosa infections. J. Med. Chem. 2016, 59, 5929–5969. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Xie, Y.; Zhang, Y.; Liu, J.; Ding, Y.; Wu, M.; Wang, X.; Deng, X. Novel therapeutic strategies for treating Pseudomonas aeruginosa infection. Expert Opin. Drug Discov. 2020, 15, 1403–1423. [Google Scholar] [CrossRef] [PubMed]
- Gambello, M.J.; Iglewski, B.H. Cloning and characterization of the Pseudomonas aeruginosa lasR gene, a transcriptional activator of elastase expression. J. Bacteriol. 1991, 173, 3000–3009. [Google Scholar] [CrossRef] [PubMed]
- Lau, G.W.; Hassett, D.J.; Ran, H.; Kong, F. The role of pyocyanin in Pseudomonas aeruginosa infection. Trends Mol. Med. 2004, 10, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Sun, C.; Zheng, M.; Harmer, J.R.; Yu, M.; Zhang, Y.; Peng, H.; Zhu, D.; Deng, Z.; Chen, S.L.; et al. Efficient biosynthesis of heterodimeric C(3)-aryl pyrroloindoline alkaloids. Nat. Commun. 2018, 9, 4428. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, M.E.; Chuang, K.V.; Reisman, S.E. Copper-catalyzed diastereoselective arylation of tryptophan derivatives: Total synthesis of (+)-naseseazines A and B. J. Am. Chem. Soc. 2013, 135, 5557–5560. [Google Scholar] [CrossRef]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 11th ed.; CLSI standard M07; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Zhu, H.; He, C.C.; Chu, Q.H. Inhibition of quorum sensing in Chromobacterium violaceum by pigments extracted from Auricularia auricular. Lett. Appl. Microbiol. 2011, 52, 269–274. [Google Scholar] [CrossRef]
- Luo, J.; Kong, J.L.; Dong, B.Y.; Huang, H.; Wang, K.; Wu, L.H.; Hou, C.C.; Liang, Y.; Li, B.; Chen, Y.Q. Baicalein attenuates the quorum sensing-controlled virulence factors of Pseudomonas aeruginosa and relieves the inflammatory response in P. aeruginosa-infected macrophages by downregulating the MAPK and NFkappaB signal-transduction pathways. Drug Des. Devel. Ther. 2016, 10, 183–203. [Google Scholar] [CrossRef]
- Das, M.C.; Paul, S.; Gupta, P.; Tribedi, P.; Sarkar, S.; Manna, D.; Bhattacharjee, S. 3-Amino-4-aminoximidofurazan derivatives: Small molecules possessing antimicrobial and antibiofilm activity against Staphylococcus aureus and Pseudomonas aeruginosa. J. Appl. Microbiol. 2016, 120, 842–859. [Google Scholar] [CrossRef]
- Cui, X.; Ruan, X.; Yin, J.; Wang, M.; Li, N.; Shen, D. Regulation of las and rhl quorum sensing on aerobic denitrification in Pseudomonas aeruginosa PAO1. Curr. Microbiol. 2021, 78, 659–667. [Google Scholar] [CrossRef]
- Khan, F.; Jeong, M.C.; Park, S.K.; Kim, S.K.; Kim, Y.M. Contribution of chitooligosaccharides to biofilm formation, antibiotics resistance and disinfectants tolerance of Listeria monocytogenes. Microb. Pathog. 2019, 136, 103673. [Google Scholar] [CrossRef]
- Zhao, Y.; Yu, C.; Yu, Y.; Wei, X.; Duan, X.; Dai, X.; Zhang, X. Bioinspired heteromultivalent ligand-decorated nanotherapeutic for enhanced photothermal and photodynamic therapy of antibiotic-resistant bacterial pneumonia. ACS Appl. Mater. Interfaces 2019, 11, 39648–39661. [Google Scholar] [CrossRef]
- Rahman, M.A.; Bam, M.; Luat, E.; Jui, M.S.; Ganewatta, M.S.; Shokfai, T.; Nagarkatti, M.; Decho, A.W.; Tang, C. Macromolecular-clustered facial amphiphilic antimicrobials. Nat Commun 2018, 9, 5231. [Google Scholar] [CrossRef]
- Ouyang, J.; Sun, F.; Feng, W.; Sun, Y.; Qiu, X.; Xiong, L.; Liu, Y.; Chen, Y. Quercetin is an effective inhibitor of quorum sensing, biofilm formation and virulence factors in Pseudomonas aeruginosa. J. Appl. Microbiol. 2016, 120, 966–974. [Google Scholar] [CrossRef]
- Ghilini, F.; Rodriguez Gonzalez, M.C.; Minan, A.G.; Pissinis, D.; Creus, A.H.; Salvarezza, R.C.; Schilardi, P.L. Highly stabilized nanoparticles on poly-l-lysine-coated oxidized metals: A versatile platform with enhanced antimicrobial activity. ACS Appl. Mater. Interfaces 2018, 10, 23657–23666. [Google Scholar] [CrossRef]
- Hanelt, M.; Gareis, M.; Kollarczik, B. Cytotoxicity of mycotoxins evaluated by the MTT-cell culture assay. Mycopathologia 1994, 128, 167–174. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zheng, Q.; Li, L.; Pan, L.; Zhu, H. Anti-Quorum-Sensing Activity of Tryptophan-Containing Cyclic Dipeptides. Mar. Drugs 2022, 20, 85. https://doi.org/10.3390/md20020085
Wang Y, Zheng Q, Li L, Pan L, Zhu H. Anti-Quorum-Sensing Activity of Tryptophan-Containing Cyclic Dipeptides. Marine Drugs. 2022; 20(2):85. https://doi.org/10.3390/md20020085
Chicago/Turabian StyleWang, Yinglu, Qian Zheng, Li Li, Lile Pan, and Hu Zhu. 2022. "Anti-Quorum-Sensing Activity of Tryptophan-Containing Cyclic Dipeptides" Marine Drugs 20, no. 2: 85. https://doi.org/10.3390/md20020085
APA StyleWang, Y., Zheng, Q., Li, L., Pan, L., & Zhu, H. (2022). Anti-Quorum-Sensing Activity of Tryptophan-Containing Cyclic Dipeptides. Marine Drugs, 20(2), 85. https://doi.org/10.3390/md20020085