
Two Benthic Diatoms, Nanofrustulum shiloi and Striatella unipunctata, Encapsulated in Alginate Beads, Influence the Reproductive Efficiency of Paracentrotus lividus by Modulating the Gene Expression







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Species Identification by Morphological and Molecular Analyses
2.2. Diatom’s Encapsulation
2.3. Effects of Feeding Tests on Sea Urchin Progeny
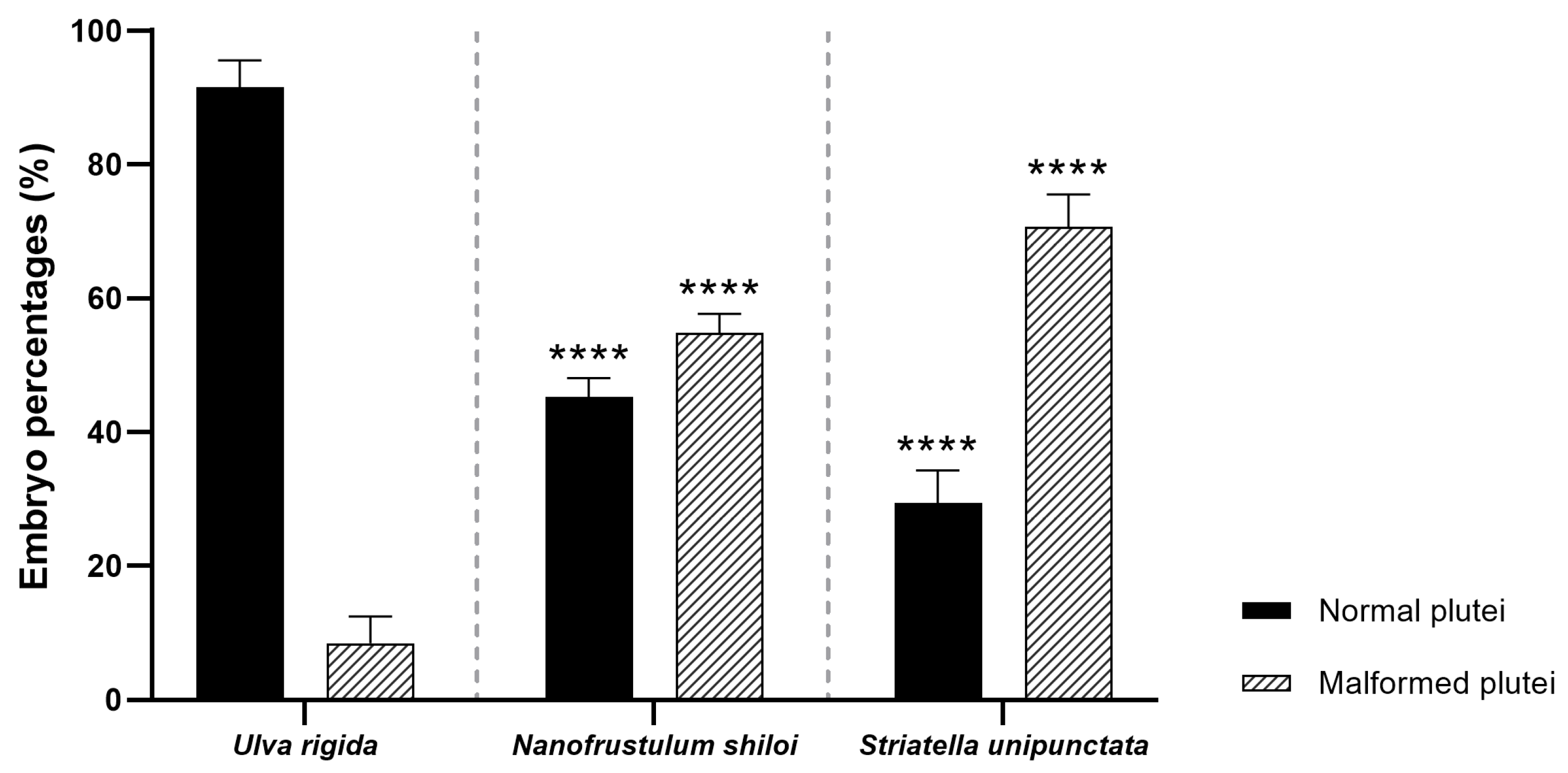
2.3.1. Fertilization/Cleavage Rates and Embryo Development
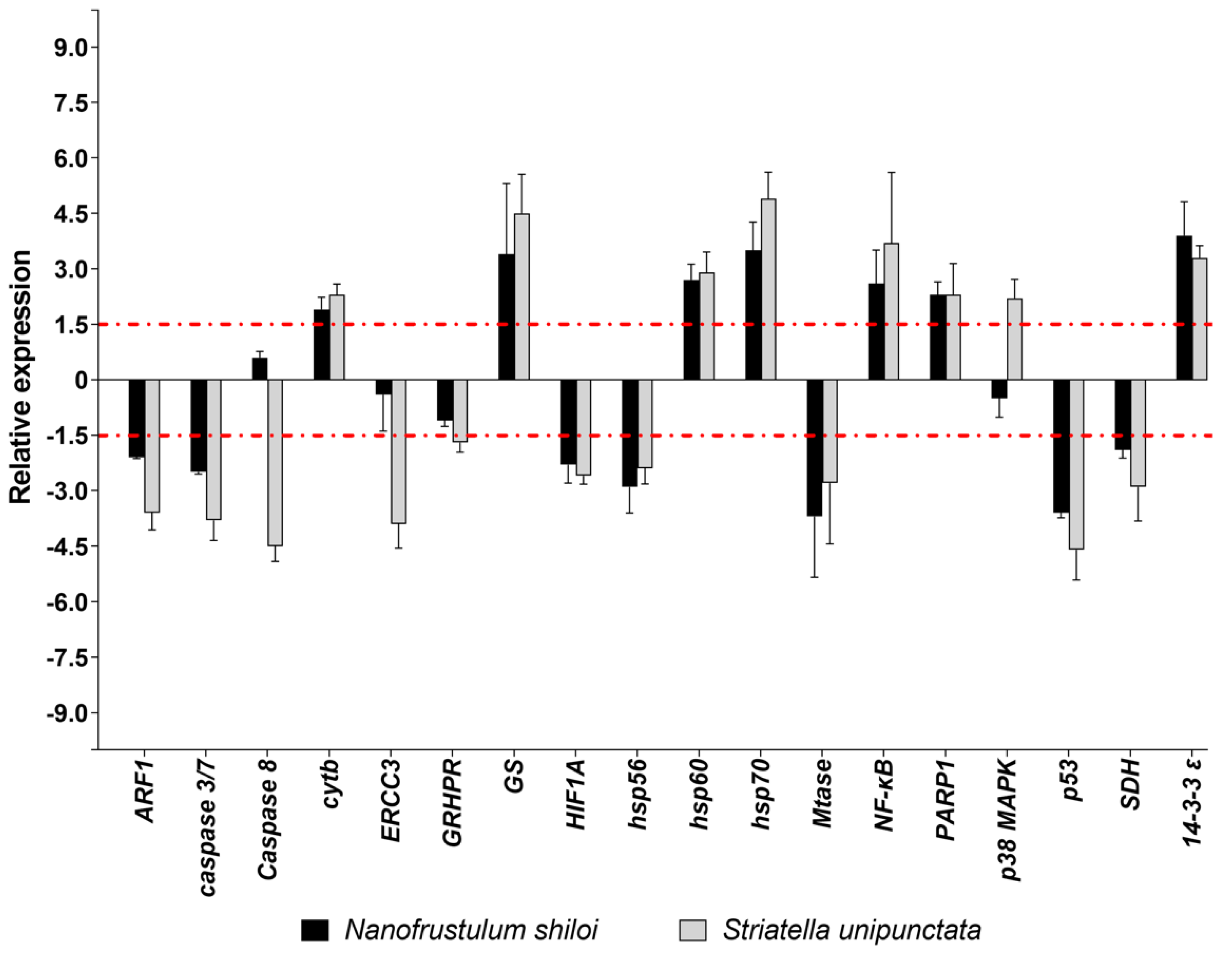
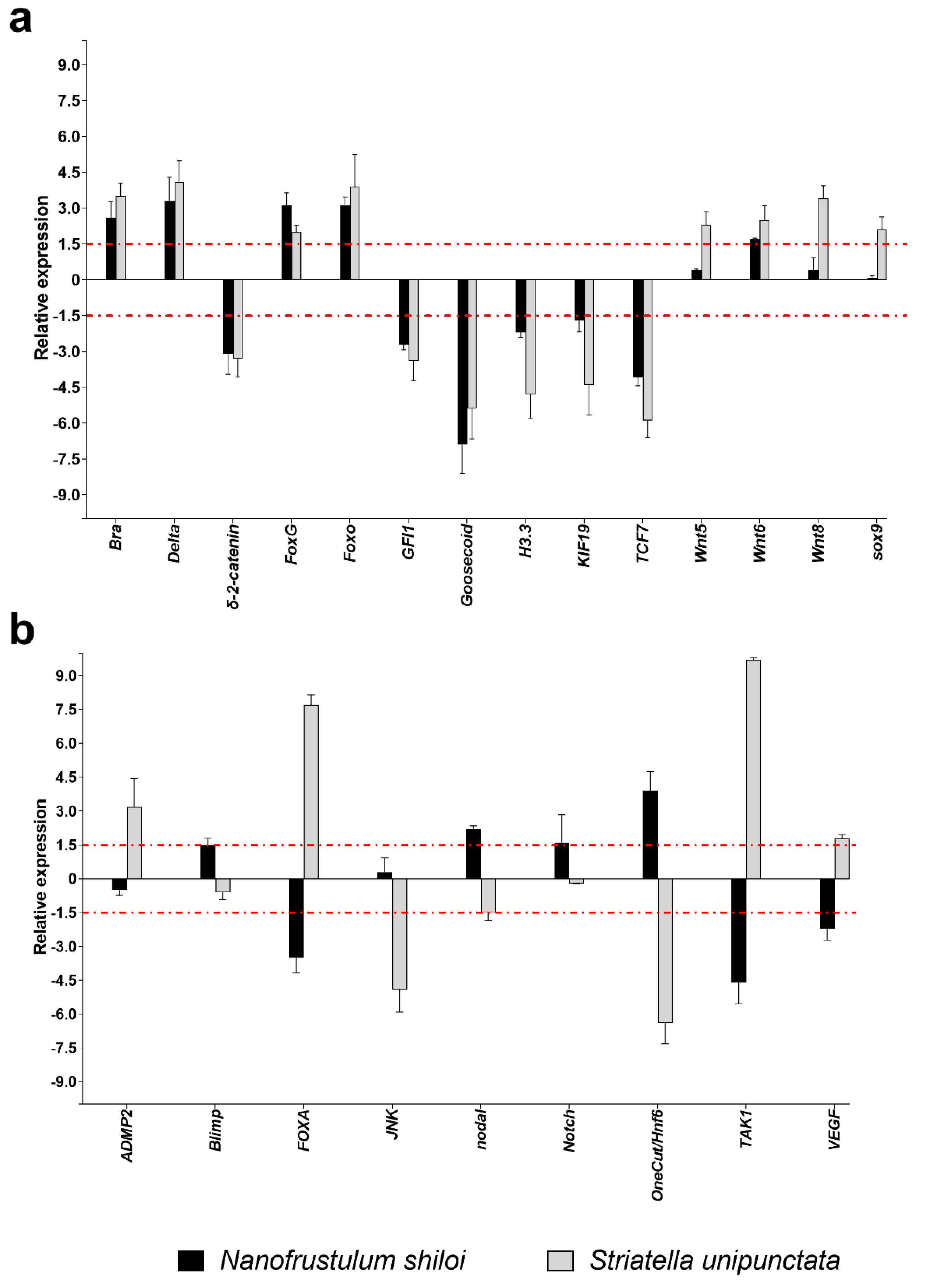
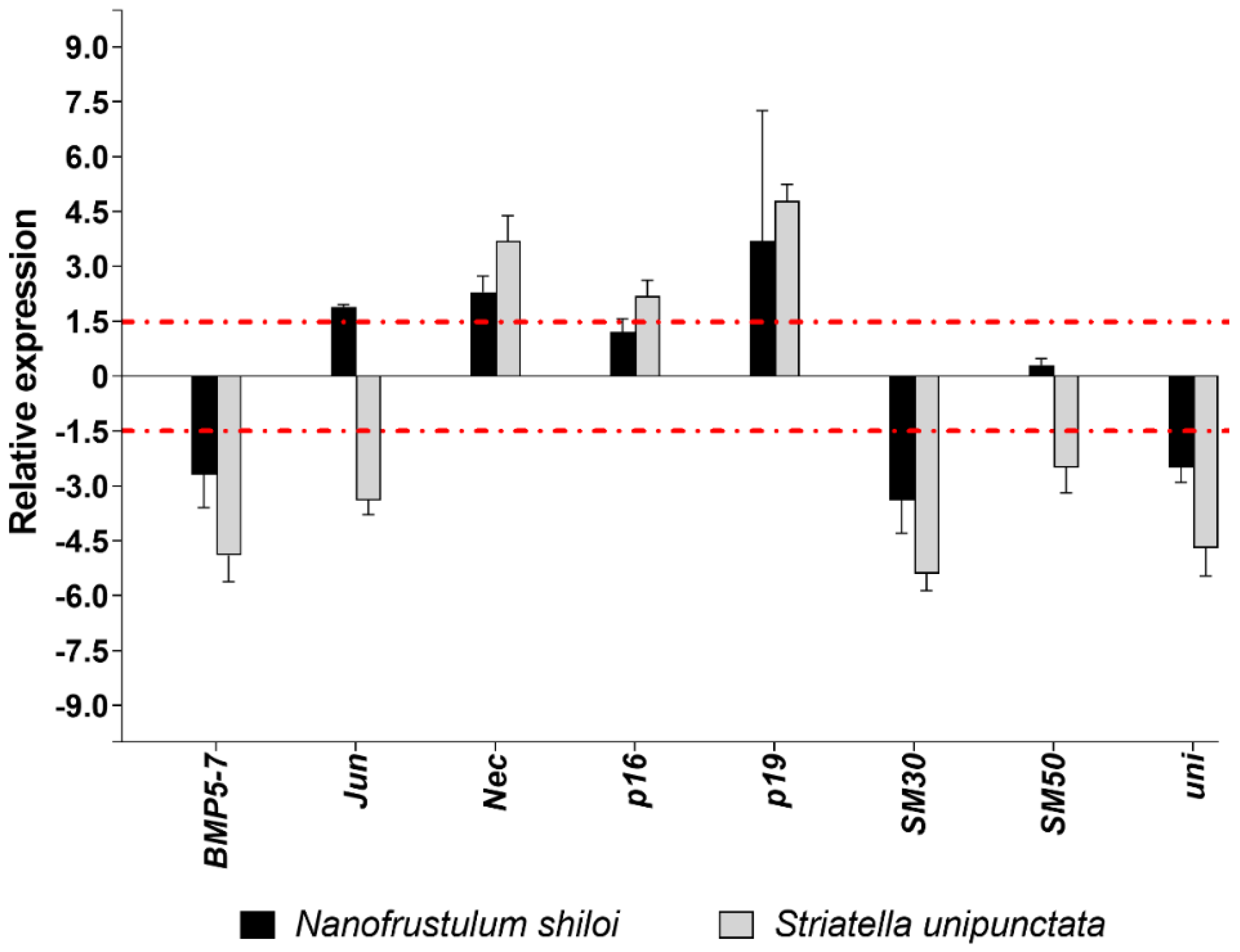
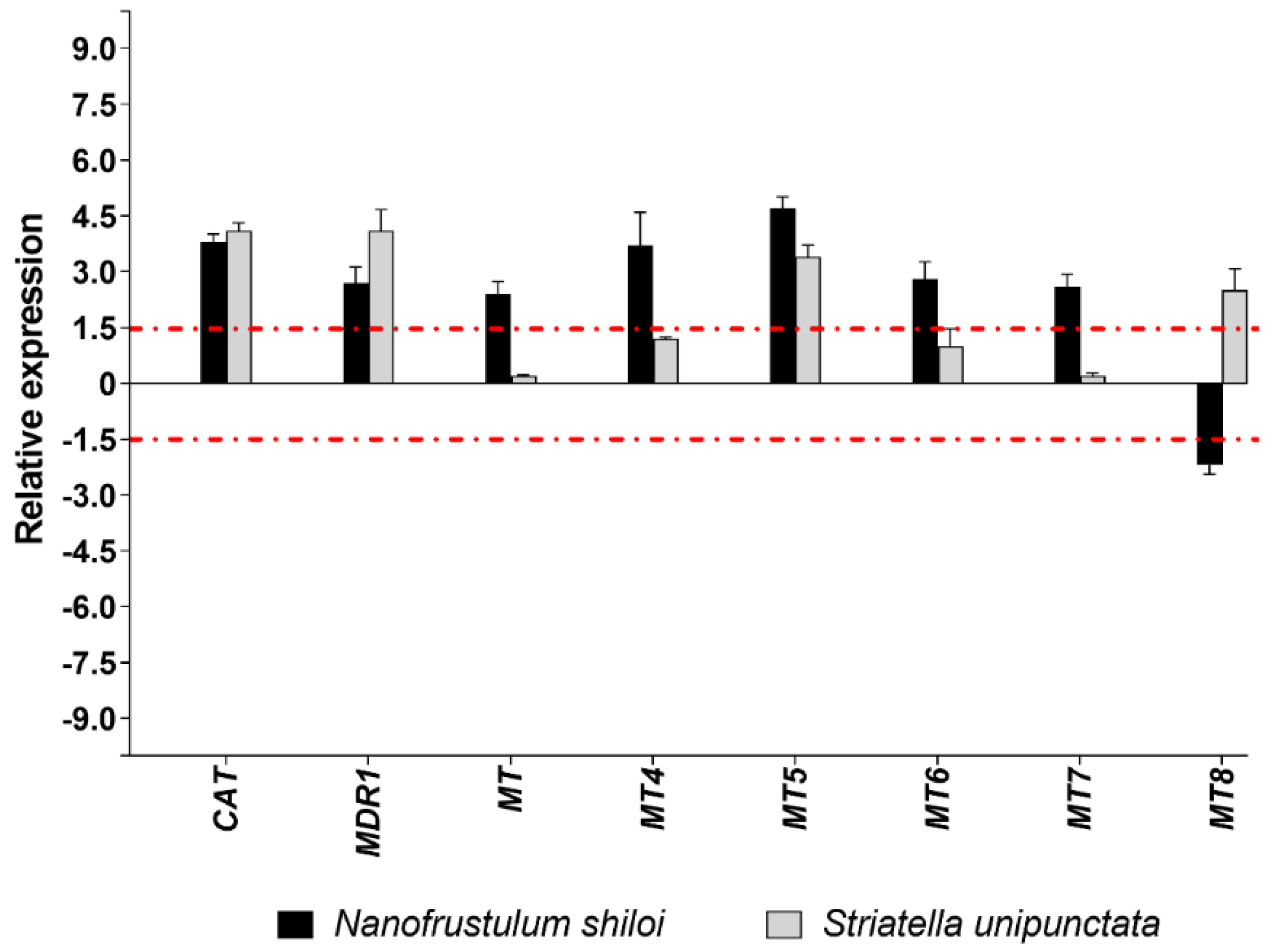
2.3.2. Gene Expression by Real Time (RT)-qPCR
3. Discussion
3.1. Effects of Feeding Tests by Morphological Observations
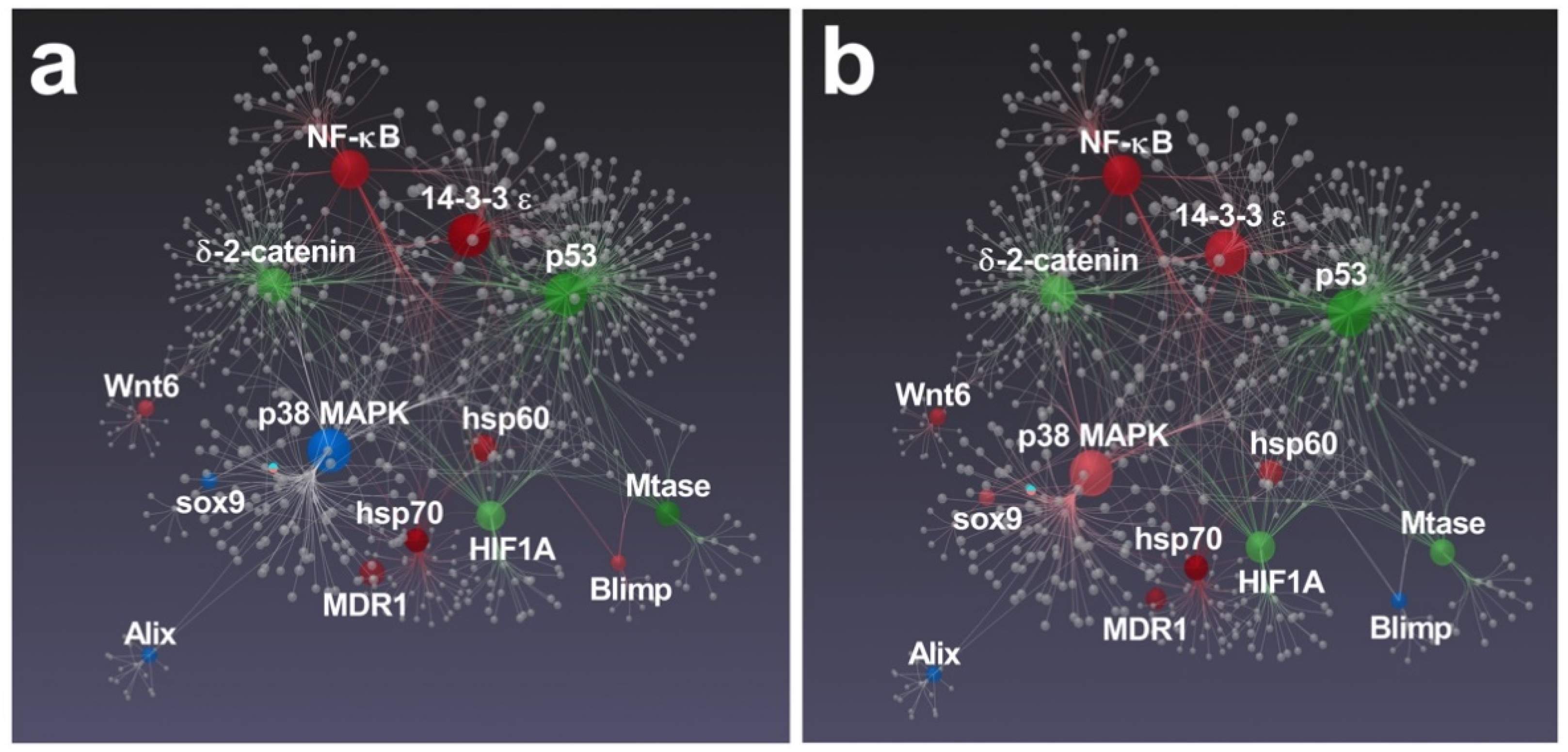
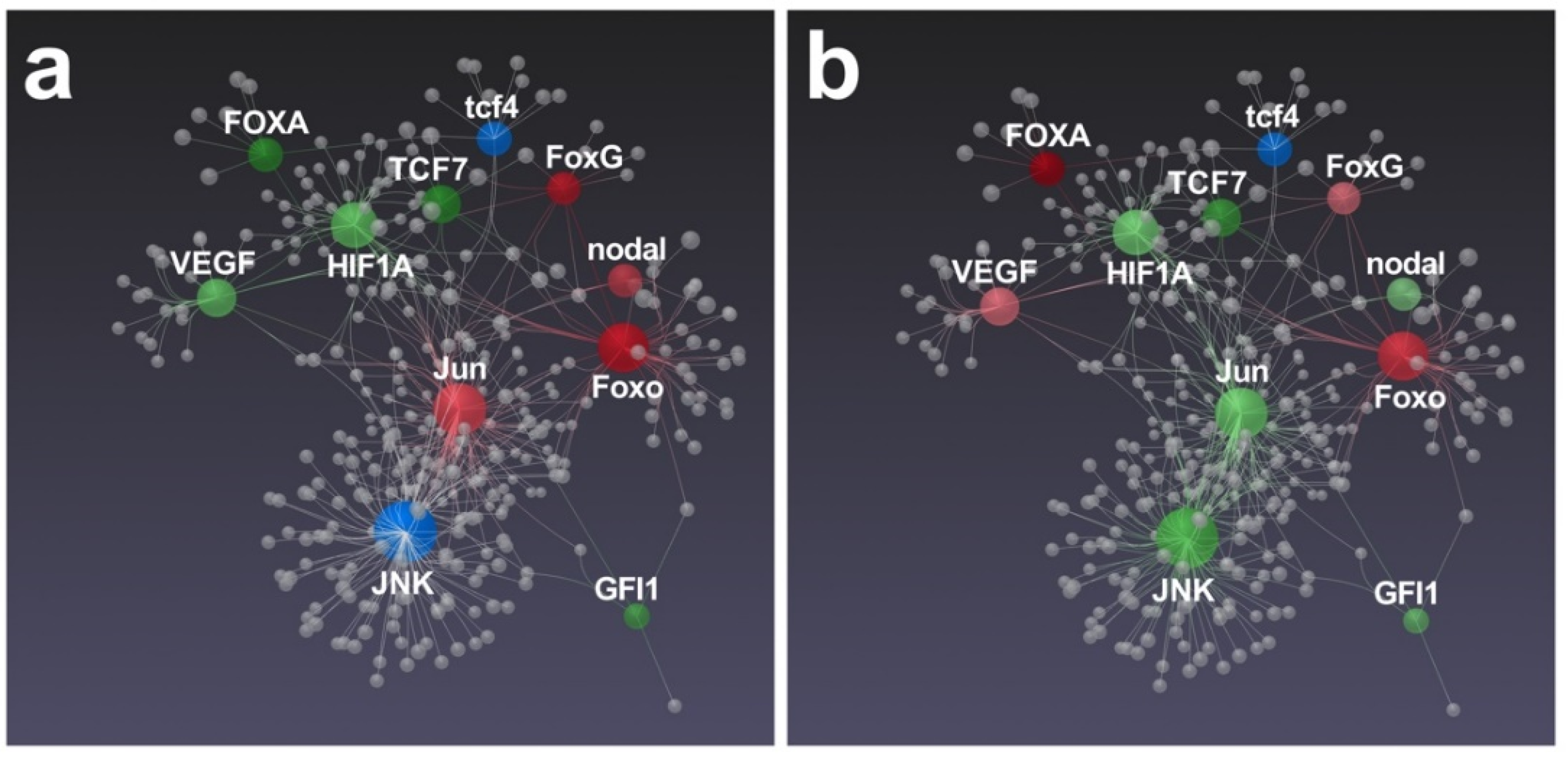
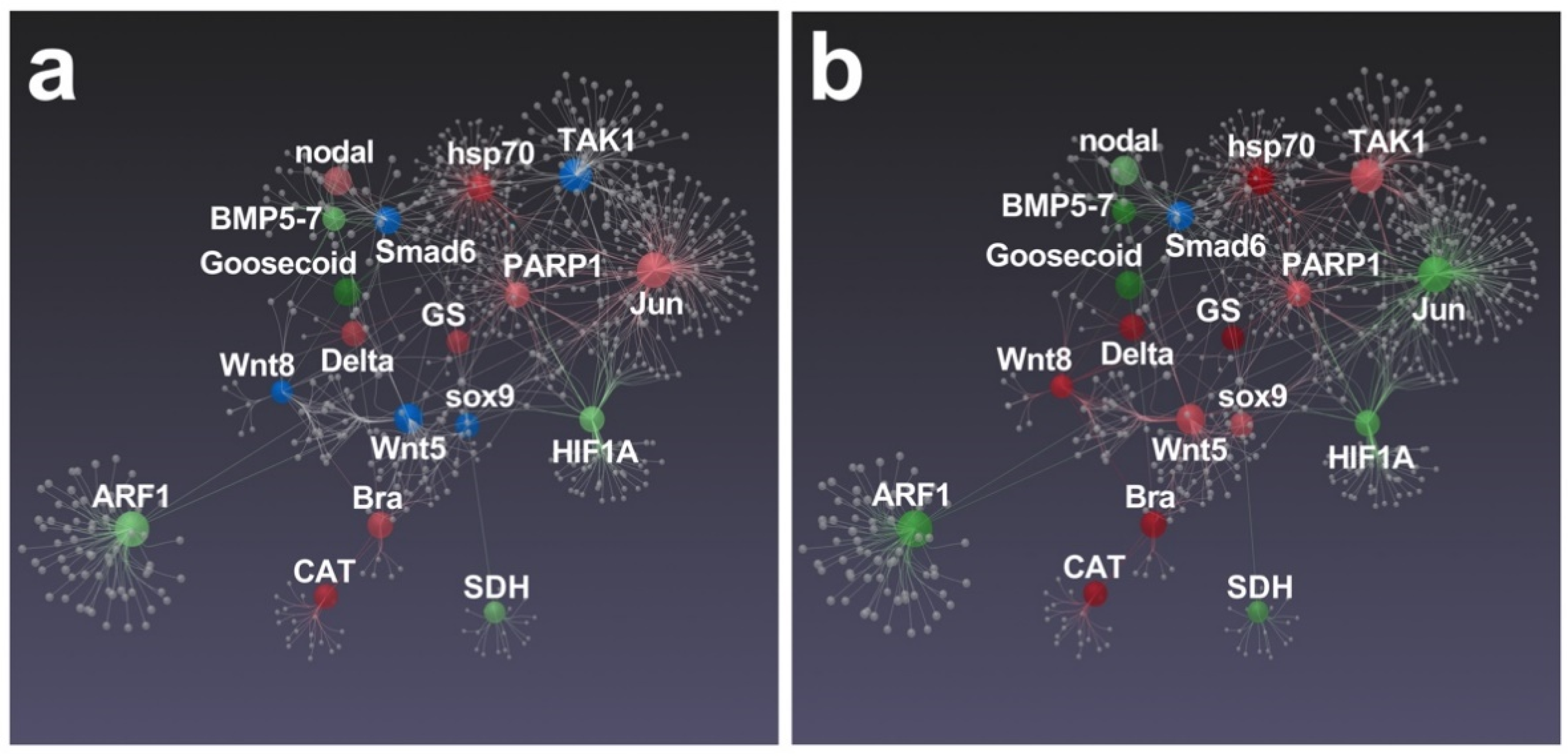
3.2. Effects of Feeding Tests on Gene Pathways
4. Materials and Methods
4.1. Ethics Statement
4.2. Isolation of S. unipunctata and Culturing
4.3. Species Characterization
4.3.1. Morphological Analysis of Frustules
4.3.2. Molecular Identification of Diatom Species
4.4. Diatom Encapsulation in Alginate Beads
4.5. Feeding, Gametes Collection, Evaluation of Fertilization/Cleavage Success and Detection of Abnormal Plutei
4.6. Molecular Analysis on Sea Urchin Plutei
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miralto, A.; Barone, G.; Romano, G.; Poulet, S.A.; Ianora, A.; Russo, G.L.; Buttino, I.; Mazzarella, G.; Laablr, M.; Cabrini, M.; et al. The insidious effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar] [CrossRef]
- Zupo, V. Effect of microalgal food on the sex reversal of Hippolyte inermis (Crustacea: Decapoda). Mar. Ecol. Prog. Ser. 2000, 201, 251–259. [Google Scholar] [CrossRef]
- Zupo, V. Strategies of sexual inversion in Hippolyte inermis Leach (Crustacea, Decapoda) from a Mediterranean seagrass meadow. J. Exp. Mar. Bio. Ecol. 1994, 178, 131–145. [Google Scholar] [CrossRef]
- Mutalipassi, M.; Maibam, C.; Zupo, V. The sex change of the caridean shrimp Hippolyte inermis Leach: Temporal development of the gonopore morphology. Zoomorphology 2018, 137, 377–388. [Google Scholar] [CrossRef]
- Estévez-Calvar, N.; Canesi, L.; Montagna, M.; Faimali, M.; Piazza, V.; Garaventa, F. Adverse effects of the SSRI antidepressant sertraline on early life stages of marine invertebrates. Mar. Environ. Res. 2017, 128, 88–97. [Google Scholar] [CrossRef]
- Maibam, C.; Fink, P.; Romano, G.; Buia, M.C.; Gambi, M.C.; Scipione, M.B.; Patti, F.P.; Lorenti, M.; Butera, E.; Zupo, V. Relevance of wound-activated compounds produced by diatoms as toxins and infochemicals for benthic invertebrates. Mar. Biol. 2014, 161, 1639–1652. [Google Scholar] [CrossRef]
- Peckol, P.; Putnam, A.B. Differential toxic effects of Ulva lactuca (Chlorophyta) on the herbivorous gastropods, Littorina littorea and L. obtusata (Mollusca). J. Phycol. 2017, 53, 361–367. [Google Scholar] [CrossRef]
- Ruocco, N.; Costantini, S.; Zupo, V.; Lauritano, C.; Caramiello, D.; Ianora, A.; Budillon, A.; Romano, G.; Nuzzo, G.; D’Ippolito, G.; et al. Toxigenic effects of two benthic diatoms upon grazing activity of the sea urchin: Morphological, metabolomic and de novo transcriptomic analysis. Sci. Rep. 2018, 8, 5622. [Google Scholar] [CrossRef]
- Ruocco, N.; Cavaccini, V.; Caramiello, D.; Ianora, A.; Fontana, A. Noxious effects of the benthic diatoms Cocconeis scutellum and Diploneis sp. on sea urchin development: Morphological and de novo transcriptomic analysis. Harmful Algae 2019, 86, 64–73. [Google Scholar] [CrossRef]
- Johnson, L.E.; Paine, R.L. Consistency in a marine algal-grazer interaction over multiple scales. J. Phycol. 2016, 52, 942–950. [Google Scholar] [CrossRef]
- Zupo, V.; Maibam, C.; Buia, M.C.; Gambi, M.C.; Patti, F.P.; Scipione, M.B.; Lorenti, M.; Fink, P. Chemoreception of the seagrass Posidonia oceanica by benthic Invertebrates is altered by seawater acidification. J. Chem. Ecol. 2015, 41, 766–779. [Google Scholar] [CrossRef]
- Jüttner, F.; Messina, P.; Patalano, C.; Zupo, V. Odour compounds of the diatom Cocconeis scutellum: Effects on benthic herbivores living on Posidonia oceanica. Mar. Ecol. Prog. Ser. 2010, 400, 63–73. [Google Scholar] [CrossRef]
- Mutalipassi, M.; Fink, P.; Maibam, C.; Porzio, L.; Buia, M.C.; Gambi, M.C.; Patti, F.P.; Scipione, M.B.; Lorenti, M.; Zupo, V. Ocean acidification alters the responses of invertebrates to wound-activated infochemicals produced by epiphytes of the seagrass Posidonia oceanica. J. Exp. Mar. Bio. Ecol. 2020, 530–531, 151435. [Google Scholar] [CrossRef]
- Cutignano, A.; Nuzzo, G.; Ianora, A.; Luongo, E.; Romano, G.; Gallo, C.; Sansone, C.; Aprea, S.; Mancini, F.; D’Oro, U.; et al. Development and application of a novel SPE-method for bioassay-guided fractionation of marine extracts. Mar. Drugs 2015, 13, 5736–5749. [Google Scholar] [CrossRef] [PubMed]
- Corral, F.M.; Vilariño, N.; Louzao, M.C.; Botana, L.M. Sensitivity improvement of an immuno-detection method for azaspiracids based on the use of microspheres coupled to a flow-fluorimetry system. Front. Mar. Sci. 2014, 1. [Google Scholar] [CrossRef]
- Gackowska, A.; Studziński, W.; Kudlek, E.; Dudziak, M.; Gaca, J. Estimation of physicochemical properties of 2-ethylhexyl-4-methoxycinnamate (EHMC) degradation products and their toxicological evaluation. Environ. Sci. Pollut. Res. 2018, 25, 16037–16049. [Google Scholar] [CrossRef]
- Rodríguez, L.P.; Vilariño, N.; Louzao, M.C.; Dickerson, T.J.; Nicolaou, K.C.; Frederick, M.O.; Botana, L.M. Microsphere-based Immunoassay for the detection of azaspiracids. Anal. Biochem. 2014, 447, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Kelly, M.S.; Cook, E.J.; Black, K.; Orr, H.; Zhu, J.X.; Dong, S.L. The effect of diet type on growth and fatty-acid composition of sea urchin larvae, I. Paracentrotus lividus (Lamarck, 1816) (Echinodermata). Aquaculture 2007, 264, 247–262. [Google Scholar] [CrossRef]
- Liu, H.; Kelly, M.S.; Cook, E.J.; Black, K.; Orr, H.; Zhu, J.X.; Dong, S.L. The effect of diet type on growth and fatty acid composition of the sea urchin larvae, II. Psammechinus miliaris (Gmelin). Aquaculture 2007, 264, 263–278. [Google Scholar] [CrossRef]
- Laurienzo, P. Marine polysaccharides in pharmaceutical applications: An overview. Mar. Drugs 2010, 8, 2435–2465. [Google Scholar] [CrossRef]
- Laurienzo, P.; Malinconico, M.; Mattia, G.; Russo, R.; La Rotonda, M.I.; Quaglia, F.; Capitani, D.; Mannina, L. Novel alginate—acrylic polymers as a platform for drug delivery. J. Biomed. Mater. Res. Part A 2006, 79, 963–973. [Google Scholar] [CrossRef]
- Guilherme, M.R.; Reis, A.V.; Paulino, A.T.; Fajardo, A.R.; Muniz, E.C.; Tambourgi, E.B. Superabsorbent hydrogel based on modified polysaccharide for removal of Pb2+ and Cu2+ from water with excellent performance. J. Appl. Polym. Sci. 2007, 105, 2903–2909. [Google Scholar] [CrossRef]
- Pourjavadi, A.; Soleyman, R.; Bardajee, G.R.; Ghavami, S. Novel superabsorbent hydrogel based on natural hybrid backbone: Optimized synthesis and its swelling behavior. Bull. Korean Chem. Soc. 2009, 30, 2680–2686. [Google Scholar] [CrossRef]
- Buttino, I.; De Rosa, G.; Carotenuto, Y.; Ianora, A.; Fontana, A.; Quaglia, F.; La Rotonda, M.I.; Miralto, A. Giant liposomes as delivery system for ecophysiological studies in copepods. J. Exp. Biol. 2006, 209, 801–809. [Google Scholar] [CrossRef][Green Version]
- Song, W.; Lima, A.C.; Mano, J.F. Bioinspired methodology to fabricate hydrogel spheres for multi-applications using superhydrophobic substrates. Soft Matter 2010, 6, 5868–5871. [Google Scholar] [CrossRef]
- Costa, A.M.S.; Alatorre-Meda, M.; Oliveira, N.M.; Mano, J.F. Biocompatible polymeric microparticles produced by a simple biomimetic approach. Langmuir 2014, 30, 4535–4539. [Google Scholar] [CrossRef]
- Macintyre, H.L.; Stutes, A.L.; Smith, W.L.; Dorsey, C.P.; Annabraham, A.; Dickey, R.W. Environmental correlates of community composition and toxicity during a bloom of Pseudo-nitzschia spp. in the northern Gulf of Mexico. J. Plankton Res. 2011, 33, 273–295. [Google Scholar] [CrossRef]
- Nenadović, T.; Šarčević, T.; Čižmek, H.; Godrijan, J.; Pfannkuchen, D.M.; Pfannkuchen, M.; Ljubešić, Z. Development of periphytic diatoms on different artificial substrates in the Eastern Adriatic Sea. Acta Bot. Croat. 2015, 74, 377–392. [Google Scholar] [CrossRef]
- Lok, A.; Metin, G.; Acarli, S.; Goulletquer, P. Harmful algal blooms (HABs) and black mussel Mytilus galloprovincialis (Linnaeus, 1758) culture in Izmir Bay (Iskele-Urla)-Turkey: Preliminary results on the annual feeding cycle using a qualitative approach. Turk. J. Fish. Aquat. Sci. 2010, 10, 527–536. [Google Scholar] [CrossRef]
- Tan, T.H.; Pin Leaw, C.; Chee, S.; Leong, Y.; Lim, L.P.; Chew, S.M.; Teng, S.T.; Lim, P.T. Marine micro-phytoplankton of Singapore, with a review of harmful microalgae in the region. Raffles Bull. Zool. 2016, 34, 78–96. [Google Scholar]
- Mabrouk, L.; Ben, B.M.; Hamza, A.; Mahfoudhi, M.; Bradai, M.N. A comparison of abundance and diversity of epiphytic microalgal assemblages on the leaves of the seagrasses Posidonia oceanica (L.) and Cymodocea nodosa (Ucria) asch in Eastern Tunisia. J. Mar. Biol. 2014, 2014, 1–10. [Google Scholar] [CrossRef]
- Kanjer, L.; Mucko, M.; Car, A.; Bosak, S. Epiphytic diatoms on Posidonia oceanica (L.) Delile leaves from eastern Adriatic Sea. Nat. Croat. 2019, 28, 1–20. [Google Scholar] [CrossRef]
- Mazzella, L.; Spinoccia, L. Epiphytic diatoms of leaf blades of the mediterranean seagrass Posidonia oceanica (L.) delile. G. Bot. Ital. 1992, 126, 752–754. [Google Scholar] [CrossRef]
- de Stefano, M.; Marino, D.; Mazzella, L. Marine taxa of Cocconeis on leaves of Posidonia oceanica, including a new species and two new varieties. Eur. J. Phycol. 2000, 35, 225–242. [Google Scholar] [CrossRef]
- Varrella, S.; Romano, G.; Ianora, A.; Bentley, M.G.; Ruocco, N.; Costantini, M. Molecular response to toxic diatom-derived aldehydes in the sea urchin Paracentrotus lividus. Mar. Drugs 2014, 12, 2089–2113. [Google Scholar] [CrossRef]
- Ruocco, N.; Nuzzo, G.; D’Ippolito, G.; Manzo, E.; Sardo, A.; Ianora, A.; Romano, G.; Iuliano, A.; Zupo, V.; Costantini, M.; et al. Lipoxygenase pathways in diatoms: Occurrence and correlation with grazer toxicity in four benthic diatoms. Mar. Drugs 2020, 18, 66. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, R.; Zou, L.; McClements, D.J. Protein encapsulation in alginate hydrogel beads: Effect of pH on microgel stability, protein retention and protein release. Food Hydrocoll. 2016, 58, 308–315. [Google Scholar] [CrossRef]
- Santagapita, P.R.; Mazzobre, M.F.; Buera, M.P. Formulation and drying of alginate beads for controlled release and stabilization of invertase. Biomacromolecules 2011, 12, 3147–3155. [Google Scholar] [CrossRef]
- Gombotz, W.R.; Wee, S.F. Protein release from alginate matrices. Adv. Drug Deliv. Rev. 1998, 31, 267–285. [Google Scholar] [CrossRef]
- Calvo, T.A.; Santagapita, P. Physicochemical characterization of alginate beads containing sugars and biopolymers. J. Qual. Reliab. Eng. 2016, 2016, 1–7. [Google Scholar] [CrossRef]
- Varrella, S.; Romano, G.; Costantini, S.; Ruocco, N.; Ianora, A.; Bentley, M.G.; Costantini, M. Toxic diatom aldehydes affect defence gene networks in sea urchins. PLoS ONE 2016, 11, e0149734. [Google Scholar] [CrossRef] [PubMed]
- Ruocco, N.; Maria Fedele, A.; Costantini, S.; Romano, G.; Ianora, A.; Costantini, M. New inter-correlated genes targeted by diatom-derived polyunsaturated aldehydes in the sea urchin Paracentrotus lividus. Ecotoxicol. Environ. Saf. 2017, 142, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Esposito, R.; Ruocco, N.; Albarano, L.; Ianora, A.; Manfra, L.; Libralato, G.; Costantini, M. Combined effects of diatom-derived oxylipins on the sea urchin Paracentrotus lividus. Int. J. Mol. Sci. 2020, 21, 719. [Google Scholar] [CrossRef] [PubMed]
- Bonaventura, R.; Poma, V.; Russo, R.; Zito, F.; Matranga, V. Effects of UV-B radiation on development and hsp70 expression in sea urchin cleavage embryos. Mar. Biol. 2006, 149, 79–86. [Google Scholar] [CrossRef]
- Russo, R.; Bonaventura, R.; Matranga, V. Time- and dose-dependent gene expression in sea urchin embryos exposed to UVB. Mar. Environ. Res. 2014, 93, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Pinsino, A.; Bergami, E.; Della Torre, C.; Vannuccini, M.L.; Addis, P.; Secci, M.; Dawson, K.A.; Matranga, V.; Corsi, I. Amino-modified polystyrene nanoparticles affect signalling pathways of the sea urchin (Paracentrotus lividus) embryos. Nanotoxicology 2017, 11, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Reinardy, H.C.; Chapman, J.; Bodnar, A.G. Induction of innate immune gene expression following methyl methanesulfonate-induced DNA damage in sea urchins. Biol. Lett. 2016, 12, 20151057. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Pinsino, A.; Costa, C.; Bonaventura, R.; Matranga, V.; Zito, F. The newly characterized Pl-jun is specifically expressed in skeletogenic cells of the Paracentrotus lividus sea urchin embryo. FEBS J. 2014, 281, 3828–3843. [Google Scholar] [CrossRef] [PubMed]
- Long, J.T.; Irwin, L.; Enomoto, A.C.; Grow, Z.; Ranck, J.; Peeler, M.T. Jun N-terminal kinase activity is required for invagination but not differentiation of the sea urchin archenteron. Genesis 2015, 53, 762–769. [Google Scholar] [CrossRef]
- Morgulis, M.; Gildor, T.; Roopin, M.; Sher, N.; Malik, A.; Lalzar, M.; Dines, M.; De-Leon, S.B.T.; Khalaily, L.; De-Leon, S.B.T. Possible cooption of a VEGF-driven tubulogenesis program for biomineralization in echinoderms. Proc. Natl. Acad. Sci. USA 2019, 116, 12353–12362. [Google Scholar] [CrossRef]
- Layous, M.; Khalaily, L.; Gildor, T.; De-Leon, S.B.T. The tolerance to hypoxia is defined by a time-sensitive response of the gene regulatory network in sea urchin embryos. Biorxiv 2020, 1–25. [Google Scholar] [CrossRef]
- Chang, W.L.; Chang, Y.C.; Lin, K.T.; Li, H.R.; Pai, C.Y.; Chen, J.H.; Su, Y.H. Asymmetric distribution of hypoxia-inducible factor α regulates dorsoventral axis establishment in the early sea urchin embryo. Development 2017, 144, 2940–2950. [Google Scholar] [CrossRef]
- Reinardy, H.C.; Bodnar, A.G. Profiling DNA damage and repair capacity in sea urchin larvae and coelomocytes exposed to genotoxicants. Mutagenesis 2015, 30, 829–839. [Google Scholar] [CrossRef]
- Coffman, J.A.; Coluccio, A.; Planchart, A.; Robertson, A.J. Oral-aboral axis specification in the sea urchin embryo III. Role of mitochondrial redox signaling via H2O2. Dev. Biol. 2009, 330, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Saudemont, A.; Haillot, E.; Mekpoh, F.; Bessodes, N.; Quirin, M.; Ro, E.; Wincker, P.; Lepage, T. Ancestral regulatory circuits governing ectoderm patterning downstream of nodal and BMP2/4 revealed by Gene Regulatory Network analysis in an echinoderm. PLoS Genet. 2010, 6, e1001259. [Google Scholar] [CrossRef] [PubMed]
- Duboc, V.; Röttinger, E.; Lapraz, F.; Besnardeau, L.; Lepage, T. Left-right asymmetry in the sea urchin embryo Is regulated by nodal signaling on the right side. Dev. Cell 2005, 9, 147–158. [Google Scholar] [CrossRef]
- Molina, M.D.; de Crozé, N.; Haillot, E.; Lepage, T. Nodal: Master and commander of the dorsal-ventral and left-right axes in the sea urchin embryo. Curr. Opin. Genet. Dev. 2013, 23, 445–453. [Google Scholar] [CrossRef]
- Croce, J.; Lhomond, G.; Gache, C. Expression pattern of Brachyury in the embryo of the sea urchin Paracentrotus lividus. Dev. Genes Evol. 2001, 211, 617–619. [Google Scholar] [CrossRef] [PubMed]
- Gross, J.M.; McClay, D.R. The role of Brachyury (T) during gastrulation movements in the sea urchin Lytechinus variegatus. Dev. Biol. 2001, 239, 132–147. [Google Scholar] [CrossRef]
- Ben Tabou de-Leon, S.; Davidson, E.H. Experimentally based sea urchin gene regulatory network and the causal explanation of developmental phenomenology. Wiley Interdiscip. Rev. Syst. Biol. Med. 2009, 1, 237–246. [Google Scholar] [CrossRef]
- Cui, M.; Siriwon, N.; Li, E.; Davidson, E.H.; Peter, I.S. Specific functions of the Wnt signaling system in gene regulatory networks throughout the early sea urchin embryo. Proc. Natl. Acad. Sci. USA 2014, 111, E5029–E5038. [Google Scholar] [CrossRef]
- Erkenbrack, E.M. Divergence of ectodermal and mesodermal gene regulatory network linkages in early development of sea urchins. Proc. Natl. Acad. Sci. USA 2016, 113, E7202–E7211. [Google Scholar] [CrossRef]
- Materna, S.C.; Davidson, E.H. A comprehensive analysis of Delta signaling in pre-gastrular sea urchin embryos. Dev. Biol. 2012, 364, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Kooistra, W.H.C.F.; De Stefano, M.; Mann, D.G.; Salma, N.; Medlin, L.K. Phylogenetic position of Toxarium, a pennate-like lineage within centric diatoms (Bacillariophyceae). J. Phycol. 2003, 39, 185–197. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef]
- Menden-Deuer, S.; Lessard, E.J. Carbon to volume relationships for dinoflagellates, diatoms, and other protist plankton. Limnol. Oceanogr. 2000, 45, 569–579. [Google Scholar] [CrossRef]
- Fabbrocini, A.; Volpe, M.G.; di Stasio, M.; D’Adamo, R.; Maurizio, D.; Coccia, E.; Paolucci, M. Agar-based pellets as feed for sea urchins (Paracentrotus lividus): Rheological behaviour, digestive enzymes and gonad growth. Aquac. Res. 2012, 43, 321–331. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Romano, G.; Costantini, M.; Buttino, I.; Ianora, A.; Palumbo, A. Nitric oxide mediates the stress response induced by diatom aldehydes in the sea urchin Paracentrotus lividus. PLoS ONE 2011, 6, e25980. [Google Scholar] [CrossRef] [PubMed]
- Ragusa, M.A.; Costa, S.; Gianguzza, M.; Roccheri, M.C.; Gianguzza, F. Effects of cadmium exposure on sea urchin development assessed by SSH and RT-qPCR: Metallothionein genes and their differential induction. Mol. Biol. Rep. 2013, 40, 2157–2167. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Soufan, O.; Ewald, J.; Hancock, R.E.W.; Basu, N.; Xia, J. NetworkAnalyst 3.0: A visual analytics platform for comprehensive gene expression profiling and meta-analysis. Nucleic Acids Res. 2019, 47, W234–W241. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glaviano, F.; Ruocco, N.; Somma, E.; De Rosa, G.; Campani, V.; Ametrano, P.; Caramiello, D.; Costantini, M.; Zupo, V. Two Benthic Diatoms, Nanofrustulum shiloi and Striatella unipunctata, Encapsulated in Alginate Beads, Influence the Reproductive Efficiency of Paracentrotus lividus by Modulating the Gene Expression. Mar. Drugs 2021, 19, 230. https://doi.org/10.3390/md19040230
Glaviano F, Ruocco N, Somma E, De Rosa G, Campani V, Ametrano P, Caramiello D, Costantini M, Zupo V. Two Benthic Diatoms, Nanofrustulum shiloi and Striatella unipunctata, Encapsulated in Alginate Beads, Influence the Reproductive Efficiency of Paracentrotus lividus by Modulating the Gene Expression. Marine Drugs. 2021; 19(4):230. https://doi.org/10.3390/md19040230
Chicago/Turabian StyleGlaviano, Francesca, Nadia Ruocco, Emanuele Somma, Giuseppe De Rosa, Virginia Campani, Pasquale Ametrano, Davide Caramiello, Maria Costantini, and Valerio Zupo. 2021. "Two Benthic Diatoms, Nanofrustulum shiloi and Striatella unipunctata, Encapsulated in Alginate Beads, Influence the Reproductive Efficiency of Paracentrotus lividus by Modulating the Gene Expression" Marine Drugs 19, no. 4: 230. https://doi.org/10.3390/md19040230
APA StyleGlaviano, F., Ruocco, N., Somma, E., De Rosa, G., Campani, V., Ametrano, P., Caramiello, D., Costantini, M., & Zupo, V. (2021). Two Benthic Diatoms, Nanofrustulum shiloi and Striatella unipunctata, Encapsulated in Alginate Beads, Influence the Reproductive Efficiency of Paracentrotus lividus by Modulating the Gene Expression. Marine Drugs, 19(4), 230. https://doi.org/10.3390/md19040230