Macroalgae as a Valuable Source of Naturally Occurring Bioactive Compounds for the Treatment of Alzheimer’s Disease


Abstract
1. Introduction
2. Methods
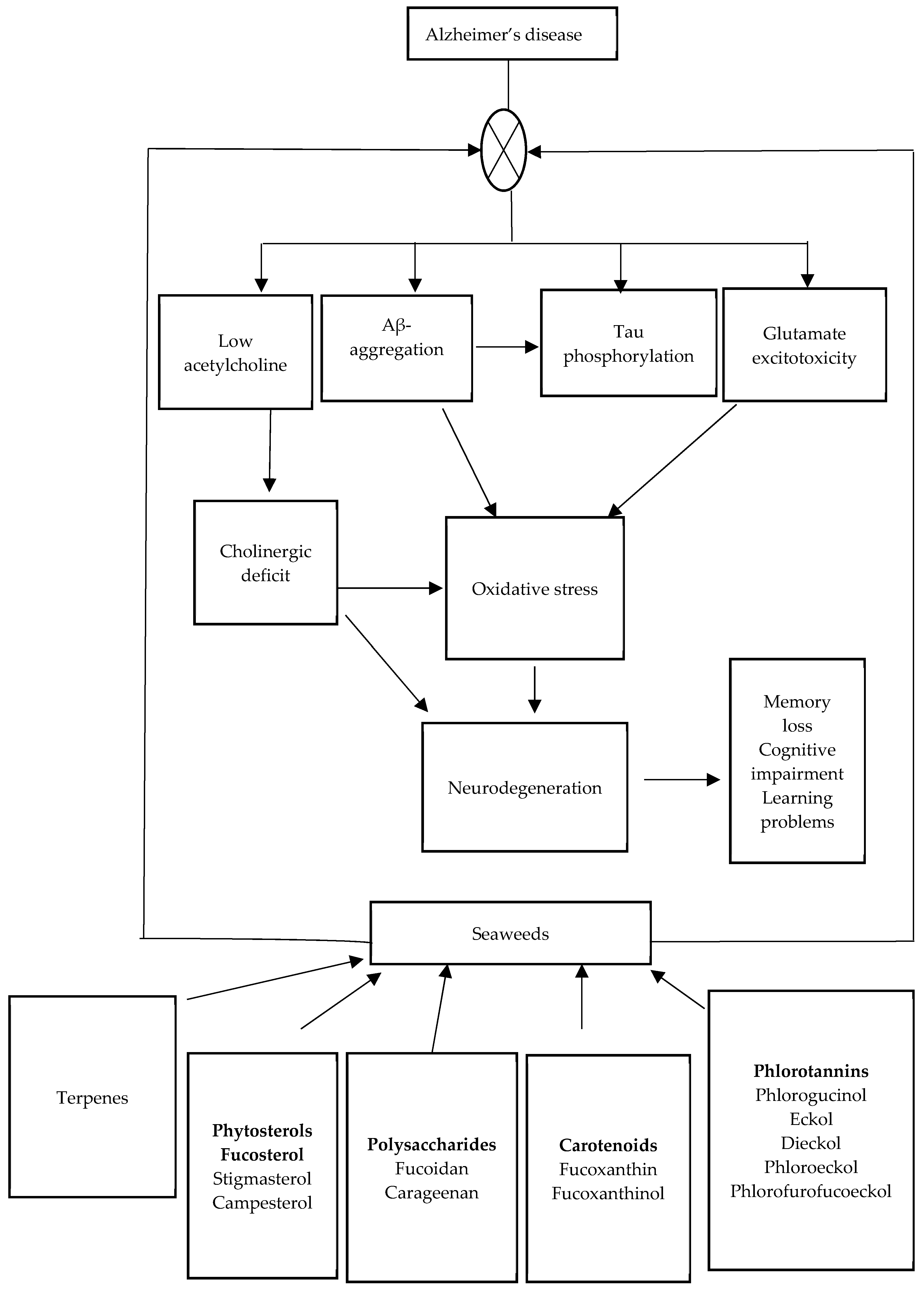
3. Etiology of AD
4. Therapeutic Role of Some Macroalgae in the Management of AD
4.1. Evidence from In Vitro Studies
4.1.1. Cholinesterase Inhibitory Activity
4.1.2. BACE-1 Inhibitory Activity
4.1.3. Action against Glutamate-Induced Neurotoxicity in Neuronal Cells
4.1.4. Protection against Aβ-Induced Neurotoxicity
4.1.5. Antioxidant Activity of Macroalgae and AD
4.2. Evidence from In Vivo Studies
4.2.1. Neuroprotective Activities of Some Macroalgal Extracts
4.2.2. Neuroprotective Effects of Macroalgal-Derived Compounds
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rengasamy, K.R.; Kulkarni, M.G.; Stirk, W.A.; van Staden, J. Advances in algal drug research with emphasis on enzyme inhibitors. Biotechnol. Adv. 2014, 32, 1364–1381. [Google Scholar] [CrossRef]
- Hong, D.D.; Hien, H.T. Nutritional analysis of Vietnamese seaweeds for food and medicineNo Title. Biofactors 2008, 22, 323–325. [Google Scholar] [CrossRef]
- Chengkui, Z.; Tseng, C.; Junfu, Z.; Chang, C.F. Chinese seaweeds in herbal medicineNo Title. Hdrobiologia 1984, 116, 152–154. [Google Scholar] [CrossRef]
- Niazi, A.K.; Kalra, S.; Irfan, A.; Islam, A. Thyroidology over the ages. Indian J. Endocrinol. Metab. 2011, 15, S121–S126. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y. Chinese Herbal Medicines: Comparisons and Characteristicstle; Churchill Livingstone: London, UK, 2009; Volume 268. [Google Scholar]
- Hamed, S.M.; El-Rhman, A.A.A.; Ibraheem, I.B.M. Role of marine macroalgae in plant protection & improvement for sustainable agriculture technology. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 104–110. [Google Scholar]
- Wells, M.I.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- Kelman, D.; Posner, E.K.; McDermid, K.J.; Tabandera, N.K.; Wright, P.R.; Wright, A.D. Antioxidant activity of Hawaiian marine algae. Mar. Drugs 2012, 10, 403–416. [Google Scholar] [CrossRef]
- Moussavou, G.; Kwak, D.H.; Obiang-Obonou, B.F.; Maranguy, C.A.; Dinzouna-Boutamba, S.; Lee, D.H.; Pissibanganga, O.G.M.; Ko, K.; Seo, J.I.; Choo, Y.K. Anticancer effects of different seaweeds on human colon and breast cancers. Mar. Drugs 2014, 12, 4898–4911. [Google Scholar] [CrossRef]
- Riahi, C.R.; Tarhouni, S.; Kharrat, R. Screening of anti-inflammatory and analgesic activities in marines macroalgae from Mediterranean Sea. Arch. Inst. Pasteur Tunis 2011, 88, 19–28. [Google Scholar]
- Zhao, C.; Yang, C.F.; Liu, B.; Lin, L.; Sarker, S.D.; Nahar, L.; Yu, H.; Cao, H.; Xiao, J. Bioactive compounds from marine macroalgae and their hypoglycemic benefits. Trends Foods Sci. Technol. 2018, 72, 1–12. [Google Scholar] [CrossRef]
- Tierney, M.S.; Croft, A.K.; Hayes, M. A review of antihypertensive and antioxidant activities in macroalgae. Bot. Mar. 2010, 53, 387–408. [Google Scholar] [CrossRef]
- Pérez, M.J.; Falque, E.; Domingue, H. Antimicrobial action of compounds from marine seaweed. Mar. Drugs 2016, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- Pangestuti, R.; Kim, S.K. Neuroprotective effects of marine algae. Mar. Drugs 2011, 9, 803–818. [Google Scholar] [CrossRef] [PubMed]
- Alghazwi, M.; Kan, Y.Q.; Zhang, W.; Gai, W.P.; Garson, M.J.; Smid, S. Neuroprotective activities of natural products from marine macroalgae during 1999–2015. J. Appl. Phycol. 2016, 28, 3599–3616. [Google Scholar] [CrossRef]
- Paris, D.; Beaulieu-Abdelahad, D.; Bachmeier, C.; Reed, J.; Ait-Ghezala, G.; Bishop, A.; Chao, J.; Mathura, V.; Crawford, F.; Mullan, M. Anatabine lowers Alzheimer’s Aβ production in vitro and in vivo. Eur. J. Pharmacol. 2011, 670, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Isik, A.T. Late onset Alzheimer’s disease in older people. Clin. Interv. Aging 2010, 5, 307–311. [Google Scholar] [CrossRef]
- Bekris, L.M.; Yu, C.; Bird, T.D.; Tsuang, T.W. Genetics of Alzheimer disease. J. Geriatr. Psychiatry Neurol. 2010, 23, 213–227. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Painter, M.M.; Bu, G.; Kanekiyo, T. Apolipoprotein E as a therapeutic target in Alzheimer’s disease: A review of basic research and clinical evidence. CNS Drugs 2016, 30, 773–789. [Google Scholar] [CrossRef]
- Liu, C.; Kanekiyo, T.; Xu, H.; Bu, G. Apolipoprotein E and Alzheimer disease: Risk, mechanisms, and Therapy. Nat. Rev. Neurol. 2013, 9, 106–118. [Google Scholar] [CrossRef]
- Kim, J.; Basak, J.M.; Holtzman, D.M. The role of apolipoprotein E in Alzheimer’s disease. Neuron 2009, 63, 287–303. [Google Scholar] [CrossRef]
- Shi, Y.; Yamada, K.; Liddelow, S.A.; Smith, S.T.; Zhao, L.; Luo, W.; Tsai, R.M.; Spina, S.; Grinberg, L.T.; Rojas, J.C.; et al. ApoE4 markedly exacerbates tau-mediated neurodegeneration in a mouse model of tauopathy. Nature 2017, 549, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhao, B. Oxidative stress and the pathogenesis of Alzheimer’s disease. Oxid. Med. Cell. Longev. 2013, 2013, 316523. [Google Scholar] [CrossRef] [PubMed]
- Haam, J.; Yakel, J.L. Cholinergic modulation of the hippocampal region and memory function. J. Neurochem. 2017, 142, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Sadigh-Eteghad, S.; Sabermarouf, B.; Majdi, A.; Talebi, M.; Farhoudi, M.; Mahmoudi, J. Amyloid-beta: A crucial factor in Alzheimer’s disease. Med. Princ. Pract. 2015, 24, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Reas, X.E. Amyloid and tau pathology in normal cognitive aging. J. Neurosci. 2017, 37, 7561–7563. [Google Scholar] [CrossRef]
- Kametani, F.; Hasegawa, M. Reconsideration of amyloid hypothesis and tau hypothesis in Alzheimer’s disease. Front. Neurosci. 2018, 12, 25. [Google Scholar] [CrossRef]
- Shal, B.; Ding, W.; Ali, H.; Kim, Y.S.; Khan, S. Anti-neuroinflammatory potential of natural products in attenuation of Alzheimer’s disease. Front. Pharmacol. 2018, 9, 548. [Google Scholar] [CrossRef]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A. Therapeutic potentials of microalgae in the treatment of Alzheimer’s disease. Molecules 2017, 22, 480. [Google Scholar] [CrossRef]
- Oboh, G.; Nwanna, E.E.; Oyeleye, S.I.; Olasehinde, T.A.; Ogunsuyi, O.B.; Boligon, A.A. In vitro neuroprotective potentials of aqueous and methanol extracts from Heinsia crinita leaves. Food Sci. Hum. Wellness 2016, 5, 95–102. [Google Scholar] [CrossRef]
- Qui, C.; Kivipelto, M.; Strauss, E. Epidemiology of Alzheimer’s disease: Occurrence, determinants, and strategies toward intervention. Dialogues Clin. Neurosci. 2009, 11, 111–128. [Google Scholar]
- Graham, W.V.; Bonito-Oliva, A.; Sakmar, T.P. Update on Alzheimer’s disease therapy and prevention strategies. Annu. Rev. Med. 2017, 68, 413–430. [Google Scholar] [CrossRef] [PubMed]
- Yiannopoulou, K.G.; Papageorgiou, S.G. Current and future treatments for Alzheimer’s disease. Ther. Adv. Neurol. Disord. 2013, 6, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Frozza, R.L.; Lourenco, M.V.; de Felice, F.G. Challenges for Alzheimer’s disease therapy: Insights from novel mechanisms beyond memory defects. Front. Neurosci. 2018, 12, 37. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Olasehinde, T.A.; Ademosun, A.O. Essential oil from lemon peels inhibit key enzymes linked to neurodegenerative conditions and pro-oxidant induced lipid peroxidation. J. Oleo Sci. 2014, 63, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Olasehinde, T.A.; Odjadjare, E.C.; Mabinya, L.V.; Olaniran, A.O.; Okoh, A.I. Chlorella sorokiniana and Chlorella minutissima exhibit antioxidant potentials, inhibit cholinesterases and modulate disaggregation of β-amyloid fibrils. Electron. J. Biotechnol. 2019, 40, 1–9. [Google Scholar] [CrossRef]
- Oboh, G.; Adewuni, T.M.; Ademosun, A.O.; Olasehinde, T.A. Sorghum stem extract modulates Na+/K+-ATPase, ecto-5′-nucleotidase, and acetylcholinesterase activities. Comp. Clin. Pathol. 2016, 25, 749–756. [Google Scholar] [CrossRef]
- Oboh, G.; Oyeleye, S.I.; Akintemi, O.A.; Olasehinde, T.A. Moringa oleifera supplemented diet modulates nootropic-related biomolecules in the brain of STZ-induced diabetic rats treated with acarbose. Metab. Brain Dis. 2018, 33, 457–466. [Google Scholar] [CrossRef]
- Oboh, G.; Adewuni, T.M.; Ademiluyi, A.O.; Olasehinde, T.A.; Ademosun, A.O. Phenolic constituents and inhibitory effects of Hibiscus sabdariffa L.(Sorrel) calyx on cholinergic, monoaminergic, and purinergic enzyme activities. J. Diet. Suppl. 2018, 15, 910–922. [Google Scholar] [CrossRef]
- Oboh, G.; Ademosun, A.O.; Ogunsuyi, O.B.; Oyedola, E.T.; Olasehinde, T.A.; Oyeleye, S.I. In vitro anticholinesterase, antimonoamine oxidase and antioxidant properties of alkaloid extracts from kola nuts (Cola acuminata and Cola nitida). J. Complement. Integr. Med. 2018. [Google Scholar] [CrossRef]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Aqueous–ethanol extracts of some South African seaweeds inhibit beta-amyloid aggregation, cholinesterases, and beta-secretase activities in vitro. J. Food Biochem. 2019, 43, e12870. [Google Scholar] [CrossRef]
- Olasehinde, T.A.; Mabinya, L.V.; Olaniran, A.O.; Okoh, A.I. Chemical characterization of sulfated polysaccharides from Gracilaria gracilis and Ulva lactuca and their radical scavenging, metal chelating, and cholinesterase inhibitory activities. Int. J. Food Prop. 2019, 22, 100–110. [Google Scholar] [CrossRef]
- Olasehinde, T.A.; Mabinya, L.V.; Olaniran, A.O.; Okoh, A.I. Chemical characterization, antioxidant properties, cholinesterase inhibitory and anti-amyloidogenic activities of sulfated polysaccharides from some seaweeds. Bioact. Carbohydr. Diet. Fibre 2019, 18, 100182. [Google Scholar] [CrossRef]
- Syad, A.N.; Rajamohamed, B.S.; Shunmugaiah, K.P.; Devi, P.K. Neuroprotective effect of the marine macroalga Gelidiella acerosa: Identification of active compounds through bioactivity-guided fractionation. Pharm. Biol. 2016, 54, 2073–2081. [Google Scholar] [CrossRef] [PubMed]
- Rengasamy, K.R.; Amoo, S.O.; Aremu, A.O.; Stirk, W.A.; Gruz, J.; Šubrtová, M.; Doležal, K.; Staden, J.V. Phenolic profiles, antioxidant capacity, and acetylcholinesterase inhibitory activity of eight South African seaweeds. J. Appl. Phycol. 2015, 27, 1599–1605. [Google Scholar] [CrossRef]
- Jung, S.H.; Young, U.M.; Inho, K.; Suengmok, C.; Daeseok, H.; Changho, L. In vitro screening for anti-dementia activities of seaweed extracts. J. Korean Soc. Food Sci. Nutr. 2016, 45, 966–997. [Google Scholar]
- Rafiquzzaman, S.M.; Ki, E.Y.; Lee, J.M.; Mohibbullah, M.; BadrulAlam, M.; Moon, S.; Kim, J.M.; Kong, S. Anti-Alzheimers and anti-inflammatory activities of a glycoprotein purified from the edible brown alga Undaria pinnatifida. Food Res. Int. 2015, 77, 118–124. [Google Scholar] [CrossRef]
- Shanmuganathan, B.; Sheeja, M.D.; Sathya, S.; Devi, P.K. Antiaggregation potential of Padina gymnospora against the toxic Alzheimer’s beta-amyloid peptide 25–35 and cholinesterase inhibitory property of its bioactive compounds. PLoS ONE 2015, 10, e0141708. [Google Scholar] [CrossRef]
- Castro-Silva, E.S.; Bello, M.; Hernández-Rodríguez, M.; Correa-Basurto, J.; Murillo-Álvarez, J.I.; Rosales-Hernández, M.C.; Muñoz-Ochoa, M. Invitro and insilico evaluation of fucosterol from Sargassum horridum as potential human acetylcholinesterase inhibitor. J. Biomol. Struc. Dyn. 2018. [Google Scholar] [CrossRef]
- Choi, B.W.; Lee, H.S.; Shin, H.; Lee, B.H. Multifunctional activity of polyphenolic compounds associated with a potential for Alzheimer’s disease therapy from Ecklonia Cava. Phytother. Res. 2015, 29, 549–553. [Google Scholar] [CrossRef]
- Sathya, M.; Premkumar, P.; Karthick, C.; Moorthi, P.; Jayachandran, K.S.; Anusuyadevi, M. BACE1 in Alzheimer’s disease. Clin. Chim. Acta 2012, 24, 171–178. [Google Scholar] [CrossRef]
- Cheng, X.; He, P.; Lee, T.; Yao, H.; Li, R.; Shen, Y. High activities of BACE1 in brains with mild cognitive impairment. Am. J. Pathol. 2014, 184, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Rockenstein, E.; Mante, M.; Alford, M.; Adame, A.; Crews, L.; Hashimoto, M.; Esposito, L.; Mucke, L.; Masliah, E. High β-secretase activity elicits neurodegeneration in transgenic mice despite reductions in amyloid-β levels. J. Biol. Chem. 2005, 280, 32957–32967. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.A.; Ali, M.Y.; Choi, R.J.; Jeong, H.O.; Chung, H.Y.; Choi, J.S. Kinetics and molecular docking studies of fucosterol and fucoxanthin, BACE1 inhibitors from brown algae Undaria pinnatifida and Ecklonia stolonifera. Food Chem. Toxicol. 2016, 89, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Seong, S.H.; Ali, M.Y.; Kim, H.R.; Jung, H.A.; Choi, J.S. BACE1 inhibitory activity and molecular docking analysis of meroterpenoids from Sargassum serratifolium. Bioorg. Med. Chem. 2017, 25, 3964–3970. [Google Scholar] [CrossRef] [PubMed]
- Wagle, A.; Seong, S.H.; Zhao, B.T.; Woo, M.H.; Jung, H.A.; Choi, J.S. Comparative study of selective in vitro and in silico BACE1 inhibitory potential of glycyrrhizin together with its metabolites, 18a-and 18b-glycyrrhetinic acid, isolated from Hizikia fusiformis. Arch. Pharm. Res. 2018, 41, 409–418. [Google Scholar] [CrossRef]
- Dong, X.; Wang, Y.; Qin, Z. 2009. Molecular mechanisms of excitotoxicity and their relevance to pathogenesis of neurodegenerative diseases. Acta Pharm. Sin. 2009, 30, 379–387. [Google Scholar] [CrossRef]
- Fernandes, F.; Barbosa, M.; Oliveira, A.P.; Azevedo, I.C.; Sousa-Pinto, I.; Valentão, P.; Andrade, P.B. The pigments of kelps (Ochrophyta) as part of the flexible response to highly variable marine environments. J. Appl. Phycol. 2016, 28, 3689–3696. [Google Scholar] [CrossRef]
- Kim, J.J.; Kang, Y.J.; Shin, S.A.; Bak, D.H.; Lee, J.W.; Lee, K.B.; Yoo, Y.C.; Kim, D.K.; Lee, B.H.; Kim, D.W.; et al. Phlorofucofuroeckol improves glutamate-induced neurotoxicity through modulation of oxidative stress-mediated mitochondrial dysfunction in PC12 cells. PLoS ONE 2016, 11, 0163433. [Google Scholar] [CrossRef]
- O’Brien, R.J.; Wong, B.C. Amyloid precursor protein processing and Alzheimer’s disease. Annu. Rev. Neurosci. 2011, 34, 185–204. [Google Scholar] [CrossRef]
- Giffin, J.C.; Richards, R.C.; Craft, C.; Jahan, N.; Leggiadro, C.; Chopin, T.; Szemerda, M.; MacKinnon, S.L.; Ewart, K.V. An extract of the marine alga Alaria esculenta modulates α-synuclein folding and amyloid formation. Neurosci. Lett. 2017, 22, 87–93. [Google Scholar] [CrossRef]
- Syad, A.N.; Devi, K.P. Assessment of anti-amyloidogenic activity of marine red alga G. acerosa against Alzheimer’s beta-amyloid peptide 25–35. Neurol. Res. 2015, 37, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Alghazwi, M.; Smid, S.; Zhang, W. In vitro protective activity of South Australian marine sponge and macroalgae extracts against amyloid beta (Aβ1–42) induced neurotoxicity in PC-12 cells. Neurotoxicol. Teratol. 2018, 68, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Gan, S.Y.; Wong, L.Z.; Wong, J.W.; Tan, E.L. Fucosterol exerts protection against amyloid β-induced neurotoxicity, reduces intracellular levels of amyloid β and enhances the mRNA expression of neuroglobin in amyloid β-induced SH-SY5Y cells. Int. J. Biol. Macromol. 2019, 121, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Alghazwi, M.; Smid, S.; Karpiniec, S.; Zhang, W. Comparative study on neuroprotective activities of fucoidans from Fucus vesiculosus and Undaria pinnatifida. Int. J. Biol. Macromol. 2019, 122, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.J.; Ahn, S.; Ryu, J.; Choi, M.; Choi, S.; Chong, Y.H.; Hyun, J.W.; Chang, M.; Kim, H.S. Phloroglucinol attenuates the cognitive deficits of the 5XFAD mouse model of Alzheimer’s disease. PLoS ONE 2015, 10, e0135686. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, J.; Huang, C.; Zhao, J.; Lin, J.; Zhou, X.; Naman, C.B.; Wang, N.; Gerwick, W.H.; Wang, Q.; et al. Eckmaxol, a phlorotannin extracted from Ecklonia maxima, produces anti-β-amyloid oligomer neuroprotective effects possibly via directly acting on glycogen synthase kinase 3β. ACS Chem. Neurosci. 2018, 9, 1349–1356. [Google Scholar] [CrossRef]
- Oh, J.H.; Choi, J.S.; Nam, T. Fucosterol from an edible brown alga Ecklonia stolonifera prevents soluble amyloid beta-induced cognitive dysfunction in aging rats. Mar. Drugs 2018, 16, 368. [Google Scholar] [CrossRef]
- Lin, J.; Yu, J.; Zhao, J.; Zhang, K.; Zheng, J.; Wang, J.; Huang, C.; Zhang, J.; Yan, X.; Gerwick, W.H.; et al. Fucoxanthin, a marine carotenoid, attenuates β-amyloid oligomer-induced neurotoxicity possibly via regulating the PI3K/Akt and the ERK pathways in SH-SY5Y cells. Oxid. Med. Cell. Longev. 2017, 2017, 6792543. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, S.; An, C.; Zhang, H.; Sun, Y.; Li, Y.; Pu, X. Neuroprotective effect of fucoxanthin on β-amyloid induced cell death. J. Chin. Pharm. Sci. 2015, 24, 467–470. [Google Scholar]
- Wei, H.; Gao, Z.; Zheng, L.; Zhang, C.; Liu, Z.; Yang, Y.; Teng, H.; Hou, L.; Yin, Y.; Zou, X. Protective effects of fucoidan on Aβ25–35 and d-Gal-induced neurotoxicity in PC12 cells and d-Gal-induced cognitive dysfunction in mice. Mar. Drugs 2017, 15, 77. [Google Scholar] [CrossRef]
- Sathya, R.; Kanaga, N.; Sankar, P.; Jeeva, S. Antioxidant properties of phlorotannins from brown seaweed Cystoseira trinodis (Forsskål) C. Agardh. Arab. J. Chem. 2017, 10, S2608–S2614. [Google Scholar] [CrossRef]
- Li, Y.; Fu, X.; Duan, D.; Liu, X.; Xu, J.; Gao, X. Extraction and identification of phlorotannins from the brown alga, Sargassum fusiforme (Harvey) setchell. Mar. Drugs 2017, 15, 49. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zhang, J.; Fan, J.; Clark, J.; Shen, P.; Li, Y.; Zhang, C. Microwave assisted extraction of phenolic compounds from four economic brown macroalgae species and evaluation of their antioxidant activities and inhibitory effects on α-amylase, α-glucosidase, pancreatic lipase and tyrosinase. Food Res. Int. 2018, 113, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Arulkumar, A.; Rosemary, T.; Paramasivam, S.; Rajendran, R.B. Phytochemical composition, in vitro antioxidant, antibacterial potential and GC-MS analysis of red seaweeds (Gracilaria corticata and Gracilaria edulis) from Palk Bay, India. Biocatal. Agric. Biotechnol. 2018, 15, 63–71. [Google Scholar] [CrossRef]
- Kosanić, M.; Ranković, B.; Stanojković, T. Biological activities of two macroalgae from Adriatic coast of Montenegro. Saudi J. Biol. Sci. 2015, 22, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Alencar, D.B.; Carvalho, C.F.T.; Rebouças, R.H.; Santos, D.R.D.; Pires-Cavalcante, K.M.; Lima, R.L.; Baracho, B.M.; Bezerra, R.M.; Viana, F.A.; Vieira, R.H.; et al. Bioactive extracts of red seaweeds Pterocladiella capillacea and Osmundaria obtusiloba (Floridophyceae: Rhodophyta) with antioxidant and bacterial agglutination potential. Asian Pac. J. Trop. Med. 2016, 9, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Agregán, R.; Munekata, P.E.; Domínguez, R.; Carballo, J.; Franco, D.; Lorenzo, J.M. Proximate composition, phenolic content and in vitro antioxidant activity of aqueous extracts of the seaweeds Ascophyllum nodosum, Bifurcaria bifurcata and Fucus vesiculosus. Effect of addition of the extracts on the oxidative stability of canola oil under accelerated storage conditions. Food Res. Int. 2017, 99, 986–994. [Google Scholar]
- Pinteus, S.; Silva, J.; Alves, C.; Horta, A.; Fino, N.; Inês, A. Rodrigues susana mendes rui pedrosa. cytoprotective effect of seaweeds with high antioxidant activity from the Peniche coast (Portugal). Food Chem. 2017, 218, 591–599. [Google Scholar] [CrossRef]
- Chiboub, O.; Ktari, L.; Sifaoui, I.; López-Arencibia, A.; Batle, M.R.; Mejri, M.; Valladares, B.; Abderrabba, M.; Piñero, J.E.; Lorenzo-Morales, J. In vitro amoebicidal and antioxidant activities of some Tunisian seaweeds. Exp. Parasit. 2017, 183, 76–80. [Google Scholar] [CrossRef]
- Hifney, A.F.; Fawzy, M.A.; Abdel-Gawad, K.M.; Gomaa, M. Industrial optimization of fucoidan extraction from Sargassum sp. and its potential antioxidant and emulsifying activities. Food Hydrocoll. 2016, 54, 77–88. [Google Scholar] [CrossRef]
- Rajauria, G.; Foley, B.; Abu-Ghannam, N. Identification and characterization of phenolic antioxidant compounds from brown Irish seaweed Himanthalia elongata using LC-DAD–ESI-MS/MS. Innov. Food Sci. Emerg. Technol. 2016, 37, 261–268. [Google Scholar] [CrossRef]
- Leyton, A.; Pezoa-Conte, R.; Barriga, A.; Buschmann, A.H.; Mäki-Arvela, P.; Mikkola, J.P.; Lienqueo, M.E. Identification and efficient extraction method of phlorotannins from the brown seaweed Macrocystis pyrifera using an orthogonal experimental design. Algal Res. 2016, 16, 201–208. [Google Scholar] [CrossRef]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Neuroprotective effects of some seaweeds against Zn–induced neuronal damage in HT-22 cells via modulation of redox imbalance, inhibition of apoptosis and acetylcholinesterase activity. Metab. Brain Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Phenolic composition, antioxidant activity, anticholinesterase potential and modulatory effects of aqueous extracts of some seaweeds on β-amyloid aggregation and disaggregation. Pharm. Biol. 2019, 57, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wu, S.; Yang, W.; Kuan, A.; Chen, C. Antioxidant activities of crude extracts of fucoidan extracted from Sargassum glaucescens by a compressional-puffing hydrothermal extraction process. Food Chem. 2016, 197, 1121–1129. [Google Scholar] [CrossRef] [PubMed]
- Palanisamy, S.; Vinosha, M.; Marudhupandi, T.; Rajasekar, P.; Prabhu, N.M. Isolation of fucoidan from Sargassum polycystum brown algae: Structural characterization, in vitro antioxidant and anticancer activity. Int. J. Biol. Macromol. 2017, 102, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Hifney, A.W.; Fawzy, M.A.; Abdel-Gawad, K.M.; Gomaa, M. Upgrading the antioxidant properties of fucoidan and alginate from Cystoseira trinodis by fungal fermentation or enzymatic pretreatment of the seaweed biomass. Food Chem. 2018, 269, 387–395. [Google Scholar] [CrossRef]
- Gomaa, M.; Fawzy, M.A.; Hifney, A.F.; Abdel-Gawad, K.M. Use of the brown seaweed Sargassum latifolium in the design of alginate-fucoidan based films with natural antioxidant properties and kinetic modeling of moisture sorption and polyphenolic release. Food Hydrocoll. 2018, 82, 64–72. [Google Scholar] [CrossRef]
- Khajouei, R.A.; Keramat, J.; Hamdami, N.; Delattre, C.; Laroche, C.; Gardarin, C.; Lecerf, D.; Desbrières, J.; Djelveh, G.; Michaud, P. Extraction and characterization of an alginate from the Iranian brown seaweed Nizimuddinia zanardini. Int. J. Biol. Macromol. 2018, 118, 1073–1081. [Google Scholar] [CrossRef]
- Yao, Y.; Xiang, H.; You, L.; Cui, C.; Sun-Waterhouse, D.; Zhao, M. Hypolipidaemic and antioxidant capacities of polysaccharides obtained from Laminaria japonica by different extraction media in diet-induced mouse model. Int. J. Food Sci. Tech. 2017, 52, 2274–2281. [Google Scholar] [CrossRef]
- Kazir, M.; Abuhassira, Y.; Robin, A.; Nahor, O.; Luo, J.; Israel, A.; Golberg, A.; Livney, Y.D. Extraction of proteins from two marine macroalgae, Ulva sp. and Gracilaria sp., for food application, and evaluating digestibility, amino acid composition and antioxidant properties of the protein concentrates. Food Hydrocoll. 2019, 87, 194–203. [Google Scholar] [CrossRef]
- Mohibbullah, M.; Maqueshudul, M.; Bhuiyan, H.; Hannan, M.A.; Getachew, P.; Hong, Y.; Choi, J.; Choi, S.; Moon, S. The edible red alga Porphyra yezoensis promotes neuronal survival and cytoarchitecture in primary hippocampal neurons. Cell. Mol. Neurobiol. 2016, 36, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Rajauria, G.; Foley, B.; Abu-Ghannam, N. Characterization of dietary fucoxanthin from Himanthalia elongata brown seaweed. Food Res. Int. 2017, 99, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Lin, J.; Yu, R.; He, S.; Wang, Q.; Cui, W.; Zhang, J. Fucoxanthin prevents H2O2-induced neuronal apoptosis via concurrently activating the PI3-K/Akt cascade and inhibiting the ERK pathway. Food Nutr. Res. 2017, 61, 1304678. [Google Scholar] [CrossRef]
- Mohibbullah, M.; Haque, M.N.; Khan, M.N.; Park, I.S.; Moon, I.S.; Hong, Y. Neuroprotective effects of fucoxanthin and its derivative fucoxanthinol from the phaeophyte Undaria pinnatifida attenuate oxidative stress in hippocampal neurons. J. Appl. Phycol. 2018. [Google Scholar] [CrossRef]
- Syad, A.N.; Devi, P.K. Gelidiella acerosa protects against Aβ 25–35-induced toxicity and memory impairment in Swiss Albino mice: An in vivo report. Pharm. Biol. 2017, 55, 1423–1435. [Google Scholar]
- Um, Y.M.; Lim, D.W.; Son, H.J.; Cho, S.; Lee, C. Phlorotannin-rich fraction from Ishige foliacea brown seaweed prevents the scopolamine-induced memory impairment via regulation of ERK-CREB-BDNF pathway. J. Funct. Foods 2018, 40, 110–116. [Google Scholar] [CrossRef]
- Choi, J.Y.; Mohibbullah, M.; Park, I.S.; Moon, I.S.; Hong, Y.K. An ethanol extract from the phaeophyte Undaria pinnatifida improves learning and memory impairment and dendritic spine morphology in hippocampal neurons. J. Appl. Phycol. 2018, 30, 129–136. [Google Scholar] [CrossRef]
- Hu, P.; Li, Z.; Chen, M.; Sun, Z.; Ling, Y.; Jiang, J.; Huang, C. Structural elucidation and protective role of a polysaccharide from Sargassum fusiforme on ameliorating learning and memory deficiencies in mice. Carbohydr. Polym. 2016, 139, 150–158. [Google Scholar] [CrossRef]
- Wang, X.; Yi, K.; Zhao, Y. Fucoidan inhibits amyloid-β-induced toxicity in transgenic Caenorhabditis elegans by reducing the accumulation of amyloid-β and decreasing the production of reactive oxygen species. Food Funct. 2018, 9, 552–560. [Google Scholar] [CrossRef]
- Lin, J.; Huang, L.; Yu, J.; Xiang, S.; Wang, J.; Zhang, J.; Yan, X.; Cui, W.; He, S.; Wang, Q. Fucoxanthin, a marine carotenoid, reverses scopolamine-induced cognitive impairments in mice and inhibits acetylcholinesterase in vitro. Mar. Drugs 2016, 14, 67. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Liu, F.; Lin, J.; Chen, H.; Huang, C.; Chen, L.; Zhou, Y.; Ye, L.; Zhang, K.; Jin, J.; et al. Fucoxanthin inhibits β-amyloid assembly and attenuates β-amyloid oligomer-induced cognitive impairments. J. Agric. Food Chem. 2017, 65, 4092–4102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, H.; Fan, Y.; Gao, Y.; Li, X.; Hu, Z.; Ding, K.; Wang, Y.; Wang, X. Fucoxanthin provides neuroprotection in models of traumatic brain injury via the Nrf2-ARE and Nrf2-autophagy pathways. Sci. Rep. 2017, 7, 46763. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.J.; Mahmood, U.; Kim, H.; Choi, M.; Choi, Y.; Lee, J.; Cho, J.; Hyun, J.; Kim, Y.S.; Chang, M.; et al. Phloroglucinol ameliorates cognitive impairments by reducing the amyloid β peptide burden and pro-inflammatory cytokines in the hippocampus of 5XFAD mice. Free Radic. Biol. Med. 2018, 126, 221–234. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Class of Compounds | Components | Algal Source | Mechanism of Action | Reference |
|---|---|---|---|---|
| Crude extracts | Benezene:ethyl acetate fraction | G. acerosa | Inhibition of AChE | [44] |
| Methanol extracts | E. cava | Inhibition of AChE and BACE-1 | ||
| E. kurome | ||||
| M. simples | Inhibition of AChE | |||
| Phlorotannins | Phlorofucofuroeckol | E. cava | Inhibition AChE, BChE, and BACE-1 | [46] |
| Polysaccharides | Purified glycoprotein | U. pinnatifida | Inhibition of AChE, BChE and BACE-1`` | [47] |
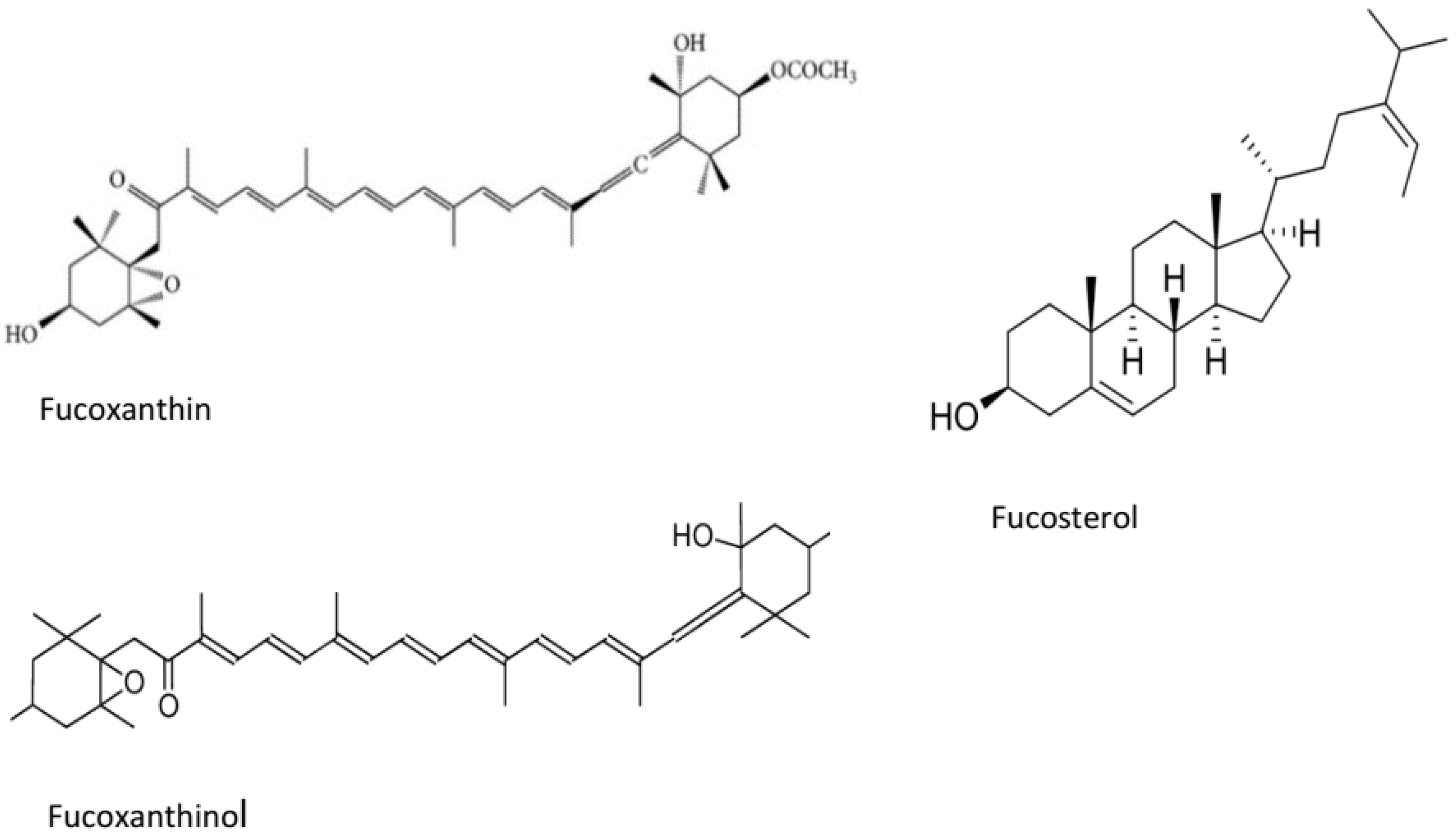
| Sterol | Fucosterol | S. horridum | Inhibition of AChE | [49] |
| Carotenoids | Fucoxanthin | U. pinnatifida E. stolonifera | Inhibition of BACE-1 | [54] |
| Triterpenoid-saponin | Sarahydroquinoic acid | S. serratifolium | Inhibition of BACE-1 | [55] |
| Glycyrrhizin | H. fusiformis | Inhibition of AChE, BChE, and BACE-1 | [56] | |
| 18α-glycyrrhetinic acid | ||||
| 18β-glycyrrhetinic acid |
| Class of Compounds | Components | Algal Source | Mechanism of Action | Reference |
|---|---|---|---|---|
| Crude extracts | Aqueous extracts | A. esculenta | Inhibition of amyloid formation | [61] |
| Acetone extracts | P. gymnospora | Anti-aggregation and dis-aggregation of amyloid fibrils | [48] | |
| Ether/benzene extracts | G. acerosa | Prevention of Aβ25–35 formation and dis-aggregation of pre-formed fibrils | [62] | |
| Phlorotannins | Phloroglucinol | E. cava | Inhibition of Aβ-induced-cytotoxicity and protection against ROS accumulation in HT-22 cells | [64] |
| Eckmaxol | E. maxima | Prevention of Aβ-induced neuronal apoptosis and decrease in intracellular ROS | [67] | |
| Phytosterol | Fucosterol | Padina australis | Reduction of APP mRNA and inhibition of Aβ-induced neurotoxicity | [64] |
| E. stolonifera | Attenuation of Aβ-induced cognitive dysfunction | [68] | ||
| Carotenoid | Fucoxanthin | Sargassum horneri | Attenuation of Aβ-oligomer-induced neurotoxicity in SYH-SY5Y cells | [69] |
| Attenuation of Aβ-induced neurotoxicity in PC12-cells | [70] | |||
| Sulfated polysaccharides | Fucoidan | U. pinnatifida F. vesiculosus | Protection against Aβ1–42-induced neuronal death in PC-12 cells | [65] |
| Inhibition of Aβ25–35-induced neurotoxicity in PC-12 cells | [71] |
| Class of Compounds | Components | Algal Source | Mechanism of Action | Reference |
|---|---|---|---|---|
| Crude extracts | Methanol extract | Ascophyllum nodosum | ABTS and DPPH scavenging activity | [74] |
| Laminaria japonica | Ferric reducing antioxidant property | |||
| Lessonia trabeculate | ||||
| Lessonia nigrescens | ||||
| Gracilaria edulis | DPPH, ABTS, and NO radical scavenging activities | [75] | ||
| Gracilaria corticata | ||||
| Myelophycus simplex | ABTS radical scavenging activity | [46] | ||
| Ecklonia cava | Attenuation of H202-induced oxidative damage in SH-SY5Y cells | |||
| E. kurome | ||||
| Acetone extract | Ulva lactuca | DPPH and superoxide anion scavenging activity | [76] | |
| Entermorpha intestinalis | Ferric reducing antioxidant property | |||
| Ethanol/hexane extract | Pterocladiella capillacea | DPPH radical scavenging activity | [77] | |
| Osmindaria obtusiloba | Metal chelating activity | |||
| Aqueous extract | Ascophyllum nodosum | DPPH, ABTS, and hydroxyl radical scavenging activities | [78] | |
| Bifurcaria bifurcate | Ferric reducing antioxidant capacity | |||
| Fucus vesiculosus | Inhibition of lipid oxidation | |||
| Phlorotannins | Phlorotannin extract | Macrocytis pyrifera | DPPH radical scavenging activity | [83] |
| Polysaccharides | Fucoidan | Sargassum glaucescens | ABTS and DPPH scavenging and metal chelating activities | [85] |
| Sargassum polycystum | DPPH radical scavenging activity Ferric reducing antioxidant property | [87] | ||
| Fucoidan and alginate | Cystoseira trinodis | Ferric reducing antioxidant property | [88] | |
| Fucoidan and alginate | Sargassum latifolium | Hydroxyl radical scavenging activity | [89] | |
| Sodium alginate | Nizimuddinia zanardini | DPPH radical scavenging activity | [90] | |
| Polysaccharides | Laminaria japonica | DPPH and oxygen radical scavenging activity | [91] | |
| Fucoidan | U. pinnatifida F. vesiculosus | Attenuation of hydrogen peroxide-induced oxidative stress and apoptosis in PC-12 cells | [65] | |
| Activation of superoxide dismutase and glutathione in Aβ-induced neurotoxicity in PC-12 cells | [71] | |||
| Proteins | Protein extracts | Ulva spp. | Ferric reducing antioxidant property | [92] |
| Gracilaria spp. | Oxygen radical absorption capacity | |||
| Carotenoids | Fucoxanthin | Himanthalia elongata | Ferric reducing antioxidant property DPPH radical scavenging activity | [94] |
| Sargassum horneri | Attenuation of H2O2-induced neuronal apoptosis and intracellular ROS | [95] | ||
| Reduced malondialdehyde levels and SOD activity in Aβ-induced cell death in PC12 cells | [70] | |||
| Fucoxanthinol | Undaria pinnatifida | Attenuation of oxidative stress in rats’ hippocampal neurons | [96] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Macroalgae as a Valuable Source of Naturally Occurring Bioactive Compounds for the Treatment of Alzheimer’s Disease. Mar. Drugs 2019, 17, 609. https://doi.org/10.3390/md17110609
Olasehinde TA, Olaniran AO, Okoh AI. Macroalgae as a Valuable Source of Naturally Occurring Bioactive Compounds for the Treatment of Alzheimer’s Disease. Marine Drugs. 2019; 17(11):609. https://doi.org/10.3390/md17110609
Chicago/Turabian StyleOlasehinde, Tosin A., Ademola O. Olaniran, and Anthony I. Okoh. 2019. "Macroalgae as a Valuable Source of Naturally Occurring Bioactive Compounds for the Treatment of Alzheimer’s Disease" Marine Drugs 17, no. 11: 609. https://doi.org/10.3390/md17110609
APA StyleOlasehinde, T. A., Olaniran, A. O., & Okoh, A. I. (2019). Macroalgae as a Valuable Source of Naturally Occurring Bioactive Compounds for the Treatment of Alzheimer’s Disease. Marine Drugs, 17(11), 609. https://doi.org/10.3390/md17110609