A Novel High-Mannose Specific Lectin from the Green Alga Halimeda renschii Exhibits a Potent Anti-Influenza Virus Activity through High-Affinity Binding to the Viral Hemagglutinin


Abstract
:1. Introduction
2. Results
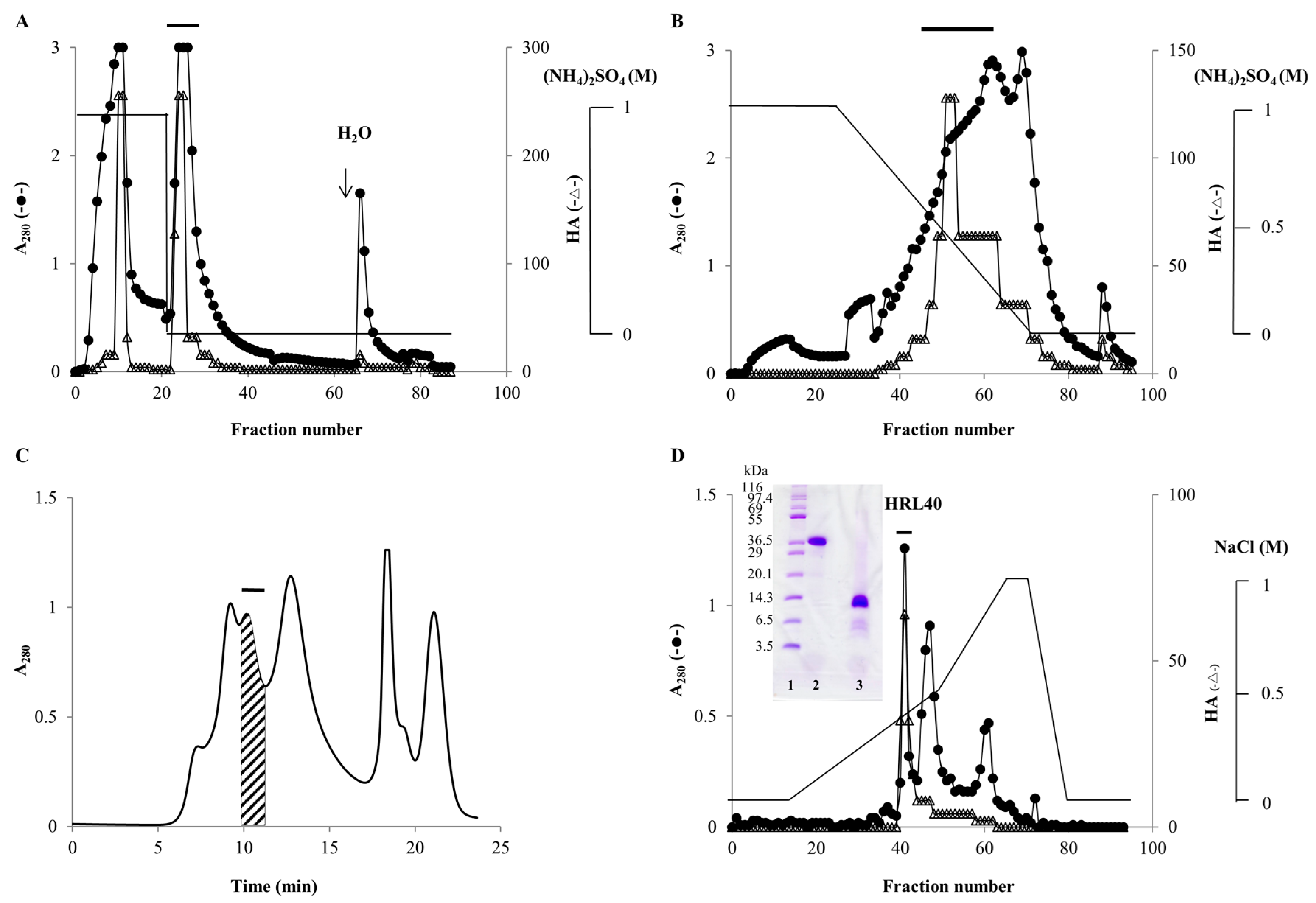
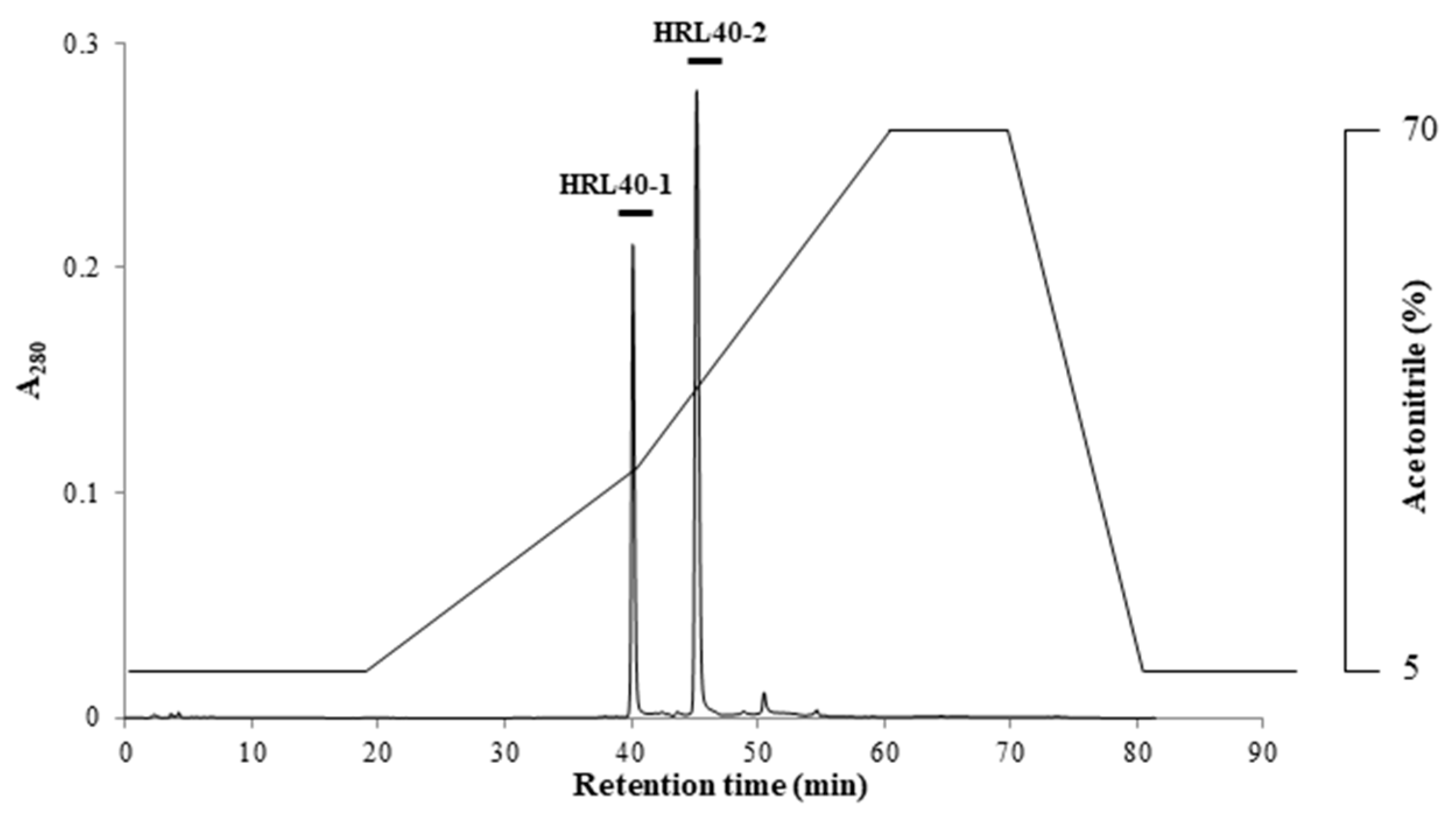
2.1. Purification of HM-Specific Lectin HRL40
2.2. Hemagglutination-Inhibition Test
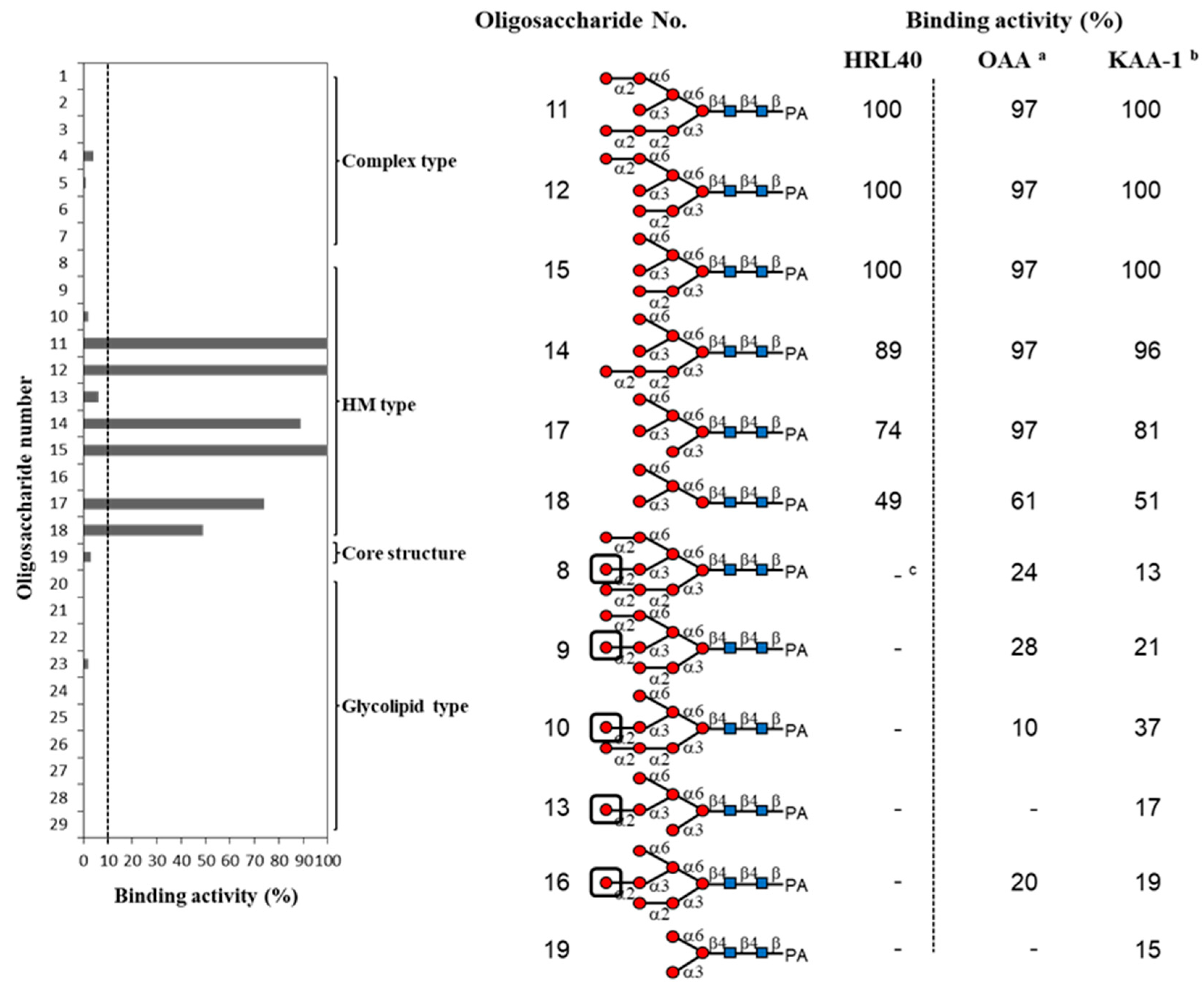
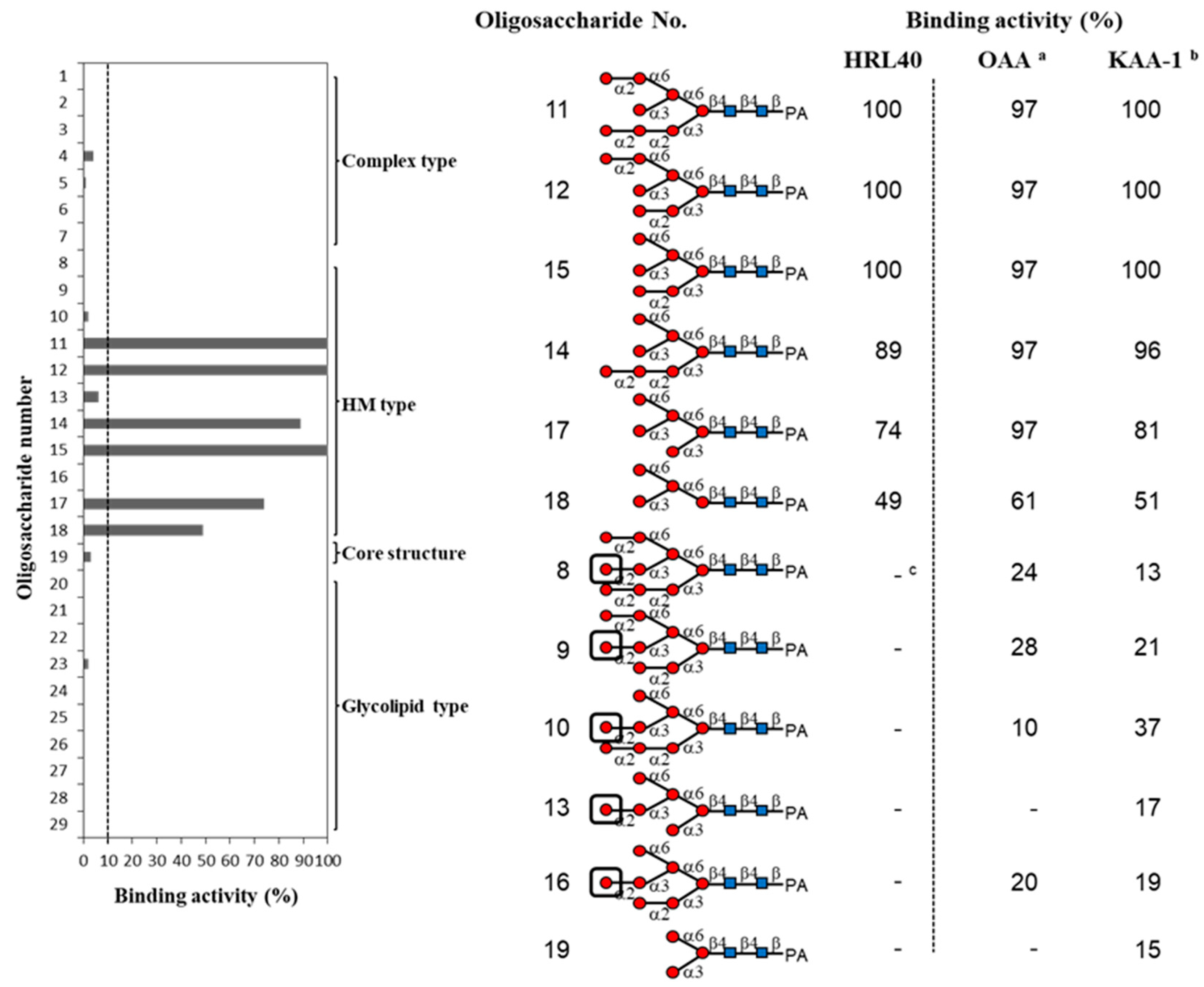
2.3. Oligosaccharide-Binding Specificity of HRL40
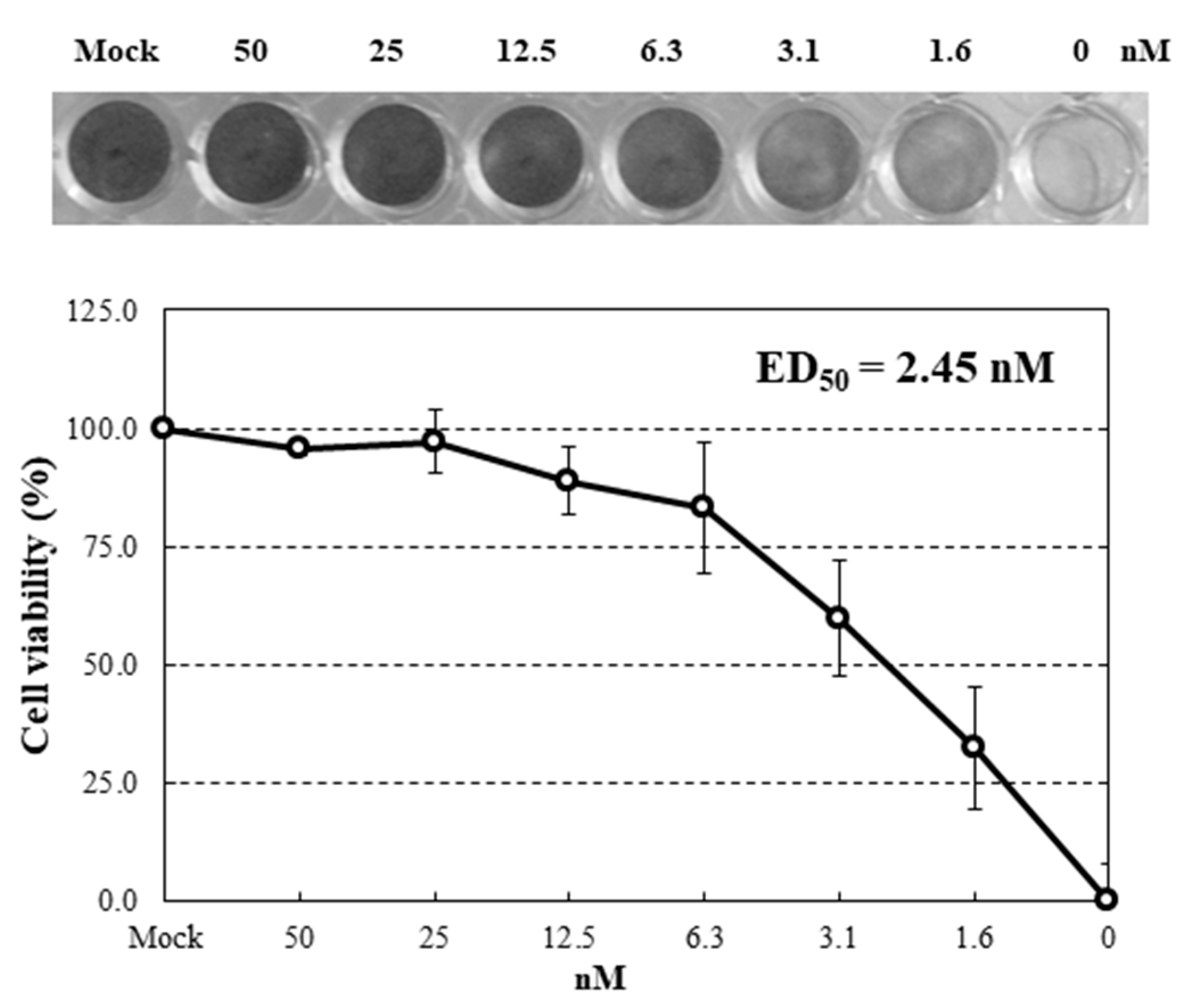
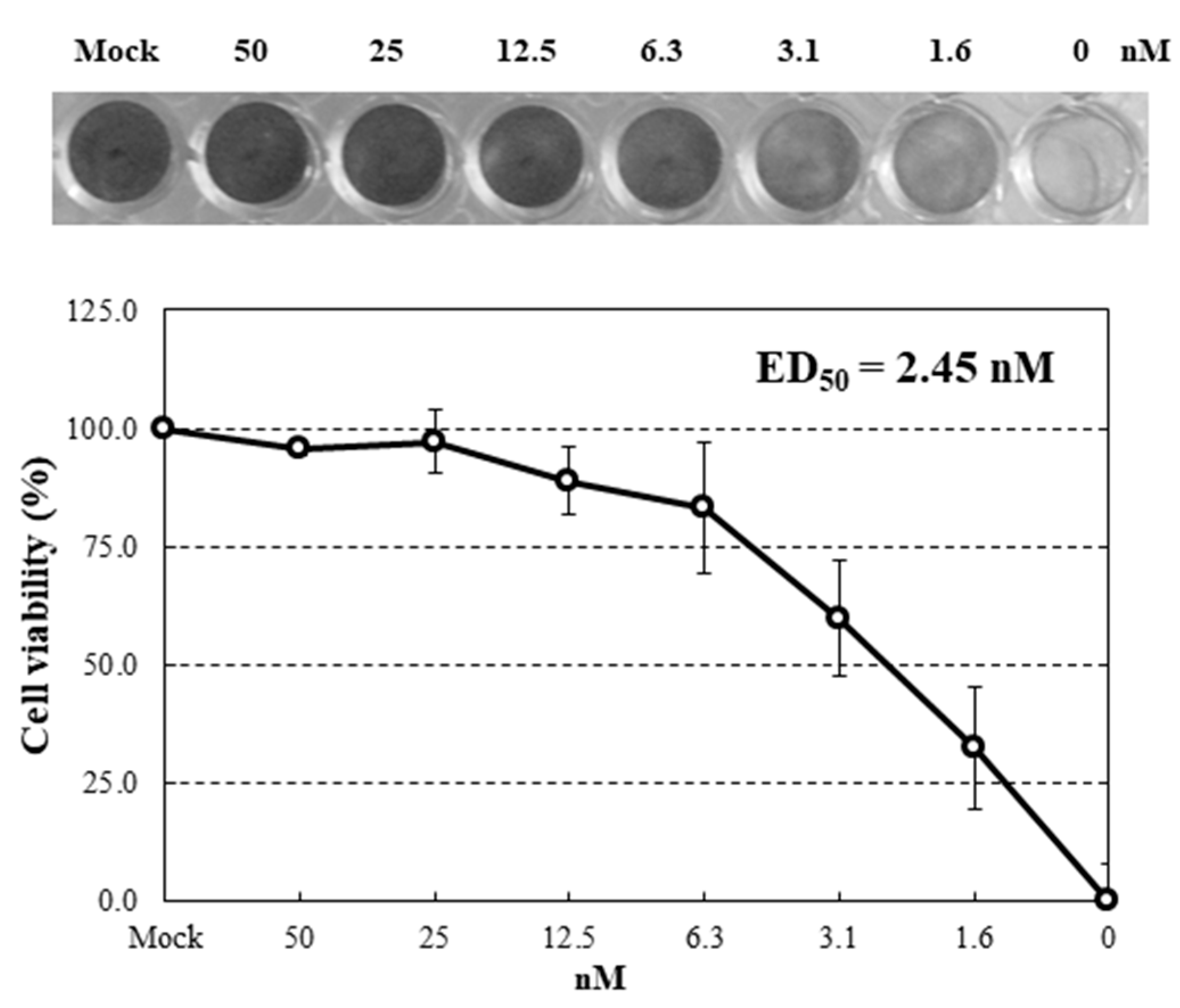
2.4. Inhibition of Virus Infection by HRL40
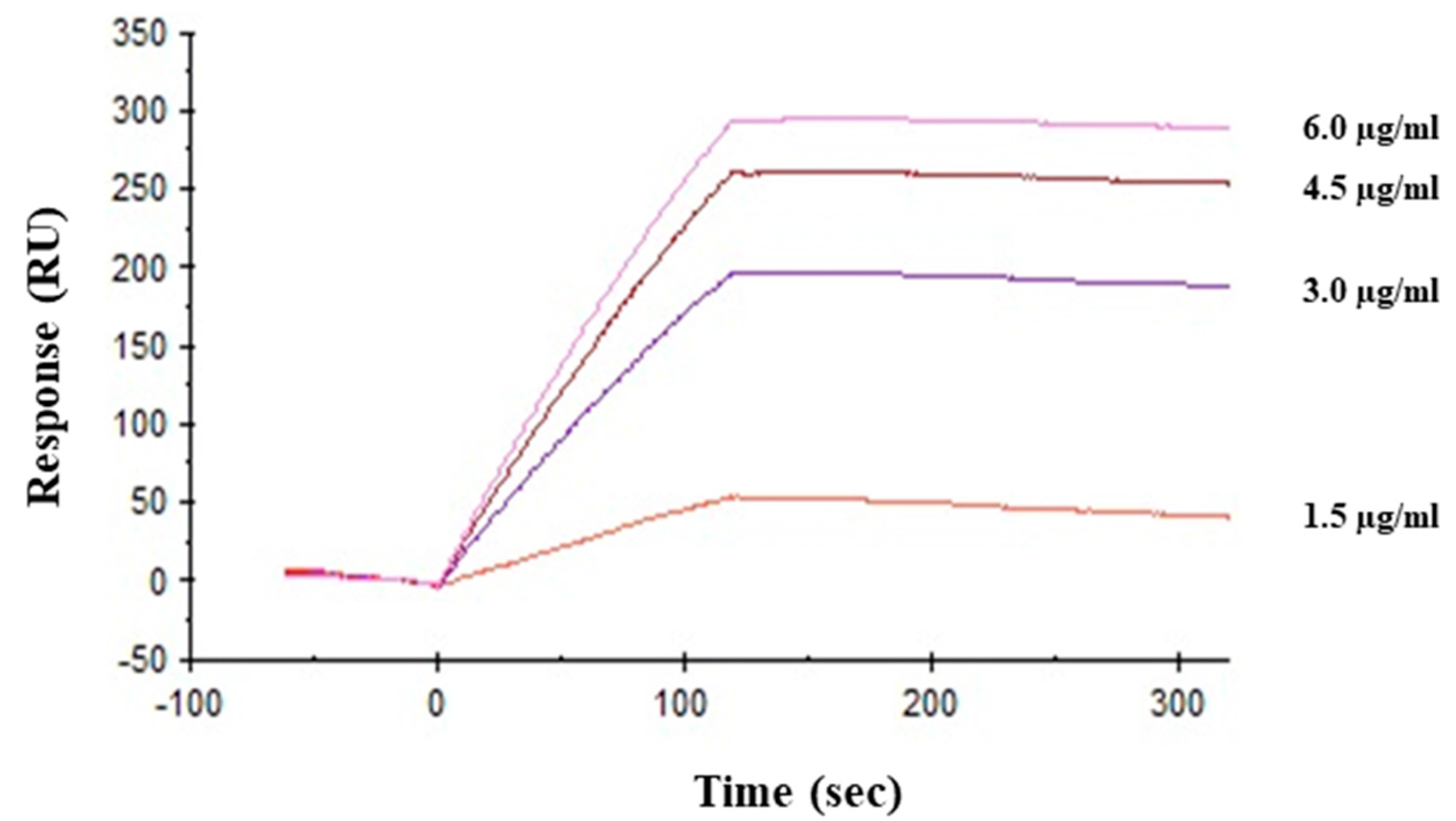
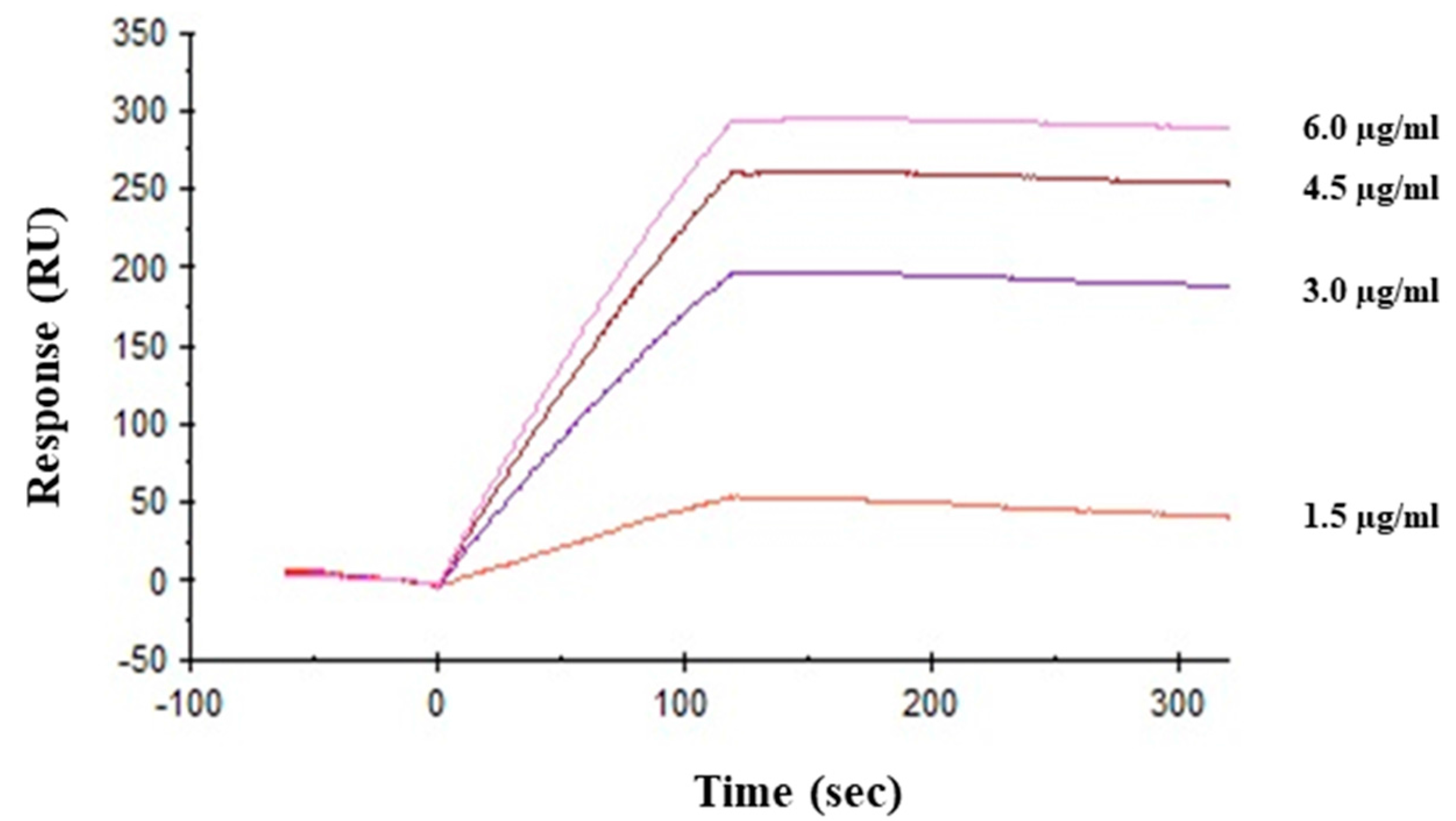
2.5. Interaction between HRL40 and Influenza Virus Envelope Hemagglutinin
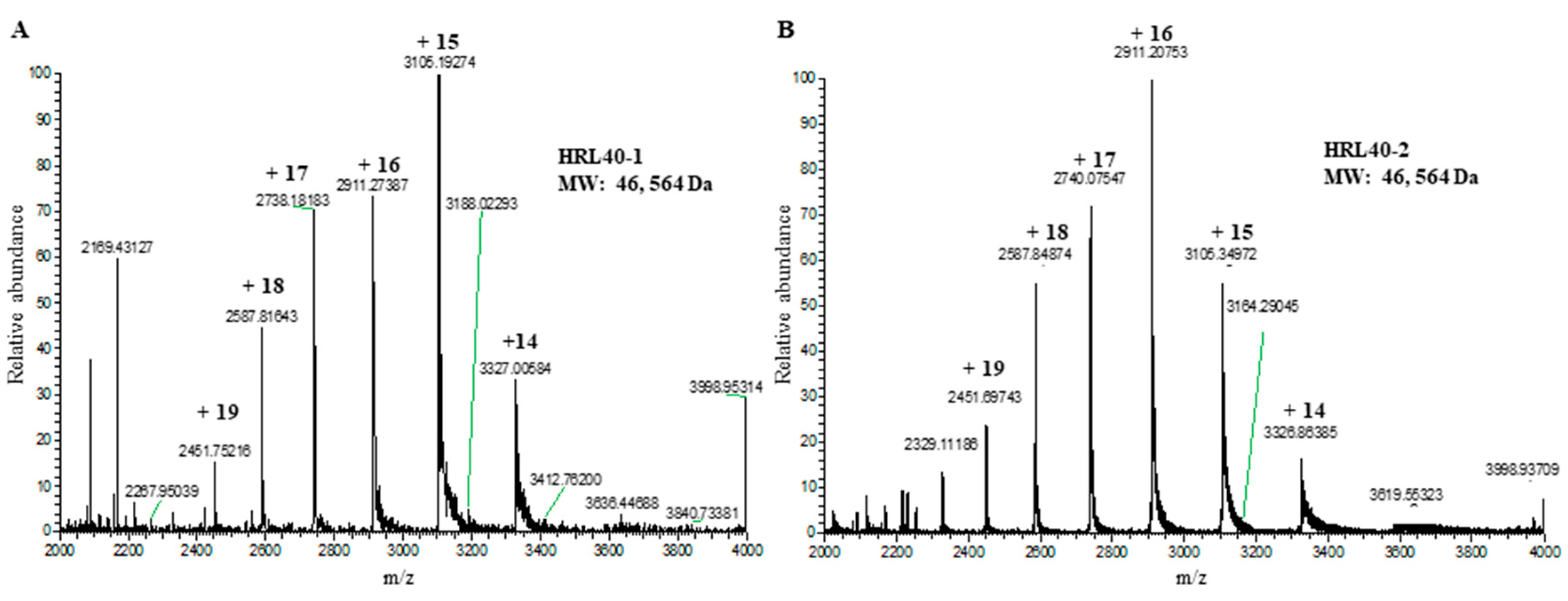
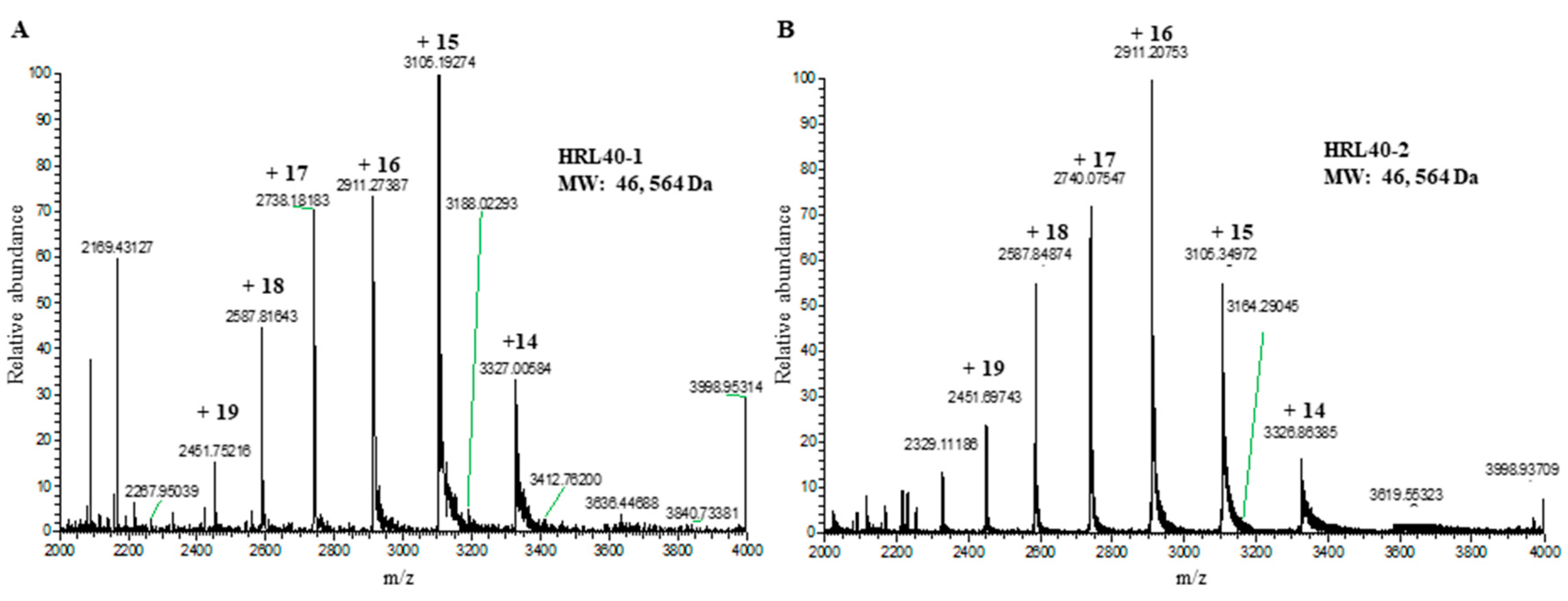
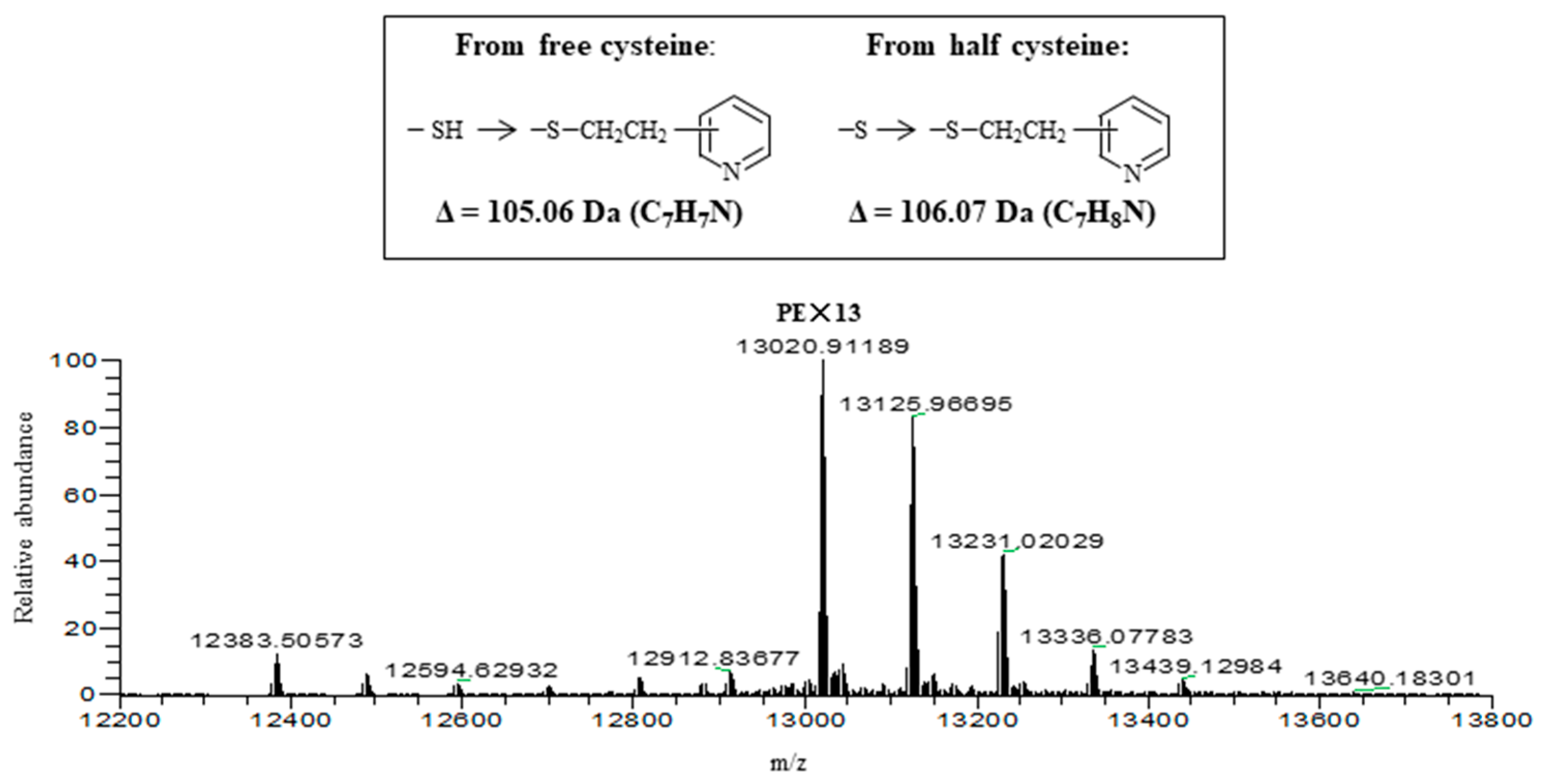
2.6. Molecular Structure of HRL40
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Extraction and Purification of Lectin HRL40
4.3. Determination of Protein Content
4.4. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
4.5. Hemagglutination and Hemagglutination-Inhibition Assays
4.6. Oligosaccharide-Binding Analysis
4.7. Molecular Weight Determination of HRL40
4.8. Preparation of S-Pyridylethylated Lectins
4.9. Anti-Influenza Activity Test
4.10. Surface Plasmon Resonance Analysis of Interaction between HRL40 and Influenza Viral Hemagglutinin
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sharon, N.; Lis, H. Lectins, 2nd ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Hori, K.; Matsubara, K.; Miyazawa, K. Primary structures of two hemagglutinins from the marine red alga, Hypnea japonica. Biochim. Biophys. Acta 2000, 1474, 226–236. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, F.-D.; Zhang, Y.-J.; Wu, Z.-J.; Lin, Q.-Y.; Xie, L.-H. Molecular characterization of a new lectin from the marine alga Ulva pertusa. Acta Biochim. Biophys. Sin. 2004, 36, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; O’Keefe, B.R.; Sowder, R.C., II; Bringans, S.; Gardella, R.; Berg, S.; Cochran, P.; Turpin, J.A.; Buckheit, R.W.; McMahon, J.B.; et al. Isolation and characterization of griffithsin, a novel HIV-inactivating protein, from the red alga Griffithsia sp. J. Biol. Chem. 2005, 280, 9345–9353. [Google Scholar] [CrossRef] [PubMed]
- Nagano, C.S.; Debray, H.; Nascimento, K.S.; Pinto, V.P.; Cavada, B.S.; Saker-Sampaio, S.; Farias, W.R.; Sampaio, A.H.; Calvete, J.J. HCA and HML isolated from the red marine algae Hypnea cervicornis and Hypnea musciformis define a novel lectin family. Protein Sci. 2005, 14, 2167–2176. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Sato, Y.; Ito, K.; Fujiwara, Y.; Iwamoto, Y.; Makino, H.; Kawakabo, A. Strict specificity for high mannose-type N-glycans and primary structure of a red alga Eucheuma serra lectin. Glycobiology 2007, 17, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, S.; Nakamura-Tsuruta, S.; Tateno, H.; Hirabayashi, J.; Matsubara, K.; Hori, K. Strict binding specificity of small-sized lectins from the red alga Hypnea japonica for core (α1-6) fucosylated N-glycans. Biosci. Biotechnol. Biochem. 2009, 73, 912–920. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Hirayama, M.; Morimoto, K.; Yamamoto, N.; Okuyama, S.; Hori, K. High mannose-binding lectin with preference for the cluster of alpha 1–2-mannose from the green alga Boodlea coacta is a potent entry inhibitor of HIV-1 and influenza viruses. J. Biol. Chem. 2011, 286, 19446–19458. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Morimoto, K.; Hirayama, M.; Hori, K. High mannose-specific lectin (KAA-2) from the red alga Kappaphycus alvarezii potently inhibits influenza virus infection in a strain-independent manner. Biochem. Biophys. Res. Commun. 2011, 405, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Hung, L.D.; Sato, Y.; Hori, K. High-mannose N-glycan-specific lectin from the red alga Kappaphycus striatum (Carrageenophyte). Phytochemistry 2011, 72, 855–861. [Google Scholar] [CrossRef] [PubMed]
- Praseptiangga, D.; Hirayama, M.; Hori, K. Purification, characterization, and cDNA cloning of a novel lectin from the green alga, Codium barbatum. Biosci. Biotechnol. Biochem. 2012, 76, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, M.; Shibata, H.; Imamura, K.; Sakaguchi, T.; Hori, K. High-mannose specific lectin and its recombinants from a carrageenophyte Kappaphycus alvarezii represent a potent anti-HIV activity through high-affinity binding to the viral envelope glycprotein gp120. Mar. Biotechnol. 2016, 18, 144–160. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Morimoto, K.; Kubo, T.; Sakaguchi, T.; Nishizono, A.; Hirayama, M.; Hori, K. Entry inhibition of influenza viruses with high mannose binding lectin ESA-2 from the red alga Eucheuma serra through the recognition of viral hemagglutinin. Mar. Drugs 2015, 13, 3454–3465. [Google Scholar] [CrossRef] [PubMed]
- Boyd, M.R.; Gustafson, K.R.; McMahon, J.B.; Shoemaker, R.H.; O’Keefe, B.R.; Mori, T.; Gulakowski, R.J.; Wu, L.; Rivera, M.I.; Laurencot, C.M.; et al. Discovery of cyanovirin-N, a novel human immunodeficiency virus-inactivating protein that binds viral surface envelope glycoprotein gp120: Potential applications to microbicide development. Antimicrob. Agents Chemother. 1997, 41, 1521–1530. [Google Scholar] [PubMed]
- Sato, Y.; Okuyama, S.; Hori, K. Primary structure and carbohydrate binding specificity of a potent anti-HIV lectin isolated from the filamentous cyanobacterium, Osillatoria agardhii. J. Biol. Chem. 2007, 282, 11021–11029. [Google Scholar] [CrossRef] [PubMed]
- Huskens, D.; Férir, G.; Vermeire, K.; Kehr, J.C.; Balzarini, J.; Dittmann, E.; Schols, D. Microvirin, a novel α(1,2)-mannose-specific lectin isolated from Microcystis aeruginosa, has anti-HIV-1 activity comparable with that of cyanovirin-N but a much higher safety profile. J. Biol. Chem. 2010, 285, 24845–24854. [Google Scholar] [CrossRef] [PubMed]
- Bewley, C.A.; Cai, M.; Ray, S.; Ghirlando, R.; Yamaguchi, M.; Muramoto, K. New carbohydrate specificity and HIV-1 fusion blocking activity of the cyanobacterial protein MVL: NMR, ITC and sedimentation equilibrium studies. J. Mol. Biol. 2004, 339, 901–914. [Google Scholar] [CrossRef] [PubMed]
- Bokesch, H.R.; O’Keefe, B.R.; McKee, T.C.; Pannell, L.K.; Patterson, G.M.; Gardella, R.S.; Sowder, R.C.; Turpin, J.; Watson, K.; Buckheit, R.W., Jr.; et al. A potent novel anti-HIV protein from the cultured cyanobacterium Scytonema varium. Biochemistry 2003, 42, 2578–2584. [Google Scholar] [CrossRef] [PubMed]
- Ziółkowska, N.E.; Wlodawer, A. Structural studies of algal lectins with anti-HIV activity. Acta Biochim. Pol. 2006, 53, 617–626. [Google Scholar] [PubMed]
- Huskens, D.; Schols, D. Algal lectins as potential HIV microbicide candidates. Mar. Drugs 2012, 10, 1476–1497. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.A.; Ramessar, K.; O’Keefe, B.R. Antiviral lectins: Selective inhibitors of viral entry. Antiviral Res. 2017, 142, 37–54. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, B.R.; Smee, D.F.; Turpin, J.A.; Saucedo, C.J.; Gustafson, K.R.; Mori, T.; Blakeslee, D.; Buckheit, R.; Boyd, M.R. Potent anti-influenza activity of cyanovirin-N and interactions with viral hemagglutinin. Antimicrob. Agents Chemother. 2003, 47, 2518–2525. [Google Scholar] [CrossRef] [PubMed]
- Mir-Sherari, S.Y.; Ashford, D.A.; Harvey, D.J.; Dwek, R.A.; Schulza, I.T. The glycosylation of the influenza A virus hemagglutinin by mammalian cells. A site-specific study. J. Biol. Chem. 1997, 272, 4027–4036. [Google Scholar] [CrossRef]
- Skehel, J.J.; Wiley, D.C. Receptor binding and membrane fusion in virus entry: The influenza hemagglutinin. Annu. Rev. Biochem. 2000, 69, 531–569. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Gaschen, B.; Blay, W.; Foley, B.; Haigwood, N.; Kuiken, C.; Korber, B. Tracking global patterns of N-linked glycosylation site variation in highly variable viral glycoproteins: HIV, SIV, and HCV envelopes and influenza hemagglutinin. Glycobiology 2004, 14, 1229–1246. [Google Scholar] [CrossRef] [PubMed]
- Férir, G.; Gordts, S.C.; Schols, D. HIV-1 and its resistance to peptidic carbohydrate-binding agents (CBAs): An overview. Molecules 2014, 19, 21085–21112. [Google Scholar] [CrossRef] [PubMed]
- Koharudin, L.M.; Gronenborn, A.M. Antiviral lectins as potential HIV microbicides. Curr. Opin. Virol. 2014, 7, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Hirayama, M. Centrifugal ultrafiltration-HPLC method for interaction analysis between lectins and sugars. Methods Mol. Biol. 2014, 1200, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Hori, K. Cloning, expression, and characterization of a novel anti-HIV lectin from the cultured cyanobacterium, Oscillatoria agardhii. Fish. Sci. 2009, 75, 743–753. [Google Scholar] [CrossRef]
- Hori, K. Potential of algal lectins as functional materials. Biosci. Ind. 2013, 71, 129–133. [Google Scholar]
- Romeo, J.M.; Esmon, B.; Zusman, D.R. Nucleotide sequence of the myxobacterial hemagglutinin gene contains four homologous domains. Proc. Natl. Acad. Sci. USA 1986, 83, 6332–6336. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Morimoto, K.; Kubo, T.; Yanagihara, K.; Seyama, T. High mannose-binding antiviral lectin PFL from Pseudomonas fluorescens Pf0-1 promotes cell death of gastric cancer cell MKN28 via interaction with alpha2-integrin. PLoS ONE 2012, 7, e45922. [Google Scholar] [CrossRef] [PubMed]
- Whitley, M.J.; Furey, W.; Kollipara, S.; Gronenborn, A.M. Burkholderia oklahomensis agglutinin is a canonical two-domain OAA-family lectin: Structures, carbohydrate binding, and anti-HIV activity. FEBS J. 2013, 280, 2056–2067. [Google Scholar] [CrossRef] [PubMed]
- Koharudin, L.M.; Kollipara, S.; Aiken, C.; Gronenborn, A.M. Structural insights into the anti-HIV activity of the Oscillatoria agardhii agglutinin homolog lectin family. J. Biol. Chem. 2012, 287, 33796–33811. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, K.; Sato, Y. Anti-influenza virus activity of high-mannose binding lectins derived from genus Pseudomonas. Virus Res. 2016, 223, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Koharudin, L.M.; Furey, W.; Gronenborn, A.M. Novel fold and carbohydrate specificity of the potent anti-HIV cyanobacterial lectin from Oscillatoria agardhii. J. Biol. Chem. 2011, 286, 1588–1597. [Google Scholar] [CrossRef] [PubMed]
- Koharudin, L.M.; Gronenborn, A.M. Structural basis of the anti-HIV activity of the cyanobacterium Oscillatoria agardhii agglutinin. Structure 2011, 19, 1170–1181. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Miyazawa, K.; Ito, K. Hypnins, low-molecular weight peptidic agglutinins isolated from a marine red alga, Hypnea japonica. Biochim. Biophys. Acta 1986, 873, 228–236. [Google Scholar] [CrossRef]
- Schägger, H.; Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef]
- Hori, K.; Miyazawa, K.; Ito, K. Preliminary characterization of agglutinins from seven marine algal species. Bull. Jpn. Soc. Sci. Fish 1986, 52, 323–331. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Procedure | Volume (mL) | Protein a (mg/mL) | HA b | THA c | MAC d (μg/mL) |
|---|---|---|---|---|---|
| Extraction | 4000 | 7.0 | 32 | 128,000 | 218.8 |
| Salting-out | 100 | 20.0 | 1024 | 102,400 | 19.5 |
| HP e (stepwise) | 94 | 3.2 | 128 | 12,032 | 25.0 |
| HP (gradient) | 75 | 1.6 | 64 | 4800 | 25.0 |
| GF f | 26 | 0.5 | 64 | 1664 | 7.8 |
| IE g | 1 | 1.3 | 64 | 64 | 20.3 |
| Sugars and Glycoproteins a | Salting-Out | HP b (Stepwise) | HP (Gradient) | GF c | HRL40 |
|---|---|---|---|---|---|
| Sugars (mM) | |||||
| Monosaccharides d | >100 | >100 | >100 | >100 | >100 |
| Disaccharide | |||||
| Lactose | >100 | >100 | >100 | >100 | >100 |
| Glycoproteins (μg/mL) | |||||
| N-glycan-linked | |||||
| Complex type | |||||
| Transferrin | >1000 | >1000 | >1000 | >1000 | >1000 |
| Asialo-transferrin | >1000 | >1000 | >1000 | >1000 | >1000 |
| High-mannose type | |||||
| Yeast mannan | >1000 | >1000 | 31.3 | 31.3 | 15.6 |
| Complex and high-mannose types | |||||
| Porcine thyroglobulin (PTG) | 250 | 250 | 62.5 | 125 | 62.5 |
| Asialo-PTG | 250 | 500 | 250 | 500 | 250 |
| N/O-glycan-linked | |||||
| Fetuin | >1000 | >1000 | 1000 | >1000 | >1000 |
| Asialo-fetuin | >1000 | >1000 | >1000 | >1000 | 1000 |
| O-glycan-linked | |||||
| Bovine submaxillary mucin (BSM) | >1000 | >1000 | 1000 | >1000 | 1000 |
| Asialo-BSM | >1000 | >1000 | 1000 | 1000 | 1000 |
| ka (M−1s−1) | kd (s−1) | KA (M−1) | KD (M) | |
|---|---|---|---|---|
| HRL40 | 7.14 × 104 | 2.63 × 10−6 | 2.71 × 1010 | 3.69 × 10−11 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, J.; Hirayama, M.; Sato, Y.; Morimoto, K.; Hori, K. A Novel High-Mannose Specific Lectin from the Green Alga Halimeda renschii Exhibits a Potent Anti-Influenza Virus Activity through High-Affinity Binding to the Viral Hemagglutinin. Mar. Drugs 2017, 15, 255. https://doi.org/10.3390/md15080255
Mu J, Hirayama M, Sato Y, Morimoto K, Hori K. A Novel High-Mannose Specific Lectin from the Green Alga Halimeda renschii Exhibits a Potent Anti-Influenza Virus Activity through High-Affinity Binding to the Viral Hemagglutinin. Marine Drugs. 2017; 15(8):255. https://doi.org/10.3390/md15080255
Chicago/Turabian StyleMu, Jinmin, Makoto Hirayama, Yuichiro Sato, Kinjiro Morimoto, and Kanji Hori. 2017. "A Novel High-Mannose Specific Lectin from the Green Alga Halimeda renschii Exhibits a Potent Anti-Influenza Virus Activity through High-Affinity Binding to the Viral Hemagglutinin" Marine Drugs 15, no. 8: 255. https://doi.org/10.3390/md15080255
APA StyleMu, J., Hirayama, M., Sato, Y., Morimoto, K., & Hori, K. (2017). A Novel High-Mannose Specific Lectin from the Green Alga Halimeda renschii Exhibits a Potent Anti-Influenza Virus Activity through High-Affinity Binding to the Viral Hemagglutinin. Marine Drugs, 15(8), 255. https://doi.org/10.3390/md15080255