

Whole Genome Sequencing of a Chlamydia trachomatis Strain Responsible for a Case of Rectal Lymphogranuloma Venereum in Italy

 , ,
, ,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Strain Isolation and LGV Diagnosis
2.2. Whole Genome Sequencing and Subsequent Analyses
2.3. Data Availability
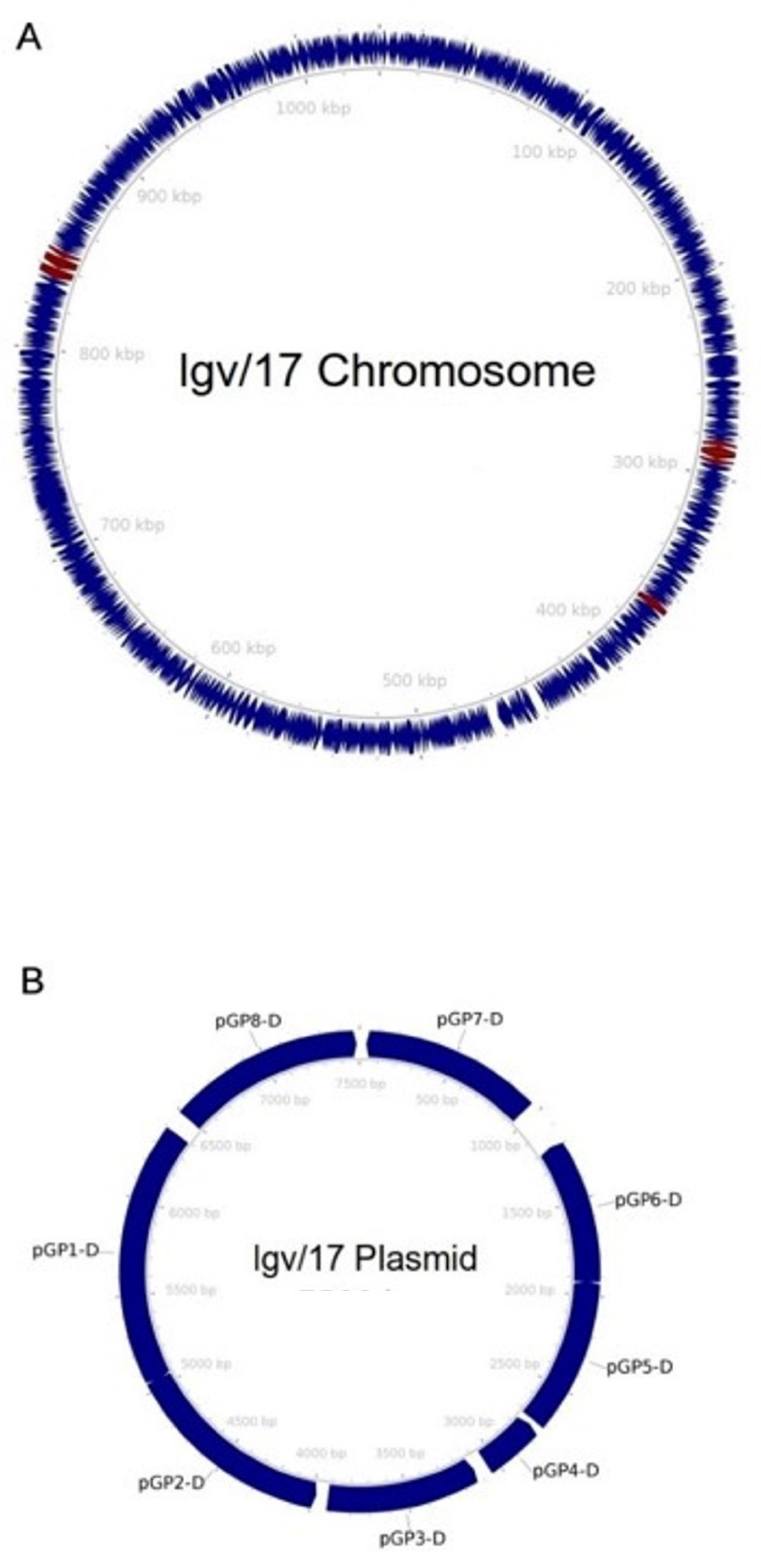
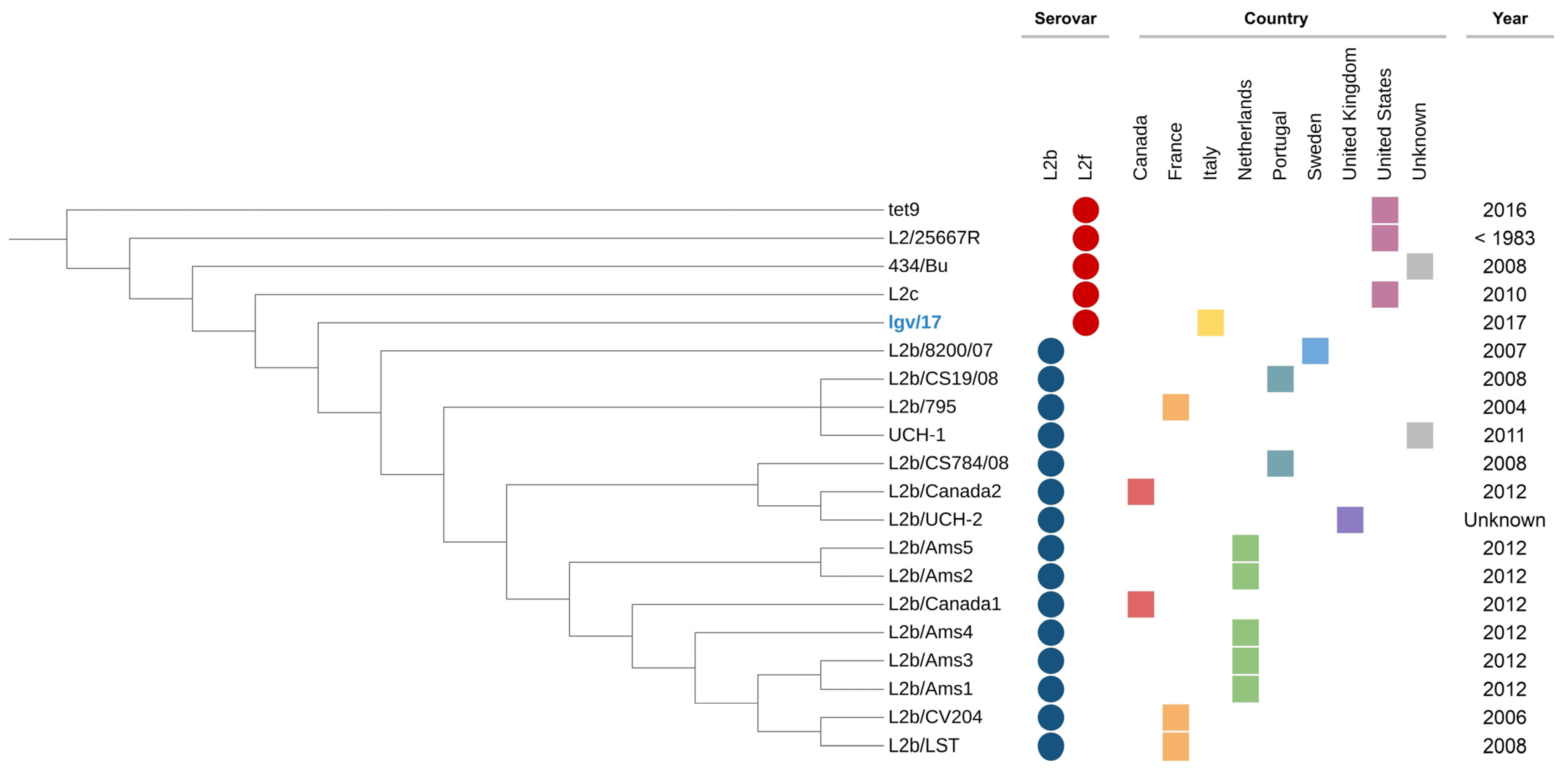
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stary, G.; Meyer, T.; Bangert, C.; Kohrgruber, N.; Gmeinhart, B.; Kirnbauer, R.; Jantschitsch, C.; Rieger, A.; Stary, A.; Geusau, A. New Chlamydia trachomatis L2 strains identified in a recent outbreak of lymphogranuloma venereum in Vienna, Austria. Sex Transm. Dis. 2008, 35, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Marangoni, A.; Foschi, C.; Tartari, F.; Gaspari, V.; Re, M.C. Lymphogranuloma venereum genovariants in men having sex with men in Italy. Sex Transm. Infect. 2021, 97, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Childs, T.; Simms, I.; Alexander, S.; Eastick, K.; Hughes, G.; Field, N. Rapid increase in lymphogranuloma venereum in men who have sex with men, United Kingdom, 2003 to September 2015. Euro Surveill. 2015, 20, 30076. [Google Scholar] [CrossRef] [PubMed]
- De Vrieze, N.H.; de Vries, H.J. Lymphogranuloma venereum among men who have sex with men. An epidemiological and clinical review. Expert. Rev. Anti. Infect. Ther. 2014, 12, 697–704. [Google Scholar] [CrossRef]
- Savage, E.J.; van de Laar, M.J.; Gallay, A.; van der Sande, M.; Hamouda, O.; Sasse, A.; Hoffmann, S.; Diez, M.; Borrego, M.J.; Lowndes, C.M.; et al. European Surveillance of Sexually Transmitted Infections (ESSTI) network. Lymphogranuloma venereum in Europe, 2003–2008. Euro Surveill. 2009, 14, 19428. [Google Scholar] [CrossRef]
- Foschi, C.; Marangoni, A.; D’Antuono, A.; Nardini, P.; Compri, M.; Bellavista, S.; Filippini, A.; Bacchi Reggiani, M.L.; Cevenini, R. Prevalence and predictors of Lymphogranuloma venereum in a high risk population attending a STD outpatients clinic in Italy. BMC Res. Notes 2014, 7, 225. [Google Scholar] [CrossRef]
- Hughes, Y.; Chen, M.Y.; Fairley, C.K.; Hocking, J.S.; Williamson, D.; Ong, J.J.; De Petra, V.; Chow, E.P.F. Universal lymphogranuloma venereum (LGV) testing of rectal chlamydia in men who have sex with men and detection of asymptomatic LGV. Sex. Transm. Infect. 2022, 98, 582–585. [Google Scholar] [CrossRef]
- Christerson, L.; de Vries, H.J.; de Barbeyrac, B.; Gaydos, C.A.; Henrich, B.; Hoffmann, S.; Schachter, J.; Thorvaldsen, J.; Vall-Mayans, M.; Klint, M.; et al. Typing of lymphogranuloma venereum Chlamydia trachomatis strains. Emerg. Infect. Dis. 2010, 16, 1777–1779. [Google Scholar] [CrossRef]
- Isaksson, J.; Carlsson, O.; Airell, Å.; Strömdahl, S.; Bratt, G.; Herrmann, B. Lymphogranuloma venereum rates increased and Chlamydia trachomatis genotypes changed among men who have sex with men in Sweden 2004-2016. J. Med. Microbiol. 2017, 66, 1684–1687. [Google Scholar] [CrossRef]
- Borges, V.; Cordeiro, D.; Salas, A.I.; Lodhia, Z.; Correia, C.; Isidro, J.; Fernandes, C.; Rodrigues, A.M.; Azevedo, J.; Alves, J.; et al. Chlamydia trachomatis: When the virulence-associated genome backbone imports a prevalence-associated major antigen signature. Microb. Genom. 2019, 5, e000313. [Google Scholar] [CrossRef]
- Rodríguez-Domínguez, M.; Puerta, T.; Menéndez, B.; González-Alba, J.M.; Rodríguez, C.; Hellín, T.; Vera, M.; González-Sainz, F.J.; Clavo, P.; Villa, M.; et al. Clinical and epidemiological characterization of a lymphogranuloma venereum outbreak in Madrid, Spain: Co-circulation of two variants. Clin. Microbiol. Infect. 2014, 20, 219–225. [Google Scholar] [CrossRef]
- Morré, S.A.; Spaargaren, J.; Fennema, J.S.; de Vries, H.J.; Coutinho, R.A.; Peña, A.S. Real-time polymerase chain reaction to diagnose lymphogranuloma venereum. Emerg. Infect. Dis. 2005, 11, 1311–1312. [Google Scholar] [CrossRef]
- Petrovay, F.; Balla, E.; Németh, I.; Gönczöl, É. Genotyping of Chlamydia trachomatis from the endocervical specimens of high-risk women in Hungary. J. Med. Microbiol. 2009, 58 Pt 6, 760–764. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Elwell, C.; Mirrashidi, K.; Engel, J. Chlamydia cell biology and pathogenesis. Nat. Rev. Microbiol. 2016, 14, 385–400. [Google Scholar] [CrossRef]
- Foschi, C.; Bortolotti, M.; Polito, L.; Marangoni, A.; Zalambani, C.; Liparulo, I.; Fato, R.; Bolognesi, A. Insights into penicillin-induced Chlamydia trachomatis persistence. Microb. Pathog. 2020, 142, 104035. [Google Scholar] [CrossRef]
- Marangoni, A.; Foschi, C.; Nardini, P.; Compri, M.; Cevenini, R. Evaluation of the Versant CT/GC DNA 1.0 assay (kPCR) for the detection of extra-genital Chlamydia trachomatis and Neisseria gonorrhoeae infections. PLoS ONE 2015, 10, e0120979. [Google Scholar] [CrossRef]
- Gallo Vaulet, L.; Entrocassi, C.; Corominas, A.I.; Rodriguez Fermepin, M. Distribution study of Chlamydia trachomatis genotypes in symptomatic patients in Buenos Aires, Argentina: Association between genotype E and neonatal conjunctivitis. BMC Res. Notes 2010, 3, 34. [Google Scholar] [CrossRef]
- Marangoni, A.; Fiorino, E.; Gilardi, F.; Aldini, R.; Scotti, E.; Nardini, P.; Foschi, C.; Donati, M.; Montagnani, M.; Cevenini, M.; et al. Chlamydia pneumoniae acute liver infection affects hepatic cholesterol and triglyceride metabolism in mice. Atherosclerosis 2015, 41, 471–479. [Google Scholar] [CrossRef]
- Marziali, G.; Marangoni, A.; Foschi, C.; Re, M.C.; Calonghi, N. Effect of Sugars on Chlamydia trachomatis Infectivity. Pathogens 2020, 9, 298. [Google Scholar] [CrossRef]
- Gaibani, P.; Amadesi, S.; Lazzarotto, T.; Ambretti, S. Genome characterization of a Klebsiella pneumoniae co-producing OXA-181 and KPC-121 resistant to ceftazidime/avibactam, meropenem/vaborbactam, imipenem/relebactam and cefiderocol isolated from a critically ill patient. J. Glob. Antimicrob. Resist. 2022, 30, 262–264. [Google Scholar] [CrossRef]
- Vasilevsky, S.; Stojanov, M.; Greub, G.; Baud, D. Chlamydial polymorphic membrane proteins: Regulation, function and potential vaccine candidates. Virulence 2016, 7, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Comanducci, M.; Ricci, S.; Ratti, G. The structure of a plasmid of Chlamydia trachomatis believed to be required for growth within mammalian cells. Mol. Microbiol. 1988, 2, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Patiño, L.H.; Camargo, M.; Muñoz, M.; Ríos-Chaparro, D.I.; Patarroyo, M.A.; Ramírez, J.D. Unveiling the Multilocus Sequence Typing (MLST) Schemes and Core Genome Phylogenies for Genotyping Chlamydia trachomatis. Front. Microbiol. 2018, 9, 1854. [Google Scholar] [CrossRef] [PubMed]
- Feodorova, V.A.; Saltykov, Y.V.; Kolosova, A.A.; Rubanik, L.V.; Poleshchuk, N.N.; Motin, V.L. Emergence of Novel Chlamydia trachomatis Sequence Types among Chlamydia Patients in the Republic of Belarus. Microorganisms 2022, 10, 478. [Google Scholar] [CrossRef]
- Pannekoek, Y.; Morelli, G.; Kusecek, B.; Morré, S.A.; Ossewaarde, J.M.; Langerak, A.A.; van der Ende, A. Multi locus sequence typing of Chlamydiales: Clonal groupings within the obligate intracellular bacteria Chlamydia trachomatis. BMC Microbiol. 2008, 8, 42. [Google Scholar] [CrossRef]
- Dean, D.; Bruno, W.J.; Wan, R.; Gomes, J.P.; Devignot, S.; Mehari, T.; de Vries, H.J.; Morré, S.A.; Myers, G.; Read, T.D.; et al. Predicting phenotype and emerging strains among Chlamydia trachomatis infections. Emerg Infect Dis. 2009, 15, 1385–1394. [Google Scholar] [CrossRef]
- Bom, R.J.; Christerson, L.; Schim van der Loeff, M.F.; Coutinho, R.A.; Herrmann, B.; Bruisten, S.M. Evaluation of high-resolution typing methods for Chlamydia trachomatis in samples from heterosexual couples. J. Clin. Microbiol. 2011, 49, 2844–2853. [Google Scholar] [CrossRef]
- Tan, C.; Hsia, R.C.; Shou, H.; Haggerty, C.L.; Ness, R.B.; Gaydos, C.A.; Dean, D.; Scurlock, A.M.; Wilson, D.P.; Bavoil, P.M. Chlamydia trachomatis-infected patients display variable antibody profiles against the nine-member polymorphic membrane protein family. Infect. Immun. 2009, 77, 3218–3226. [Google Scholar] [CrossRef]
- Gomes, J.P.; Hsia, R.C.; Mead, S.; Borrego, M.J.; Dean, D. Immunoreactivity and differential developmental expression of known and putative Chlamydia trachomatis membrane proteins for biologically variant serovars representing distinct disease groups. Microbes Infect. 2005, 7, 410–420. [Google Scholar] [CrossRef]
- Tan, C.; Hsia, R.C.; Shou, H.; Carrasco, J.A.; Rank, R.G.; Bavoil, P.M. Variable expression of surface-exposed polymorphic membrane proteins in in vitro-grown Chlamydia trachomatis. Cell Microbiol. 2010, 12, 174–187. [Google Scholar] [CrossRef]
- Kiselev, A.O.; Stamm, W.E.; Yates, J.R.; Lampe, M.F. Expression, processing, and localization of PmpD of Chlamydia trachomatis Serovar L2 during the chlamydial developmental cycle. PLoS ONE 2007, 2, e568. [Google Scholar] [CrossRef]
- Gong, S.; Yang, Z.; Lei, L.; Shen, L.; Zhong, G. Characterization of Chlamydia trachomatis plasmid-encoded open reading frames. J. Bacteriol. 2013, 195, 3819–3826. [Google Scholar] [CrossRef]
- Carlson, J.H.; Whitmire, W.M.; Crane, D.D.; Wicke, L.; Virtaneva, K.; Sturdevant, D.E.; Kupko, J.J., III; Porcella, S.F.; Martinez-Orengo, N.; Heinzen, R.A.; et al. The Chlamydia trachomatis plasmid is a transcriptional regulator of chromosomal genes and a virulence factor. Infect. Immun. 2008, 76, 2273–2283. [Google Scholar] [CrossRef]
- Song, L.; Carlson, J.H.; Whitmire, W.M.; Kari, L.; Virtaneva, K.; Sturdevant, D.E.; Watkins, H.; Zhou, B.; Sturdevant, G.L.; Porcella, S.F.; et al. Chlamydia trachomatis plasmid-encoded Pgp4 is a transcriptional regulator of virulence-associated genes. Infect. Immun. 2013, 81, 636–644. [Google Scholar] [CrossRef]
- Wang, Y.; Cutcliffe, L.T.; Skilton, R.J.; Persson, K.; Bjartling, C.; Clarke, I.N. Transformation of a plasmid-free, genital tract isolate of Chlamydia trachomatis with a plasmid vector carrying a deletion in CDS6 revealed that this gene regulates inclusion phenotype. Pathog. Dis. 2013, 67, 100–103. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gene | Description | Location |
|---|---|---|
| pmpA | polymorphic membrane protein PmpA | 845,095–848,022 |
| pmpB | polymorphic membrane protein PmpB | 839,707–844,956 |
| pmpC | polymorphic membrane protein PmpC | 834,207–839,531 |
| pmpD | polymorphic membrane protein PmpD | 364,682–369,274 |
| pmpE | polymorphic membrane protein PmpE | 291,344–294,241 |
| pmpF | polymorphic membrane protein PmpF | 288,243–291,341 |
| pmpG | polymorphic membrane protein PmpG | 285,037–288,078 |
| pmpH | polymorphic membrane protein PmpH | 281,977–285,006 |
| pmpI | polymorphic membrane protein PmpI | 278,235–280,871 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marangoni, A.; Amadesi, S.; Djusse, M.E.; Foschi, C.; Gaspari, V.; Lazzarotto, T.; Gaibani, P. Whole Genome Sequencing of a Chlamydia trachomatis Strain Responsible for a Case of Rectal Lymphogranuloma Venereum in Italy. Curr. Issues Mol. Biol. 2023, 45, 1852-1859. https://doi.org/10.3390/cimb45030119
Marangoni A, Amadesi S, Djusse ME, Foschi C, Gaspari V, Lazzarotto T, Gaibani P. Whole Genome Sequencing of a Chlamydia trachomatis Strain Responsible for a Case of Rectal Lymphogranuloma Venereum in Italy. Current Issues in Molecular Biology. 2023; 45(3):1852-1859. https://doi.org/10.3390/cimb45030119
Chicago/Turabian StyleMarangoni, Antonella, Stefano Amadesi, Marielle Ezekielle Djusse, Claudio Foschi, Valeria Gaspari, Tiziana Lazzarotto, and Paolo Gaibani. 2023. "Whole Genome Sequencing of a Chlamydia trachomatis Strain Responsible for a Case of Rectal Lymphogranuloma Venereum in Italy" Current Issues in Molecular Biology 45, no. 3: 1852-1859. https://doi.org/10.3390/cimb45030119
APA StyleMarangoni, A., Amadesi, S., Djusse, M. E., Foschi, C., Gaspari, V., Lazzarotto, T., & Gaibani, P. (2023). Whole Genome Sequencing of a Chlamydia trachomatis Strain Responsible for a Case of Rectal Lymphogranuloma Venereum in Italy. Current Issues in Molecular Biology, 45(3), 1852-1859. https://doi.org/10.3390/cimb45030119