
Mapping QTLs for Super-Earliness and Agro-Morphological Traits in RILs Population Derived from Interspecific Crosses between Pisum sativum × P. fulvum

 ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Phenotyping
2.3. Genotyping
2.4. Genetic Mapping
2.5. QTL Analyses
3. Results
3.1. Phenotypic Characteristics
3.2. Genetic Mapping
3.3. QTLs Analyses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Davis, P.H. Flora of Turkey and the East Aegean Islands.; Davis, P.H., Ed.; University Press: Edinburgh, Scotland, 1970; Volume 3. [Google Scholar]
- Ladizinsky, G.; Abbo, S. The Search for Wild Relatives of Cool Season Legumes; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Smỳkal, P.; Coyne, C.J.; Ambrose, M.J.; Maxted, N.; Schaefer, H.; Blair, M.W.; Berger, J.; Greene, S.L.; Nelson, M.N.; Besharat, N. Legume Crops Phylogeny and Genetic Diversity for Science and Breeding. Crit. Rev. Plant Sci. 2015, 34, 43–104. [Google Scholar] [CrossRef]
- Mendel, G. Experiments in Plant Hybridization (1865). In Verhandlungen Naturforschenden Ver. Brünn Available Online; 1996; Available online: http://old.esp.org/foundations/genetics/classical/gm-65-a.pdf (accessed on 20 December 2022).
- FAOSTAT. Food and Agriculture Organisation Statistics Database. Available online: https://search.library.wisc.edu/database/UWI12320 (accessed on 20 December 2022).
- Dahl, W.J.; Foster, L.M.; Tyler, R.T. Review of the Health Benefits of Peas (Pisum sativum L.). Br. J. Nutr. 2012, 108, S3–S10. [Google Scholar] [CrossRef] [PubMed]
- Smỳkal, P.; Aubert, G.; Burstin, J.; Coyne, C.J.; Ellis, N.T.; Flavell, A.J.; Ford, R.; Hỳbl, M.; Macas, J.; Neumann, P. Pea (Pisum sativum L.) in the Genomic Era. Agronomy 2012, 2, 74–115. [Google Scholar] [CrossRef]
- Toker, C.; Yadav, S.S. Legumes Cultivars for Stress Environments. In Climate change and management of cool season grain legume crops; Springer: Berlin/Heidelberg, Germany, 2010; pp. 351–376. [Google Scholar]
- Blum, A. Plant Breeding for Stress Environments; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Toker, C.; Lluch, C.; Tejera, N.A.; Serraj, R.; Siddique, K.H.M. 23 Abiotic Stresses. Chickpea Breed. Manag. 2007, 474. Available online: https://www.researchgate.net/profile/Cengiz-Toker/publication/285964620_Conventional_breeding_methods/links/5b19399a45851587f2987c0c/Conventional-breeding-methods.pdf#page=500 (accessed on 20 December 2022).
- Masson-Delmotte, V.; Zhai, P.; Pörtner, H.-O.; Roberts, D.C.; Skea, J.; Shukla, P.R.; Pirani, A.; Moufouma-Okia, W.; Péan, C.; Pidcock, R. Global Warming of 1.5 °C: Summary for Policy Makers; IPCC: Geneva, Switzerland, 2018. [Google Scholar]
- Jiang, Y.; Lindsay, D.L.; Davis, A.R.; Wang, Z.; MacLean, D.E.; Warkentin, T.D.; Bueckert, R.A. Impact of Heat Stress on Pod-Based Yield Components in Field Pea (Pisum sativum L.). J. Agron. Crop Sci. 2020, 206, 76–89. [Google Scholar] [CrossRef]
- Parihar, A.K.; Dixit, G.P.; Bohra, A.; Sen Gupta, D.; Singh, A.K.; Kumar, N.; Singh, D.; Singh, N.P. Genetic Advancement in Dry Pea (Pisum sativum L.): Retrospect and Prospect. In Accelerated Plant Breeding, Volume 3; Springer: Berlin/Heidelberg, Germany, 2020; pp. 283–341. [Google Scholar]
- Tafesse, E.G.; Gali, K.K.; Lachagari, V.R.; Bueckert, R.; Warkentin, T.D. Genome-Wide Association Mapping for Heat Stress Responsive Traits in Field Pea. Int. J. Mol. Sci. 2020, 21, 2043. [Google Scholar] [CrossRef]
- Kreplak, J.; Madoui, M.-A.; Cápal, P.; Novák, P.; Labadie, K.; Aubert, G.; Bayer, P.E.; Gali, K.K.; Syme, R.A.; Main, D.; et al. A Reference Genome for Pea Provides Insight into Legume Genome Evolution. Nat. Genet. 2019, 51, 1411–1422. [Google Scholar] [CrossRef] [PubMed]
- Dirlewanger, E.; Isaac, P.G.; Ranade, S.; Belajouza, M.; Cousin, R.; De Vienne, D. Restriction Fragment Length Polymorphism Analysis of Loci Associated with Disease Resistance Genes and Developmental Traits in Pisum sativum L. Theor. Appl. Genet. 1994, 88, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Irzykowska, L.; Wolko, B.; Swiecicki, W.K. Interval Mapping of QTLs Controlling Some Morphological Traits in Pea. Cell. Mol. Biol. Lett. 2002, 7, 417–422. [Google Scholar]
- Tar’an, B.; Warkentin, T.; Somers, D.J.; Miranda, D.; Vandenberg, A.; Blade, S.; Woods, S.; Bing, D.; Xue, A.; DeKoeyer, D. Quantitative Trait Loci for Lodging Resistance, Plant Height and Partial Resistance to Mycosphaerella Blight in Field Pea (Pisum sativum L.). Theor. Appl. Genet. 2003, 107, 1482–1491. [Google Scholar] [CrossRef]
- Prioul, S.; Frankewitz, A.; Deniot, G.; Morin, G.; Baranger, A. Mapping of Quantitative Trait Loci for Partial Resistance to Mycosphaerella Pinodes in Pea (Pisum sativum L.), at the Seedling and Adult Plant Stages. Theor. Appl. Genet. 2004, 108, 1322–1334. [Google Scholar] [CrossRef] [PubMed]
- Timmerman-Vaughan, G.M.; Frew, T.J.; Butler, R.; Murray, S.; Gilpin, M.; Falloon, K.; Johnston, P.; Lakeman, M.B.; Russell, A.; Khan, T. Validation of Quantitative Trait Loci for Ascochyta Blight Resistance in Pea (Pisum sativum L.), Using Populations from Two Crosses. Theor. Appl. Genet. 2004, 109, 1620–1631. [Google Scholar] [CrossRef] [PubMed]
- Loridon, K.; McPhee, K.; Morin, J.; Dubreuil, P.; Pilet-Nayel, M.-L.; Aubert, G.; Rameau, C.; Baranger, A.; Coyne, C.; Lejeune-Henaut, I. Microsatellite Marker Polymorphism and Mapping in Pea (Pisum sativum L.). Theor. Appl. Genet. 2005, 111, 1022–1031. [Google Scholar] [CrossRef]
- Pilet-Nayel, M.-L.; Muehlbauer, F.J.; McGee, R.J.; Kraft, J.M.; Baranger, A.; Coyne, C.J. Consistent Quantitative Trait Loci in Pea for Partial Resistance to Aphanomyces Euteiches Isolates from the United States and France. Phytopathology 2005, 95, 1287–1293. [Google Scholar] [CrossRef] [PubMed]
- Fondevilla, S.; Satovic, Z.; Rubiales, D.; Moreno, M.T.; Torres, A.M. Mapping of Quantitative Trait Loci for Resistance to Mycosphaerella Pinodes in Pisum sativum subsp. syriacum. Mol. Breed. 2008, 21, 439–454. [Google Scholar] [CrossRef]
- Dumont, E.; Fontaine, V.; Vuylsteker, C.; Sellier, H.; Bodèle, S.; Voedts, N.; Devaux, R.; Frise, M.; Avia, K.; Hilbert, J.-L. Association of Sugar Content QTL and PQL with Physiological Traits Relevant to Frost Damage Resistance in Pea under Field and Controlled Conditions. Theor. Appl. Genet. 2009, 118, 1561–1571. [Google Scholar] [CrossRef]
- Fondevilla, S.; Fernández-Aparicio, M.; Satovic, Z.; Emeran, A.A.; Torres, A.M.; Moreno, M.T.; Rubiales, D. Identification of Quantitative Trait Loci for Specific Mechanisms of Resistance to Orobanche Crenata Forsk. in Pea (Pisum sativum L.). Mol. Breed. 2010, 25, 259–272. [Google Scholar] [CrossRef]
- Ubayasena, L.; Bett, K.; Tar’an, B.; Vijayan, P.; Warkentin, T. Genetic Control and QTL Analysis of Cotyledon Bleaching Resistance in Green Field Pea (Pisum sativum L.). Genome 2010, 53, 346–359. [Google Scholar] [CrossRef]
- Feng, J.; Hwang, R.; Chang, K.F.; Conner, R.L.; Hwang, S.F.; Strelkov, S.E.; Gossen, B.D.; McLaren, D.L.; Xue, A.G. Identification of Microsatellite Markers Linked to Quantitative Trait Loci Controlling Resistance to Fusarium Root Rot in Field Pea. Can. J. Plant Sci. 2011, 91, 199–204. [Google Scholar] [CrossRef]
- Jha, A.B.; Tar’an, B.; Stonehouse, R.; Warkentin, T.D. Identification of QTLs Associated with Improved Resistance to Ascochyta Blight in an Interspecific Pea Recombinant Inbred Line Population. Crop Sci. 2016, 56, 2926–2939. [Google Scholar] [CrossRef]
- Timmerman-Vaughan, G.M.; Moya, L.; Frew, T.J.; Murray, S.R.; Crowhurst, R. Ascochyta Blight Disease of Pea (Pisum sativum L.): Defence-Related Candidate Genes Associated with QTL Regions and Identification of Epistatic QTL. Theor. Appl. Genet. 2016, 129, 879–896. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Gali, K.K.; Tar’an, B.; Warkentin, T.D.; Bueckert, R.A. Pea Phenology: Crop Potential in a Warming Environment. Crop Sci. 2017, 57, 1540–1551. [Google Scholar] [CrossRef]
- Krajewski, P.; Bocianowski, J.; Gaw\lowska, M.; Kaczmarek, Z.; Pniewski, T.; Święcicki, W.; Wolko, B. QTL for Yield Components and Protein Content: A Multienvironment Study of Two Pea (Pisum sativum L.) Populations. Euphytica 2012, 183, 323–336. [Google Scholar] [CrossRef]
- Ma, Y.; Coyne, C.J.; Grusak, M.A.; Mazourek, M.; Cheng, P.; Main, D.; McGee, R.J. Genome-Wide SNP Identification, Linkage Map Construction and QTL Mapping for Seed Mineral Concentrations and Contents in Pea (Pisum sativum L.). BMC Plant Biol. 2017, 17, 43. [Google Scholar] [CrossRef] [PubMed]
- Gali, K.K.; Sackville, A.; Tafesse, E.G.; Lachagari, V.R.; McPhee, K.; Hybl, M.; Mikić, A.; Smỳkal, P.; McGee, R.; Burstin, J. Genome-Wide Association Mapping for Agronomic and Seed Quality Traits of Field Pea (Pisum sativum L.). Front. Plant Sci. 2019, 10, 1538. [Google Scholar] [CrossRef]
- Hamon, C.; Coyne, C.J.; McGee, R.J.; Lesné, A.; Esnault, R.; Mangin, P.; Hervé, M.; Le Goff, I.; Deniot, G.; Roux-Duparque, M. QTL Meta-Analysis Provides a Comprehensive View of Loci Controlling Partial Resistance to Aphanomyces Euteichesin Four Sources of Resistance in Pea. BMC Plant Biol. 2013, 13, 1–19. [Google Scholar] [CrossRef]
- Ferrari, B.; Romani, M.; Aubert, G.; Boucherot, K.; Burstin, J.; Pecetti, L.; Huart, M.; Klein, A.; Annicchiarico, P. Association of SNP Markers with Agronomic and Quality Traits of Field Pea in Italy. Czech J. Genet. Plant Breed. 2016, 52, 83–93. [Google Scholar] [CrossRef]
- Gali, K.K.; Liu, Y.; Sindhu, A.; Diapari, M.; Shunmugam, A.S.; Arganosa, G.; Daba, K.; Caron, C.; Lachagari, R.V.; Tar’an, B. Construction of High-Density Linkage Maps for Mapping Quantitative Trait Loci for Multiple Traits in Field Pea (Pisum sativum L.). BMC Plant Biol. 2018, 18, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Gawlowska, M.; Knopkiewicz, M.; Święcicki, W.; Boros, L.; Wawer, A. Quantitative Trait Loci for Stem Strength Properties and Lodging in Two Pea Biparental Mapping Populations. Crop Sci. 2021, 61, 1682–1697. [Google Scholar] [CrossRef]
- Irzykowska, L.; Wolko, B. Interval Mapping of QTLs Controlling Yield-Related Traits and Seed Protein Content in Pisum sativum. J. Appl. Genet. 2004, 45, 297–306. [Google Scholar]
- Timmerman-Vaughan, G.M.; Mills, A.; Whitfield, C.; Frew, T.; Butler, R.; Murray, S.; Michael, L.; John, M.; Adrian, R.; Derek, W. Linkage Mapping of QTL for Seed Yield, Yield Components, and Developmental Traits in Pea. Crop Sci. 2005, 45, 1336–1344. [Google Scholar] [CrossRef]
- Guindon, M.F.; Martin, E.; Cravero, V.; Gali, K.K.; Warkentin, T.D.; Cointry, E. Linkage Map Development by GBS, SSR, and SRAP Techniques and Yield-Related QTLs in Pea. Mol. Breed. 2019, 39, 1–16. [Google Scholar] [CrossRef]
- Sadras, V.O.; Lake, L.; Kaur, S.; Rosewarne, G. Phenotypic and Genetic Analysis of Pod Wall Ratio, Phenology and Yield Components in Field Pea. Field Crop Res. 2019, 241, 107551. [Google Scholar] [CrossRef]
- Aryamanesh, N.; Zeng, Y.; Byrne, O.; Hardie, D.C.; Al-Subhi, A.M.; Khan, T.; Siddique, K.H.M.; Yan, G. Identification of Genome Regions Controlling Cotyledon, Pod Wall/Seed Coat and Pod Wall Resistance to Pea Weevil through QTL Mapping. Theor. Appl. Genet. 2014, 127, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Sari, H.; Sari, D.; Eker, T.; Toker, C. De Novo Super-Early Progeny in Interspecific Crosses Pisum sativum L. × P. fulvum Sibth. et Sm. Sci. Rep. 2021, 11, 19706. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.J. Isolation of Plant DNA from Fresh Tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Van Ooijen, J.W. Multipoint Maximum Likelihood Mapping in a Full-Sib Family of an Outbreeding Species. Genet. Res. 2011, 93, 343–349. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the Graphical Presentation of Linkage Maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef]
- Stuber, C.W.; Edwards, M.D.; Wendel, J.F. Molecular Marker-Facilitated Investigations of Quantitative Trait Loci in Maize. II. Factors Influencing Yield and Its Component Traits1. Crop Sci. 1987, 27, 639–648. [Google Scholar] [CrossRef]
- Takuno, S.; Terauchi, R.; Innan, H. The Power of QTL Mapping with RILs. PLoS ONE 2012, 7, e46545. [Google Scholar] [CrossRef]
- Onyekachi, O.G.; Bonifeca, O.O.; Gemlack, N.F.; Nicholas, N. Abiotic and Biotic Stress in Plants; BoD—Books on Demand: Norderstedt, Germany, 2019; ISBN 978-1-78923-811-2. [Google Scholar]
- Schiermeier, Q. Eat Less Meat: UN Climate-Change Report Calls for Change to Human Diet. Nature 2019, 572, 291–293. [Google Scholar] [CrossRef] [PubMed]
- FAO. The Future of Food and Agriculture–Trends and Challenges. Annual Report; Food Agriculture Organization: Rome, Italy, 2017; pp. 1–180. [Google Scholar]
- FAO; IFAD; UNICEF; WFP; WHO. The State of Food Security and Nutrition in the World 2018: Building Climate Resilience for Food Security and Nutrition; Food & Agriculture Organization: Rome, Italy, 2018; ISBN 978-92-5-130571-3. [Google Scholar]
- White, O.E. Studies of Inheritance in Pisum. II. The Present State of Knowledge of Heredity and Variation in Peas. Proc. Am. Philos. Soc. 1917, 56, 487–588. [Google Scholar]
- Murfet, I.C. Flowering in Pisum. Three Distinct Phenotypic Classes Determined by the Interaction of a Dominant Early and a Dominant Late Gene. Heredity 1971, 26, 243–257. [Google Scholar] [CrossRef]
- King, W.M.; Murfet, I.C. Flowering in Pisum: A Sixth Locus, Dne. Ann. Bot. 1985, 56, 835–846. [Google Scholar] [CrossRef]
- Arumingtyas, E.L.; Murfet, I.C. Flowering in Pisum: A Further Gene Controlling Response to Photoperiod. J. Hered. 1994, 85, 12–17. [Google Scholar] [CrossRef]
- Weller, J.L.; Ortega, R. Genetic Control of Flowering Time in Legumes. Front. Plant Sci. 2015, 6, 207. [Google Scholar] [CrossRef]
- Lejeune-Hénaut, I.; Hanocq, E.; Béthencourt, L.; Fontaine, V.; Delbreil, B.; Morin, J.; Petit, A.; Devaux, R.; Boilleau, M.; Stempniak, J.-J.; et al. The Flowering Locus Hr Colocalizes with a Major QTL Affecting Winter Frost Tolerance in Pisum sativum L. Theor. Appl. Genet. 2008, 116, 1105–1116. [Google Scholar] [CrossRef] [PubMed]
- Liew, L.C.; Hecht, V.; Laurie, R.E.; Knowles, C.L.; Vander Schoor, J.K.; Macknight, R.C.; Weller, J.L. DIE NEUTRALIS and LATE BLOOMER 1 Contribute to Regulation of the Pea Circadian Clock. Plant Cell 2009, 21, 3198–3211. [Google Scholar] [CrossRef]
- Burstin, J.; Marget, P.; Huart, M.; Moessner, A.; Mangin, B.; Duchene, C.; Desprez, B.; Munier-Jolain, N.; Duc, G. Developmental Genes Have Pleiotropic Effects on Plant Morphology and Source Capacity, Eventually Impacting on Seed Protein Content and Productivity in Pea. Plant Physiol. 2007, 144, 768–781. [Google Scholar] [CrossRef]
- Foucher, F.; Morin, J.; Courtiade, J.; Cadioux, S.; Ellis, N.; Banfield, M.J.; Rameau, C. DETERMINATE and LATE FLOWERING Are Two TERMINAL FLOWER1/CENTRORADIALIS Homologs That Control Two Distinct Phases of Flowering Initiation and Development in Pea. Plant Cell 2003, 15, 2742–2754. [Google Scholar] [CrossRef]
- Fondevilla, S.; Almeida, N.F.; Satovic, Z.; Rubiales, D.; Vaz Patto, M.C.; Cubero, J.I.; Torres, A.M. Identification of Common Genomic Regions Controlling Resistance to Mycosphaerella pinodes, Earliness and Architectural Traits in Different Pea Genetic Backgrounds. Euphytica 2011, 182, 43–52. [Google Scholar] [CrossRef]
- Timmerman-Vaughan, G.M.; McCallum, J.A.; Frew, T.J.; Weeden, N.F.; Russell, A.C. Linkage Mapping of Quantitative Trait Loci Controlling Seed Weight in Pea (Pisum sativum L.). Theor. Appl. Genet. 1996, 93, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Tar’an, B.; Warkentin, T.; Somers, D.J.; Miranda, D.; Vandenberg, A.; Blade, S.; Bing, D. Identification of Quantitative Trait Loci for Grain Yield, Seed Protein Concentration and Maturity in Field Pea (Pisum sativum L.). Euphytica 2004, 136, 297–306. [Google Scholar] [CrossRef]
- Valderrama, M.R.; Román, B.; Satovic, Z.; Rubiales, D.; Cubero, J.I.; Torres, A.M. Locating Quantitative Trait Loci Associated with Orobanche Crenata Resistance in Pea. Weed Res. 2004, 44, 323–328. [Google Scholar] [CrossRef]
- Fondevilla, S.; Torres, A.M.; Moreno, M.T.; Rubiales, D. Identification of a New Gene for Resistance to Powdery Mildew in Pisum fulvum, a Wild Relative of Pea. Breed. Sci. 2007, 57, 181–184. [Google Scholar] [CrossRef]




{kind=link}
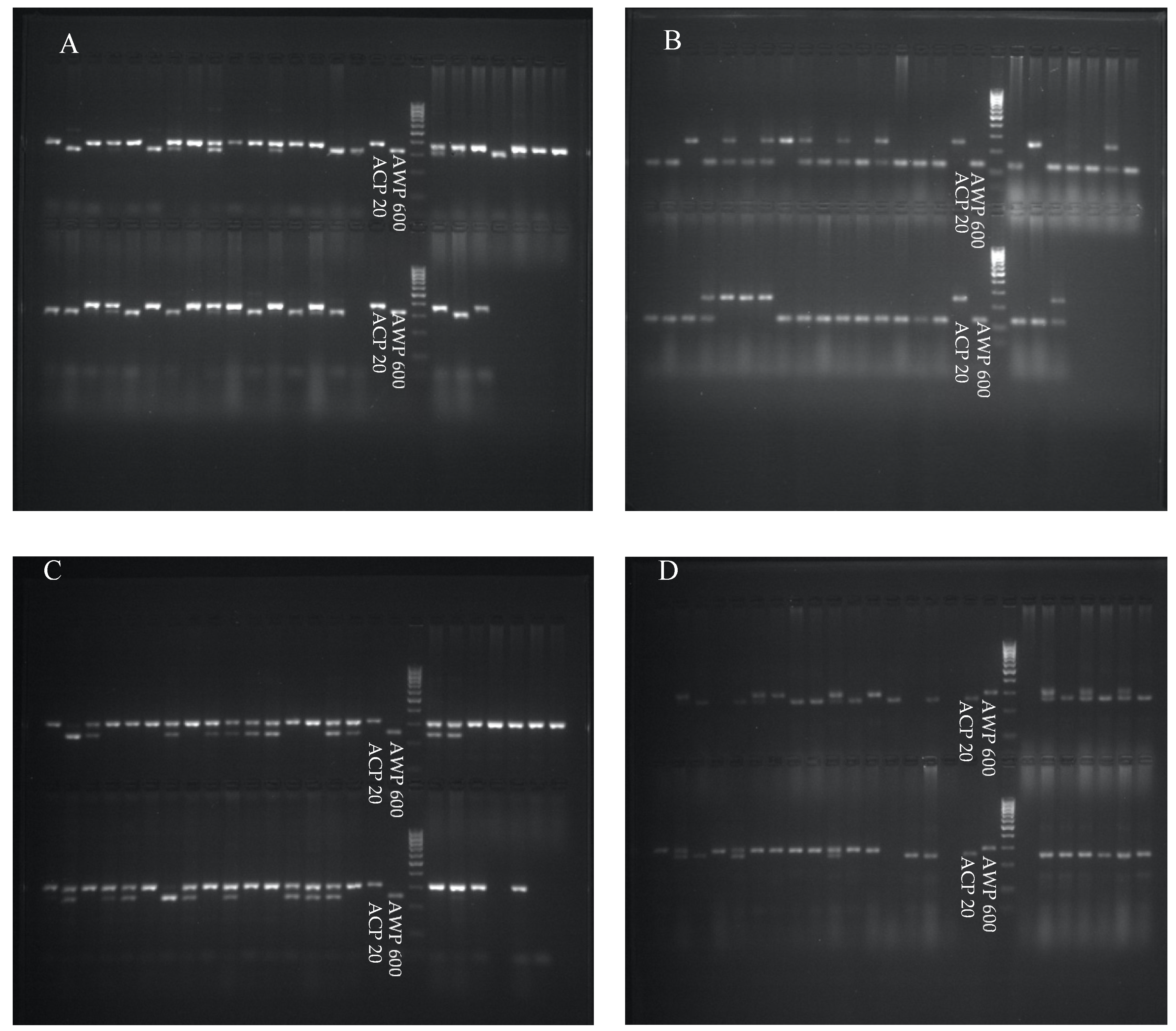{kind=link}
{kind=link}
| Primers | Linkage Groups | Forward (5′-3′) | Reverse (3′-5′) |
|---|---|---|---|
| AD147 | LG1 | AGCCCAAGTTTCTTCTGAATCC | AAATTCGCAGAGCGTTTGTTAC |
| AA474 | LG1 | GCCCACACAAGTGGTTCTATAAAT | ATTAGTCGTTTTTCTGAAACATCAAAG |
| AA67 | LG1 | CCCATGTGAAATTCTCTTGAAGA | GCATTTCACTTGATGAAATTTCG |
| AC75 | LG1 | CGCTCACCAAATGTAGATGATAA | TCATGCATCAATGAAAGTGATAAA |
| D21 | LG1 | TATTCTCCTCCAAAATTTCCTT | GTCAAAATTAGCCAAATTCCTC |
| AB28 | LG1 | CCTGAGTCATCACATAGGAGAT | GCAGAAGTATTTGACTTGATGGAA |
| AA205 | LG2 | TACGCAATCATAGAGTTTGGAA | AATCAAGTCAATGAAACAAGCA |
| AB149 | LG2 | ACAAAGGATGATGAAAGACCCG | TCATTACTCAAAGAATGCACCCAC |
| AB33 | LG2 | CATTGAATTTGTGGGAGAAAGG | TGTGGATGTTGCAATTTCGT |
| AA504 | LG2 | TGAGTGCAGTTGCAATTTCG | TCAGATGAAGAGCATGTGGG |
| AD148 | LG2 | GAAACATCATTGTGTCTTCTTG | TTCCATCACTTGATTGATAAAC |
| AA372.1 | LG2 | GAGTGACCAAAGTTTTGTGAA | CCTTGAACCCATTTTTAAGAGT |
| D23 | LG2 | ATGGTTGTCCCAGGATAGATAA | GAAAACATTGGAGAGTGGAGTA |
| AA153 | LG2 | TTTGATAGTCCGACTTTTCCAT | GTGACAAAAGAATTCAAAACGC |
| AD73 | LG3 | CAGCTGGATTCAATCATTGGTG | ATGAGTAATCCGACGATGCCTT |
| AD270 | LG3 | CTCATCTGATGCGTTGGATTAG | AGGTTGGATTTGTTGTTTGTTG |
| AA5 | LG3 | TGCCAATCCTGAGGTATTAACACC | CATTTTTGCAGTTGCAATTTCGT |
| AA122 | LG4 | GGGTCTGCATAAGTAGAAGCCA | AAGGTGTTTCCCCTAGACATCA |
| AD61 | LG4 | CTCATTCAATGATGATAATCCTA | ATGAGGTACTTGTGTGAGATAAA |
| A9 | LG4 | GTGCAGAAGCATTTGTTCAGAT | CCCACATATATTTGGTTGGTCA |
| AA315 | LG4 | AGTGGGAAGTAAAAGGTGTAG | TTTCACTAGATGATATTTCGTT |
| AA92 | LG4 | AAGGTCTGAAGCTGAACCTGAAGG | GCAGCCCACAGAAGTGCTTCAA |
| AB45 | LG4 | ATTACACCAACAATCTCCCACT | TGTAGAAGCATTTGGGTAGTTG |
| AB23 | LG5 | TCAGCCTTTATCCTCCGAACTA | GAACCCTTGTGCAGAAGCATTA |
| AC58 | LG5 | TCCGCAATTTGGTAACACTG | CGTCCATTTCTTTTATGCTGAG |
| AA399 | LG5 | CCATTGGTATATGAAAGATCGCT | TCCCAATTAATATGGCTAGGCT |
| PSGAPA-1 | LG5 | GACATTGTTGCCAATAACTGG | GGTTCTGTTCTCAATACAAG |
| AA163.2 | LG5 | TAGTTTCCAATTCAATCGACCA | AGTGTATTGTAAATGCACAAGGG |
| AD55.2 | LG5 | AACACATTAACTAAGTCCACAC | AAACCTATCACTTTAGAAACCT |
| AA99 | LG5 | AACAATAACATGGCAAAGATT | ACCTTGCGATATAATTGATG |
| AD51 | LG6 | ATGAAGTAGGCATAGCGAAGAT | GATTAAATAAAGTTCGATGGCG |
| AB20 | LG6 | TTGCATCCCACACAAGTGGT | ACCTCCAGGTTCTGCCTTATCT |
| AC76a | LG6 | CCCAATCCAATAAATAAAGAAA | AATGGTTGTTATGCCATTTT |
| AD159 | LG6 | AGCTTGGAACCACAAGATTAGT | GTGAATGATAATTCTCACCCTC |
| AA285 | LG6 | TCGCCTAATCTAGATGAGAATA | CTTAACATTTTAGGTCTTGGAG |
| AC74 | LG6 | CCTTAGTGTTCTTCAACTC | ACAGAACCAAGTTATCAATA |
| AD146 | LG7 | TGCTCAAGTCAATATATGAAGA | CAAGCAAATAGTTGTTTTGTTA |
| AA19 | LG7 | GCAGTTGTTGTACCCTAAAATT | TGTATTAGATGAAATTTTGTTTCTC |
| AA339 | LG7 | GTGTAGAAGTATTTTACTTGATG | CATCTATTGAAGGAAAATTAT |
| AD237 | LG7 | AGATCATTTGGTGTCATCAGTG | TGTTTAATACAACGTGCTCCTC |
| AA206 | LG7 | CTGAGAACTCAACGCTCAGACG | CGAGGGTCGAGTTCTGAGATTT |
| AA90 | LG7 | CCCTTACCATATTTCGTTTCT | TGCGACTCCATTCTAGTATTG |
| Traits | ACP 20 | AWP 600 | F4 | |||
|---|---|---|---|---|---|---|
| Min-Max | Min-Max | Min-Max | ||||
| Days to flowering | 50–56 | 53.10.7 | 115–119 | 116.70.7 | 24–103 | 54.31.8 |
| Days to pod setting | 59–63 | 61.70.4 | 127–130 | 128.30.6 | 30–114 | 63.11.9 |
| Plant height (cm) | 62–117 | 89.88.1 | 39–43 | 410.7 | 15–266.7 | 98.86.1 |
| First pod height (cm) | 17–40 | 27.83.9 | 5–6 | 5.30.2 | 2–75.4 | 14.91.4 |
| No of Internodes | 8–9 | 8.80.2 | 2–3 | 2.50.2 | 1.6–11.7 | 5.50.2 |
| No of pods per plant | 4–21 | 11.32.5 | 21–24 | 22.80.5 | 2–197 | 44.44.8 |
| No of seeds per pod | 5–8 | 6.70.4 | 2–3 | 2.70.2 | 1–7 | 3.10.1 |
| Pod length (cm) | 9–11 | 10.30.3 | 4–4.5 | 4.20.1 | 2.8–9 | 50.1 |
| Biological yield (g/plant) | 16.4–265.6 | 96.438.7 | 26.7–28.7 | 27.50.3 | 2–371.1 | 91.58.6 |
| Seed yield (g/plant) | 9.6–102.5 | 40.914.8 | 3.7–4.3 | 40.1 | 0.4–94.9 | 20.72.2 |
| Harvest index | 35.2–58.3 | 473.7 | 13.9–15.6 | 14.60.3 | 0.6–58.4 | 27.31.4 |
| LGs | Map Length (cM) | No. of Markers | Resolution of Markers (cM) |
|---|---|---|---|
| LG1 | 27.6 cM | 6 | 4.6 cM |
| LG2 | 46.7 cM | 8 | 5.8 cM |
| LG3 | 25.3 cM | 3 | 8.4 cM |
| LG4 | 38.4 cM | 6 | 6.4 cM |
| LG5 | 39.6 cM | 7 | 5.6 cM |
| LG6 | 44.7 cM | 7 | 6.3 cM |
| LG7 | 40.3 cM | 7 | 5.7 cM |
| Total | 262.6 cM | 44 | 5.9 cM |
| Characters | QTL | Linkage Group (LG) | Map Position (cM) | R2 * | Flanking Markers | Genetic Effect | Gene Action *** | |
|---|---|---|---|---|---|---|---|---|
| Add ** | Dom | |d|/|a| | ||||||
| Days to flowering | FLO2.1 | LG2 | 0–2 cM | 0.13 | AA205 | 6.5 | −6.9 | 1.06 D |
| FLO2.2 | LG2 | 30–32 cM | 0.13 | AA372.1 | 6.5 | −4.6 | 0.71 PD | |
| Plant height | PH3.1 | LG3 | 0–12 cM | 0.16 | AD73; AD270 | −28.6 | 47.2 | 1.65 OD |
| Pods per plant | NP2.1 | LG2 | 0–2 cM | 0.16 | AA205 | 10.4 | −17.9 | 1.72 OD |
| NP2.2 | LG2 | 26–30 cM | 0.13 | AD148 | 10.6 | −24.3 | 2.29 OD | |
| NP2.3 | LG2 | 46–46 cM | 0.13 | AA153 | 11.0 | −3.3 | 0.30 PD | |
| NP4.1 | LG4 | 38–38 cM | 0.14 | AB45 | 12.7 | 11.2 | 0.88 D | |
| NP5.1 | LG5 | 38–38 cM | 0.14 | AA99 | 10.0 | −23.1 | 2.31 OD | |
| NP6.1 | LG6 | 2–12 cM | 0.17 | AB20; AD51-2 | 9.2 | 463.5 | 50.38 OD | |
| Seeds per pod | NS6.1 | LG6 | 16–16 cM | 0.13 | AC76a | 1.8 | 19.8 | 11.0 OD |
| Pod length | PL4.1 | LG4 | 36–36 cM | 0.12 | AA92 | 0.42 | 0.47 | 1.1 D |
| Harvest index | HX3.1 | LG3 | 0–8 cM | 0.13 | AD73 | 8.0 | −3.2 | 0.39 PD |
| HX5.1 | LG5 | 38–38 cM | 0.12 | AA99 | 3.6 | −15.8 | 4.38 OD | |
| HX6.1 | LG6 | 8–8 cM | 0.12 | AD51-2 | 4.0 | −300.2 | 75.0 OD | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sari, H.; Eker, T.; Tosun, H.S.; Mutlu, N.; Celik, I.; Toker, C. Mapping QTLs for Super-Earliness and Agro-Morphological Traits in RILs Population Derived from Interspecific Crosses between Pisum sativum × P. fulvum. Curr. Issues Mol. Biol. 2023, 45, 663-676. https://doi.org/10.3390/cimb45010044
Sari H, Eker T, Tosun HS, Mutlu N, Celik I, Toker C. Mapping QTLs for Super-Earliness and Agro-Morphological Traits in RILs Population Derived from Interspecific Crosses between Pisum sativum × P. fulvum. Current Issues in Molecular Biology. 2023; 45(1):663-676. https://doi.org/10.3390/cimb45010044
Chicago/Turabian StyleSari, Hatice, Tuba Eker, Hilal Sule Tosun, Nedim Mutlu, Ibrahim Celik, and Cengiz Toker. 2023. "Mapping QTLs for Super-Earliness and Agro-Morphological Traits in RILs Population Derived from Interspecific Crosses between Pisum sativum × P. fulvum" Current Issues in Molecular Biology 45, no. 1: 663-676. https://doi.org/10.3390/cimb45010044
APA StyleSari, H., Eker, T., Tosun, H. S., Mutlu, N., Celik, I., & Toker, C. (2023). Mapping QTLs for Super-Earliness and Agro-Morphological Traits in RILs Population Derived from Interspecific Crosses between Pisum sativum × P. fulvum. Current Issues in Molecular Biology, 45(1), 663-676. https://doi.org/10.3390/cimb45010044