
The Negative Impact of Triptolide on the Immune Function of Human Natural Killer Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
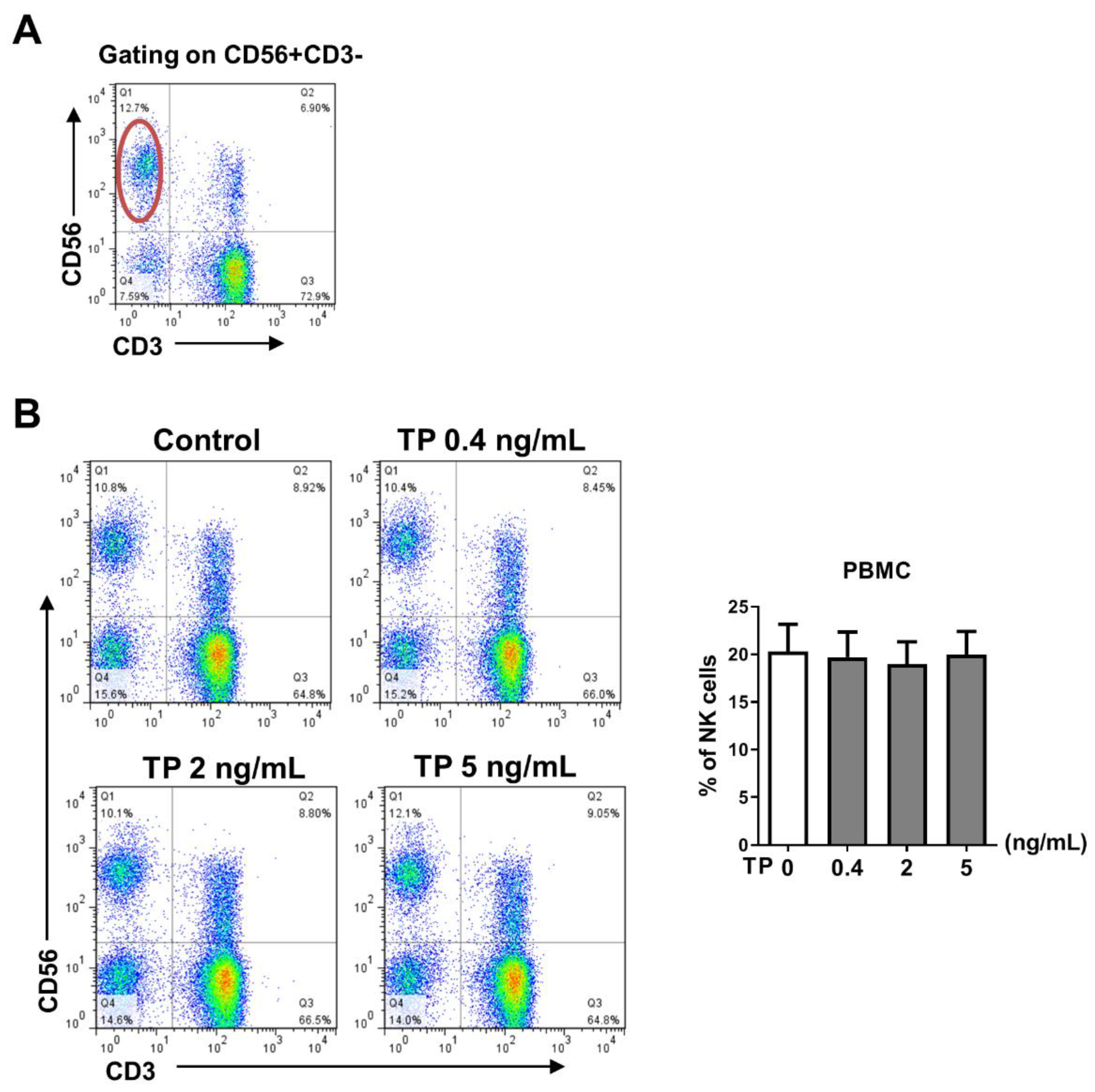
2.1. TP Did Not Influence the Proportion of Peripheral Blood NK Cells in Normal Human PBMCs
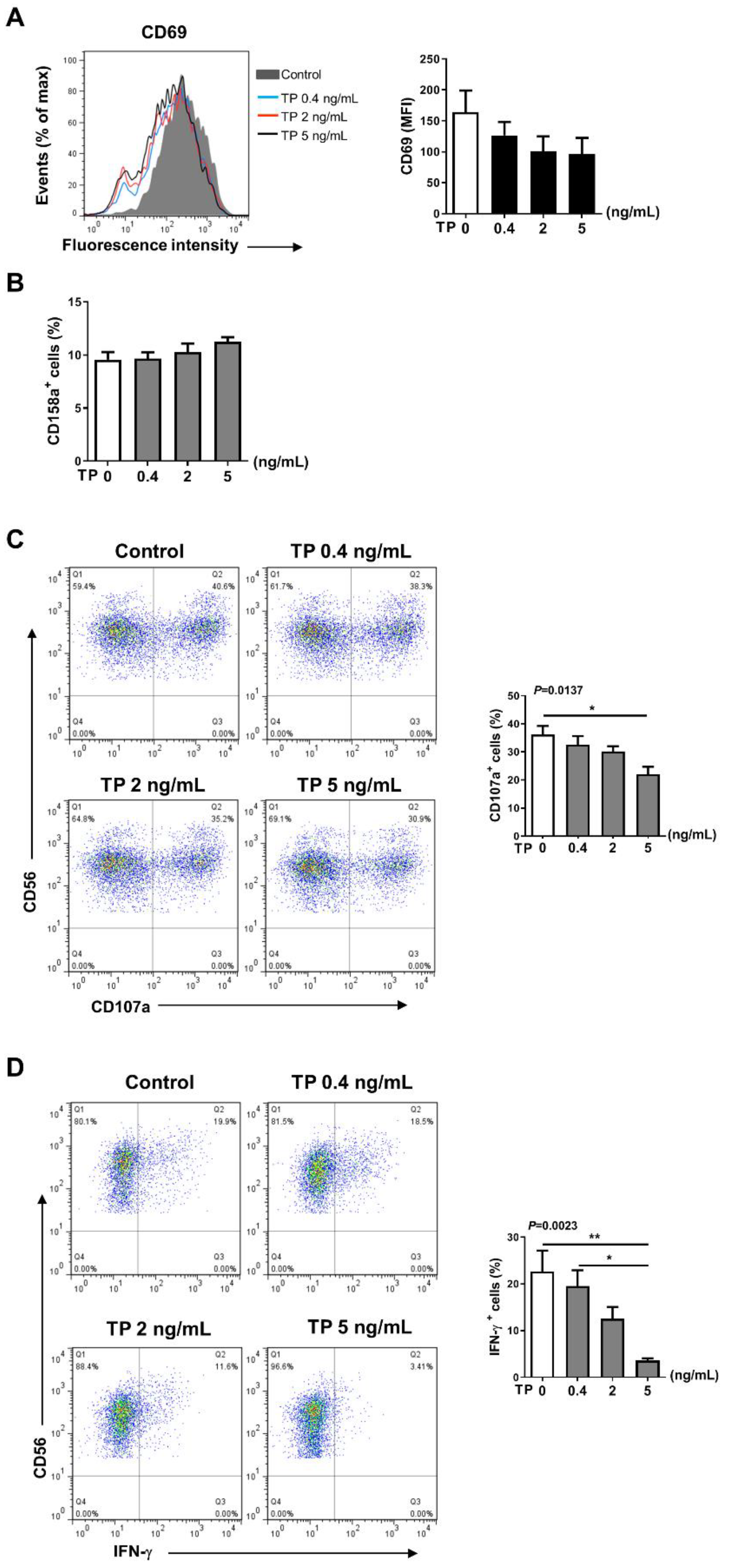
2.2. TP Inhibited the Activity and Function of NK Cells in Normal Human PBMCs
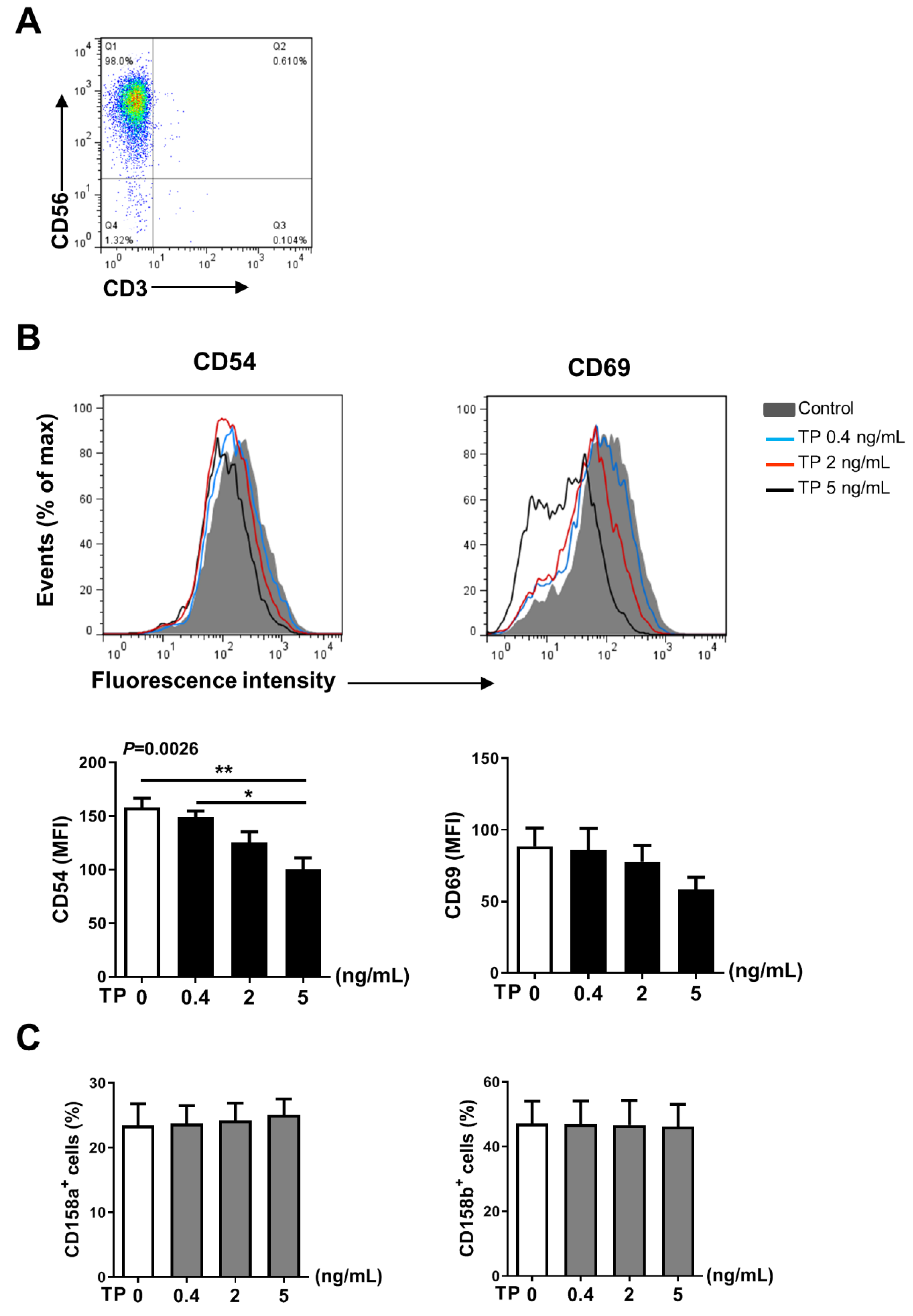
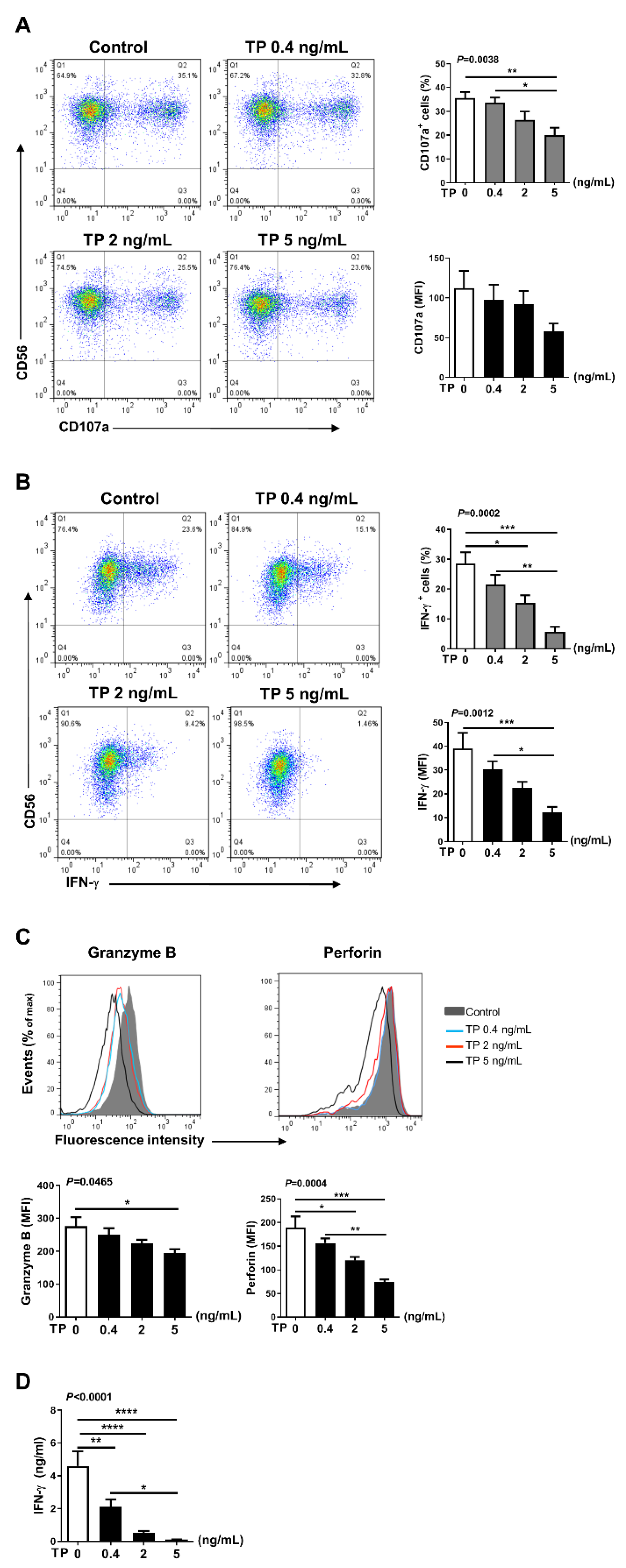
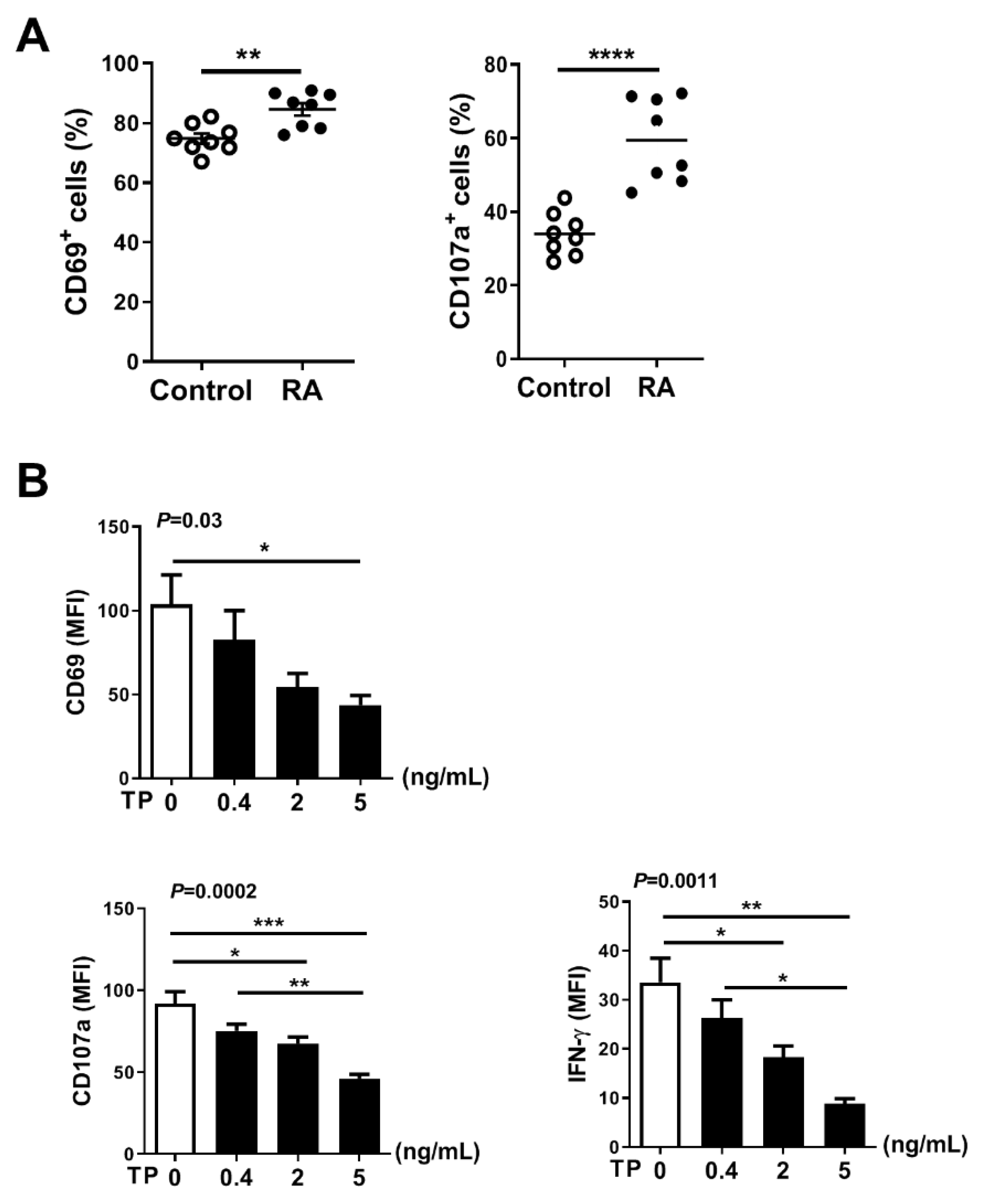
2.3. TP Had Negative Effects on Human Purified NK Cell Activity and Function
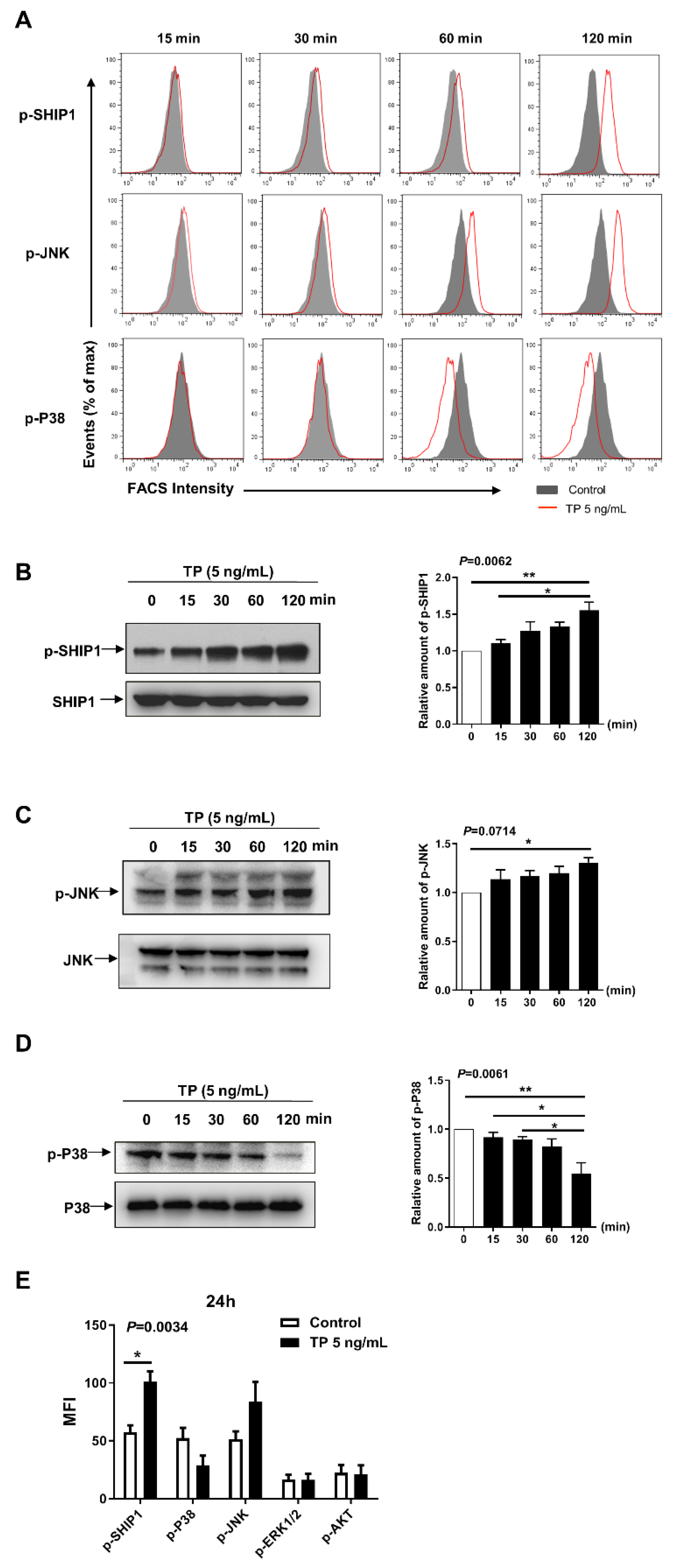
2.4. Effects of TP on Intracellular Signalling in NK Cells
2.5. TP Impaired NK Cell Function of Rheumatoid Arthritis (RA) Patients without Treatment
3. Discussion
4. Materials and Methods
4.1. Preparation of Human PBMCs and NK Cells
4.2. Cell Culture
4.3. Human NK Cells Phenotypic and Functional Analyses
4.4. Interferon-γ (IFN-γ) Synthesis in NK Cells
4.5. Western Blot
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tong, L.; Zhao, Q.; Datan, E.; Lin, G.; Minn, I.; Pomper, M.G.; Yu, B.; Romo, D.; He, Q.; Liu, J.O. Triptolide: Reflections on two decades of research and prospects for the future. Nat. Prod. Rep. 2021, 38, 843–860. [Google Scholar] [CrossRef]
- Wang, J.; Chu, Y.; Zhou, X. Inhibitory effect of Triperygium wilfordii polyglucoside on dipeptidyl peptidase I in vivo and in vitro. Biomed. Pharmacother. 2017, 96, 466–470. [Google Scholar] [CrossRef]
- Fan, D.; Guo, Q.; Shen, J.; Zheng, K.; Lu, C.; Zhang, G.; Lu, A.; He, X. The Effect of Triptolide in Rheumatoid Arthritis: From Basic Research towards Clinical Translation. Int. J. Mol. Sci. 2018, 19, 376. [Google Scholar] [CrossRef]
- Chen, X.; Murakami, T.; Oppenheim, J.J.; Howard, O.M. Triptolide, a constituent of immunosuppressive Chinese herbal medicine, is a potent suppressor of dendritic-cell maturation and trafficking. Blood 2005, 106, 2409–2416. [Google Scholar] [CrossRef] [PubMed]
- Rao, Q.; Ma, G.; Li, M.; Wu, H.; Zhang, Y.; Zhang, C.; Ma, Z.; Huang, L. Targeted delivery of triptolide by dendritic cell-derived exosomes for colitis and rheumatoid arthritis therapy in murine models. Br. J. Pharmacol. 2023, 180, 330–346. [Google Scholar] [CrossRef] [PubMed]
- Rao, Q.; Ma, G.C.; Wu, H.; Li, M.; Xu, W.; Wang, G.J.; Wang, D.; Zhang, C.E.; Ma, Z.J.; Zhang, Z.T. Dendritic cell combination therapy reduces the toxicity of triptolide and ameliorates colitis in murine models. Drug Deliv. 2022, 29, 679–691. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Zhou, W.; Li, K.; Sacks, S.H. Triptolide is a potent suppressant of C3, CD40 and B7h expression in activated human proximal tubular epithelial cells. Kidney Int. 2002, 62, 1291–1300. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhang, Y.; Liu, X.; Wu, X.; Huang, L.; Gao, W. Triptolide: Pharmacological spectrum, biosynthesis, chemical synthesis and derivatives. Theranostics 2021, 11, 7199–7221. [Google Scholar] [CrossRef]
- Huang, G.; Yuan, K.; Zhu, Q.; Zhang, S.; Lu, Q.; Zhu, M.; Sheng, H.; Yu, R.; Luo, G.; Xu, A. Triptolide inhibits the inflammatory activities of neutrophils to ameliorate chronic arthritis. Mol. Immunol. 2018, 101, 210–220. [Google Scholar] [CrossRef]
- Morandi, F.; Yazdanifar, M.; Cocco, C.; Bertaina, A.; Airoldi, I. Engineering the Bridge between Innate and Adaptive Immunity for Cancer Immunotherapy: Focus on gammadelta T and NK Cells. Cells 2020, 9, 1757. [Google Scholar] [CrossRef]
- Ferretti, E.; Carlomagno, S.; Pesce, S.; Muccio, L.; Obino, V.; Greppi, M.; Solari, A.; Setti, C.; Marcenaro, E.; Della, C.M.; et al. Role of the Main Non HLA-Specific Activating NK Receptors in Pancreatic, Colorectal and Gastric Tumors Surveillance. Cancers 2020, 12, 3705. [Google Scholar] [CrossRef]
- Vitale, M.; Cantoni, C.; Della Chiesa, M.; Ferlazzo, G.; Carlomagno, S.; Pende, D.; Falco, M.; Pessino, A.; Muccio, L.; De Maria, A.; et al. An Historical Overview: The Discovery of How NK Cells Can Kill Enemies, Recruit Defense Troops, and More. Front. Immunol. 2019, 10, 1415. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.J.; Hsu, C.Y.; Kuo, M.L.; Lee, P.T.; Hsiao, H.S.; Chen, J.Y. Phenotypic and functional characterization of natural killer cells in rheumatoid arthritis-regulation with interleukin-15. Sci. Rep. 2020, 10, 5858. [Google Scholar] [CrossRef] [PubMed]
- Kucuksezer, U.C.; Aktas, C.E.; Esen, F.; Tahrali, I.; Akdeniz, N.; Gelmez, M.Y.; Deniz, G. The Role of Natural Killer Cells in Autoimmune Diseases. Front. Immunol. 2021, 12, 622306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.L.; Colmenero, P.; Purath, U.; Teixeira, D.M.C.; Hueber, W.; Klareskog, L.; Tarner, I.H.; Engleman, E.G.; Soderstrom, K. Natural killer cells trigger differentiation of monocytes into dendritic cells. Blood 2007, 110, 2484–2493. [Google Scholar] [CrossRef] [PubMed]
- Yamin, R.; Berhani, O.; Peleg, H.; Aamar, S.; Stein, N.; Gamliel, M.; Hindi, I.; Scheiman-Elazary, A.; Gur, C. High percentages and activity of synovial fluid NK cells present in patients with advanced stage active Rheumatoid Arthritis. Sci. Rep. 2019, 9, 1351. [Google Scholar] [CrossRef]
- Louis, C.; Souza-Fonseca-Guimaraes, F.; Yang, Y.; D Silva, D.; Kratina, T.; Dagley, L.; Hediyeh-Zadeh, S.; Rautela, J.; Masters, S.L.; Davis, M.J.; et al. NK cell-derived GM-CSF potentiates inflammatory arthritis and is negatively regulated by CIS. J. Exp. Med. 2020, 217, e20191421. [Google Scholar] [CrossRef]
- Yang, Y.; Day, J.; Souza Fonseca Guimaraes, F.; Wicks, I.P.; Louis, C. Natural killer cells in inflammatory autoimmune diseases. Clin. Transl. Immunol. 2021, 10, e1250. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhao, Y.; Zheng, Y. Therapeutic potential of triptolide in autoimmune diseases and strategies to reduce its toxicity. Chin. Med. 2021, 16, 114. [Google Scholar] [CrossRef]
- Liu, C.; Min, X.; Wang, N.; Wang, J.; Ma, N.; Dong, X.; Zhang, B.; Wu, W.; Li, Z.; Zhou, W.; et al. Complement Receptor 3 Has Negative Impact on Tumor Surveillance through Suppression of Natural Killer Cell Function. Front. Immunol. 2017, 8, 1602. [Google Scholar] [CrossRef]
- Liu, G.; Hu, Y.; Jin, S.; Jiang, Q. Genetic variant rs763361 regulates multiple sclerosis CD226 gene expression. Proc. Natl. Acad. Sci. USA 2017, 114, E906–E907. [Google Scholar] [CrossRef]
- Zitti, B.; Bryceson, Y.T. Natural killer cells in inflammation and autoimmunity. Cytokine Growth Factor Rev. 2018, 42, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Dean, J.W.; Peters, L.D.; Fuhrman, C.A.; Seay, H.R.; Posgai, A.L.; Stimpson, S.E.; Brusko, M.A.; Perry, D.J.; Yeh, W.; Newby, B.N.; et al. Innate inflammation drives NK cell activation to impair Treg activity. J. Autoimmun. 2020, 108, 102417. [Google Scholar] [CrossRef]
- Zhang, C.; Tian, Z. NK cell subsets in autoimmune diseases. J. Autoimmun. 2017, 83, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Ogawa, E.; Okuyama, R. Role of Innate Immune Cells in Psoriasis. Int. J. Mol. Sci. 2020, 21, 6604. [Google Scholar] [CrossRef]
- Pauls, S.D.; Marshall, A.J. Regulation of immune cell signaling by SHIP1: A phosphatase, scaffold protein, and potential therapeutic target. Eur. J. Immunol. 2017, 47, 932–945. [Google Scholar] [CrossRef] [PubMed]
- Min, X.; Liu, C.; Wei, Y.; Wang, N.; Yuan, G.; Liu, D.; Li, Z.; Zhou, W.; Li, K. Expression and regulation of complement receptors by human natural killer cells. Immunobiology 2014, 219, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Min, X.; Liu, C.; Cao, B.; Zhang, T.; Yang, X.; Ma, N.; Wang, N.; Li, K. Human CD3+CD56+NKT-like cells express a range of complement receptors and C3 activation has negative effects on these cell activity and effector function. Hum. Immunol. 2021, 82, 625–633. [Google Scholar] [CrossRef]
- Wang, N.; Wu, W.; Qiang, C.; Ma, N.; Wu, K.; Liu, D.; Wang, J.X.; Yang, X.; Xue, L.; Diao, T.Y.; et al. Protective Role of Collectin 11 in a Mouse Model of Rheumatoid Arthritis. Arthritis Rheumatol. 2021, 73, 1430–1440. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, N.; Min, X.; Ma, N.; Zhu, Z.; Cao, B.; Wang, Y.; Yong, Q.; Huang, J.; Li, K. The Negative Impact of Triptolide on the Immune Function of Human Natural Killer Cells. Pharmaceuticals 2023, 16, 458. https://doi.org/10.3390/ph16030458
Wang N, Min X, Ma N, Zhu Z, Cao B, Wang Y, Yong Q, Huang J, Li K. The Negative Impact of Triptolide on the Immune Function of Human Natural Killer Cells. Pharmaceuticals. 2023; 16(3):458. https://doi.org/10.3390/ph16030458
Chicago/Turabian StyleWang, Na, Xiaoyun Min, Ning Ma, Zhuoran Zhu, Bo Cao, Yuan Wang, Qing Yong, Jingjin Huang, and Ke Li. 2023. "The Negative Impact of Triptolide on the Immune Function of Human Natural Killer Cells" Pharmaceuticals 16, no. 3: 458. https://doi.org/10.3390/ph16030458
APA StyleWang, N., Min, X., Ma, N., Zhu, Z., Cao, B., Wang, Y., Yong, Q., Huang, J., & Li, K. (2023). The Negative Impact of Triptolide on the Immune Function of Human Natural Killer Cells. Pharmaceuticals, 16(3), 458. https://doi.org/10.3390/ph16030458