

The Influence of Protein Charge and Molecular Weight on the Affinity of Aptamers


Abstract
1. Introduction
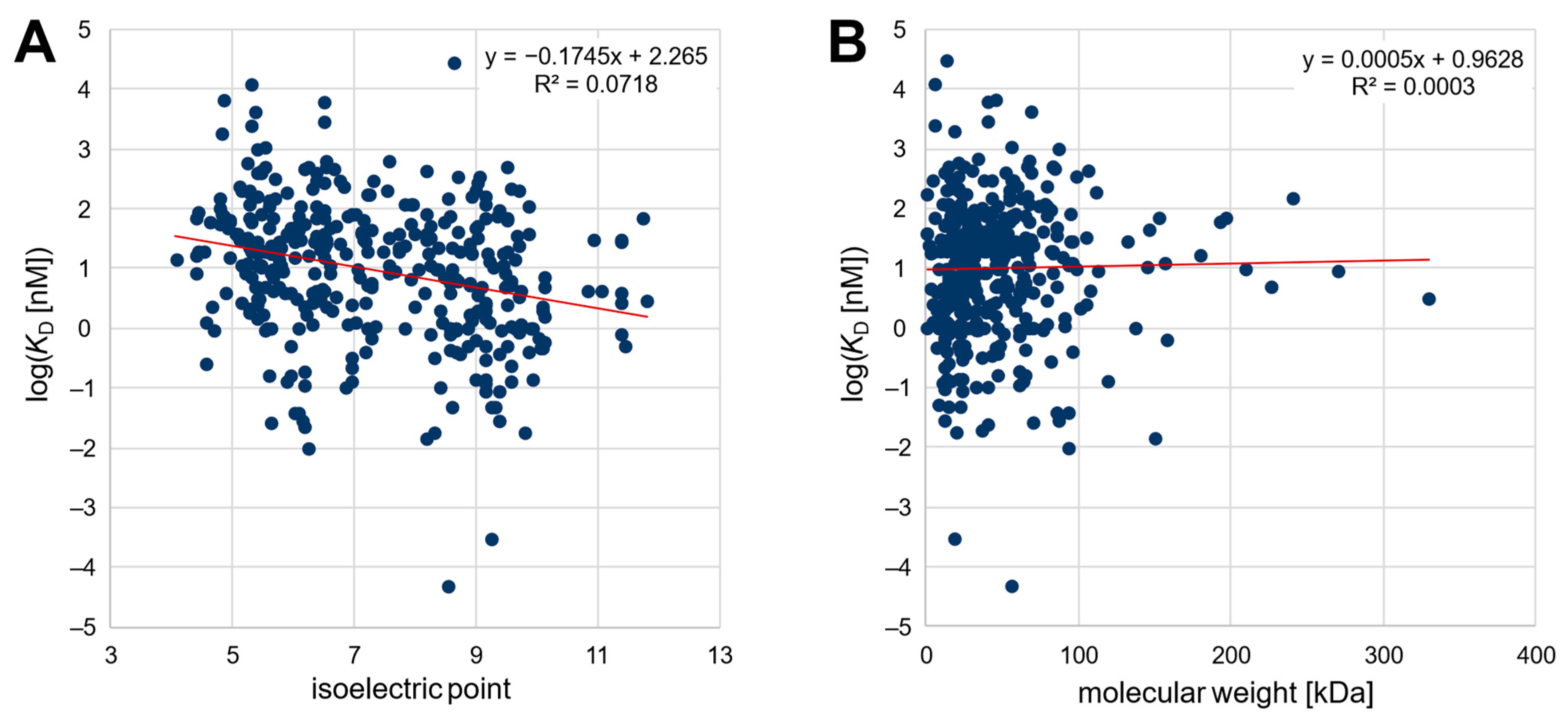
2. Results and Discussion
3. Materials and Methods
3.1. Electrophoretic Mobility Shift Assay
3.2. Determination of Dissociation Constants via Fluorescence Assay after Bead-Based Affinity Chromatography
3.3. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kluβmann, S. The Aptamer Handbook: Functional Oligonucleotides and Their Applications; John Wiley & Sons: Denver, CO, USA, 2006. [Google Scholar]
- McKeague, M.; Giamberardino, A.; DeRosa, M.C. Advances in aptamer-based biosensors for food safety. In Environmental Biosensors; IntechOpen: London, UK, 2011. [Google Scholar]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Robertson, D.L.; Joyce, G.F. Selection in vitro of an RNA enzyme that specifically cleaves single-stranded DNA. Nature 1990, 344, 467–468. [Google Scholar] [CrossRef] [PubMed]
- Drees, A.; Fischer, M. High-Throughput Selection and Characterisation of Aptamers on Optical Next-Generation Sequencers. Int. J. Mol. Sci. 2021, 22, 9202. [Google Scholar] [CrossRef] [PubMed]
- Thiel, W.H. Galaxy workflows for web-based bioinformatics analysis of aptamer high-throughput sequencing data. Mol. Ther.-Nucleic Acids 2016, 5, e345. [Google Scholar] [CrossRef] [PubMed]
- McKeague, M.; McConnell, E.M.; Cruz-Toledo, J.; Bernard, E.D.; Pach, A.; Mastronardi, E.; Zhang, X.; Beking, M.; Francis, T.; Giamberardino, A. Analysis of in vitro aptamer selection parameters. J. Mol. Evol. 2015, 81, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Hirose, K.; Tsuchida, M.; Asakura, H.; Wakui, K.; Yoshimoto, K.; Iida, K.; Sato, M.; Shibukawa, M.; Suganuma, M.; Saito, S. A single-round selection of selective DNA aptamers for mammalian cells by polymer-enhanced capillary transient isotachophoresis. Analyst 2017, 142, 4030–4038. [Google Scholar] [CrossRef]
- Wu, Z.; Shen, H.; Hu, J.; Fu, Q.; Yao, C.; Yu, S.; Xiao, W.; Tang, Y. Aptamer-based fluorescence-quenching lateral flow strip for rapid detection of mercury (II) ion in water samples. Anal. Bioanal. Chem. 2017, 409, 5209–5216. [Google Scholar] [CrossRef]
- Cruz-Toledo, J.; McKeague, M.; Zhang, X.; Giamberardino, A.; McConnell, E.; Francis, T.; DeRosa, M.C.; Dumontier, M. Aptamer base: A collaborative knowledge base to describe aptamers and SELEX experiments. Database 2012, 2012, bas006. [Google Scholar] [CrossRef]
- Hermann, T.; Patel, D.J. Adaptive recognition by nucleic acid aptamers. Science 2000, 287, 820–825. [Google Scholar] [CrossRef]
- Ahmad, K.M.; Oh, S.S.; Kim, S.; McClellen, F.M.; Xiao, Y.; Soh, H.T. Probing the limits of aptamer affinity with a microfluidic SELEX platform. PLoS ONE 2011, 6, e27051. [Google Scholar] [CrossRef] [PubMed]
- Welfle, K.; Misselwitz, R.; Hausdorf, G.; Höhne, W.; Welfle, H. Conformation, pH-induced conformational changes, and thermal unfolding of anti-p24 (HIV-1) monoclonal antibody CB4-1 and its Fab and Fc fragments. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 1999, 1431, 120–131. [Google Scholar] [CrossRef]
- Voordouw, G.; Milo, C.; Roche, R.S. Role of bound calcium ions in thermostable, proteolytic enzymes. Separation of intrinsic and calcium ion contributions to the kinetic thermal stability. Biochemistry 1976, 15, 3716–3724. [Google Scholar]
- Hianik, T.; Ostatná, V.; Sonlajtnerova, M.; Grman, I. Influence of ionic strength, pH and aptamer configuration for binding affinity to thrombin. Bioelectrochemistry 2007, 70, 127–133. [Google Scholar] [CrossRef]
- Neves, M.A.; Reinstein, O.; Saad, M.; Johnson, P.E. Defining the secondary structural requirements of a cocaine-binding aptamer by a thermodynamic and mutation study. Biophys. Chem. 2010, 153, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; German, I.; Buchanan, D.; Kennedy, R.T. Retention and separation of adenosine and analogues by affinity chromatography with an aptamer stationary phase. Anal. Chem. 2001, 73, 5415–5421. [Google Scholar] [CrossRef]
- Perelson, A.S. Mathematical approaches in immunology. In Theory and Control of Dynamical Systems; World Scientific Publishing Company: Stockholm, Sweden, 1992; pp. 200–230. [Google Scholar]
- Perelson, A.S.; Oster, G.F. Theoretical studies of clonal selection: Minimal antibody repertoire size and reliability of self-non-self discrimination. J. Theor. Biol. 1979, 81, 645–670. [Google Scholar] [CrossRef]
- Carothers, J.M.; Goler, J.A.; Kapoor, Y.; Lara, L.; Keasling, J.D. Selecting RNA aptamers for synthetic biology: Investigating magnesium dependence and predicting binding affinity. Nucleic Acids Res. 2010, 38, 2736–2747. [Google Scholar] [CrossRef]
- Hünniger, T.; Wessels, H.; Fischer, C.; Paschke-Kratzin, A.; Fischer, M. Just in time-selection: A rapid semiautomated SELEX of DNA aptamers using magnetic separation and BEAMing. Anal. Chem. 2014, 86, 10940–10947. [Google Scholar]
- Tran, D.T.; Janssen, K.P.; Pollet, J.; Lammertyn, E.; Anné, J.; Van Schepdael, A.; Lammertyn, J. Selection and characterization of DNA aptamers for egg white lysozyme. Molecules 2010, 15, 1127–1140. [Google Scholar] [CrossRef]
- Jarmoskaite, I.; AlSadhan, I.; Vaidyanathan, P.P.; Herschlag, D. How to measure and evaluate binding affinities. Elife 2020, 9, e57264. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: The Universal Protein knowledgebase in 2023. Nucleic Acids Res. 2023, 51, D523–D531. [CrossRef] [PubMed]
- Duvaud, S.; Gabella, C.; Lisacek, F.; Stockinger, H.; Ioannidis, V.; Durinx, C. Expasy, the Swiss Bioinformatics Resource Portal, as designed by its users. Nucleic Acids Res. 2021, 49, W216–W227. [Google Scholar] [CrossRef] [PubMed]
- Illarionov, B.; Illarionova, V.; Lee, J.; van Dongen, W.; Vervoort, J. Expression and properties of the recombinant lumazine (riboflavin) protein from Photobacterium Leiognathi. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1994, 1201, 251–258. [Google Scholar] [CrossRef]
- Frohnmeyer, E.; Frisch, F.; Falke, S.; Betzel, C.; Fischer, M. Highly affine and selective aptamers against cholera toxin as capture elements in magnetic bead-based sandwich ELAA. J. Biotechnol. 2018, 269, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.; Lei, L.; Egli, M. Label-Free Electrophoretic Mobility Shift Assay (EMSA) for Measuring Dissociation Constants of Protein-RNA Complexes. Curr. Protoc. Nucleic Acid Chem. 2019, 76, e70. [Google Scholar] [CrossRef]
- Fischer, C.; Klockmann, S.; Wessels, H.; Hünniger, T.; Schrader, J.; Paschke-Kratzin, A.; Fischer, M. Aptamer-based trapping of phytosphingosine in urine samples. J. Biotechnol. 2016, 238, 30–34. [Google Scholar] [CrossRef]
- Holleman, A.F.; Wieberg, E.; Wiberg, N. Lehrbuch der Anorganischen Chemie, 91th–100th ed.; Walter de Gruyter GmbH & Co KG: Berlin, Germany, 2019. [Google Scholar]
- Davies, D.R.; Gelinas, A.D.; Zhang, C.; Rohloff, J.C.; Carter, J.D.; O’Connell, D.; Waugh, S.M.; Wolk, S.K.; Mayfield, W.S.; Burgin, A.B. Unique motifs and hydrophobic interactions shape the binding of modified DNA ligands to protein targets. Proc. Natl. Acad. Sci. USA 2012, 109, 19971–19976. [Google Scholar] [CrossRef]


{kind=link}
| pI * | MW [kDa] * | Protein | Oligonucleotide | KD |
|---|---|---|---|---|
| 11.4 | 14.3 | Lysozyme C | 1 2 | 14 nM 8 nM |
| 9.59 | 53.7 | Cytochrome C | 1 2 | 1.4 µM 1.1 µM |
| 8.75 | 25.7 | Chymotrypsin | 1 2 | 1.4 µM 3.3 µM |
| 7.36 | 17.6 | Myoglobin | 1 2 | n.d. n.d. |
| 4.6–6.5 | 25.8 | Trypsin-1 | 1 2 | n.d. n.d. |
| 5.82 | 69.3 | BSA | 1 2 | n.d. n.d. |
| 4.1–5.8 | 19–25 | Casein | 1 2 | n.d. n.d. |
| 5.36 | 68.8 | PEA | 1 2 | n.d. n.d. |
| 5.35 | 42.7 | Ovalbumin | 1 2 | n.d. n.d. |
| 4.94 | 12.8 | Lectin A | 1 2 | n.d. n.d. |
| 4.44 | 20.0 | Lumazine Protein | 1 2 | n.d. n.d. |
| 3.87 | 36.0 | Pepsin | 1 2 | n.d. n.d. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drees, A.; Trinh, T.L.; Fischer, M. The Influence of Protein Charge and Molecular Weight on the Affinity of Aptamers. Pharmaceuticals 2023, 16, 457. https://doi.org/10.3390/ph16030457
Drees A, Trinh TL, Fischer M. The Influence of Protein Charge and Molecular Weight on the Affinity of Aptamers. Pharmaceuticals. 2023; 16(3):457. https://doi.org/10.3390/ph16030457
Chicago/Turabian StyleDrees, Alissa, Tung Lam Trinh, and Markus Fischer. 2023. "The Influence of Protein Charge and Molecular Weight on the Affinity of Aptamers" Pharmaceuticals 16, no. 3: 457. https://doi.org/10.3390/ph16030457
APA StyleDrees, A., Trinh, T. L., & Fischer, M. (2023). The Influence of Protein Charge and Molecular Weight on the Affinity of Aptamers. Pharmaceuticals, 16(3), 457. https://doi.org/10.3390/ph16030457