
A New Species of the Medically Important Scorpion Genus, Hemiscorpius Peters, 1861 (Hemiscorpiidae), from Southern Iran

 , ,
, , 
Abstract
1. Introduction
2. Material and Methods
2.1. Morphology
2.2. Molecular Systematics
3. Results
3.1. Molecular Systematics

3.2. Taxonomy

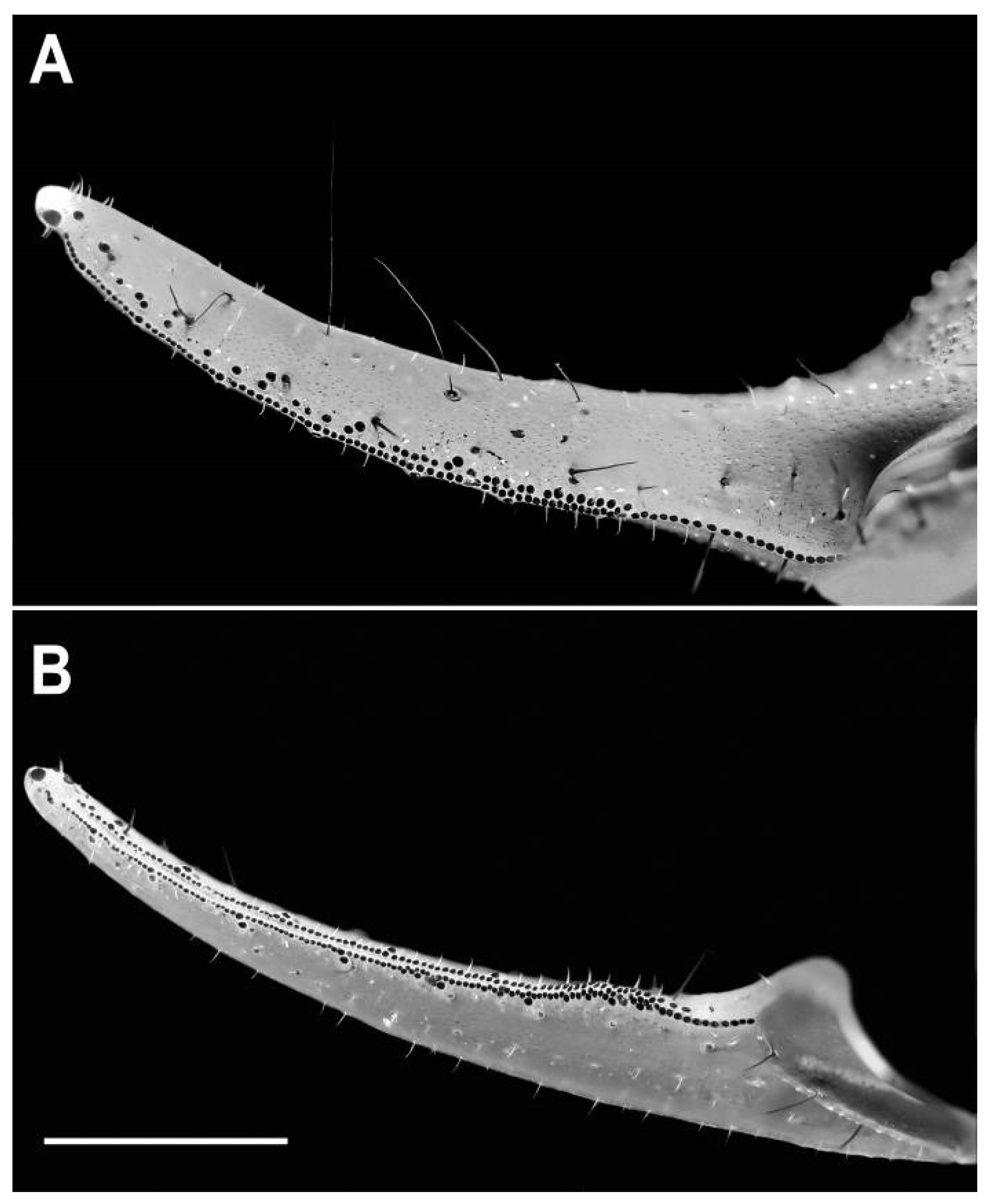
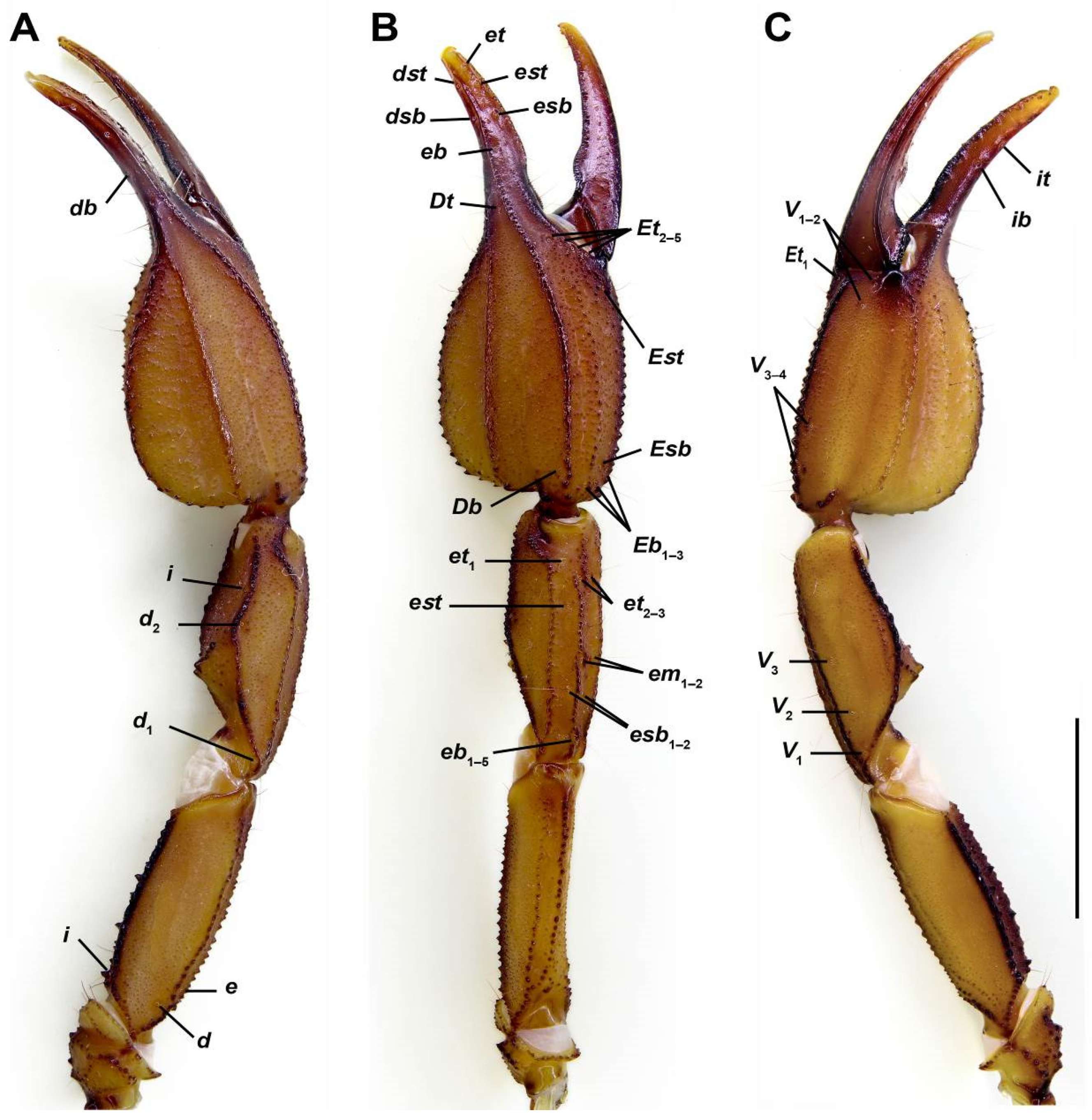


4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lourenço, W.R. The genus Hemiscorpius Peters, 1861 (Scorpiones: Hemiscorpiidae) in East Africa, and description of a new species from Somalia. Entomol. Mitt. zool. Mus. Hamburg. 2011, 15, 275–285. [Google Scholar]
- Prendini, L. Phylogeny and classification of the superfamily Scorpionoidea Latreille 1802 (Chelicerata, Scorpiones): An exemplar approach. Cladistics 2000, 16, 1–78. [Google Scholar] [CrossRef] [PubMed]
- Monod, L.; Lourenço, W.R. Hemiscorpiidae (Scorpiones) from Iran, with descriptions of two new species and notes on biogeography and phylogenetic relationships. Rev. suisse Zool. 2005, 112, 869–941. [Google Scholar] [CrossRef]
- Shahi, M.; Barahoei, H. Morphological study of Hemiscorpius Peters, 1861 (Scorpiones: Hemiscorpiidae) in Hormozgan Province, southern Iran. Arch. Razi Inst. 2023, 78, 1588–1600. [Google Scholar] [CrossRef]
- Barahoei, H.; Navidpour, S.; Aliabadian, M.; Siahsarvie, R.; Mirshamsi, O. Scorpions of Iran (Arachnida: Scorpiones): Annotated checklist, DELTA database and identification key. J. Insect Biodivers. Syst. 2020, 6, 375–474. [Google Scholar] [CrossRef]
- Shahi, M.; Rafinejad, J.; Az-Khosravi, L.; Moosavy, S. First report of death due to Hemiscorpius acanthocercus envenomation in Iran: Case report. Electron. Physician 2015, 7, 1234–1238. [Google Scholar]
- Akbari, H.; Mesbahzadeh, T.; Zehtabian, G. Prediction of climate change in arid and semi-arid regions of the western basin of Jazmourian wetland. Environ. Res. 2021, 11, 27–43. [Google Scholar]
- Salari, M.; Sampour, M. First two records of Hemiscorpius species (Scorpiones: Hemiscorpiidae) from Kerman Province, southeast of Iran. Asian J. Appl. Sci. 2017, 5, 101–108. [Google Scholar]
- Adeli-Sardou, M.; Shahi, M.; Dehghan, H.; Ahmadyousefi-Sarhadi, M.; Falah, G.; Barahoei, H. Geographical distribution of scorpions (Arachnida: Scorpiones) in southern regions of Kerman Province, Iran. Biol. Bull. 2024, 51, 644–654. [Google Scholar] [CrossRef]
- Dehghani, R.; Mazaheri Tehrani, A.; Ghadami, F.; Sanaei-Zadeh, H. Methods for collecting and capturing scorpions: A review. Int. J. Zool. Res. 2016, 4, 65–72. [Google Scholar]
- Vachon, M. Études des caractères utilisés pour classer les familles et les genres des scorpions (Arachnides). 1. La trichobothriotaxie en arachnologie. Sigles trichobothriaux et types de trichobothriotaxie chez les scorpions. Bull. Mus. Natl. Hist. Nat. 1974, 140, 857–958. [Google Scholar]
- Loria, S.F.; Prendini, L. Homology of the lateral eyes of Scorpiones: A six-ocellus model. PLoS ONE 2014, 9, e112913. [Google Scholar] [CrossRef]
- González-Santillán, E.; Prendini, L. Redefinition and generic revision of the north American vaejovid scorpion subfamily Syntropinae Kraepelin, 1905, with descriptions of six new genera. Bull. Am. Mus. Nat. Hist. 2013, 382, 1–71. [Google Scholar] [CrossRef]
- Stahnke, H.L. Scorpion nomenclature and mensuration. Entomol. News 1970, 81, 297–316. [Google Scholar]
- Sissom, W.D. Systematics, biogeography and paleontology. In The Biology of Scorpions; Polis, G.A., Ed.; Stanford University Press: Stanford, CA, USA, 1990; pp. 31–80. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial Cytochrome c Oxidase Subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Barahoei, H.; Prendini, L.; Navidpour, S.; Tahir, H.M.; Aliabadian, M.; Siahsarvie, R.; Mirshamsi, O. Integrative systematics of the tooth-tailed scorpions, Odontobuthus (Buthidae), with descriptions of three new species from the Iranian Plateau. Zool. J. Linn. Soc. 2022, 195, 355–398. [Google Scholar] [CrossRef]
- Shahi, M.; Barahoei, H. Molecular study of Hemiscorpius Peters (Scorpiones: Hemiscorpiidae) in Hormozgan Province, south of Iran. Arch. Razi Inst. 2024, 79, 216–222. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Vakili Shahrbabaki, S.M.A.; Alikhani, T. Introduction of the flora, life form and chorotypes of plants in Jeball Barez (Jiroft). J. Plant Environ. Physiol. 2013, 8, 66–79. [Google Scholar]
- Kovařík, F.; Navidpour, S. Six new species of Orthochirus Karsch, 1892 from Iran (Scorpiones: Buthidae). Euscorpius 2020, 132, 1–41. [Google Scholar]
- Kovařík, F.; Fet, V.; Gantenbein, B.; Graham, M.R.; Yağmur, E.A.; Šťáhlavský, F.; Poverennyi, N.M.; Novruzov, N.E. A revision of the genus Mesobuthus Vachon, 1950, with a description of 14 new species (Scorpiones: Buthidae). Euscorpius 2022, 348, 1–189. [Google Scholar]
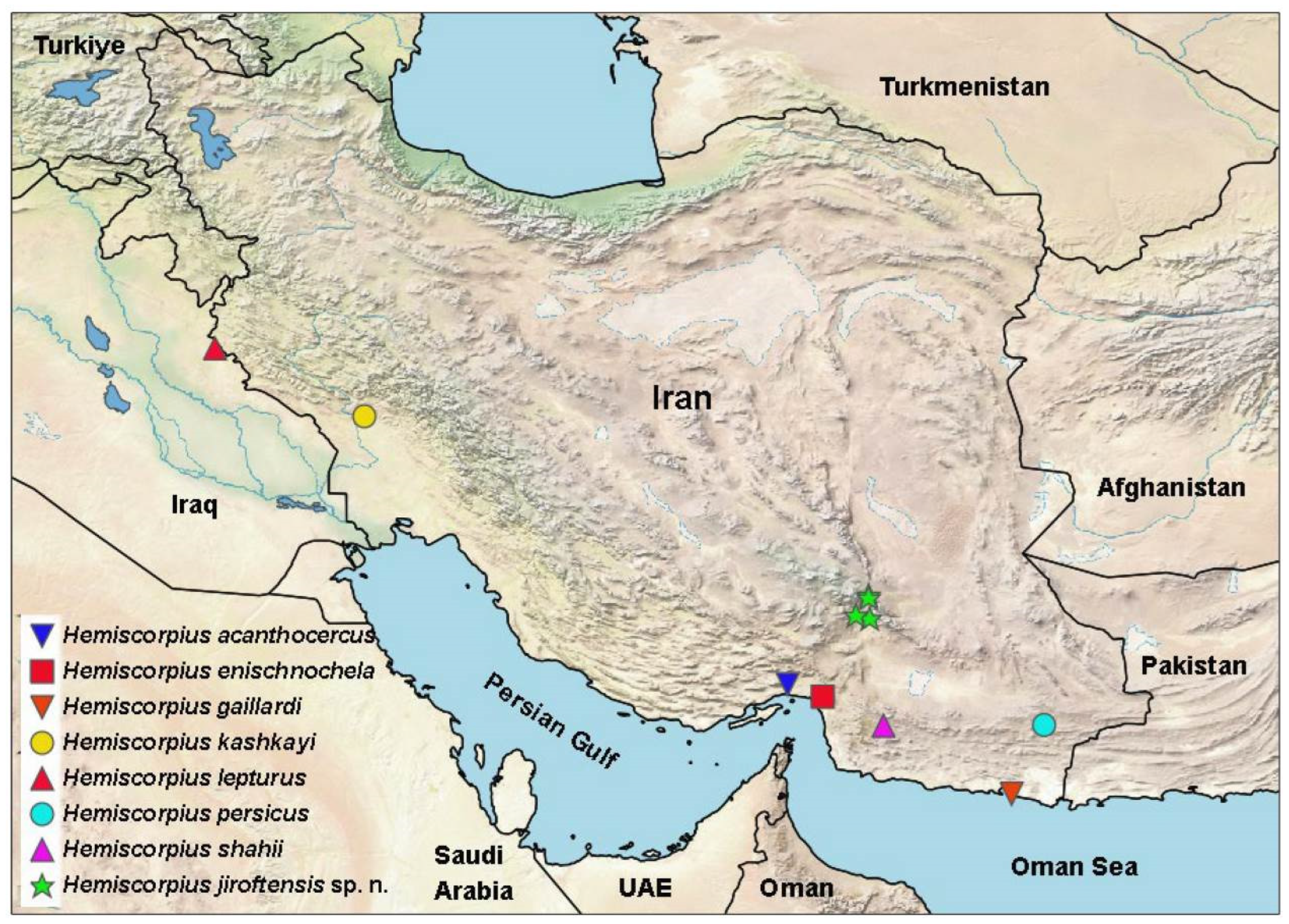

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Country | H. acanth. | H. enisch. | H. leptur. | H. leptur. | H. jirof. |
|---|---|---|---|---|---|---|
| H. acanthocercus | Iran | 0.029 | ||||
| H. enischnochela | Iran | 0.126 | 0.036 | |||
| H. lepturus | Iran | 0.147 | 0.136 | - | ||
| H. lepturus | Iraq | 0.139 | 0.129 | 0.089 | - | |
| H. jiroftensis | Iran | 0.130 | 0.105 | 0.138 | 0.115 | 0.013 |
| Characters | Specimen Code and Sex | |||||||
|---|---|---|---|---|---|---|---|---|
| JMU HD 23 | RIZ HD 44A | RIZ HD 44B | JMU HD 45A | JMU HD 45B | JMU HD 45C | JMU HD 43 | RIZ HD 44C | |
| ♂ | ♂ | ♂ | ♂ | ♂ | ♂ | ♀ | ♀ | |
| Total body length | 75.08 | 80.94 | 79.95 | 65.41 | 84.01 | 90.39 | 60.14 | 69.16 |
| Carapace length | 6.58 | 7.7 | 7.6 | 6.4 | 7.72 | 7.83 | 7.14 | 8.67 |
| Carapace anterior width | 4.41 | 5 | 4.9 | 4.42 | 4.59 | 4.97 | 4.47 | 5.39 |
| Carapace posterior width | 5.11 | 5.48 | 5.76 | 5.12 | 5.32 | 5.95 | 6.09 | 6.64 |
| Pedipalp femur length | 5.83 | 7.32 | 7.66 | 5.15 | 7.53 | 8.06 | 6.05 | 8 |
| Pedipalp femur width | 2.22 | 2.56 | 2.46 | 2.16 | 2.36 | 2.63 | 2.4 | 2.83 |
| Pedipalp patella length | 5.9 | 7.31 | 7.34 | 6.28 | 6.78 | 7.21 | 6.44 | 7.41 |
| Pedipalp patella width | 2.13 | 2.45 | 2.51 | 2.34 | 2.39 | 2.43 | 2.55 | 2.95 |
| Pedipalp chela manus length | 5.67 | 7.15 | 6.92 | 6.04 | 6.43 | 7.06 | 6.11 | 7.23 |
| Pedipalp chela manus width | 3.83 | 5.54 | 5 | 3.82 | 4.91 | 5.23 | 4.61 | 5.91 |
| Pedipalp chela manus depth | 2.39 | 2.54 | 2.8 | 2.16 | 2.22 | 2.43 | 2.95 | 3.2 |
| Pedipalp chela movable finger length | 5.82 | 7.74 | 7.34 | 6.49 | 7.22 | 7.5 | 6.47 | 8.11 |
| Pedipalp chela fixed/movable finger denticles | 7/7 | 7/7 | 7/7 | 7/7 | 7/7 | 7/7 | 6/6 | 7/7 |
| Mesosoma length | 20.98 | 22.06 | 21.69 | 15.67 | 23.27 | 22.23 | 21.72 | 22.29 |
| Metasomal segment I length | 7.23 | 7.91 | 7.79 | 6.35 | 8.16 | 8.96 | 4.23 | 5.61 |
| Metasomal segment I width | 1.77 | 2.05 | 2.13 | 1.91 | 1.78 | 2.02 | 2.4 | 2.43 |
| Metasomal segment I depth | 1.79 | 1.95 | 2 | 1.58 | 1.54 | 1.85 | 2.13 | 2.18 |
| Metasomal segment II length | 7.9 | 8.48 | 8.55 | 7.1 | 8.75 | 9.74 | 4.58 | 5.94 |
| Metasomal segment II width | 1.6 | 1.79 | 1.85 | 1.64 | 1.5 | 1.7 | 2.1 | 2.3 |
| Metasomal segment II depth | 1.77 | 1.85 | 1.9 | 1.59 | 1.55 | 1.85 | 2.09 | 2.32 |
| Metasomal segment III length | 8.14 | 8.95 | 8.72 | 7.38 | 9.04 | 10.71 | 5.03 | 6.09 |
| Metasomal segment III width | 1.47 | 1.67 | 1.74 | 1.5 | 1.45 | 1.49 | 1.99 | 1.97 |
| Metasomal segment III depth | 1.88 | 1.92 | 1.79 | 1.56 | 1.64 | 1.74 | 2.15 | 2.26 |
| Metasomal segment IV length | 8.2 | 9.14 | 9.1 | 8.09 | 9.61 | 11.07 | 5.21 | 6.41 |
| Metasomal segment IV width | 1.33 | 1.56 | 1.5 | 1.42 | 1.28 | 1.38 | 1.91 | 1.86 |
| Metasomal segment IV depth | 1.6 | 1.89 | 1.8 | 1.75 | 1.67 | 2.88 | 2.08 | 2.17 |
| Metasomal segment V length | 9.57 | 10.09 | 9.79 | 8.38 | 10.99 | 11.65 | 6.49 | 7.91 |
| Metasomal segment V width | 1.32 | 1.66 | 1.48 | 1.42 | 1.51 | 1.38 | 1.96 | 1.92 |
| Metasomal segment V depth | 1.67 | 2.3 | 2.06 | 1.66 | 1.77 | 1.95 | 2.22 | 2.18 |
| Telson length | 6.48 | 6.61 | 6.51 | 6.04 | 7.07 | 8.2 | 5.74 | 6.24 |
| Telson vesicle width | 1.65 | 1.84 | 1.88 | 1.6 | 1.61 | 1.5 | 2.58 | 2.6 |
| Telson vesicle depth | 2.01 | 2.19 | 2.1 | 1.72 | 1.91 | 1.98 | 2.59 | 2.62 |
| Pectinal teeth (sinistral/dextral) | 12/13 | 13/13 | 13/13 | 13/14 | 13/13 | 13/13 | 10/10 | 9/10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saman, E.A.G.; Barahoei, H.; Dehghan, H.; Oshaghi, M.A.; Rafinejad, J.; Azarm, A.; Prendini, L. A New Species of the Medically Important Scorpion Genus, Hemiscorpius Peters, 1861 (Hemiscorpiidae), from Southern Iran. Diversity 2025, 17, 321. https://doi.org/10.3390/d17050321
Saman EAG, Barahoei H, Dehghan H, Oshaghi MA, Rafinejad J, Azarm A, Prendini L. A New Species of the Medically Important Scorpion Genus, Hemiscorpius Peters, 1861 (Hemiscorpiidae), from Southern Iran. Diversity. 2025; 17(5):321. https://doi.org/10.3390/d17050321
Chicago/Turabian StyleSaman, Esmaeil Amiri Ghanat, Hossein Barahoei, Hossein Dehghan, Mohammad Ali Oshaghi, Javad Rafinejad, Amrollah Azarm, and Lorenzo Prendini. 2025. "A New Species of the Medically Important Scorpion Genus, Hemiscorpius Peters, 1861 (Hemiscorpiidae), from Southern Iran" Diversity 17, no. 5: 321. https://doi.org/10.3390/d17050321
APA StyleSaman, E. A. G., Barahoei, H., Dehghan, H., Oshaghi, M. A., Rafinejad, J., Azarm, A., & Prendini, L. (2025). A New Species of the Medically Important Scorpion Genus, Hemiscorpius Peters, 1861 (Hemiscorpiidae), from Southern Iran. Diversity, 17(5), 321. https://doi.org/10.3390/d17050321