
An Integrative Analysis of the Specific Distinctness of Valvata (Cincinna) ambigua Westerlund, 1873 and Valvata (Cincinna) piscinalis (Müller, 1774) (Gastropoda: Valvatidae)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction

2. Materials and Methods
2.1. Materials
2.2. Morphometric Analyses
2.3. DNA Extraction, Amplification and Sequencing
2.4. Alignment and Analyses of DNA Sequences
3. Results
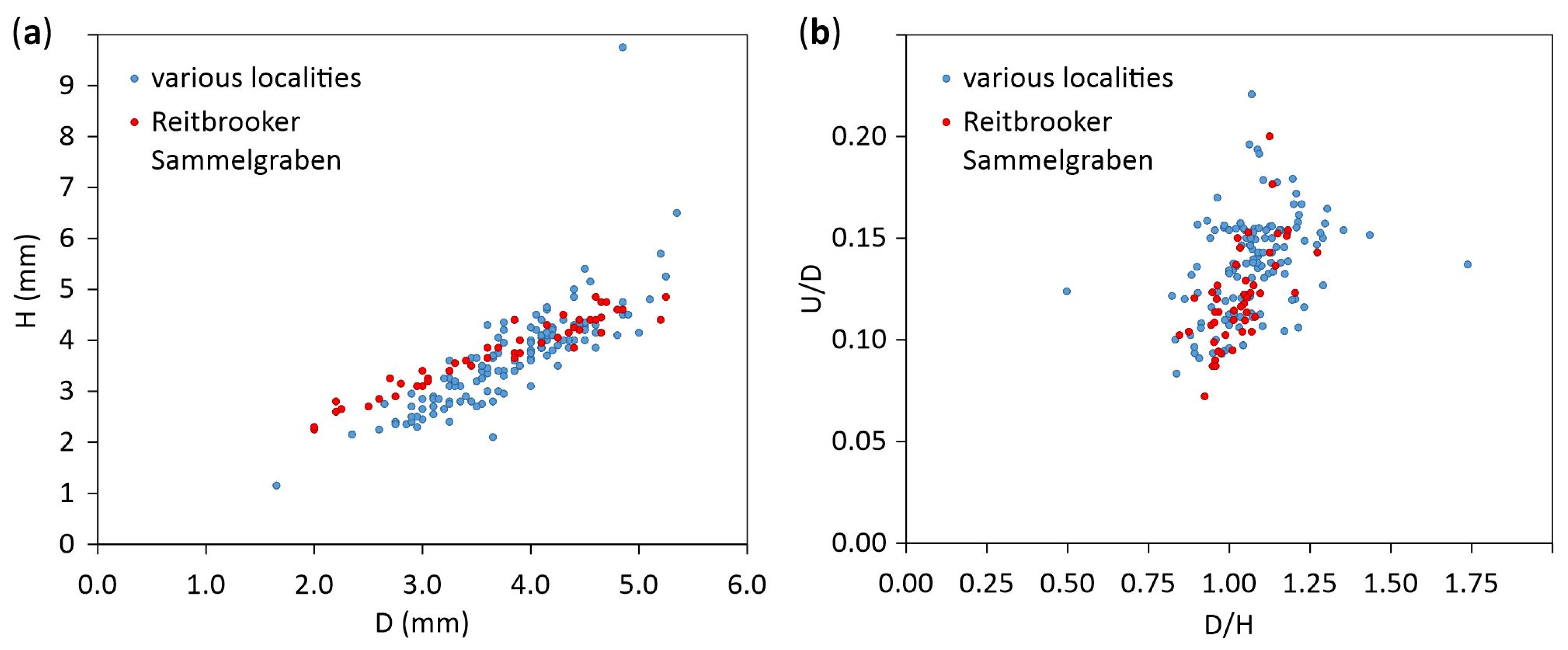
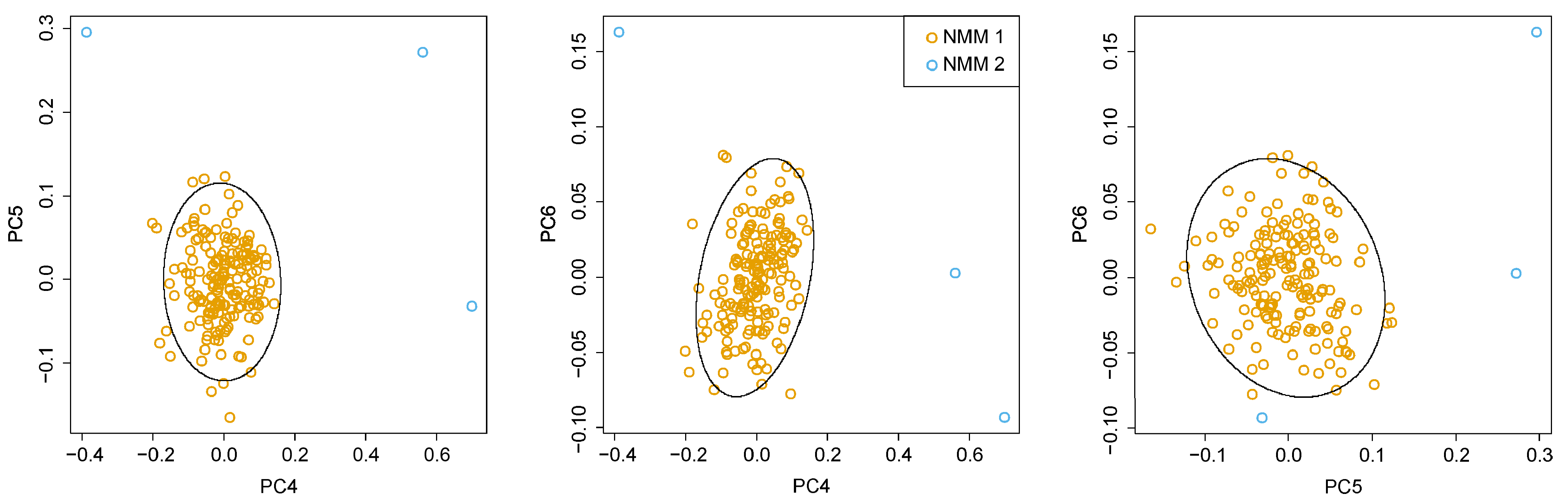
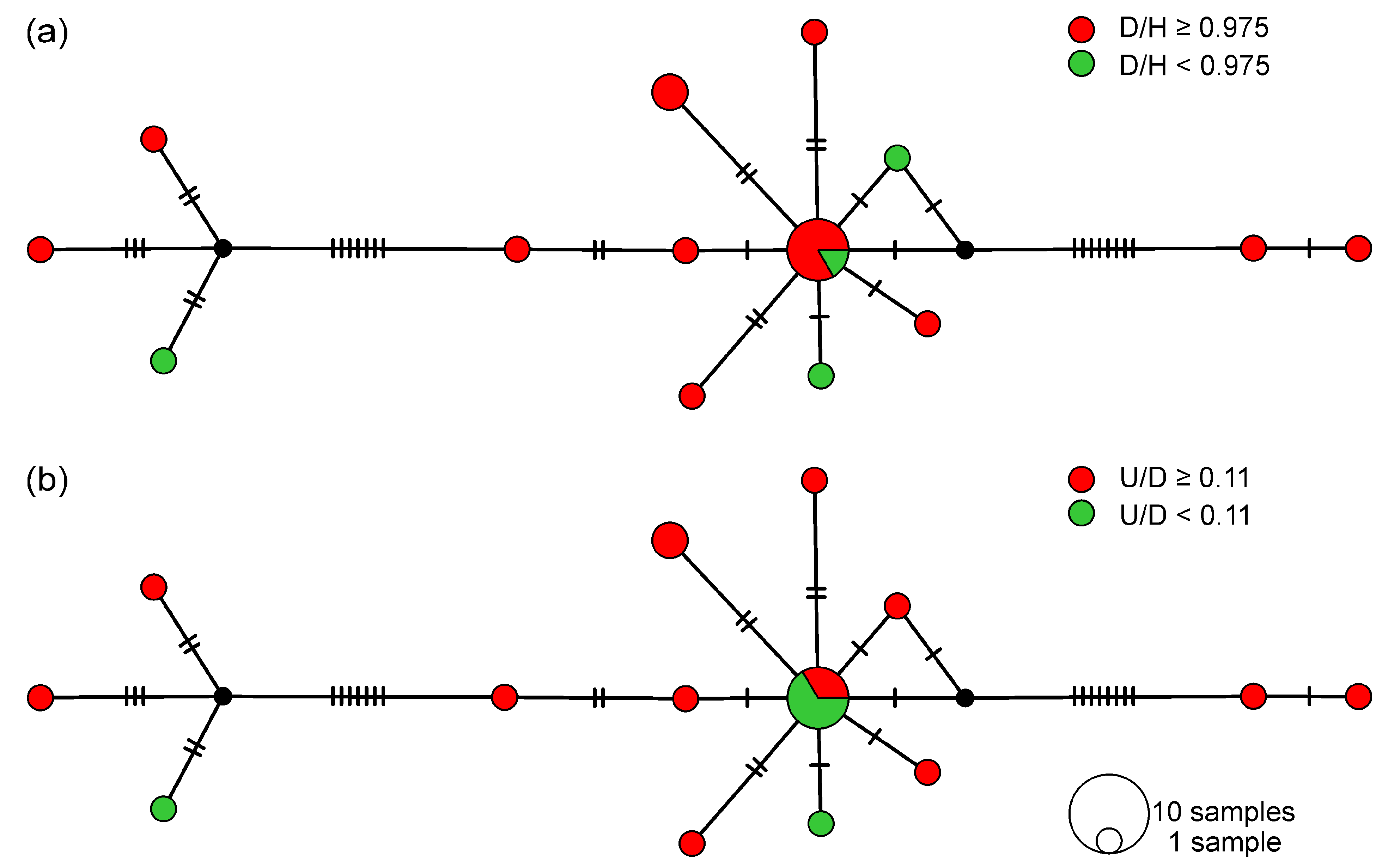
3.1. Shell Morphology
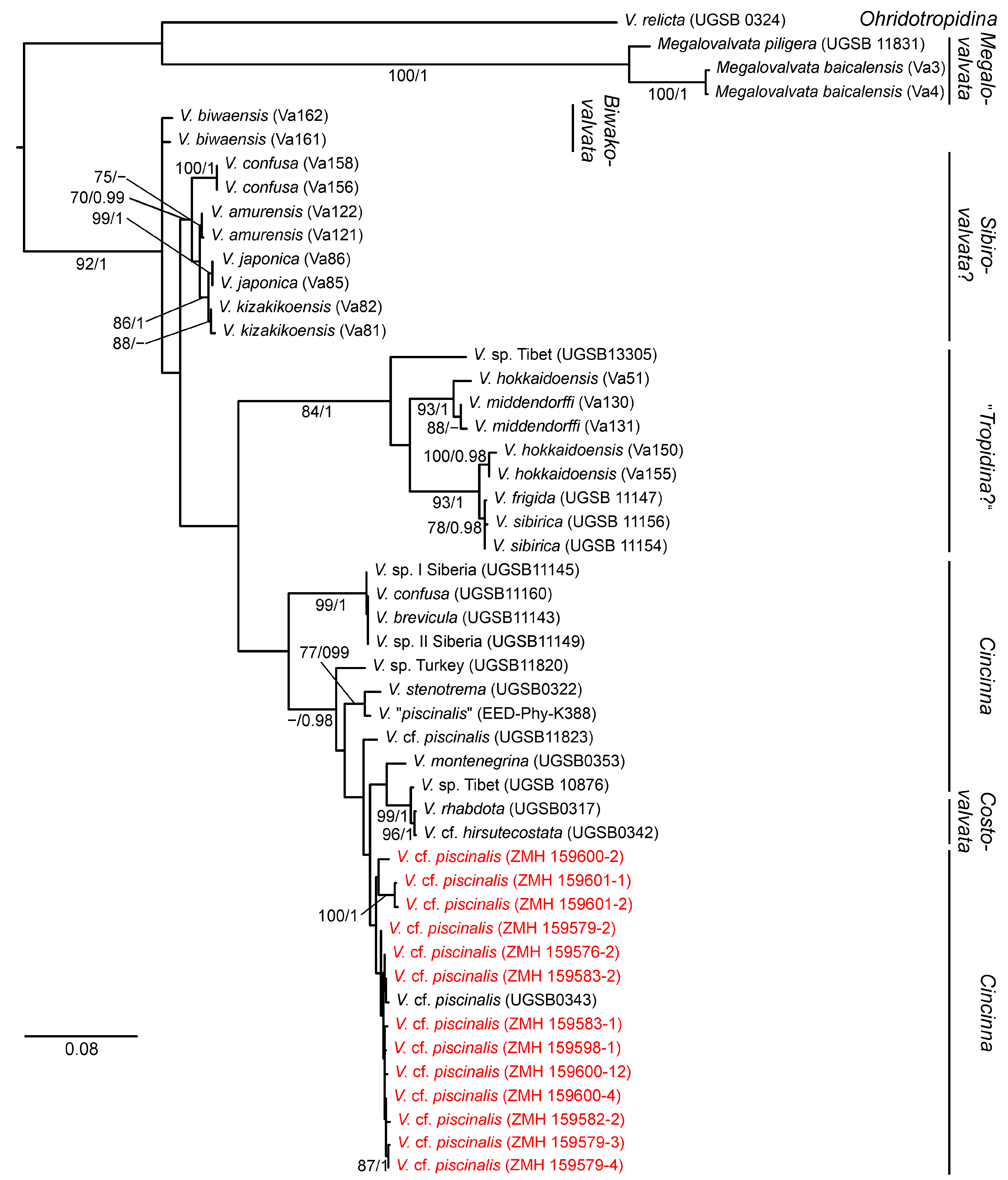
3.2. Molecular Phylogeny
4. Discussion
4.1. Delimitation of Valvata (Cincinna) ambigua Westerlund, 1873 and Valvata (Cincinna) piscinalis (Müller, 1774)
4.2. Phylogenetic Relationships
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Strong, E.E.; Gargominy, O.; Ponder, W.F.; Bouchet, P. Global diversity of gastropods (Gastropoda; Mollusca) in freshwater. Hydrobiologia 2008, 595, 149–166. [Google Scholar] [CrossRef]
- Haszprunar, G. A nomenclator of extant and fossil taxa of the Valvatidae (Gastropoda, Ectobranchia). ZooKeys 2014, 377, 1–172. [Google Scholar] [CrossRef] [PubMed]
- Falkner, G.; Bank, R.A.; von Proschwitz, T. Check-list of the non-marine molluscan species-group taxa of the states of Northern, Atlantic and Central Europe (CLECOM I). Heldia 2001, 4, 1–76. [Google Scholar]
- Glöer, P. Süsswassergastropoden Nord- und Mitteleuropas. Bestimmungsschlüssel, Lebensweise, Verbreitung. In Die Tierwelt Deutschlands und der Angrenzenden Meeresteile; ConchBooks: Hackenheim, Germany, 2002; Volume 73. [Google Scholar]
- Kantor, Y.I.; Sysoev, A.V. Katalog Mollyuskov Rossii i Sopregelnykh Stran; Tovarishchestvo nauchnykh izdaniy KMK: Moskva, Russia, 2005. [Google Scholar]
- Vinarski, M.V.; Kantor, Y.I. Analytical Catalogue of Fresh and Brackish Water Molluscs of Russia and Adjacent Countries; A.N. Severtsov Institute of Ecology and Evolution of Russian Academy of Sciences: Moskva, Russia, 2016. [Google Scholar]
- Glöer, P. The Freshwater Gastropods of the West-Palaearctis. Volume I. Fresh- and Brackish Waters except Spring and Subterranean Snails; P. Glöer: Hetlingen, Germany, 2019. [Google Scholar]
- Anistratenko, V.V. Opredelitel’ grebnezhabernykh mollyuskov (Gastropoda, Pectinibranchia) Fauny Ukrainy. Chast’ 2. Presnovodiye i nazemnye. Vestn. Zool. Suppl. 1998, 8, 67–117. [Google Scholar]
- Starobogatov, Y.I.; Prozorova, L.A.; Bogatov, V.V.; Saenko, E.M. Mollyuski. In Opredelitel’ Presnovodnykh Bespozvonochnykh Rossii i Sopredel’nykh Territoriy; Tsalolikhin, S.J., Ed.; Nauka: Saint Petersburg, Russia, 2004; Volume 6, pp. 9–492. [Google Scholar]
- Westerlund, C.A. Fauna Molluscorum Terrestrium et Fluviatilium Sveciae, Norvegiae et Daniae. Sveriges, Norges och Danmarks Land- Och Sötvatten-Mollusker. II. Sötvattenmollusker; A. Bonnier: Stockholm, Sweden, 1873. [Google Scholar]
- Westerlund, C.A. Synopsis molluscorum extramarinorum Scandinaviae (Sueciae, Norvegiae, Daniae & Fenniae). Acta Soc. Fauna Flora Fenn. 1897, 13, 1–238. [Google Scholar]
- Krivosheina, L.B.; Starobogatov, Y.I. Sostav i zoogeograficheskaya kharakteristika presnovodnoy malakofauny gornoy chasti basseyna berkhnego Irtysha. Zool. Zhurnal 1973, 22, 348–355. [Google Scholar]
- Glöer, P.; Diercking, R. Atlas der Süßwassermollusken. Rote Liste, Verbreitung, Ökologie, Bestand und Schutz; Behörde für Stadtentwicklung und Umwelt Hamburg: Hamburg, Germany, 2010. [Google Scholar]
- Vinarski, M.V.; Glöer, P.; Andreyeva, S.I.; Lazutkina, E.A. Taxonomic notes on Euro-Siberian molluscs. 5. Valvata (Cincinna) ambigua Westerlund 1873(7)—A distinct species of the group of Valvata piscinalis O.F. Müller 1774. J. Conchol. 2013, 41, 295–303. [Google Scholar]
- McLachlan, G.; Peel, D. Finite Mixture Models; John Wiley & Sons: New York, NY, USA, 2000. [Google Scholar]
- Ezard, T.H.G.; Pearson, P.N.; Purvis, A. Algorithmic approaches to aid species’ delimitation in multidimensional morphospace. BMC Evol. Biol. 2010, 10, 175. [Google Scholar] [CrossRef]
- Guillot, G.; Renaud, S.; Ledevin, R.; Michaux, J.; Claude, J. A unifying model for the analysis of phenotypic, genetic, and geographic data. Syst. Biol. 2012, 61, 897–911. [Google Scholar] [CrossRef]
- Cadena, C.D.; Zapata, F.; Jiménez, I. Issues and perspectives in species delimitation using phenotypic data: Atlantean evolution in Darwin’s finches. Syst. Biol. 2018, 67, 181–194. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing, Version 4.2.0. R Foundation for Statistical Computing. 2022. Available online: http://CRAN.R-project.org/ (accessed on 18 June 2024).
- Scrucca, L.; Raftery, A.E. clustvarsel: A package implementing variable selection for Gaussian model-based clustering in R. J. Stat. Softw. 2018, 84, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Scrucca, L.; Fop, M.; Murphy, T.B.; Raftery, A.E. mclust 5: Clustering, classification and density estimation using Gaussian finite mixture models. R J. 2016, 8, 289–317. [Google Scholar] [CrossRef] [PubMed]
- Sokolov, E.P. An improved method for DNA isolation from mucopolysaccharide-rich molluscan tissues. J. Molluscan Stud. 2000, 66, 573–575. [Google Scholar] [CrossRef]
- Scheel, B.; Hausdorf, B. Survival and differentiation of subspecies of the land snail Charpentieria itala in mountain refuges in the Southern Alps. Mol. Ecol. 2012, 21, 3794–3808. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Marine Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Palumbi, S.R.; Martin, A.P.; Romano, S.; McMillan, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR; Department of Zoology, University of Hawaii: Honolulu, HI, USA, 1991. [Google Scholar]
- Hauswald, A.K.; Albrecht, C.; Wilke, T. Testing two contrasting evolutionary patterns in ancient lakes: Species flock versus species scatter in valvatid gastropods of Lake Ohrid. Hydrobiologia 2008, 615, 169–179. [Google Scholar] [CrossRef]
- Dinapoli, A.; Klussmann-Kolb, A. The long way to diversity—Phylogeny and evolution of the Heterobranchia (Mollusca: Gastropoda). Mol. Phylogenet. Evol. 2010, 55, 60–76. [Google Scholar] [CrossRef] [PubMed]
- Clewing, C.; von Oheimb, P.V.; Vinarski, M.; Wilke, T.; Albrecht, C. Freshwater mollusc diversity at the roof of the world: Phylogenetic and biogeographical affinities of Tibetan Plateau Valvata. J. Molluscan Stud. 2014, 80, 452–455. [Google Scholar] [CrossRef]
- Saito, T.; Prozorova, L.; Sitnikova, T.; Surenkhorloo, P.; Hirano, T.; Morii, Y.; Chiba, S. Molecular phylogeny of glacial relict species: A case of freshwater Valvatidae molluscs (Mollusca: Gastropoda) in North and East Asia. Hydrobiologia 2018, 818, 105–118. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New models and efficient methods for phylogenetic inference in the genomic era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Patzner, R.A.; Glöer, P. Vorkommen von Valvata (Cincinna) ambigua Westerlund 1873 im Süden von Deutschland. Mitteilungen Dtsch. Malakozool. Ges. 2020, 102, 51–52. [Google Scholar]
- Borleis, F. Wiederfund von Pisidium moitessierianum Paladilhe 1866 sowie Nachweise von Valvata ambigua Westerlund 1873 und Corbicula fluminea (O. F. Müller 1774) in Leipzig. Mitteilungen Dtsch. Malakozool. Ges. 2021, 104, 65–68. [Google Scholar]
- Smythe, A.B.; Forgrave, K.; Patti, A.; Hochberg, R.; Litvaitis, M.K. Untangling the ecology, taxonomy, and evolution of Chaetogaster limnaei (Oligochaeta: Naididae) species complex. J. Parasitol. 2015, 101, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Mack, J.M.; Klinth, M.; Martinsson, S.; Lu, R.; Stormer, H.; Hanington, P.; Proctor, H.C.; Erséus, C.; Bely, A.E. Cryptic carnivores: Intercontinental sampling reveals extensive novel diversity in a genus of freshwater annelids. Mol. Phylogenet. Evol. 2023, 182, 107748. [Google Scholar] [CrossRef]
- Buse, A. The relationship of Chaetogaster limnaei (Oligochaeta: Naididae) with a variety of gastropod species. J. Anim. Ecol. 1974, 43, 821–837. [Google Scholar] [CrossRef]
- Conn, D.B.; Ricciardi, A.; Babapulle, M.N.; Klein, K.A.; Rosen, D.A. Chaetogaster limnaei (Annelida: Oligochaeta) as a parasite of the zebra mussel Dreissena polymorpha, and the quagga mussel Dreissena bugensis (Mollusca: Bivalvia). Parasitol. Res. 1996, 82, 1–7. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schäffer, M.; Hausdorf, B. An Integrative Analysis of the Specific Distinctness of Valvata (Cincinna) ambigua Westerlund, 1873 and Valvata (Cincinna) piscinalis (Müller, 1774) (Gastropoda: Valvatidae). Diversity 2024, 16, 419. https://doi.org/10.3390/d16070419
Schäffer M, Hausdorf B. An Integrative Analysis of the Specific Distinctness of Valvata (Cincinna) ambigua Westerlund, 1873 and Valvata (Cincinna) piscinalis (Müller, 1774) (Gastropoda: Valvatidae). Diversity. 2024; 16(7):419. https://doi.org/10.3390/d16070419
Chicago/Turabian StyleSchäffer, Maibritt, and Bernhard Hausdorf. 2024. "An Integrative Analysis of the Specific Distinctness of Valvata (Cincinna) ambigua Westerlund, 1873 and Valvata (Cincinna) piscinalis (Müller, 1774) (Gastropoda: Valvatidae)" Diversity 16, no. 7: 419. https://doi.org/10.3390/d16070419
APA StyleSchäffer, M., & Hausdorf, B. (2024). An Integrative Analysis of the Specific Distinctness of Valvata (Cincinna) ambigua Westerlund, 1873 and Valvata (Cincinna) piscinalis (Müller, 1774) (Gastropoda: Valvatidae). Diversity, 16(7), 419. https://doi.org/10.3390/d16070419