

Severe Disturbance of Chinese Pangolins Caused by Free-Ranging Domestic Dogs in Unprotected Areas

 and
and
Abstract
1. Introduction
2. Materials and Methods
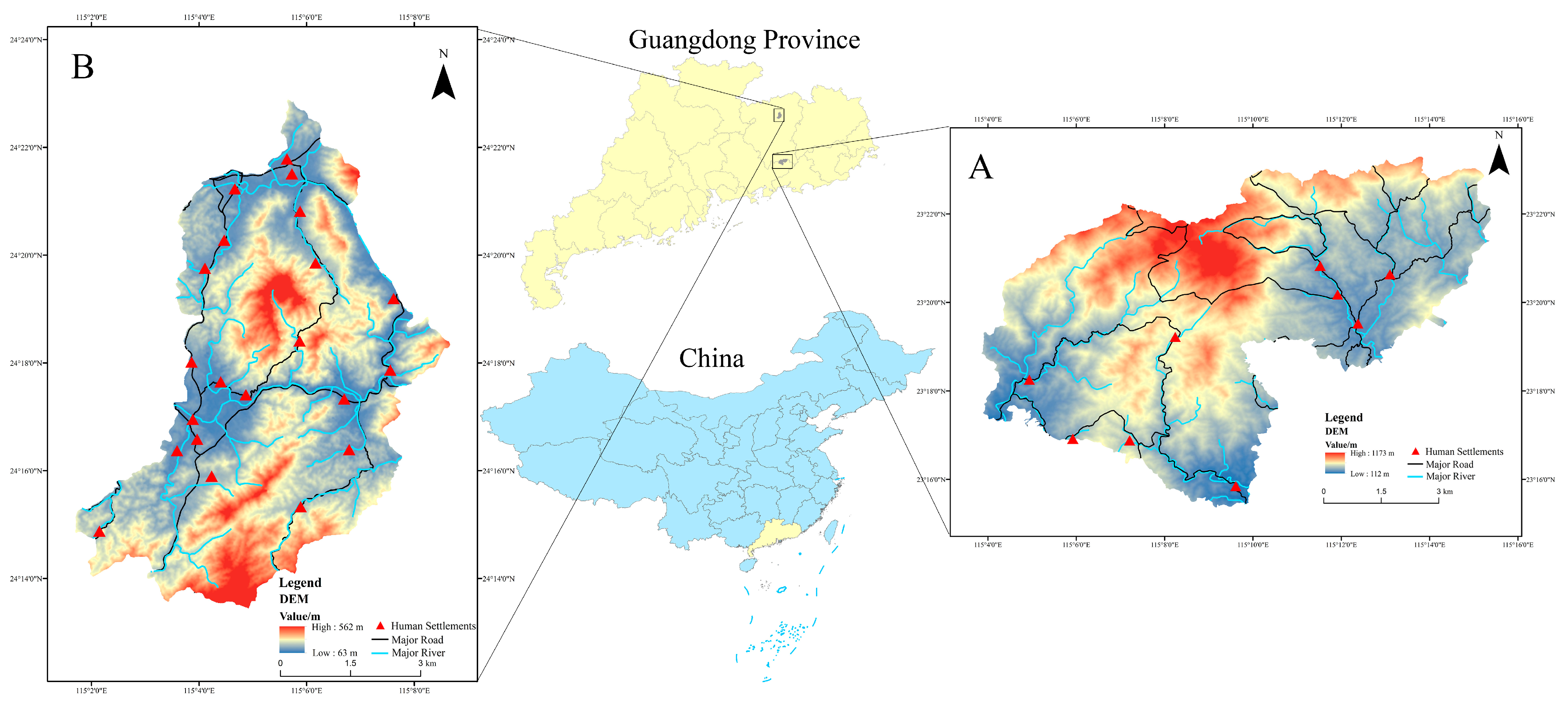
2.1. Study Area and Data Acquisition
2.1.1. Occurrences of Chinese Pangolins and Activity Rhythms
2.1.2. GPS Collars Tracking Free-Ranging Domestic Dogs
2.1.3. Environmental Variables
2.2. Constructing a Habitat Model for Chinese Pangolins
2.3. Spatial Relationship between Chinese Pangolins and Domestic Dogs
2.4. Temporal Analysis of Chinese Pangolins and Domestic Dogs
3. Results
3.1. Potentially Suitable Habitats of Chinese Pangolins
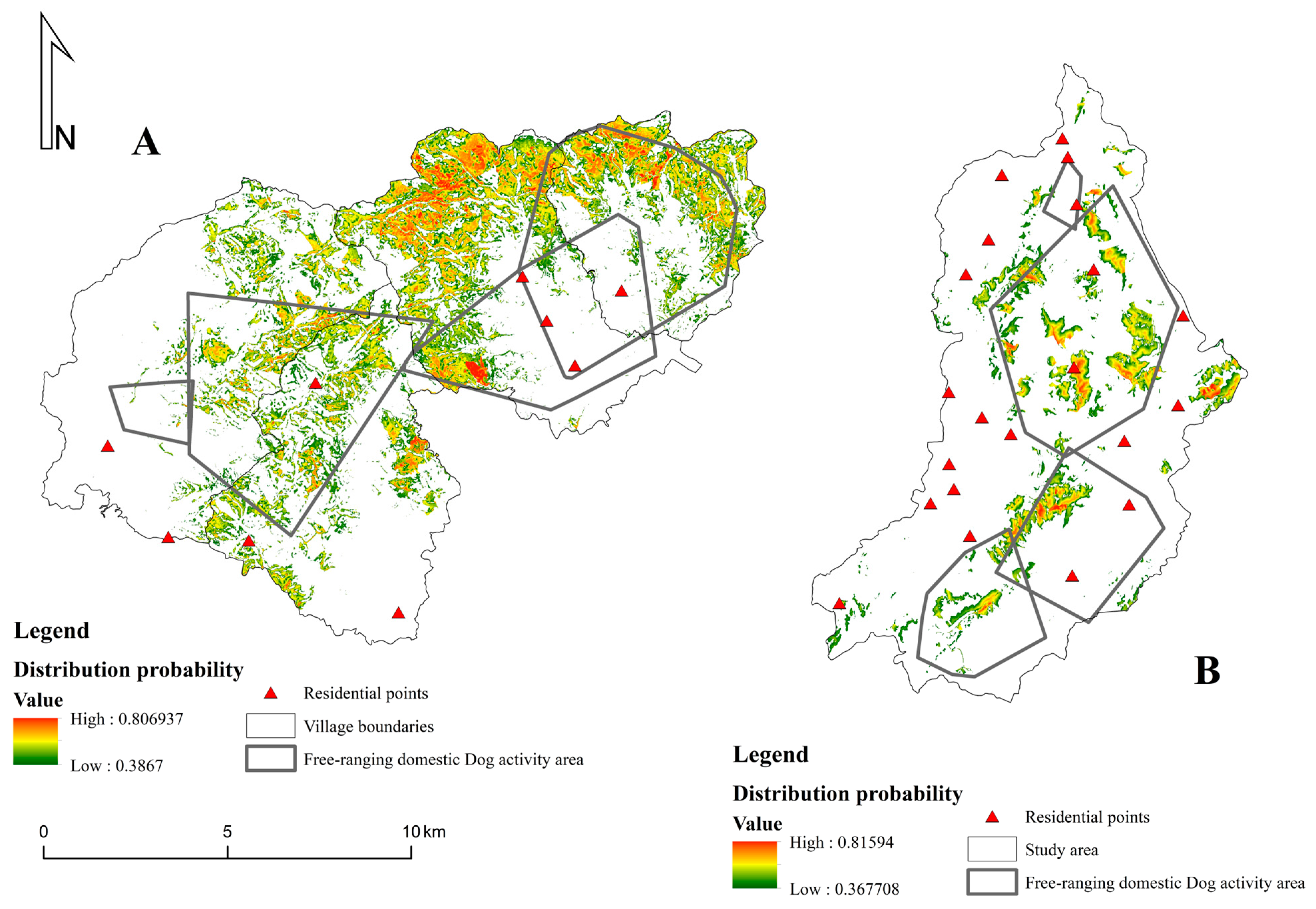
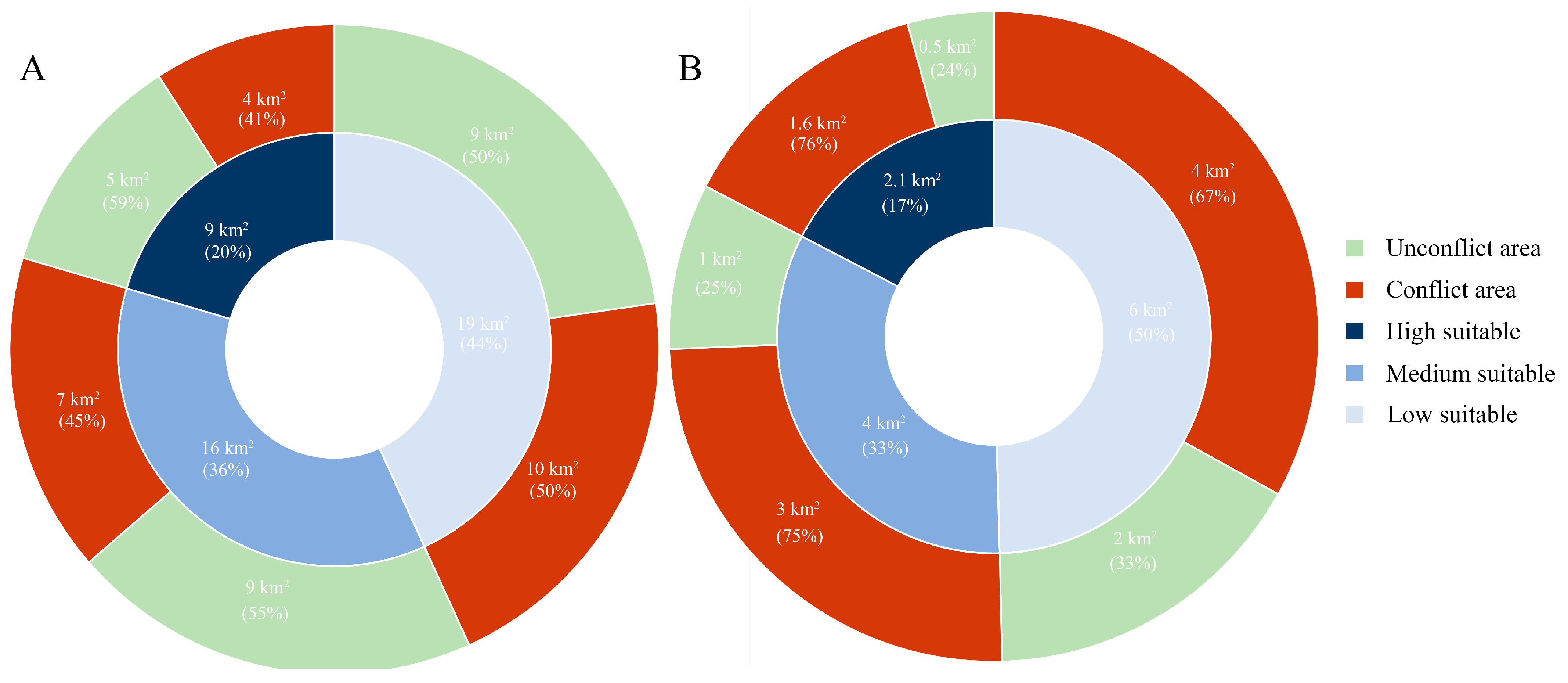
3.2. Spatial Conflicts between Chinese Pangolins and Free-Ranging Domestic Dogs
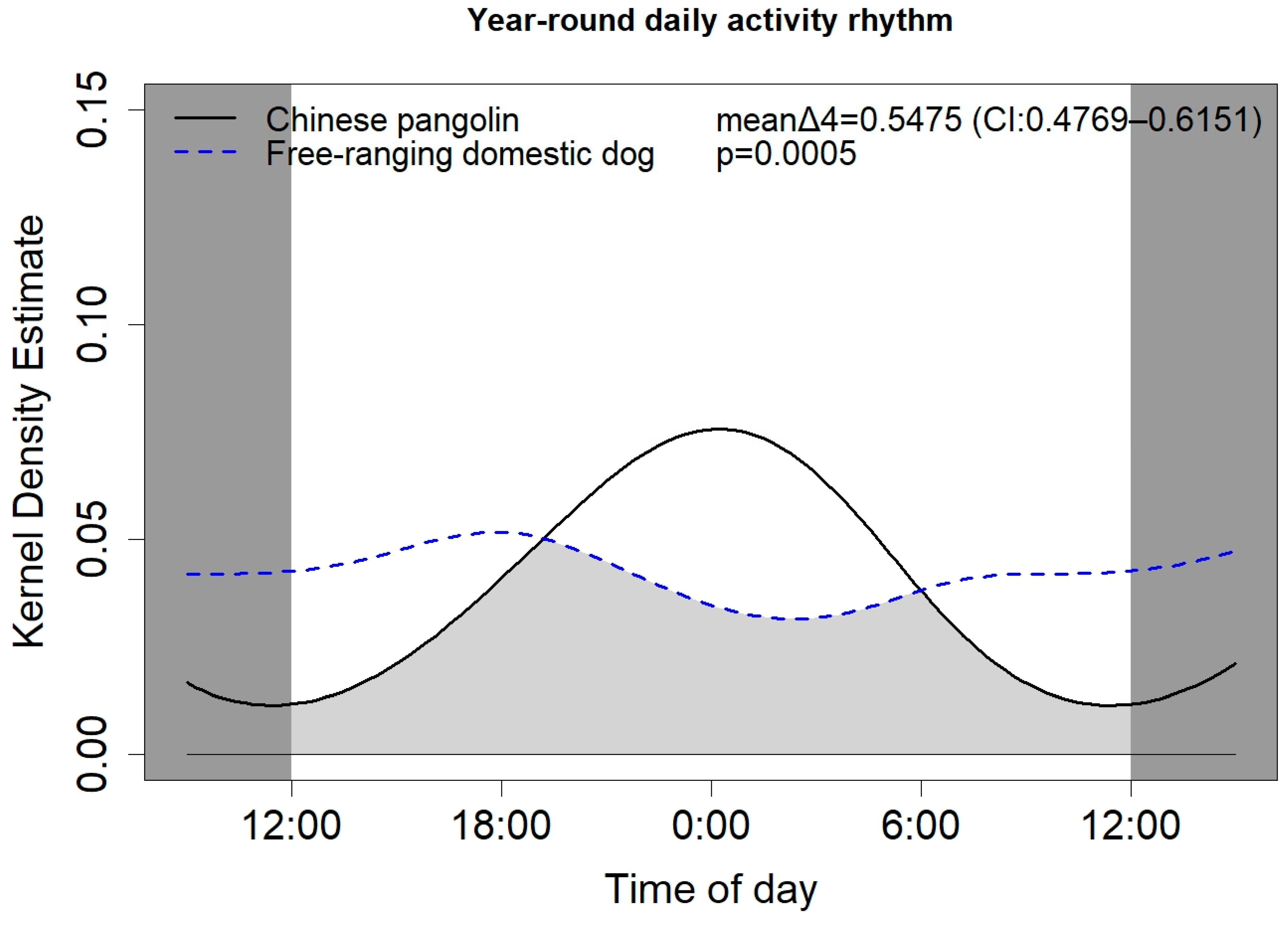
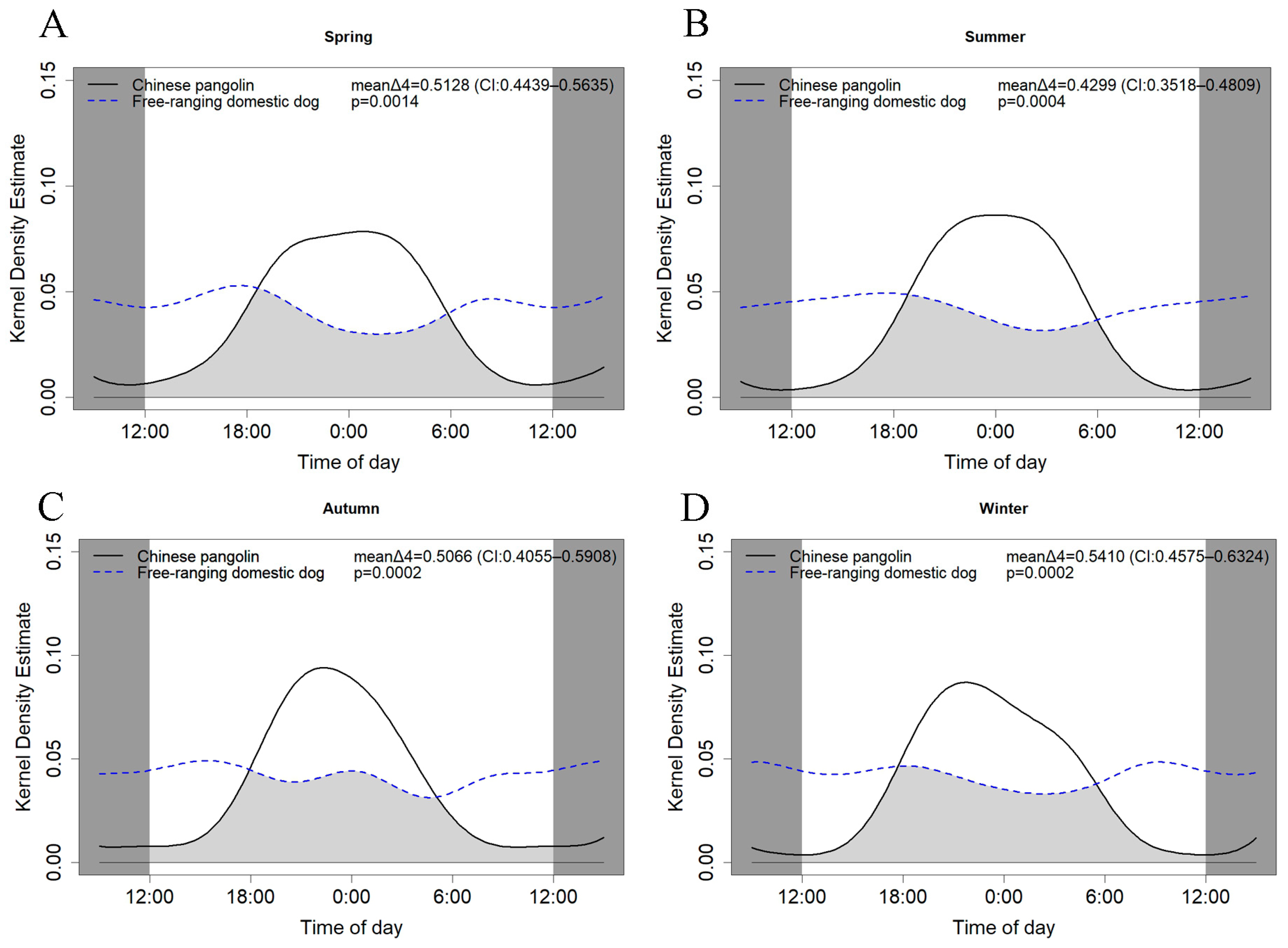
3.3. Temporal Niche Differentiation between Chinese Pangolins and Free-Ranging Domestic Dogs
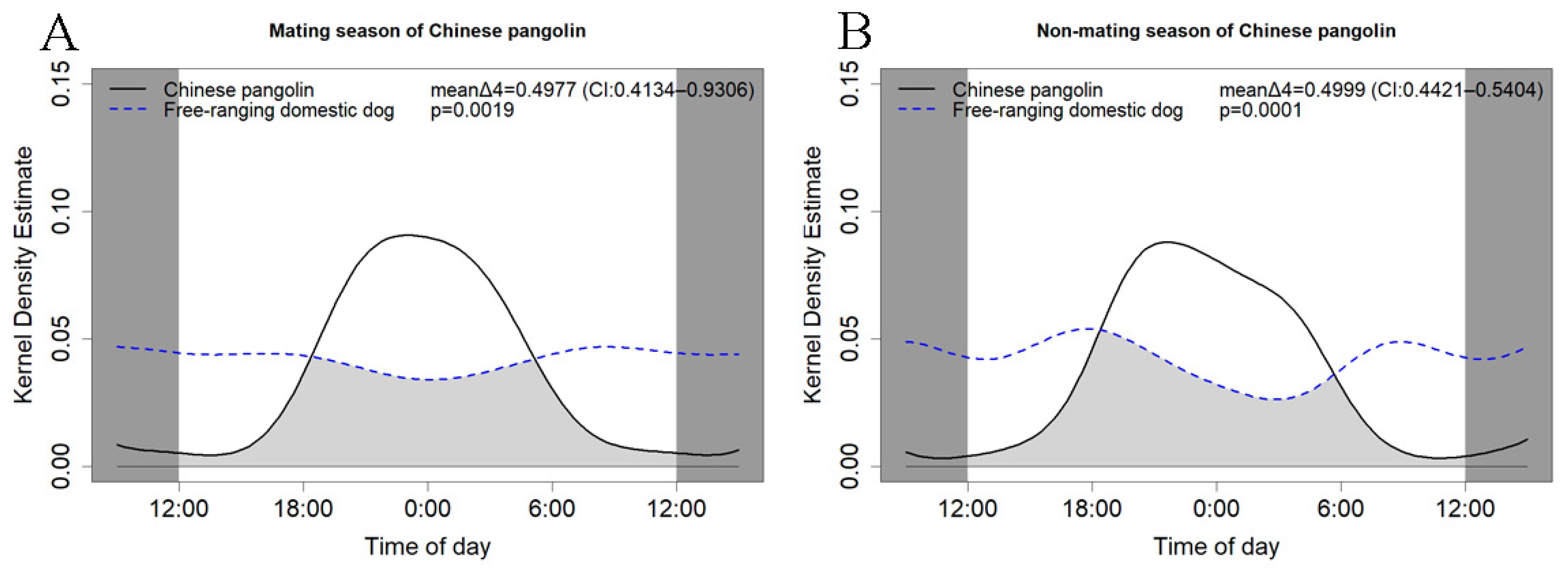
3.4. Attraction–Avoidance Ratio between Chinese Pangolins and Domestic Dogs
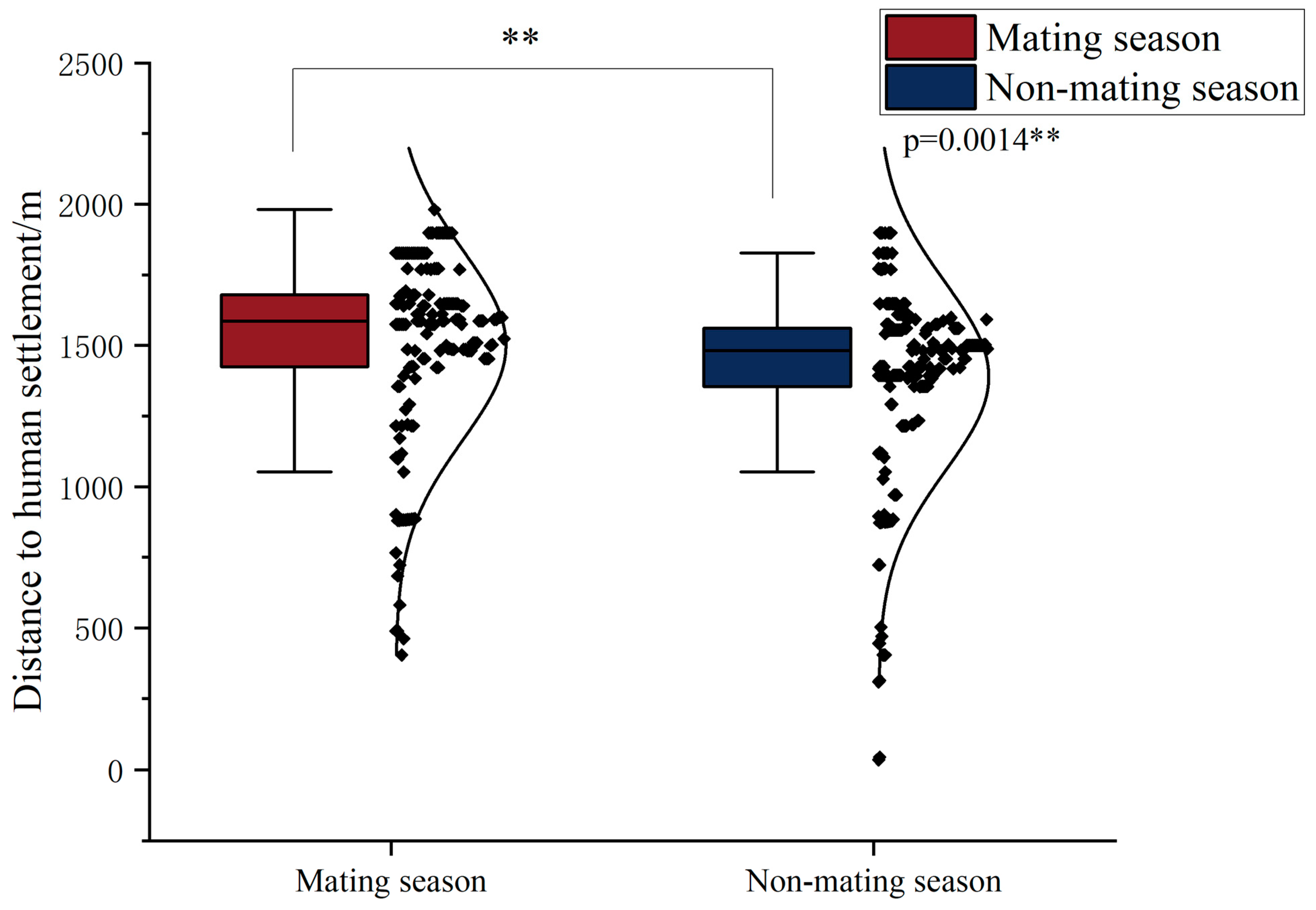
3.5. Distance to Human Settlement in Different Seasons
4. Discussion
4.1. Characteristics of Chinese Pangolin Habitats
4.2. Spatial Conflicts between Chinese Pangolins and Free-Ranging Domestic Dogs
4.3. Characteristics and Differences in Activity Rhythm between Chinese Pangolins and Free-Ranging Domestic Dogs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hughes, J.; Macdonald, D.W. A review of the interactions between free-roaming domestic dogs and wildlife. Biol. Conserv. 2013, 157, 341–351. [Google Scholar] [CrossRef]
- Ferreira, J.P.; Leitão, I.; Santos-Reis, M.; Revilla, E. Human-related factors regulate the spatial ecology of domestic cats in sensitive areas for conservation. PLoS ONE 2011, 6, e25970. [Google Scholar] [CrossRef] [PubMed]
- Doherty, T.S.; Dickman, C.R.; Glen, A.S.; Newsome, T.M.; Nimmo, D.G.; Ritchie, E.G.; Vanak, A.T.; Wirsing, A.J. The global impacts of domestic dogs on threatened vertebrates. Biol. Conserv. 2017, 210, 56–59. [Google Scholar] [CrossRef]
- Driscoll, C.A.; Macdonald, D.W. Top dogs: Wolf domestication and wealth. J. Biol. 2010, 9, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Vilà, C.; Savolainen, P.; Maldonado, J.E.; Amorim, I.R.; Rice, J.E.; Honeycutt, R.L.; Crandall, K.A.; Lundeberg, J.; Wayne, R.K. Multiple and ancient origins of the domestic dog. Science 1997, 276, 1687–1689. [Google Scholar] [CrossRef] [PubMed]
- Zapata-Ríos, G.; Branch, L.C. Altered activity patterns and reduced abundance of native mammals in sites with feral dogs in the high Andes. Biol. Conserv. 2016, 193, 9–16. [Google Scholar] [CrossRef]
- Jin, Y.P.; Zhang, X.K.; Ma, Y.S.; Qiao, Y.C.; Liu, X.B.; Zhao, K.H.; Zhang, C.L.; Lin, D.G.; Fu, X.L.; Xu, X.R.; et al. Canine distemper viral infection threatens the giant panda population in China. Oncotarget 2017, 8, 113910–113919. [Google Scholar] [CrossRef]
- Zhang, L.N.; Wang, K.; An, F.Y.; Zhang, D.L.; Zhang, H.L.; Xu, X.L.; Guo, C.; Yan, H.; Kuang, Y.; Zhang, Z.; et al. Fatal canine parvovirus type 2a and 2c infections in wild Chinese pangolins (Manis pentadactyla) in southern China. Transbound. Emerg. Dis. 2022, 69, 4002–4008. [Google Scholar]
- Wang, S.L.; Tu, Y.C.; Lee, M.S.; Wu, L.H.; Chen, T.Y.; Wu, C.H.; Tsao, E.H.; Chin, S.C.; Li, W.T. Fatal canine parvovirus-2 (CPV-2) infection in a rescued free-ranging Taiwanese pangolin (Manis pentadactyla pentadactyla). Transbound. Emerg. Dis. 2020, 67, 1074–1081. [Google Scholar] [CrossRef]
- Atickem, A.; Bekele, A.; Williams, S.D. Competition between domestic dogs and Ethiopian wolf (Canis simensis) in the Bale Mountains National Park, Ethiopia. Afr. J. Ecol. 2010, 48, 401–407. [Google Scholar] [CrossRef]
- Home, C.; Bhatnagar, Y.V.; Vanak, A.T. Canine Conundrum: Domestic dogs as an invasive species and their impacts on wildlife in India. Anim. Conserv. 2018, 21, 275–282. [Google Scholar] [CrossRef]
- Marshall, H.E.; Gore, M.L.; Ngoprasert, D.; Savini, T. Free-ranging dogs and their owners: Evaluating demographics, husbandry practices and local attitudes towards canine management and dog–wildlife conflict. Integr. Conserv. 2024, 2, 255–270. [Google Scholar] [CrossRef]
- Loss, S.R.; Will, T.; Marra, P.P. The impact of free-ranging domestic cats on wildlife of the United States. Nat. Commun. 2013, 4, 1396. [Google Scholar] [CrossRef]
- Marshall, H.E.; Niti, S.; Dusit, N.; Tommaso, S. The spatial and temporal displacement of native species by domestic dogs. Glob. Ecol. Conserv. 2023, 44, e02504. [Google Scholar] [CrossRef]
- Wu, S.B.; Sun, N.C.M.; Zhang, F.H.; Yu, Y.S.; Ades, G.; Suwal, T.L.; Jiang, Z.G. Chinese pangolin Manis pentadactyla (Linnaeus, 1758). In Pangolins; Elsevier: Amsterdam, The Netherlands, 2020; pp. 49–70. [Google Scholar]
- Wu, S.B.; Liu, N.F.; Ma, G.Z.; Tang, M.; Chen, H.; Xu, Z.R. A Current Situation of Ecology Study on Pangolins. Chin. J. Zool. 2004, 39, 46–52. [Google Scholar]
- Wu, S.B.; Liu, N.F.; Ma, G.Z.; Xu, Z.R.; Chen, H. Habitat selection by Chinese pangolin (Manis pentadactyla) in winter in Dawuling Natural Reserve. Mammalia 2003, 67, 493–502. [Google Scholar] [CrossRef]
- Zhou, Z.M.; Zhao, H.; Zhang, Z.X.; Wang, Z.H.; Wang, H. Allometry of scales in Chinese pangolins (Manis pentadactyla) and Malayan pangolins (Manis javanica) and application in judicial expertise. Zool. Res. 2012, 33, 271–275. [Google Scholar] [CrossRef]
- Challender, D. Asian Pangolins: How behavioural research can contribute to their conservation. In Proceedings of the Workshop on Trade and Conservation of Pangolins Native to South and Southeast Asia, Singapore, 1 July 2008; p. 95. [Google Scholar]
- Challender, D.W.S.; Harrop, S.R.; MacMillan, D.C. Understanding markets to conserve trade-threatened species in CITES. Biol. Conserv. 2015, 187, 249–259. [Google Scholar] [CrossRef]
- Wu, S.B.; MA, G.Z.; Tang, M.; Chen, H.; Liu, N.F. The status and conservation strategy of pangolin resource in China. J. Od Nat. Resour. 2002, 17, 174–180. [Google Scholar]
- Tian, F.; Li, J.; Liu, W.; Liu, Y.; Hu, Y.; Tu, Q.; Li, Y.; Bai, Y.; Shi, M.; Que, T.; et al. Virome in healthy pangolins reveals compatibility with multiple potentially zoonotic viruses. Zool. Res. 2022, 43, 977–988. [Google Scholar] [CrossRef]
- Challender, D.W.; Nash, H.; Waterman, C. Pangolins: Science, Society and Conservation; Academic Press: Cambridge, MA, USA, 2019; p. 71. [Google Scholar]
- Butler, J.; Du Toit, J. Diet of free-ranging domestic dogs (Canis familiaris) in rural Zimbabwe: Implications for wild scavengers on the periphery of wildlife reserves. Anim. Conserv. Forum 2002, 5, 29–37. [Google Scholar] [CrossRef]
- Butler, J.; Du Toit, J.; Bingham, J. Free-ranging domestic dogs (Canis familiaris) as predators and prey in rural Zimbabwe: Threats of competition and disease to large wild carnivores. Biol. Conserv. 2004, 115, 369–378. [Google Scholar] [CrossRef]
- Gompper, M.E. Free-Ranging Dogs and Wildlife Conservation; Oxford University Press: New York, NY, USA, 2014; pp. 73–86. [Google Scholar]
- Ikeya, K. Hunting with dogs among the San in the central Kalahari. Afr. Study Monogr. 1994, 15, 119–134. [Google Scholar]
- Zhang, F.H.; Wang, W.H.; Mahmood, A.; Wu, S.B.; Li, J.Q.; Xu, N. Observations of Chinese pangolins (Manis pentadactyla) in mainland China. Glob. Ecol. Conserv. 2021, 26, e01460. [Google Scholar] [CrossRef]
- Sun, N.C.M.; Arora, B.; Lin, J.S.; Lin, W.C.; Chi, M.J.; Chen, C.C.; Pei, C.J.C. Mortality and morbidity in wild Taiwanese pangolin (Manis pentadactyla pentadactyla). PLoS ONE 2019, 14, e0198230. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, T.; Irshad, N.; Hussain, R.; Akrim, F.; Hussain, I.; Anwar, M.; Rais, M.; Nadeem, M.S. Breeding habits of the Indian pangolin (Manis crassicaudata) in Potohar Plateau, Pakistan. Mammalia 2016, 80, 231–234. [Google Scholar] [CrossRef]
- Hassan, M.; Sulaiman, M.H.; Lian, C.J. The prevalence and intensity of Amblyomma javanense infestation on Malayan Pangolins (Manis javanica Desmarest) from Peninsular Malaysia. Acta Trop. 2013, 126, 142–145. [Google Scholar] [CrossRef]
- Khatri-Chhetri, R.; Wang, H.C.; Chen, C.C.; Shih, H.C.; Liao, H.C.; Sun, C.M.; Khatri-Chhetri, N.; Wu, H.Y.; Pei, K.J.C. Surveillance of ticks and associated pathogens in free-ranging Formosan pangolins (Manis pentadactyla pentadactyla). Ticks Tick-Borne Dis. 2016, 7, 1238–1244. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Wang, M.; Nie, Y.; Swaisgood, R.R.; Wei, W.; Zhou, W.; Zhang, Z.; Wang, G.; Wei, F. Stable seasonal migration patterns in giant pandas. Zool. Res. 2023, 44, 341–348. [Google Scholar] [CrossRef]
- Sun, N.C.M.; Pei, K.J.C.; Lin, J.-S. Attaching tracking devices to pangolins: A comprehensive case study of Chinese pangolin Manis pentadactyla from southeastern Taiwan. Glob. Ecol. Conserv. 2019, 20, e00700. [Google Scholar] [CrossRef]
- Griss, S.; Riemer, S.; Warembourg, C.; Sousa, F.M.; Wera, E.; Berger-Gonzalez, M.; Alvarez, D.; Bulu, P.M.; Hernández, A.L.; Roquel, P. If they could choose: How would dogs spend their days? Activity patterns in four populations of domestic dogs. Appl. Anim. Behav. Sci. 2021, 243, 105449. [Google Scholar] [CrossRef]
- Abdelaal, M.; Fois, M.; Fenu, G.; Bacchetta, G. Using Maxent modeling to predict the potential distribution of the endemic plant Rosa arabica Crép. in Egypt. Ecol. Inform. 2019, 50, 68–75. [Google Scholar] [CrossRef]
- Abolmaali, S.M.-R.; Tarkesh, M.; Bashari, H. Maxent modeling for predicting suitable habitats and identifying the effects of climate change on a threatened species, Daphne mucronata, in central Iran. Ecol. Inform. 2018, 43, 116–123. [Google Scholar] [CrossRef]
- Seaman, D.E.; Millspaugh, J.J.; Kernohan, B.J.; Brundige, G.C.; Raedeke, K.J.; Gitzen, R.A. Effects of sample size on kernel home range estimates. J. Wildl. Manag. 1999, 739–747. [Google Scholar] [CrossRef]
- Long, J.A.; Nelson, T.A. Time geography and wildlife home range delineation. J. Wildl. Manag. 2012, 76, 407–413. [Google Scholar] [CrossRef]
- Tenorio, F.M.B.; Fernandez, D.A.P.; de Luna, M.C.T.; de Guia, A.P.O.; Balatibat, J.B.; Baril, J.A.; Aurellado, M.E.B. Cat’s Paw: Tracking the Home Range of Domestic Cats in Mount Makiling Forest Reserve. Philipp. J. Sci. 2024, 153, 353–361. [Google Scholar]
- Chen, Y.X.; Yu, Y.; Li, C.; Xiao, Z.S.; Zhou, G.W.; Zhang, Z.J.; Wang, X.W.; Xiang, Z.F.; Chang, J.; Li, M. Population and conservation status of a transboundary group of black snub-nosed monkeys (Rhinopithecus strykeri) between China and Myanmar. Zool. Res. 2022, 43, 523–527. [Google Scholar] [PubMed]
- Sun, N.C.M.; Pei, K.J.C.; Wu, L.Y. Long term monitoring of the reproductive behavior of wild Chinese pangolin (Manis pentadactyla). Sci. Rep. 2021, 11, 18116. [Google Scholar] [CrossRef]
- Chen, Y.X.; Xiao, Z.S.; Zhang, L.; Wang, X.W.; Li, M.; Xiang, Z.F. Activity rhythms of coexisting red serow and Chinese serow at Mt. Gaoligong as identified by camera traps. Animals 2019, 9, 1071. [Google Scholar] [CrossRef]
- You, Z.Y.; Lu, B.G.; Du, B.B.; Liu, W.; Jiang, Y.; Ruan, G.F.; Yang, N. Spatio-Temporal Niche of Sympatric Tufted Deer (Elaphodus cephalophus) and Sambar (Rusa unicolor) Based on Camera Traps in the Gongga Mountain National Nature Reserve, China. Animals 2022, 12, 2694. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.Q.; Liu, Y.L.; He, C.W.; Li, T.T.; Li, Q.L.; Ma, C.X.; Wang, D.J.; Li, S. Determining the daily activity pattern of Chinese mountain cat (Felis bieti): A comparative study based on camera-trapping and satellite collar tracking data. Biodivers. Sci. 2022, 30, 22081. [Google Scholar] [CrossRef]
- Ridout, M.S.; Linkie, M. Estimating overlap of daily activity patterns from camera trap data. J. Agric. Biol. Environ. Stat. 2009, 14, 322–337. [Google Scholar] [CrossRef]
- Parsons, A.W.; Bland, C.; Forrester, T.; Baker-Whatton, M.C.; Schuttler, S.G.; McShea, W.J.; Costello, R.; Kays, R. The ecological impact of humans and dogs on wildlife in protected areas in eastern North America. Biol. Conserv. 2016, 203, 75–88. [Google Scholar] [CrossRef]
- Weng, Y.; McShea, W.; Diao, Y.X.; Yang, H.B.; Zhang, X.F.; Gu, B.J.; Bu, H.L.; Wang, F. The incursion of free-ranging dogs into protected areas: A spatio-temporal analysis in a network of giant panda reserves. Biol. Conserv. 2022, 265, 109423. [Google Scholar] [CrossRef]
- Tiller, L.N.; Humle, T.; Amin, R.; Deere, N.J.; Lago, B.O.; Leader-Williams, N.; Sinoni, F.K.; Sitati, N.; Walpole, M.; Smith, R.J. Changing seasonal, temporal and spatial crop-raiding trends over 15 years in a human-elephant conflict hotspot. Biol. Conserv. 2021, 254, 108941. [Google Scholar] [CrossRef]
- Yun, J.H.; Kim, J.H.; Oh, K.H.; Lee, B.Y. Vertical distribution of vascular plants in Jungsanri, Mt. Jiri by temperature gradient. Korean J. Environ. Ecol. 2010, 24, 680–707. [Google Scholar]
- Wang, J.J.; Soininen, J.; Zhang, Y.; Wang, B.X.; Yang, X.; Shen, J. Patterns of elevational beta diversity in micro-and macroorganisms. Glob. Ecol. Biogeogr. 2012, 21, 743–750. [Google Scholar] [CrossRef]
- McCain, C.M. Global analysis of bird elevational diversity. Glob. Ecol. Biogeogr. 2009, 18, 346–360. [Google Scholar] [CrossRef]
- Underwood, A. Structure of a rocky intertidal community in New South Wales: Patterns of vertical distribution and seasonal changes. J. Exp. Mar. Biol. Ecol. 1981, 51, 57–85. [Google Scholar] [CrossRef]
- Houghton, D.; Stubbings, H. On the vertical distribution of Elminius modestus Darwin. J. Anim. Ecol. 1963, 32, 193–201. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Jackson, R.B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol. Appl. 2000, 10, 423–436. [Google Scholar] [CrossRef]
- Gao, H.Y.; Dou, H.L.; Wei, S.C.; Sun, S.; Zhang, Y.L.; Hua, Y. Local chronicles reveal the effect of anthropogenic and climatic impacts on local extinctions of Chinese pangolins (Manis pentadactyla) in mainland China. Ecol. Evol. 2022, 12, e9388. [Google Scholar] [CrossRef]
- Hammerschlag, N.; Fallows, C.; Meÿer, M.; Seakamela, S.M.; Orndorff, S.; Kirkman, S.; Kotze, D.; Creel, S. Loss of an apex predator in the wild induces physiological and behavioural changes in prey. Biol. Lett. 2022, 18, 20210476. [Google Scholar] [CrossRef] [PubMed]
- Ordiz, A.; Bischof, R.; Swenson, J.E. Saving large carnivores, but losing the apex predator? Biol. Conserv. 2013, 168, 128–133. [Google Scholar] [CrossRef]
- Kjellander, P.; Hewison, A.J.M.; Liberg, O.; Angibault, J.M.; Bideau, E.; Cargnelutti, B. Experimental evidence for density-dependence of home-range size in roe deer (Capreolus capreolus L.): A comparison of two long-term studies. Oecologia 2004, 139, 478–485. [Google Scholar] [CrossRef]
- Madsen, T. Movements, home range size and habitat use of radio-tracked grass snakes (Natrix natrix) in southern Sweden. Copeia 1984, 3, 707–713. [Google Scholar] [CrossRef]
- Wang, X.; Jia-Li Zhang, J.; Pan, H.; Chen, Y.; Mao, S.; Qi, J.; Shen, Y.; Zhang, M.; Xiang, Z.; Li, M. Unique characteristics of gut microbiota in black snub-nosed monkeys (Rhinopithecus strykeri) reveal an enzymatic mechanism of adaptation to dietary vegetation. Zool. Res. 2023, 44, 357–360. [Google Scholar] [CrossRef]
- Salafsky, N.; Salzer, D.; Stattersfield, A.J.; Hilton-Taylor, C.; Neugarten, R.; Butchart, S.H.; Collen, B.; Cox, N.; Master, L.L.; O’Connor, S.; et al. A standard lexicon for biodiversity conservation: Unified classifications of threats and actions. Conserv. Biol. 2008, 22, 897–911. [Google Scholar] [CrossRef] [PubMed]
- Pablo-Rodríguez, N.; Aurioles-Gamboa, D.; Montero-Muñoz, J.L. Niche overlap and habitat use at distinct temporal scales among the California sea lions (Zalophus californianus) and Guadalupe fur seals (Arctocephalus philippii townsendi). Mar. Mammal Sci. 2016, 32, 466–489. [Google Scholar] [CrossRef]
- Molloy, S.; Burleigh, A.; Dürr, S.; Ward, M.P. Roaming behaviour of dogs in four remote Aboriginal communities in the Northern Territory, Australia: Preliminary investigations. Aust. Vet. J. 2017, 95, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.Y.; Dou, H.L.; Wang, K.; Zhang, Y.Q.; Hua, Y. Ensemble SDMs reveal the effect of environmental suitability and nature reserves on conserving Chinese pangolins in Guangdong, China. J. Nat. Conserv. 2024, 79, 126617. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Variables | Range | |
|---|---|---|
| Topography | Elevation | 63–1173 m |
| Slope | 0–66° | |
| Aspect | 0–360° | |
| Vegetation | Forest type | (1) Pure coniferous forest; (2) pure broad-leaved forest; (3) relatively pure coniferous forest; (4) relatively pure broad-leaved forest; (5) mixed coniferous forest; (6) mixed coniferous broad-leaved forest; (7) mixed broad-leaved forest |
| Age of forest | (1) Young forest; (2) middle-aged forest; (3) near-mature forest; (4) mature forest; (5) overmature forest | |
| Human Interference | Distance to road | 0–3352 m |
| Distance to human settlement | 11–5976 m | |
| Water Source Distance | Distance to major river | 0–2194 m |
| Region | Variables | Percent Contribution/% | Permutation Importance/% |
|---|---|---|---|
| Wuqinzhang forest area | Forest type | 22.4 | 13.8 |
| Elevation | 16.3 | 16.1 | |
| Distance to road | 15.2 | 23.5 | |
| Slope | 13 | 14.4 | |
| Forest age | 10.4 | 10.7 | |
| Aspect | 9.5 | 10.6 | |
| Distance to major river | 6.9 | 4.5 | |
| Distance to human settlement | 6.3 | 6.1 | |
| Heping forest area | Elevation | 40 | 58.1 |
| Forest age | 21.3 | 12.9 | |
| Forest type | 20.7 | 10.2 | |
| Distance to road | 7.5 | 7.2 | |
| Aspect | 4.9 | 3.6 | |
| Distance to major river | 4.1 | 5.4 | |
| Distance to human settlement | 1.1 | 1.4 | |
| Slope | 0.4 | 1.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Gao, H.; Dou, H.; Yang, J.; Wang, J.; Xiang, Z.; Hua, Y. Severe Disturbance of Chinese Pangolins Caused by Free-Ranging Domestic Dogs in Unprotected Areas. Diversity 2024, 16, 386. https://doi.org/10.3390/d16070386
Zhang Y, Gao H, Dou H, Yang J, Wang J, Xiang Z, Hua Y. Severe Disturbance of Chinese Pangolins Caused by Free-Ranging Domestic Dogs in Unprotected Areas. Diversity. 2024; 16(7):386. https://doi.org/10.3390/d16070386
Chicago/Turabian StyleZhang, Yulin, Haiyang Gao, Hongliang Dou, Jinzhen Yang, Jingxin Wang, Zuofu Xiang, and Yan Hua. 2024. "Severe Disturbance of Chinese Pangolins Caused by Free-Ranging Domestic Dogs in Unprotected Areas" Diversity 16, no. 7: 386. https://doi.org/10.3390/d16070386
APA StyleZhang, Y., Gao, H., Dou, H., Yang, J., Wang, J., Xiang, Z., & Hua, Y. (2024). Severe Disturbance of Chinese Pangolins Caused by Free-Ranging Domestic Dogs in Unprotected Areas. Diversity, 16(7), 386. https://doi.org/10.3390/d16070386