
And Yet They Differ: Reconsiderations of Diversity within Dactylochelifer latreillii (Arachnida: Pseudoscorpiones)


Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. Laboratory Procedures and Phylogenetic Reconstruction
2.3. Morphometric Analysis
2.4. Geometric Morphometrics
2.5. Genitalic Morphology
3. Results
3.1. Habitat Segregation in Northern Germany
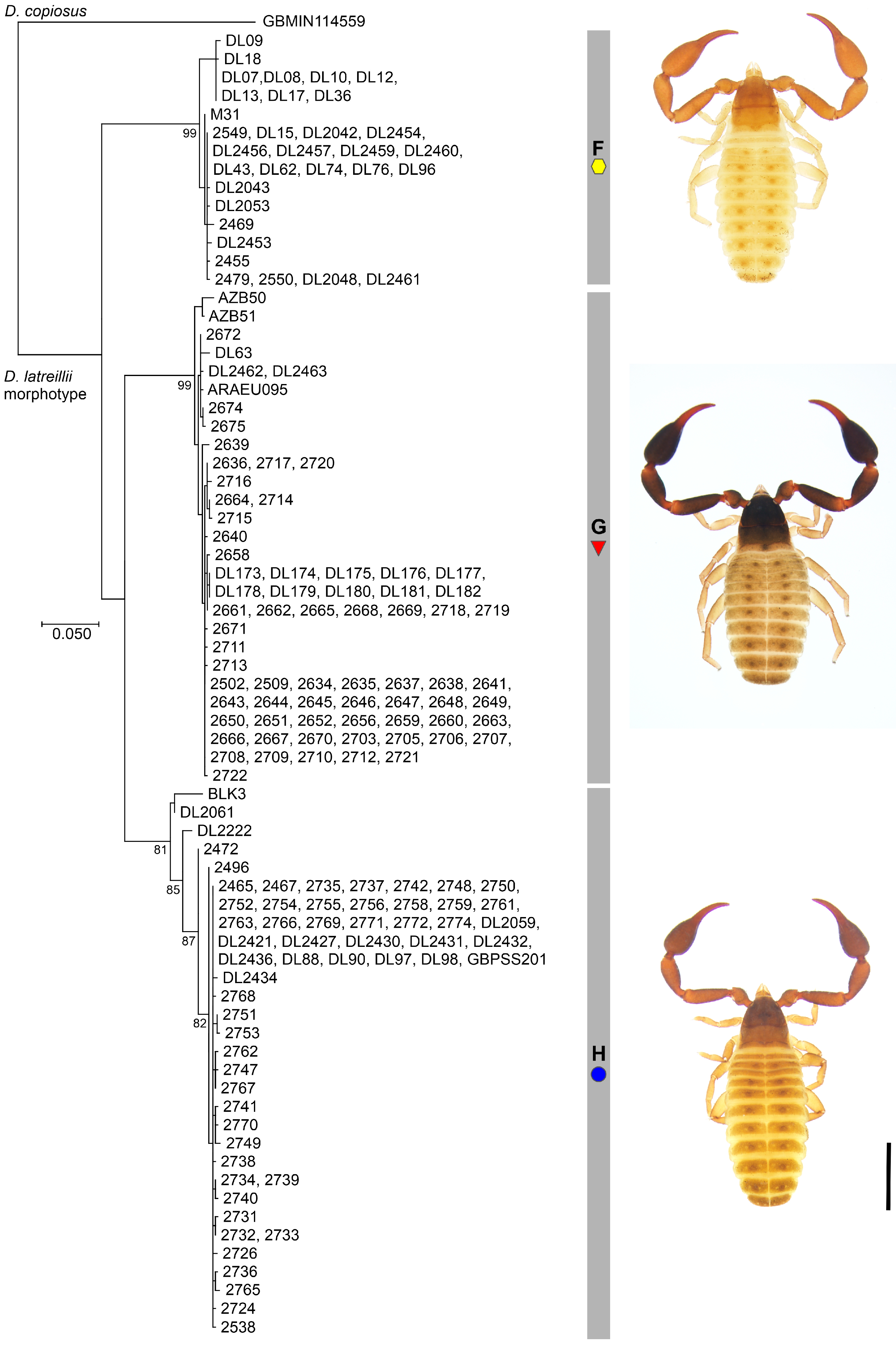
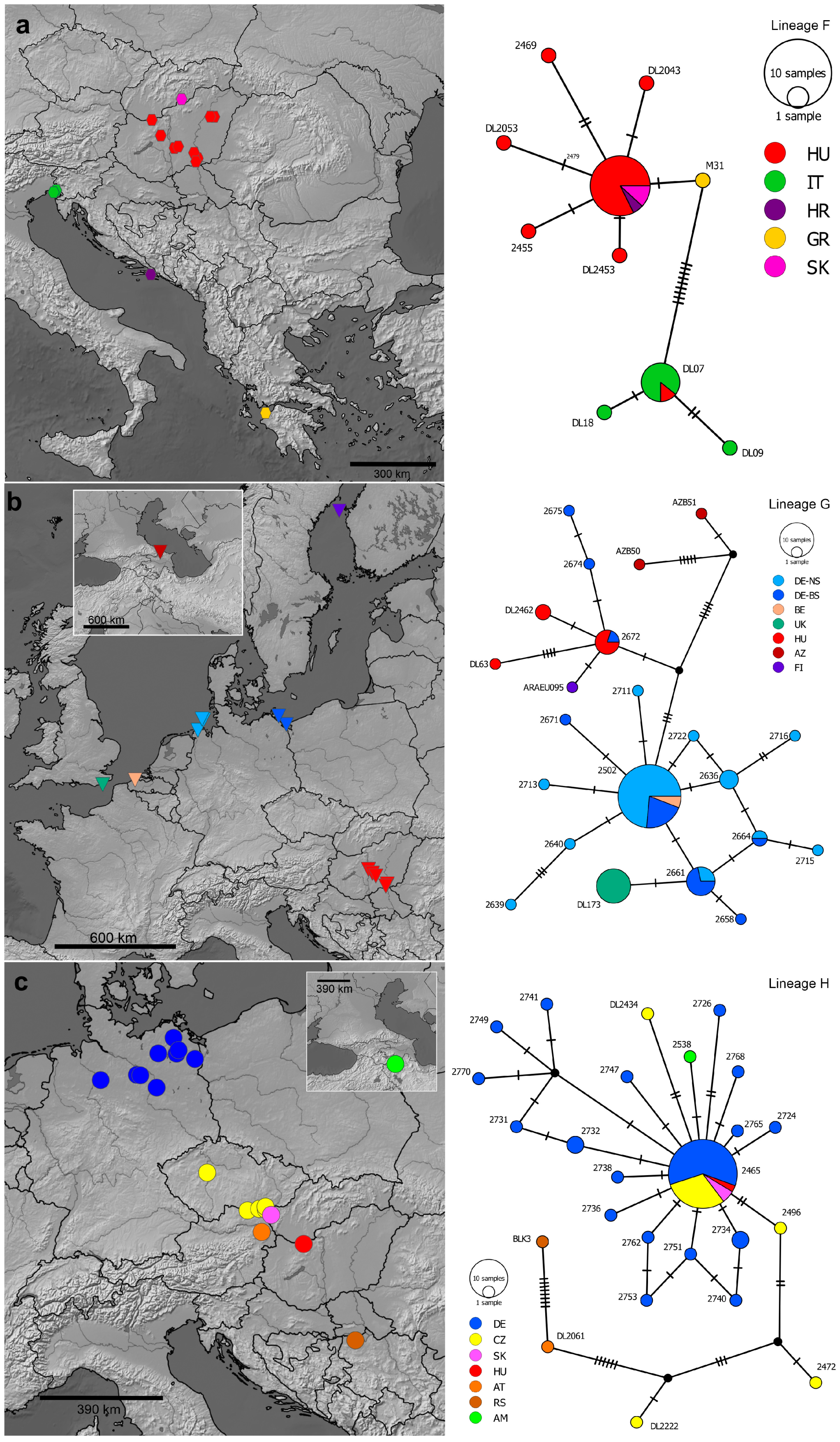
3.2. Molecular Phylogeny and Mitochondrial Divergence
3.3. Morphological Variation
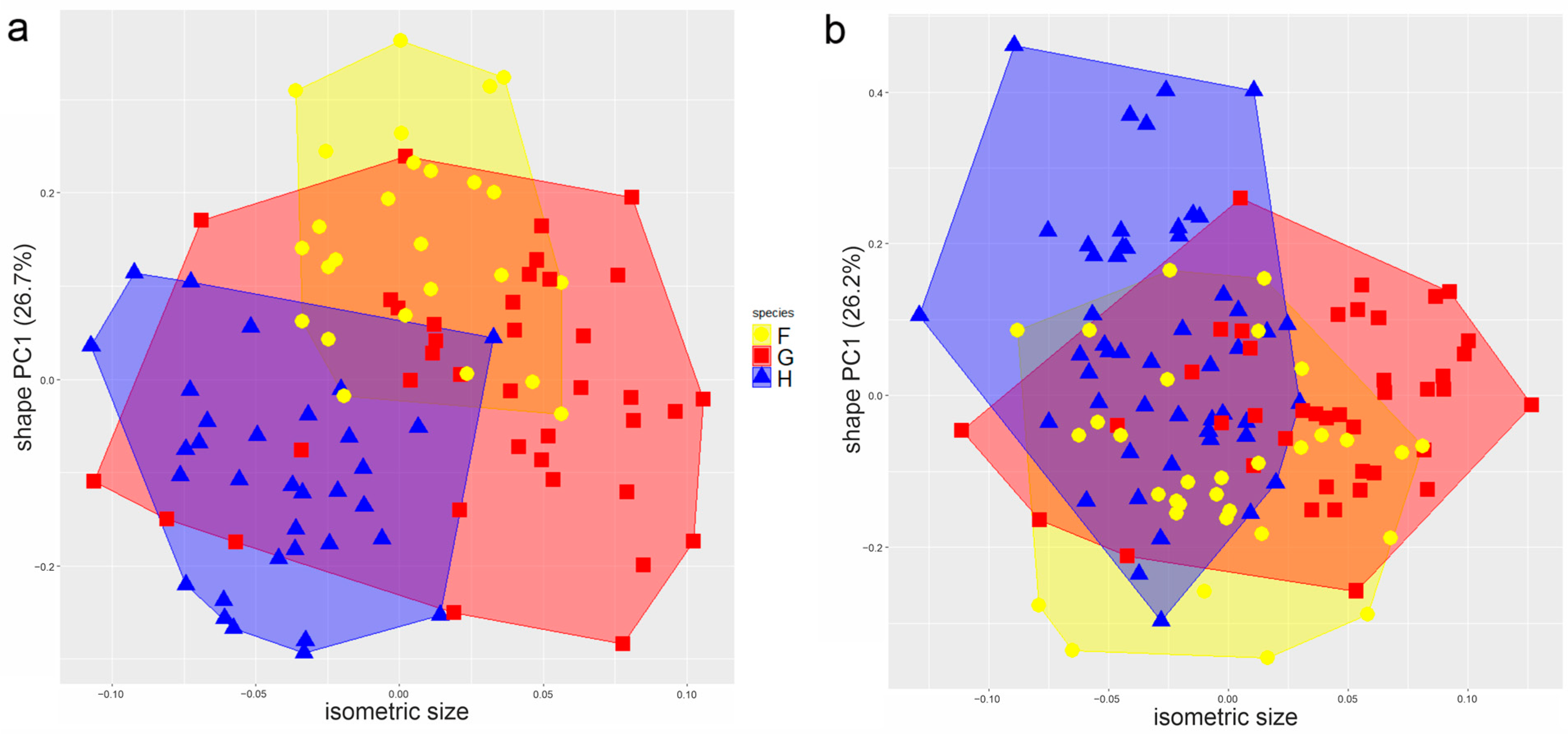
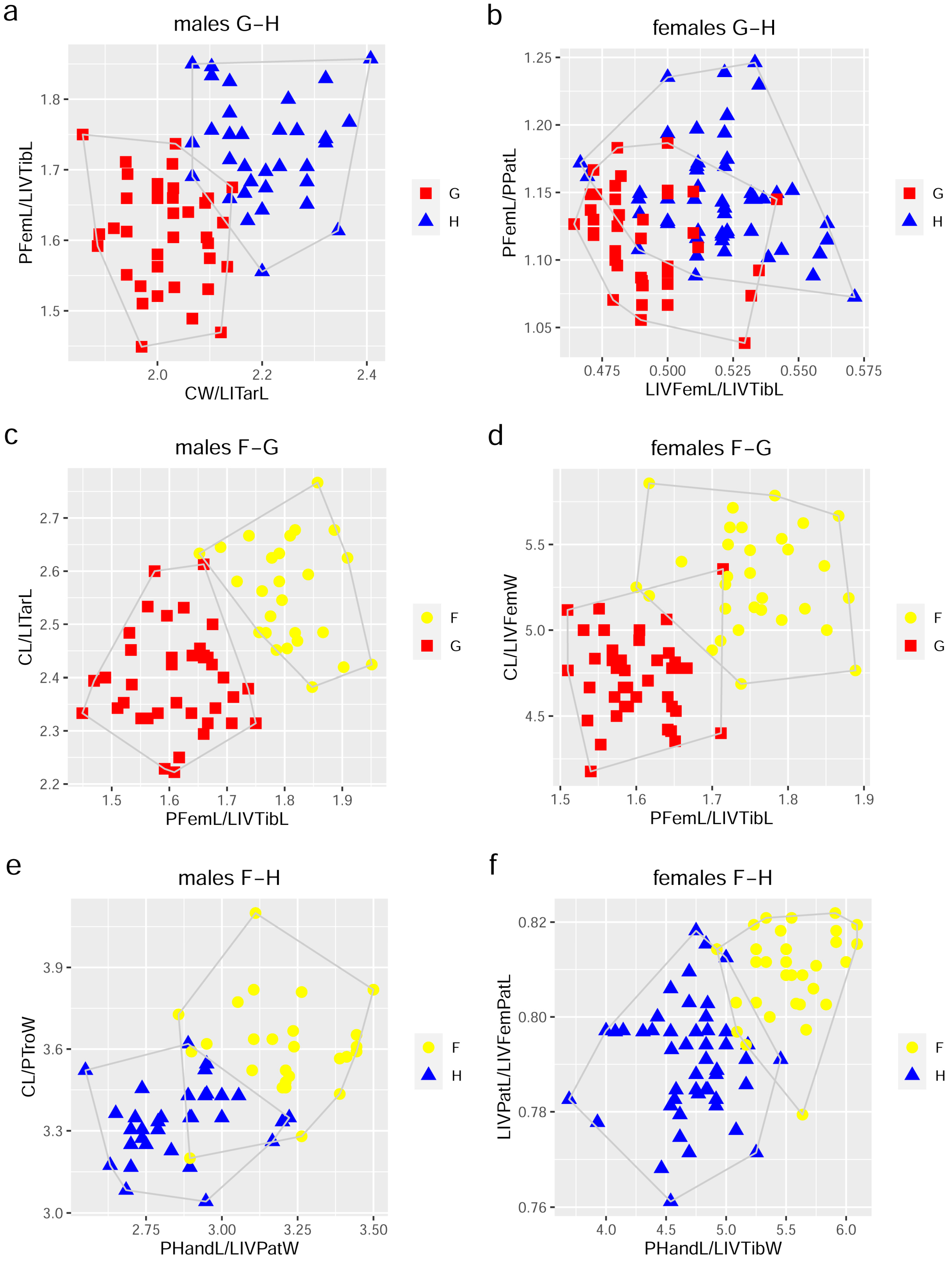
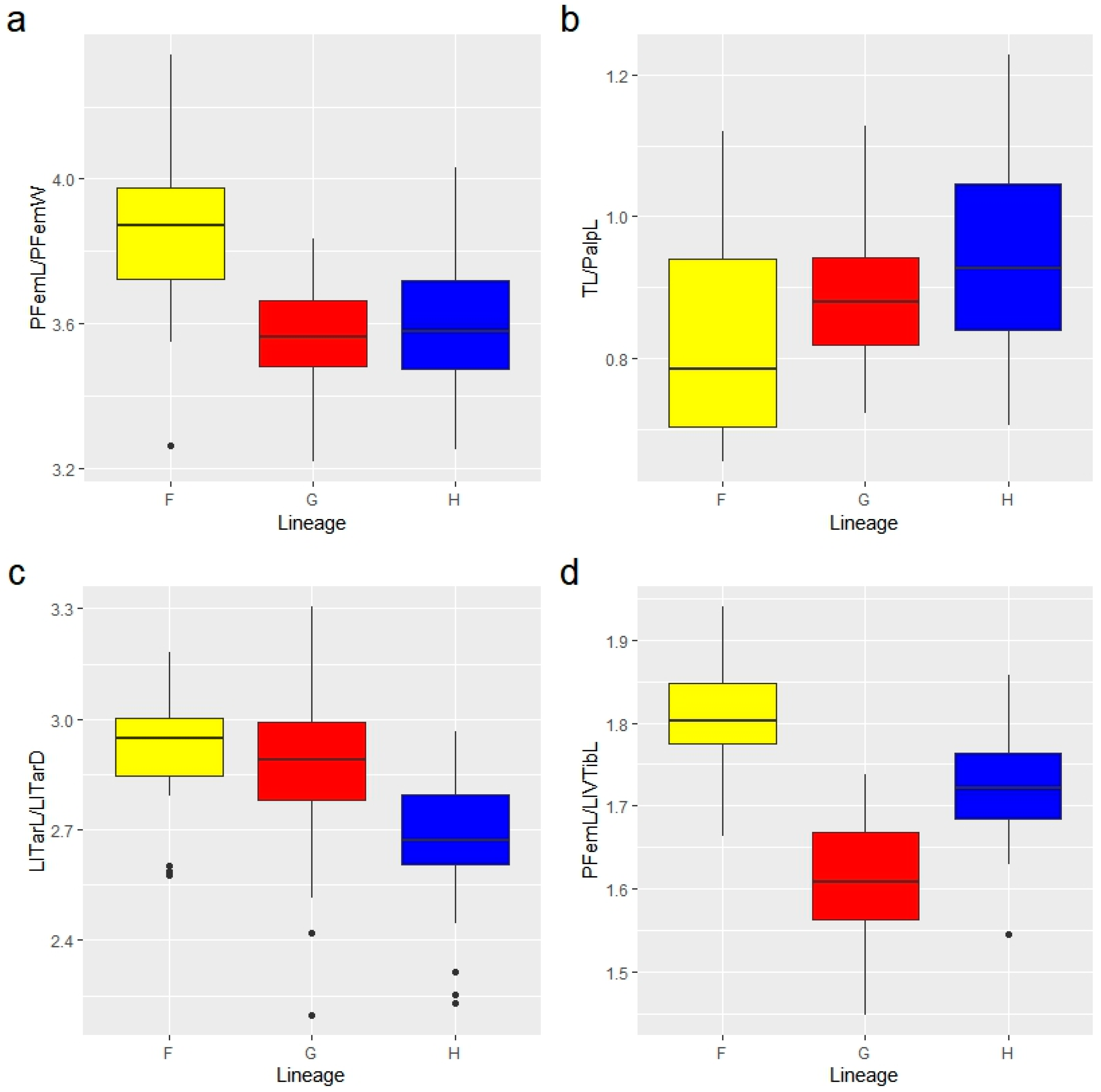
3.3.1. Multivariate Ratio Analysis
3.3.2. Shape Variation in Male Tarsus I
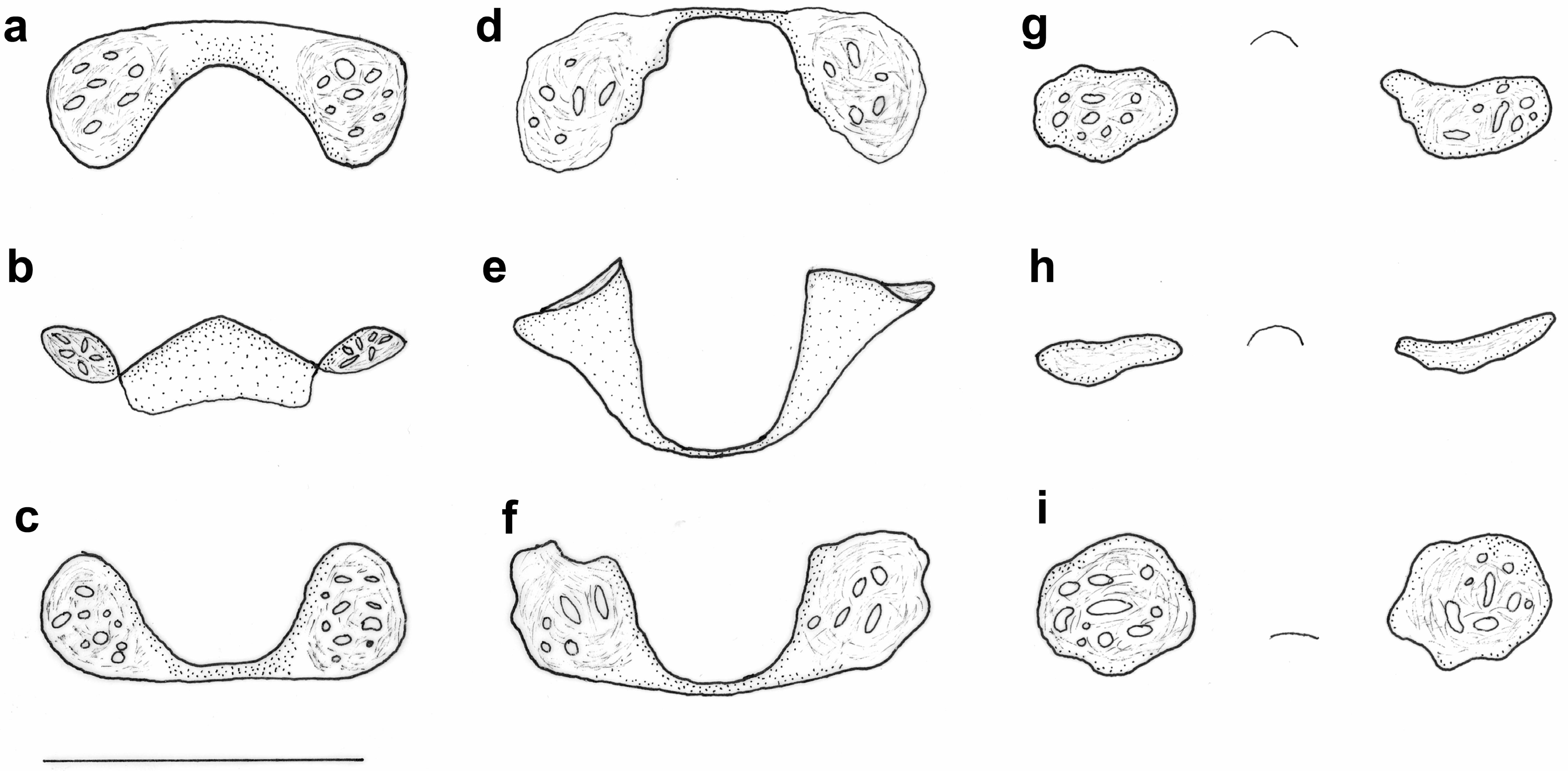
3.3.3. Genitalic Morphology
3.4. Taxonomy
- Cheliferidae Risso, 1827
- Dactylocheliferini Beier, 1932
- Dactylochelifer Beier, 1932
- Dactylochelifer latreillii species complex
- Dactylochelifer latreillii (Leach, 1817) (Figure S3)
- Chelifer latreillii Leach, 1817 (Holotype male from Britain, BMNH, dry collection of Leach “135a”, not examined).
- Dactylochelifer latreillii septentrionalis Beier, 1932 syn. nov. (Holotype male from Nordfriesische Insel: Norderney, September 1911, R. Heymans leg., ZMHB 31926, examined).
- Dactylochelifer degeerii (C.L. Koch, 1835) (Figure S4)
- Chelifer degeerii C.L. Koch, 1835 (type material from Regensburg, Bavaria, Germany, probably lost).
- Chelifer fabricii C.L. Koch, 1835 (type material from Regensburg, Bavaria, Germany, probably lost).
- Chelifer angustus C.L. Koch, 1836 (type material from Regensburg, Bavaria, Germany, probably lost).
- Chelifer schaefferi C.L. Koch, 1839 (type material from Bavaria, Germany, probably lost).
- Chelifer brevipalpis Canestrini, 1876 (type material from Bosco di Cervarese, Padova, Veneto, Italy, probably lost [77].
- Dactylochelifer ninnii (Canestrini, 1876) (Figure S5)
- Chelifer ninnii Canestrini, 1876 (type material from Valle Dogado, Laguna di Venezia, Italy, present in Canestrini collection at Zoology Museum of Padova University [77], image examined).
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agapow, P.M.; Bininda-Emonds, O.R.; Crandall, K.A.; Gittleman, J.L.; Mace, G.M.; Marshall, J.C.; Purvis, A. The impact of species concept on biodiversity studies. Q. Rev. Biol. 2004, 79, 161–179. [Google Scholar] [CrossRef]
- Mallet, J. Species, Concepts of. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Academic Press: London, UK; Washington, DC, USA, 2013; pp. 679–691. ISBN 9780123847201. [Google Scholar] [CrossRef]
- Aldhebiani, A.Y. Species concept and speciation. Saudi J. Biol. Sci. 2018, 25, 437–440. [Google Scholar] [CrossRef]
- Darwin, C. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life; John Murray: London, UK, 1859. [Google Scholar]
- Dobzhansky, T. Genetics and the Origin of Species; Columbia University Press: New York, NY, USA, 1937. [Google Scholar]
- Mayr, E. Systematics and the Origin of Species; Columbia University Press: New York, NY, USA, 1942. [Google Scholar]
- Sokal, R.R.; Crovello, T.J. The biological species concept: A critical evaluation. Am. Nat. 1970, 104, 127–153. [Google Scholar] [CrossRef]
- Zachos, F.E. Species Concepts in Biology; Springer: Cham, Switzerland, 2016; ISBN 978-3-319-44964-7. [Google Scholar] [CrossRef]
- Tautz, D.; Arctander, P.; Minelli, A.; Thomas, R.H.; Vogler, A.P. A plea for DNA taxonomy. Trends Ecol. Evol. 2003, 18, 70–74. [Google Scholar] [CrossRef]
- Padial, J.M.; Miralles, A.; De la Riva, I.; Vences, M. The integrative future of taxonomy. Front. Zool. 2010, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Schlick-Steiner, B.C.; Steiner, F.M.; Seifert, B.; Stauffer, C.; Christian, E.; Crozier, R.H. Integrative taxonomy: A multisource approach to exploring biodiversity. Ann. Rev. Entomol. 2010, 55, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Muster, C.; Michalik, P. Cryptic diversity in ant-mimic Micaria spiders (Araneae, Gnaphosidae) and a tribute to early naturalists. Zool. Scr. 2020, 49, 197–209. [Google Scholar] [CrossRef]
- Wilcox, T.P.; Hugg, L.; Zeh, J.A.; Zeh, D.W. Mitochondrial DNA sequencing reveals extreme genetic differentiation in a cryptic species complex of neotropical pseudoscorpions. Mol. Phyl. Evol. 1997, 7, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Fišer, C.; Robinson, C.T.; Malard, F. Cryptic species as a window into the paradigm shift of the species concept. Mol. Ecol. 2018, 27, 613–635. [Google Scholar] [CrossRef] [PubMed]
- Christophoryová, J.; Krajčovičová, K.; Šťáhlavský, F.; Španiel, S.; Opatova, V. Integrative taxonomy approach reveals cryptic diversity within the phoretic pseudoscorpion genus Lamprochernes (Pseudoscorpiones: Chernetidae). Insects 2023, 14, 122. [Google Scholar] [CrossRef]
- Ohira, H.; Kaneko, S.; Faulks, L.; Tsutsumi, T. Unexpected species diversity within Japanese Mundochthonius pseudoscorpions (Pseudoscorpiones: Chthoniidae) and the necessity for improved species diagnosis revealed by molecular and morphological examination. Invertebr. Syst. 2018, 32, 259–277. [Google Scholar] [CrossRef]
- Harvey, M.S. The smaller arachnid orders: Diversity, descriptions and distributions from Linnaeus to the present (1758 to 2007). Zootaxa 2007, 1668, 363–380. [Google Scholar] [CrossRef]
- Muster, C.; Spelda, J.; Rulik, B.; Thormann, J.; von der Mark, L.; Astrin, J.J. The dark side of pseudoscorpion diversity: The German Barcode of Life campaign reveals high levels of undocumented diversity in European false scorpions. Ecol. Evol. 2021, 11, 13815–13829. [Google Scholar] [CrossRef]
- Hlebec, D.; Harms, D.; Kučinić, M.; Harvey, M.S. Integrative taxonomy of the pseudoscorpion family Chernetidae (Pseudoscorpiones: Cheliferoidea): Evidence for new range-restricted species in the Dinaric Karst. Zool. J. Linn. Soc. 2023, zlad083, 1–26. [Google Scholar] [CrossRef]
- Hlebec, D.; Podnar, M.; Kučinić, M.; Harms, D. Molecular analyses of pseudoscorpions in a subterranean biodiversity hotspot reveal cryptic diversity and microendemism. Sci. Rep. 2023, 13, 430. [Google Scholar] [CrossRef]
- Mahnert, V. Neobisium carcinoides (Hermann, 1804) (Pseudoscorpionida, Neobisiidae)–une espèce polymorphe? Comptes Rendus du Xème Colloque Européen d’Arachnologie. Bull. Soc. Sci. Bretagne 1988, 59, 161–174. [Google Scholar]
- Poinar, G.O., Jr.; Ćurčić, B.P.; Cokendolpher, J.C. Arthropod phoresy involving pseudoscorpions in the past and present. Acta Arachnol. 1998, 47, 79–96. [Google Scholar] [CrossRef]
- Opatova, V.; Šťáhlavský, F. Phoretic or not? Phylogeography of the pseudoscorpion Chernes hahnii (Pseudoscorpiones: Chernetidae). J. Arachnol. 2018, 46, 104–113. [Google Scholar] [CrossRef]
- Harvey, M.S.; Lopes, P.C.; Goldsmith, G.R.; Halajian, A.; Hillyer, M.J.; Huey, J.A. A novel symbiotic relationship between sociable weaver birds (Philetairus socius) and a new cheliferid pseudoscorpion (Pseudoscorpiones: Cheliferidae) in southern Africa. Invertebr. Syst. 2015, 29, 444–456. [Google Scholar] [CrossRef]
- Just, P.; Šťáhlavský, F.; Bogusch, P.; Astapenková, A.; Opatova, V. Dispersal capabilities do not parallel ecology and cryptic speciation in European Cheliferidae pseudoscorpions (Pseudoscorpiones: Cheliferidae). Diversity 2023, 15, 1040. [Google Scholar] [CrossRef]
- World Pseudoscorpiones Catalog. Natural History Museum Bern. Available online: http://wac.nmbe.ch (accessed on 13 November 2023).
- Kew, H.W. On the pairing of Pseudoscorpiones. Proc. Zool. Soc. 1912, 82, 376–390. [Google Scholar] [CrossRef]
- Weygoldt, P. Vergleichende Untersuchungen zur Fortpflanzungsbiologie der Pseudoskorpione. Z. Morphol. Ökol. Tiere 1966, 56, 39–92. [Google Scholar] [CrossRef]
- Kirchmair, G.; Raspotnig, G. Mating behavior of Dactylochelifer latreillii latreillii (Pseudoscorpiones: Cheliferidae): A quantitative study. J. Arachnol. 2021, 49, 198–204. [Google Scholar] [CrossRef]
- Legg, G. The structure of encysted sperm of some British Pseudoscorpiones (Arachnida). J. Zool. 1973, 170, 429–440. [Google Scholar] [CrossRef]
- Weygoldt, P. Vergleichend-embryologische Untersuchungen an Pseudoscorpionen V. Das Embryonalstadium mit seinem Pumporgan bei verschiedenen Arten und sein Wert als taxonomisches Merkmal. Z. Zool. Syst. Evolutionforsch. 1971, 9, 3–29. [Google Scholar] [CrossRef]
- Vachon, M. Sur le développement postembryonnaire des Pseudoscorpions (quatrième note). Les formules chaetotaxiques des pattes-mâchoires. Bull. Mus. Natl. Hist. Nat. 1936, 8, 77–83. [Google Scholar]
- Gabbutt, P.D. Pseudoscorpions: Growth and trichobothria. Bull. Mus. Natl. Hist. Nat. 1970, 41 (Suppl. 1), 135–140. [Google Scholar]
- Gilbert, O. Observations on the feeding of some British false scorpions. Proc. Zool. Soc. 1951, 121, 547–555. [Google Scholar]
- Scheuring, L. Die Augen der Arachnoideen. Zool. Jahrb. Abt. Anat. Ontog. Tiere 1913, 33, 553–636. [Google Scholar]
- Gilbert, O. Studies of the histology of the mid-gut of the Chelonethi or Pseudoscorpiones. Q. J. Microsc. Sci. 1952, 93, 31–45. [Google Scholar] [CrossRef]
- Vachon, M. Recherches anatomiques et biologiques sur la réproduction et le développement des Pseudoscorpions. Ann. Sci. Nat. Zool. 1938, 1, 1–207. [Google Scholar]
- Gabbutt, P.D. The external morphology of the pseudoscorpion Dactylochelifer latreillei. J. Zool. 1970, 160, 313–335. [Google Scholar] [CrossRef]
- Simon, E. Les Ordres des Chernetes, Scorpiones et Opiliones. In Les Arachnides de France; Librairie Encyclopédique de Roret: Paris, France, 1879; Volume 7, pp. 1–332. [Google Scholar]
- Simon, E. Étude sur les Arachnides recueillis en Tunisie en 1883 et 1884 par Mm. A. Letourneux, M. Sédillot et Valery Mayet. In Exploration Scientifique de la Tunisie; Imprimerie Nationale: Paris, France, 1885; pp. 1–55. [Google Scholar]
- Pavesi, P. Materiali per lo studio della fauna Tunisina raccolti da G. e L. Doria. Ann. Mus. Civ. Stor. Nat. 1884, 20, 446–486. [Google Scholar]
- Ellingsen, E. Materiali per una fauna dell’Arcipelago Toscano. VIII. Isola del Giglio. Notes on pseudoscorpions. Ann. Mus. Civ. Stor. Nat. 1908, 3, 668–670. [Google Scholar]
- Ellingsen, E. Contributions to the knowledge of the pseudoscorpions from material belonging to the Museo Civico in Genova. Ann. Mus. Civ. Stor. Nat. 1909, 4, 205–220. [Google Scholar]
- Beier, M. Zur Kenntnis der Cheliferidae (Pseudoscorpionidea). Zool. Anz. 1932, 100, 53–67. [Google Scholar]
- Beier, M. Pseudoscorpionidea II. Subord. C. Cheliferinea. In Das Tierreich; Schulze, F.E., Kükenthal, W., Eds.; Walter de Gruyter & Co: Leipzig, Germany; Berlin, Germany, 1932; Volume 58, pp. i–xxi, 1–294. [Google Scholar]
- Mahnert, V. Über einige Atemnidae und Cheliferidae Griechenlands (Pseudoscorpiones). Mitt. Schweiz. Entomol. Ges. 1977, 50, 67–74. [Google Scholar]
- van den Tooren, D. The subspecies of the pseudoscorpion Dactylochelifer latreillei in the Netherlands (Pseudoscorpiones: Cheliferidae). Ned. Faun. Med. 2005, 23, 77–89. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit 1 from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Roslin, T.; Somervuo, P.; Pentinsaari, M.; Hebert, P.D.; Agda, J.; Ahlroth, P.; Anttonen, P.; Aspi, J.; Blagoev, G.; Blanco, S.; et al. A molecular-based identification resource for the arthropods of Finland. Mol. Ecol. Res. 2022, 22, 803–822. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Baur, H.; Kranz-Baltensperger, Y.; Cruaud, A.; Rasplus, J.-Y.; Timokhov, A.V.; Gokhman, V.E. Morphometric analysis and taxonomic revision of Anisopteromalus Ruschka (Hymenoptera: Chalcidoidea: Pteromalidae)–an integrative approach. Syst. Entomol. 2014, 39, 691–709. [Google Scholar] [CrossRef] [PubMed]
- Baur, H.; Leuenberger, C. Analysis of ratios in multivariate morphometry. Syst. Biol. 2011, 60, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Baur, H.; Leuenberger, C. Multivariate Ratio Analysis (MRA): R-Scripts and Tutorials for Calculating Shape PCA, Ratio Spectra and LDA Ratio Extractor (1.05). Zenodo. Available online: https://doi.org/10.5281/zenodo.4250142 (accessed on 30 November 2023).
- Shipunov, A.; Murrell, P.; D’Orazio, M.; Turner, S.; Altshuler, E.; Rau, R.; Beck, M.W.; Gibb, S.; Qiu, W.; Paradis, E.; et al. Package ‘shipunov’: Miscellaneous Functions from Alexey Shipunov, R package version 1.17.1. Available online: https://cran.r-project.org/web/packages/shipunov/index.html (accessed on 24 November 2023).
- Rohlf, F.J. TPSUTIL, Version 1.40; Department of Ecology and Evolution, State University of New York: Albany, NY, USA, 2008.
- Adams, D.; Collyer, M.; Kaliontzopoulou, A. Geomorph: Geometric Morphometric Analyses of 2D and 3D Landmark Data. Available online: https://cran.r-project.org/web/packages/geomorph/index.html (accessed on 10 November 2023).
- Rohlf, F.J.; Slice, D.E. Extensions of the Procrustes method for optimal superimposition of landmarks. Syst. Zool. 1990, 39, 40–59. [Google Scholar] [CrossRef]
- Adams, D.C.; Collyer, M.L. Multivariate comparative methods: Evaluations, comparisons, and recommendations. Syst. Biol. 2018, 67, 14–31. [Google Scholar] [CrossRef]
- Inkscape Project. Available online: https://inkscape.org (accessed on 24 November 2023).
- Kraus, O. Hoyer’s Gemisch statt Polyvinyl-Lactophenol. Mikrokosmos 1984, 2, 54–55. [Google Scholar]
- Mahnert, V. Zwei neue Dactylochelifer-Arten aus Spanien und von Mallorca (Pseudoscorpiones). Eos 1978, 52, 149–157. [Google Scholar]
- Hoff, C.C. The pseudoscorpions of Illinois. Bull. Ill. Nat. Hist. Surv. 1949, 24, 407–498. [Google Scholar] [CrossRef]
- Schawaller, W. Eine neue Dactylochelifer-Art aus Spanien (Prov. Tarragona). Eos 1987, 63, 277–280. [Google Scholar]
- Dashdamirov, S.; Schawaller, W. Pseudoscorpions from Middle Asia, Part 4 (Arachnida: Pseudoscorpiones). Stuttg. Beitr. Nat. kd. A Biol. 1995, 522, 1–24. [Google Scholar]
- Nassirkhani, M.; Snegovaya, N.; Chumachenko, Y.A. Description of a new pseudoscorpion species, Dactylochelifer zaragozai n. sp. (Pseudoscorpiones: Cheliferidae) from North Caucasian region of European Russia. Israel J. Ecol. Evol. 2022, 68, 13–20. [Google Scholar] [CrossRef]
- Mahnert, V. Sigles trichobothriaux chez les pseudoscorpions (Arachnida). Mem. Atti Soc. Tosc. Sci. Nat. Pisa B 1981, 88, 185–192. [Google Scholar]
- Kew, H.W. A synopsis of the false scorpions of Britain and Ireland. Proc. R. Ir. Acad. Sect. B 1911, 29, 38–64. [Google Scholar]
- Legg, G. British Pseudoscorpion Recording Scheme. Available online: https://www.chelifer.com/pseudos/pseudoscorpions.htm (accessed on 17 November 2023).
- Kaisila, J. A revision of the pseudoscorpion fauna of eastern Fennoscandia. Ann. Entomol. Fenn. 1949, 15, 72–92. [Google Scholar]
- Tullgren, A. Notiser rörande arter af Arachnidgrupperna Chelonethi och Phalangidea. Entomol. Tidskr. 1906, 27, 214–218. [Google Scholar]
- Gärdenfors, U.; Wilander, P. Sveriges klokrypare med nyckel till arterna. Entomol. Tidskr. 1992, 113, 20–35. [Google Scholar]
- Tóth, T. Salt-affected soils and their native vegetation in Hungary. In Sabkha Ecosystems: Volume III: Africa and Southern Europe; Öztürk, M., Böer, B., Barth, H.J., Clüsener-Godt, M., Khan, M.A., Breckle, S.-W., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 113–132. [Google Scholar]
- Pickard-Cambridge, O. On the British species of false-scorpions. Proc. Dorset Nat. Hist. Archaeol. Soc. 1892, 13, 199–231. [Google Scholar]
- Kew, H.W. An historical account of the pseudoscorpion-fauna of the British islands. J. Quekett Microsc. Club 1916, 13, 117–136. [Google Scholar]
- Judson, M.L.I. Catalogue of the pseudoscorpion types (Arachnida: Chelonethi) in the Natural History Museum, London. Occas. Pap. Syst. Entomol. 1997, 11, 1–54. [Google Scholar]
- Guariento, L.A.; Bonvicini, M.C.; Ballarin, L.; Devincenzo, U.; Gardini, G.; Moretto, E.; Pantini, P.; Nicolosi, P. Giovanni Canestrini’s heritage at the Zoology Museum of Padova University (Italy): A rediscovery of his arachnological collections and described species. Arachnol. Mitt. 2018, 55, 36–41. [Google Scholar] [CrossRef]
- Ressl, F.; Beier, M. Zur Ökologie, Biologie und Phänologie der heimischen Pseudoskorpione. Zool. Jahrb. Abt. Syst. Geogr. Biol. Tiere 1958, 86, 1–26. [Google Scholar]
- Beier, M. Ordnung Pseudoscorpionidea (Afterskorpione). Bestimmungsbücher zur Bodenfauna Europas; Akademie-Verlag: Berlin, Germany, 1963; pp. 1–313. [Google Scholar]
- Drogla, R.; Lippold, K. Zur Kenntnis der Pseudoskorpion-Fauna von Ostdeutschland (Arachnida, Pseudoscorpiones). Arachnol. Mitt. 2004, 27–28, 1–54. [Google Scholar] [CrossRef]
- Krajčovičová, K.; Christophoryova, J.; Lučeničová, T. Pseudoscorpions (Arachnida: Pseudoscorpiones) found in bird nests and in bat guano in Slovakia and Germany. Munis Entomol. Zool. 2015, 10, 428–434. [Google Scholar]
- Christophoryová, J.; Jajcayová, D.; Krajčovičová, K. Pseudoscorpions (Arachnida: Pseudoscorpiones) living in tree microhabitats in Slovakia. Klapalekiana 2017, 53, 283–297. [Google Scholar]
- Koch, C.L. Die Arachniden. Getreu Nach der Natur Abgebildet und Beschrieben, Band 10; C.H. Zeh’sche Buchhandlung: Nürnberg, Germany, 1843; pp. 1–142. [Google Scholar]
- Christophoryová, J.; Krumpálova, Z.; Krištofík, J.; Országhová, Z. Association of pseudoscorpions with different types of bird nests. Biologia 2011, 66, 669–677. [Google Scholar] [CrossRef]
- Turienzo, P.; di Iorio, O.; Mahnert, V. Global checklist of pseudoscorpions (Arachnida) found in birds’ nests. Rev. Suisse Zool. 2010, 117, 557–598. [Google Scholar]
- Christophoryová, J.; Kaňuchová, A.; Krajčovičová, K. Faunistic survey of pseudoscorpions (Arachnida: Pseudoscorpiones) collected from compost heaps in Slovakia. Klapalekiana 2017, 53, 11–19. [Google Scholar]
- Nagel, P.; Grieder, L. Herrich-Schäffer’s Deutschlands Insecten … (1829–1844), the associate publication of Koch’s Deutschlands Crustaceen … (1835–1844), and Heft 110 (1823) of Panzer’s Faunae Insectorum Germanicae …: Bibliographic notes and dating. Sherbornia 2019, 5, 1–156. [Google Scholar]
- Koch, C.L. Deutschlands Crustaceen, Myriapoden und Arachniden, 2; Friedrich Pustet: Regensburg, Germany, 1835. [Google Scholar]
- Koch, C.L. Deutschlands Crustaceen, Myriapoden und Arachniden, 7; Friedrich Pustet: Regensburg, Germany, 1836. [Google Scholar]
- Koch, C.L. Übersicht des Arachnidensystems, 2; C.H. Zeh’sche Buchhandlung: Nürnberg, Germany, 1839. [Google Scholar]
- Canestrini, G. Osservazioni aracnologiche. Atti Soc. Veneto-Trentina Sci. Nat. 1876, 3, 206–232. [Google Scholar]
- Dítě, Z.; Šuvada, R.; Tóth, T.; Jun, P.E.; Píš, V.; Dítě, D. Current condition of Pannonic salt steppes at their distribution limit: What do indicator species reveal about habitat quality? Plants 2021, 10, 530. [Google Scholar] [CrossRef] [PubMed]
- Canestrini, J. Chernetides Italici. In Acari, Myriapoda et Scorpiones hucusque in Italia reperta A. Berlese; Berlese, A., Ed.; Biodiversity Heritage Library: Padova, Italy, 1883; fascicolo VII. [Google Scholar]
- Lucas, H. Histoire Naturelle des Animaux Articulés. Crustacés, Arachnides, Myriapodes et Hexapodes. Exploration Scientifique de l’Algérie Pendant les Années 1840, 1841, 1842. Sciences Physiques. Zoologie; Imprimerie Nationale: Paris, France, 1849; Volume 2. [Google Scholar]
- Beier, M. Due nuovi pseudoscorpioni della Tunisia. Boll. Del Lab. Di Zool. Gen. E Agrar. Del R. Ist. Super. Agrar. Di Portici 1930, 24, 95–98. [Google Scholar]
- Beier, M. Pseudoscorpione aus Marocco nebst einer Art von der Insel Senafir. Bull. Soc. Sci. Nat. Maroc. 1930, 10, 70–77. [Google Scholar]
- Callaini, G. Su alcune specie di Cheliferidae della regione Mediterranea (Arachnida, Pseudoscorpionida). (Notulae Chernetologicae. XXII). Boll. Mus. Civ. Stor. Nat. 1987, 13, 273–294. [Google Scholar]
- Lazzeroni, G. Sur la faune de pseudoscorpions de la région apenninique méridionale (Recherches sur les Pseudoscorpions. III). Mem. Mus. Civ. Stor. Nat. 1969, 16, 321–344. [Google Scholar]
- Beier, M. Zoologische Forschungsreise nach den Jonischen Inseln und dem Peloponnes. I. und II. Teil. Sitzungsber. Akad. Wiss. Wien 1929, 128, 425–456. [Google Scholar]
- Weygoldt, P. Vergleichende Untersuchungen zur Fortpflanzungsbiologie der Pseudoscorpione II. Z. Zool. Syst. Evolutionforsch. 1970, 8, 241–259. [Google Scholar] [CrossRef]
- Weygoldt, P. The Biology of Pseudoscorpions; Harvard University Press: Cambridge, MA, USA, 1969; 145p. [Google Scholar]
- Bickford, D.; Lohman, D.J.; Sodhi, N.S.; Ng, P.K.; Meier, R.; Winker, K.; Ingram, K.K.; Das, I. Cryptic species as a window on diversity and conservation. Trends Ecol. Evol. 2007, 22, 148–155. [Google Scholar] [CrossRef]
- Klausen, F.E. The male genitalia of the family Atemnidae (Pseudoscorpiones). J. Arachnol. 2005, 33, 641–662. [Google Scholar] [CrossRef]
- Romero-Ortiz, C.; Sarmiento, C.E. A comparative study of the male genitalia of the Cacodemoniini (Pseudoscorpiones: Withiidae). J. Arachnol. 2021, 49, 108–121. [Google Scholar] [CrossRef]
- Legg, G. The genitalia and associated glands of five British species belonging to the family Chthoniidae (Pseudoscorpiones: Arachnida). J. Zool. 1975, 177, 99–121. [Google Scholar] [CrossRef]
- Legg, G. The genitalia and associated glands of five British species belonging to the family Neobisiidae (Pseudoscorpiones: Arachnida). J. Zool. 1975, 177, 123–151. [Google Scholar] [CrossRef]
- Heurtault, J. Pseudoscorpions de la région du Tibesti (Sahara méridionale). IV. Cheliferidae. Bull. Mus. Natl. Hist. Nat. 1971, 42, 685–707. [Google Scholar]
- Gao, Z.-Z.; Zhang, F. A new species of the genus Dactylochelifer Beier from Shanxi, China (Pseudoscorpiones: Cheliferidae). ISRN Zool. 2012, 2012, 583830. [Google Scholar] [CrossRef]
- Heneberg, P.; Bogusch, P.; Astapenková, A. Reed galls serve as an underestimated but critically important resource for an assemblage of aculeate hymenopterans. Biol. Conserv. 2014, 172, 146–154. [Google Scholar] [CrossRef]
- Bogusch, P.; Macek, J.; Janšta, P.; Kubík, Š.; Řezáč, M.; Holý, K.; Malenovský, I.; Baňař, P.; Mikát, M.; Astapenková, A.; et al. Industrial and post-industrial habitats serve as critical refugia for pioneer species of newly identified arthropod assemblages associated with reed galls. Biodivers. Conserv. 2016, 25, 827–863. [Google Scholar] [CrossRef]
- Atlas der Spinnentiere Europas. Arachnologische Gesellschaft. Available online: https://atlas.arages.de/ (accessed on 25 November 2023).
- Muster, C.; Blick, T. Rote Liste und Gesamtartenliste der Pseudoskorpione (Arachnida: Pseudoscorpiones) Deutschlands. Natursch. Biol. Vielfalt 2016, 70, 539–561. [Google Scholar]
- Schawaller, W. Pseudoskorpione aus der Sowjetunion, Teil 2 (Arachnida: Pseudoscorpiones). Stuttg. Beitr. Nat. kd. A Biol. 1986, 396, 1–15. [Google Scholar]
- Beier, M. Nochmals über iberische und marokkanische Pseudoscorpione. Eos 1961, 37, 21–39. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muster, C.; Korba, J.; Bogusch, P.; Heneberg, P.; Šťáhlavský, F. And Yet They Differ: Reconsiderations of Diversity within Dactylochelifer latreillii (Arachnida: Pseudoscorpiones). Diversity 2024, 16, 137. https://doi.org/10.3390/d16030137
Muster C, Korba J, Bogusch P, Heneberg P, Šťáhlavský F. And Yet They Differ: Reconsiderations of Diversity within Dactylochelifer latreillii (Arachnida: Pseudoscorpiones). Diversity. 2024; 16(3):137. https://doi.org/10.3390/d16030137
Chicago/Turabian StyleMuster, Christoph, Jan Korba, Petr Bogusch, Petr Heneberg, and František Šťáhlavský. 2024. "And Yet They Differ: Reconsiderations of Diversity within Dactylochelifer latreillii (Arachnida: Pseudoscorpiones)" Diversity 16, no. 3: 137. https://doi.org/10.3390/d16030137
APA StyleMuster, C., Korba, J., Bogusch, P., Heneberg, P., & Šťáhlavský, F. (2024). And Yet They Differ: Reconsiderations of Diversity within Dactylochelifer latreillii (Arachnida: Pseudoscorpiones). Diversity, 16(3), 137. https://doi.org/10.3390/d16030137