

Defining Evolutionary Conservation Units in the Macedonian Crested Newt, Triturus macedonicus (Amphibia; Salamandridae), in a Biodiversity Hotspot

 ,
, 
 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Laboratory Procedures
2.2. Mitochondrial DNA Analysis
2.2.1. Phylogeography
2.2.2. Historical Demography
2.2.3. Genetic Diversity
2.3. Microsatellite Analysis
2.3.1. Population Structure and Gene Flow
2.3.2. Genetic Diversity, Differentiation, and Isolation by Distance
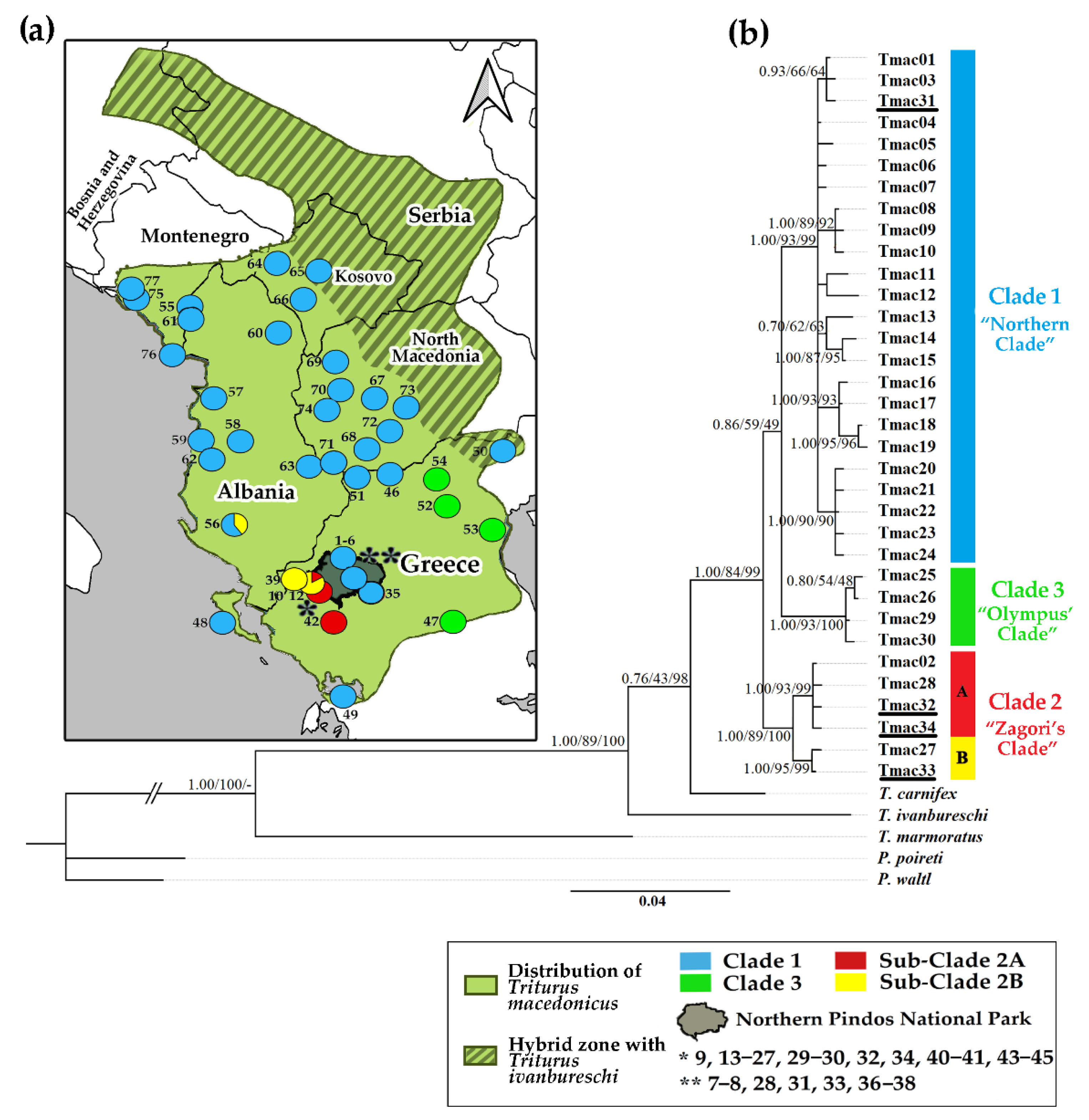
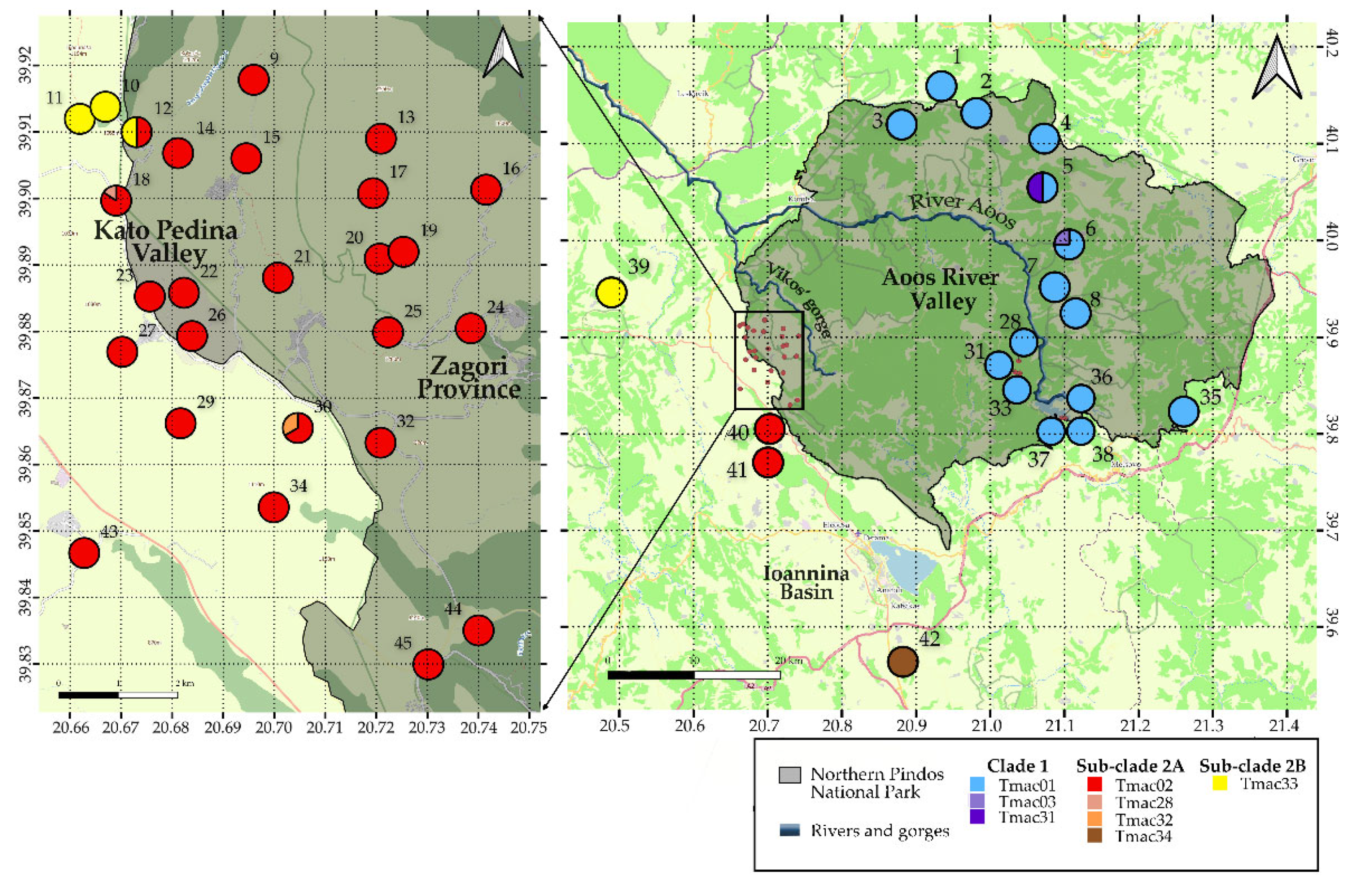
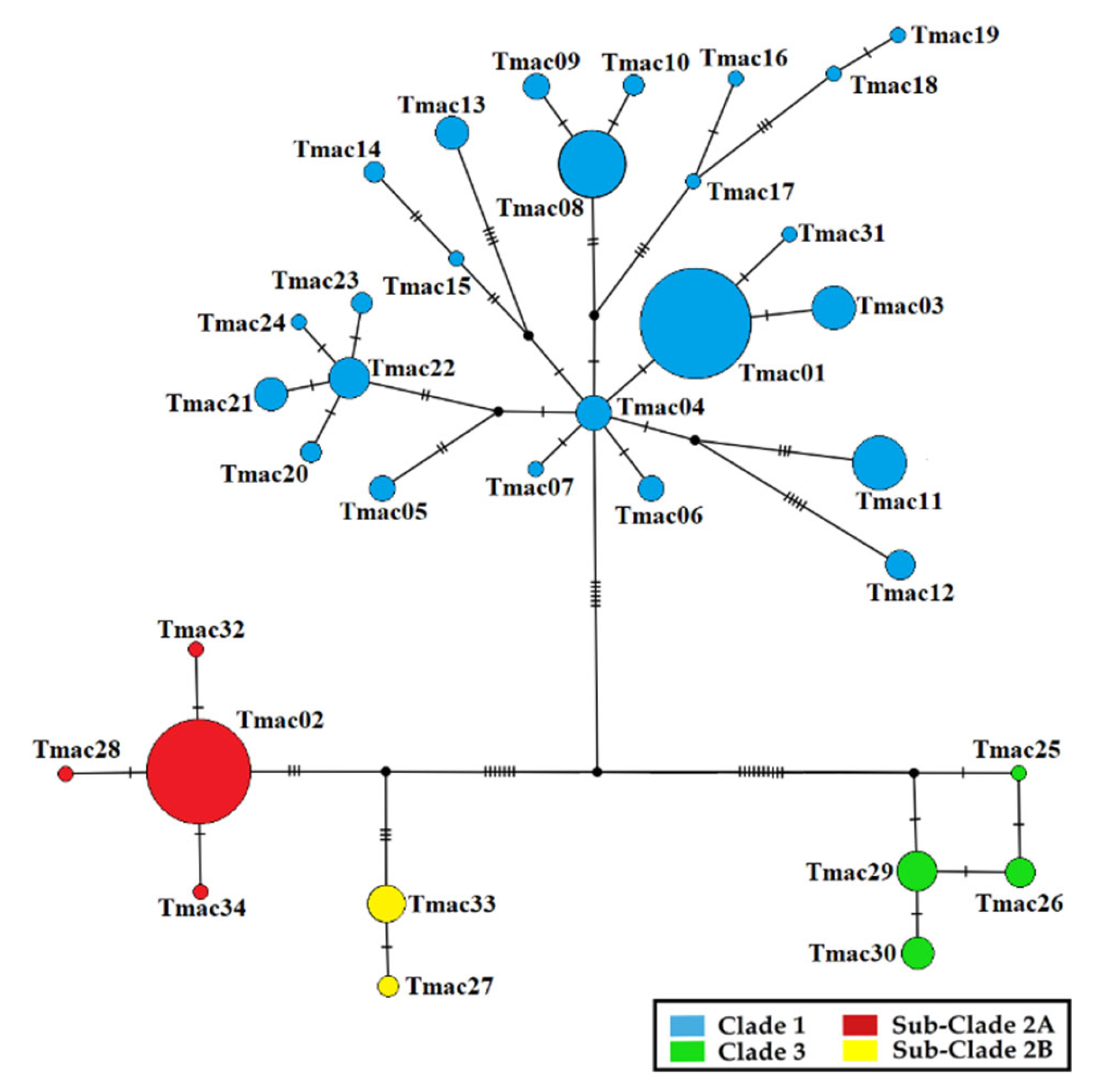
3. Results
3.1. Mitochondrial DNA
3.1.1. Phylogeography and Historical Demography
3.1.2. Genetic Diversity
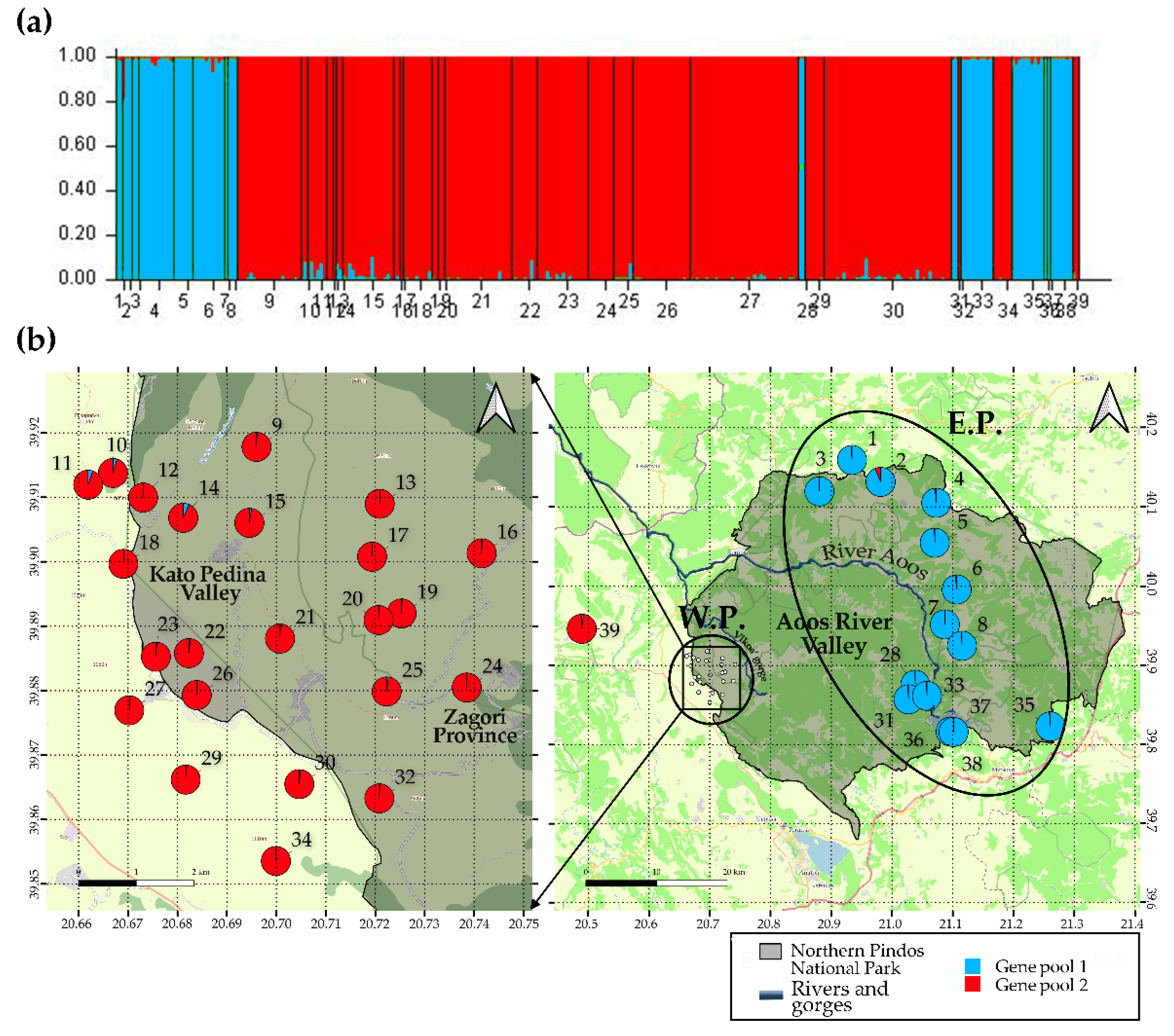
3.2. Microsatellite Analysis
3.2.1. Population Structure and Gene Flow
3.2.2. Genetic Diversity and Isolation by Distance
4. Discussion
4.1. Identification and Characterization of ESUs
4.2. Genetic Variation and Population Connectivity
4.3. Conservation Status and Management Priorities
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. Introduction to Conservation Genetics, 2nd ed.; Cambridge University Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Byrne, M. Phylogenetics and the Conservation of a Diverse and Ancient Flora. Comptes Rendus Biol. 2003, 326, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Canestrelli, D.; Bisconti, R.; Sacco, F.; Nascetti, G. What Triggers the Rising of an Intraspecific Biodiversity Hotspot? Hints from the Agile Frog. Sci. Rep. 2014, 4, 5042. [Google Scholar] [CrossRef] [PubMed]
- Ryder, O.A. Species Conservation and Systematics: The Dilemma of Subspecies. Trends Ecol. Evol. 1986, 1, 9–10. [Google Scholar] [CrossRef]
- Waples, R.S. Pacific Salmon, Oncorhynchus Spp. & the Definition of ‘Species’ under the Endangered Species Act. Mar. Fish. Rev. 1991, 53, 11–12. [Google Scholar]
- Moritz, C. Defining ‘Evolutionarily Significant Units’ for Conservation. Trends Ecol. Evol. 1994, 9, 373–375. [Google Scholar] [CrossRef] [PubMed]
- Houlahan, J.E.; Findlay, C.S.; Schmidt, B.R.; Meyer, A.H.; Kuzmin, S.L. Quantitative Evidence for Global Amphibian Population Declines. Nature 2000, 404, 752–755. [Google Scholar] [CrossRef] [PubMed]
- Wake, D.B.; Vredenburg, V.T. Are We in the Midst of the Sixth Mass Extinction? A View from the World of Amphibians. Proc. Natl. Acad. Sci. USA 2008, 105, 11466–11473. [Google Scholar] [CrossRef] [PubMed]
- IUCN. The IUCN Red List of Threatened Species. Version 2022-2; IUCN: Gland, Switzerland, 2023. [Google Scholar]
- Erwin, K.L. Wetlands and Global Climate Change: The Role of Wetland Restoration in a Changing World. Wetl. Ecol. Manag. 2009, 17, 71–84. [Google Scholar] [CrossRef]
- Cayuela, H.; Arsovski, D.; Thirion, J.-M.; Bonnaire, E.; Pichenot, J.; Boitaud, S.; Miaud, C.; Joly, P.; Besnard, A. Demographic Responses to Weather Fluctuations Are Context Dependent in a Long-Lived Amphibian. Glob. Chang. Biol. 2016, 22, 2676–2687. [Google Scholar] [CrossRef]
- Weinbach, A.; Cayuela, H.; Grolet, O.; Besnard, A.; Joly, P. Resilience to Climate Variation in a Spatially Structured Amphibian Population. Sci. Rep. 2018, 8, 14607. [Google Scholar] [CrossRef]
- Blondel, J.; Aronson, J. Biology and Wildlife of the Mediterranean Region; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Préau, C.; Bertrand, R.; Sellier, Y.; Grandjean, F.; Isselin-Nondedeu, F. Climate Change Would Prevail over Land Use Change in Shaping the Future Distribution of Triturus marmoratus in France. Anim. Conserv. 2022, 25, 221–232. [Google Scholar] [CrossRef]
- Bowler, D.E.; Benton, T.G. Causes and Consequences of Animal Dispersal Strategies: Relating Individual Behaviour to Spatial Dynamics. Biol. Rev. 2005, 80, 205–225. [Google Scholar] [CrossRef] [PubMed]
- Ronce, O. How Does It Feel to Be Like a Rolling Stone? Ten Questions About Dispersal Evolution. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 231–253. [Google Scholar] [CrossRef]
- Tromeur, E.; Rudolf, L.; Gross, T. Impact of Dispersal on the Stability of Metapopulations. J. Theor. Biol. 2016, 392, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hanski, I.; Gaggiotti, O. Metapopulation Biology. In Ecology, Genetics and Evolution of Metapopulations; Elsevier: Amsterdam, The Netherlands, 2004; pp. 3–22. [Google Scholar]
- Wright, S. Evolution and the Genetics of Populations. Vol. 4. Variability within and among Natural Populations; University of Chicago Press: Chicago, IL, USA, 1978. [Google Scholar]
- Lande, R. Risk of Population Extinction from Fixation of New Deleterious Mutations. Evolution 1994, 48, 1460–1469. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M.; Conery, J.; Bürger, R. Mutational Meltdowns In Sexual Populations. Evolution 1995, 49, 1067–1080. [Google Scholar] [CrossRef]
- Slatkin, M. Gene Flow and Genetic Drift in a Species Subject to Frequent Local Extinctions. Theor. Popul. Biol. 1977, 12, 253–262. [Google Scholar] [CrossRef]
- Semlitsch, R.D. Differentiating Migration and Dispersal Processes for Pond-Breeding Amphibians. J. Wildl. Manag. 2008, 72, 260–267. [Google Scholar] [CrossRef]
- Griffiths, I.; Kryštufek, B.; Reed, J. Balkan Biodiversity; Springer Netherlands: Dordrecht, The Netherlands, 2004. [Google Scholar]
- Sotiropoulos, K.; Danelis, T.; Theodoropoulos, A.; Toli, E.-A.; Bounas, A. Investigation of the Distribution and Conservation Status of the Priority Species of the 92/43 Macedonian Crested Newt (Triturus macedonicus) and Its Habitats, in the Protected Area of the Northern Pindos National Park. Deliverable B1: Project Final Report; University of Ioannina: Ioannina, Greece, 2022. (In Greek) [Google Scholar]
- Valakos, E.; Pafilis, P.; Sotiropoulos, K.; Lymberakis, P.; Maragou, P.; Foufopoulos, J. The Amphibians and Reptiles of Greece; Chimaira Editions: Frankfurt am Main, Germany, 2008. [Google Scholar]
- Wielstra, B.; Crnobrnja-Isailović, J.; Litvinchuk, S.N.; Reijnen, B.T.; Skidmore, A.K.; Sotiropoulos, K.; Toxopeus, A.G.; Tzankov, N.; Vukov, T.; Arntzen, J.W. Tracing Glacial Refugia of Triturus Newts Based on Mitochondrial DNA Phylogeography and Species Distribution Modeling. Front. Zool. Vol. 2013, 10, 13. [Google Scholar] [CrossRef]
- Wielstra, B.; Arntzen, J.W. Postglacial Species Displacement in Triturus Newts Deduced from Asymmetrically Introgressed Mitochondrial DNA and Ecological Niche Models. BMC Evol. Biol. 2012, 12, 161. [Google Scholar] [CrossRef]
- Wielstra, B.; Arntzen, J.W. Exploring the Effect of Asymmetric Mitochondrial DNA Introgression on Estimating Niche Divergence in Morphologically Cryptic Species. PLoS ONE 2014, 9, e95504. [Google Scholar] [CrossRef] [PubMed]
- Wielstra, B.; Arntzen, J.W. Unraveling the Rapid Radiation of Crested Newts (Triturus cristatus Superspecies) Using Complete Mitogenomic Sequences. BMC Evol. Biol. 2011, 11, 162. [Google Scholar] [CrossRef]
- Wielstra, B.; Arntzen, J.W. Extensive Cytonuclear Discordance in a Crested Newt from the Balkan Peninsula Glacial Refugium. Biol. J. Linn. Soc. 2020, 130, 578–585. [Google Scholar] [CrossRef]
- Edgar, P.; Bird, D.R. Action Plan for the Conservation of the Crested Newt Triturus cristatus Species Complex in Europe. In Proceedings of the Council of the European Union; Council of Europe: Strasbourg, France, 2006. [Google Scholar]
- IUCN SSC Amphibian Specialist Group. Triturus macedonicus. The IUCN Red List of Threatened Species 2022; IUCN: Gland, Switzerland, 2022. [Google Scholar]
- Arntzen, J.W.; Espregueira Themudo, G.; Wielstra, B. The Phylogeny of Crested Newts (Triturus cristatus Superspecies): Nuclear and Mitochondrial Genetic Characters Suggest a Hard Polytomy, in Line with the Paleogeography of the Centre of Origin. Contrib. Zool. 2007, 76, 261–278. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Wielstra, B.; Canestrelli, D. Next-Generation Phylogeography of the Italian Crested Newt Triturus carnifex Confirms Allopatric Glacial Refugia Positioned in Two Mediterranean Peninsulas; GenBank: KT364610 Direct Submission; Terrestrial Zoology, Naturalis Biodiversity Center, Darwinweg 2: Leiden, The Netherlands, 2015. [Google Scholar]
- Wielstra, B.; Burke, T.; Butlin, R.K.; Avcı, A.; Üzüm, N.; Bozkurt, E.; Olgun, K.; Arntzen, J.W. A Genomic Footprint Confirms Hybrid Zone Movement in Crested Newts. Evol. Lett. 2017, 1, 93–101. [Google Scholar] [CrossRef]
- Carvalho, S.B.; Velo-Antón, G.; Tarroso, P.; Portela, A.P.; Barata, M.; Carranza, S.; Moritz, C.; Possingham, H.P. Spatial Conservation Prioritization of Biodiversity Spanning the Evolutionary Continuum. Nat. Ecol. Evol. 2017, 1, 0151. [Google Scholar] [CrossRef]
- Gutiérrez-Rodríguez, J.; Barbosa, A.M.; Martínez-Solano, Í. Integrative Inference of Population History in the Ibero-Maghrebian Endemic Pleurodeles waltl (Salamandridae). Mol. Phylogenet Evol. 2017, 112, 122–137. [Google Scholar] [CrossRef]
- Escoriza, D.; Gutiérrez-Rodríguez, J.; Ben Hassine, J.; Martínez-Solano, I. Genetic Assessment of the Threatened Microendemic Pleurodeles poireti (Caudata, Salamandridae), with Molecular Evidence for Hybridization with Pleurodeles nebulosus. Conserv. Genet. 2016, 17, 1445–1458. [Google Scholar] [CrossRef]
- Sotiropoulos, K.; Tsaparis, D.; Eleftherakos, K.; Kotoulas, G.; Legakis, A.; Kasapidis, P. New Polymorphic Microsatellite Loci for the Macedonian Crested Newt, Triturus macedonicus, and Cross-Priming Testing in Four Other Crested Newt Species. Mol. Ecol. Resour. 2008, 8, 1402–1404. [Google Scholar] [CrossRef] [PubMed]
- Toonen, R.J.; Hughes, S.S. Increased Throughput for Fragment Analysis on an ABI PRISM 377 Automated Sequencer Using a Membrane Comb and STRand Software. Biotechniques 2001, 31, 1320–1324. [Google Scholar] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sanchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sanchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Evolutionary Trees from DNA Sequences: A Maximum Likelihood Approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F.; Nielsen, R.; Bollback, J.P. Bayesian Inference of Phylogeny and Its Impact on Evolutionary Biology. Science 2001, 294, 2310–2314. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Tavaré, S. Some Probabilistic and Statistical Problems in the Analysis of DNA Sequences. In Some Mathematical Questions in Biology—DNA Sequence Analysis; Miura, R.M., Ed.; American Mathematical Society: Providence, RI, USA, 1986; Volume 17, pp. 57–86. [Google Scholar]
- Schwarz, G. Estimating the Dimension of a Model. Ann. Stat. 1978, 6, 461–464. [Google Scholar] [CrossRef]
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree. 2007. Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 9 April 2023).
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-Joining Networks for Inferring Intraspecific Phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.W.; Bryant, D. POPART: Full-Feature Software for Haplotype Network Construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Rogers, A.R.; Harpending, H. Population Growth Makes Waves in the Distribution of Pairwise Genetic Differences. Mol. Biol. Evol. 1992, 9, 552–569. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E.L. Arlequin Suite Ver 3.5: A New Series of Programs to Perform Population Genetics Analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Tajima, F. Statistical Method for Testing the Neutral Mutation Hypothesis by DNA Polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Fu, Y.-X. Statistical Tests of Neutrality of Mutations Against Population Growth, Hitchhiking and Background Selection. Genetics 1997, 147, 915–925. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. MICRO-CHECKER: Software for Identifying and Correcting Genotyping Errors in Microsatellite Data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Goudet, J. FSTAT. 2005. Available online: https://www2.unil.ch/popgen/softwares/fstat.htm (accessed on 9 April 2023).
- Raymond, M.; Rousset, F. GENEPOP (Version 1.2): Population Genetics Software for Exact Tests and Ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of Population Structure Using Multilocus Genotype Data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of Population Structure Using Multilocus Genotype Data: Linked Loci and Correlated Allele Frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A Website and Program for Visualizing STRUCTURE Output and Implementing the Evanno Method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the Number of Clusters of Individuals Using the Software Structure: A Simulation Study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of Molecular Variance Inferred from Metric Distances among DNA Haplotypes: Application to Human Mitochondrial DNA Restriction Data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Wilson, G.A.; Rannala, B. Bayesian Inference of Recent Migration Rates Using Multilocus Genotypes. Genetics 2003, 163, 1177–1191. [Google Scholar] [CrossRef]
- Faubet, P.; Waples, R.S.; Gaggiotti, O.E. Evaluating the Performance of a Multilocus Bayesian Method for the Estimation of Migration Rates. Mol. Ecol. 2007, 16, 1149–1166. [Google Scholar] [CrossRef] [PubMed]
- Meirmans, P.G. Nonconvergence in Bayesian Estimation of Migration Rates. Mol. Ecol. Resour. 2014, 14, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic Analysis in Excel. Population Genetic Software for Teaching and Research—An Update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the Analysis of Population Structure. Evolution 1984, 38, 1358–1370. [Google Scholar] [CrossRef] [PubMed]
- Do, C.; Waples, R.S.; Peel, D.; Macbeth, G.M.; Tillett, B.J.; Ovenden, J.R. NeEstimator v2: Re-Implementation of Software for the Estimation of Contemporary Effective Population Size (Ne) from Genetic Data. Mol. Ecol. Resour. 2014, 14, 209–214. [Google Scholar] [CrossRef]
- Álvarez, I.; Gutiérrez, J.P.; Royo, L.J.; Fernández, I.; Gómez, E.; Arranz, J.J.; Goyache, F. Testing the Usefulness of the Molecular Coancestry Information to Assess Genetic Relationships in Livestock Using a Set of Spanish Sheep Breeds1. J. Anim. Sci. 2005, 83, 737–744. [Google Scholar] [CrossRef] [PubMed]
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 9 April 2023).
- Mantel, N.; Valand, R.S. A Technique of Nonparametric Multivariate Analysis. Biometrics 1970, 26, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Fay, J.C.; Wu, C.-I. Hitchhiking Under Positive Darwinian Selection. Genetics 2000, 155, 1405–1413. [Google Scholar] [CrossRef] [PubMed]
- Sotiropoulos, K.; Eleftherakos, K.; Džukić, G.; Kalezić, M.L.; Legakis, A.; Polymeni, R.M. Phylogeny and Biogeography of the Alpine Newt Mesotriton alpestris (Salamandridae, Caudata), Inferred from MtDNA Sequences. Mol. Phylogenet. Evol. 2007, 45, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Wielstra, B.; Salvi, D.; Canestrelli, D. Genetic Divergence across Glacial Refugia Despite Interglacial Gene Flow in a Crested Newt. Evol. Biol. 2021, 48, 17–26. [Google Scholar] [CrossRef]
- Avise, J.C. Phylogeography: The History and Formation of Species; Harvard University Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Hughes, P.D.; Woodward, J.C.; Gibbard, P.L.; Macklin, M.G.; Gilmour, M.A.; Smith, G.R. The Glacial History of the Pindus Mountains, Greece. J. Geol. 2006, 114, 413–434. [Google Scholar] [CrossRef]
- Telbisz, T.; Stergiou, C.L.; Mindszenty, A.; Chatzipetros, A. Geological and Geomorphological Characteristics of Vikos Gorge and Tymphi Mountain (Northern Pindos National Park, Greece) and Karst-Related Social Processes of the Region. Acta Carsolog. 2019, 48, 29–42. [Google Scholar] [CrossRef]
- Kolettas, S.X. The Lakes of Ioannina & Lapsista (Historical, Geographical-Ecological Study); Prefecture & Union of Municipalities of Ioannina: Ioannina, Greece, 2000. (In Greek) [Google Scholar]
- Arntzen, J.W.; Teunis, S.F.M. A Six Year Study on the Populations Dynamics of the Crested Newt (Triturus cristatus) Following the Colonisation of a Newly Created Pond. Herpetol. J. 1993, 3, 99–110. [Google Scholar]
- Kupfer, A.; Kneitz, S. Population Ecology of the Great Crested Newt (Triturus cristatus) in an Agricultural Landscape Dynamics, Pond Fidelity and Dispersal. Herpetol. J. 2000, 10, 165–171. [Google Scholar]
- Kalinowski, S.T. Counting Alleles with Rarefaction: Private Alleles and Hierarchical Sampling Designs. Conserv. Genet. 2004, 5, 539–543. [Google Scholar] [CrossRef]
- Jehle, R.; Wilson, G.A.; Arntzen, J.W.; Burke, T. Contemporary Gene Flow and the Spatio-Temporal Genetic Structure of Subdivided Newt Populations (Triturus cristatus, T. carmoratus). J. Evol. Biol. 2005, 18, 619–628. [Google Scholar] [CrossRef]
- Maletzky, A.; Kaiser, R.; Mikulíček, P. Conservation Genetics of Crested Newt Species Triturus cristatus and T. carnifex within a Contact Zone in Central Europe: Impact of Interspecific Introgression and Gene Flow. Diversity 2009, 2, 28–46. [Google Scholar] [CrossRef]
- Sotiropoulos, K.; Eleftherakos, K.; Tsaparis, D.; Kasapidis, P.; Giokas, S.; Legakis, A.; Kotoulas, G. Fine Scale Spatial Genetic Structure of Two Syntopic Newts across a Network of Ponds: Implications for Conservation. Conserv. Genet. 2013, 14, 385–400. [Google Scholar] [CrossRef]
- Unglaub, B.; Cayuela, H.; Schmidt, B.R.; Preißler, K.; Glos, J.; Steinfartz, S. Context-dependent Dispersal Determines Relatedness and Genetic Structure in a Patchy Amphibian Population. Mol. Ecol. 2021, 30, 5009–5028. [Google Scholar] [CrossRef] [PubMed]
- Schön, I.; Raepsaet, A.; Goddeeris, B.; Bauwens, D.; Mergeay, J.; Vanoverbeke, J.; Martens, K. High Genetic Diversity but Limited Gene Flow in Flemish Populations of the Crested Newt, Triturus cristatus. Belg. J. Zool. 2011, 141, 3–13. [Google Scholar]
- Nei, M.; Tajima, F. Genetic Drift And Estimation Of Effective Population Size. Genetics 1981, 98, 625–640. [Google Scholar] [CrossRef] [PubMed]
- Preisser, E.L.; Kefer, J.Y.; Lawrence, J.D.; Clark, T.W. Vernal Pool Conservation in Connecticut: An Assessment and Recommendations. Environ. Manag. 2000, 26, 503–513. [Google Scholar] [CrossRef]
- Vos, C.C.; Stumpel, A.H.P. Comparison of Habitat-Isolation Parameters in Relation to Fragmented Distribution Patterns in the Tree Frog (Hyla arborea). Landsc. Ecol. 1996, 11, 203–214. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, C.; Campo, D.; Pola, I.G.; Garcia-Vazquez, E. Rural Road Networks as Barriers to Gene Flow for Amphibians: Species-Dependent Mitigation by Traffic Calming. Landsc. Urban. Plan. 2012, 104, 171–180. [Google Scholar] [CrossRef]
- Asaeda, T.; Ca, V.T.; Wake, A. Heat Storage of Pavement and Its Effect on the Lower Atmosphere. Atmos. Environ. 1996, 30, 413–427. [Google Scholar] [CrossRef]
- Bray, R. Case Study: A Programme of Habitat Creation and Great Crested Newt Introduction to Restored Opencast Land for British Coal Opencast. In Proceedings of the Conservation and Management of Great Crested Newts: Proceedings of a Symposium, Richmond, UK, 11 January 1994; English Nature (ENS20): Peterborough, UK, 1994; pp. 113–125. [Google Scholar]
- Kinne, O. Successful Re-Introduction of the Newts Triturus cristatus and T. vulgaris. Endanger. Species Res. 2004, 1, 25–40. [Google Scholar] [CrossRef]
- McNeill, D.C. Translocation of a Population of Great Crested Newts (Triturus cristatus): A Scottish Case Study. PhD Thesis, University of Glasgow, Glasgow, UK, 2010. [Google Scholar]
- Furnborough, P.; Kirby, P.; Lambert, S.; Pankhurst, T.; Parker, P.; Piec, D. The Effectiveness and Cost Efficiency of Different Pond Restoration Techniques for Bearded Stonewort and Other Aquatic Taxa. Report on the Second Life for Ponds Project at Hampton Nature Reserve in Peterborough; Froglife: Cambridgeshire, UK, 2011. [Google Scholar]
- Lewis, B. An Evaluation of Mitigation Actions for Great Crested Newts at Development Sites. Ph.D. Thesis, The Durrell Institute of Conservation and Ecology, Canterbury, UK, 2012. [Google Scholar]
- Bounas, A.; Keroglidou, M.; Toli, E.; Chousidis, I.; Tsaparis, D.; Leonardos, I.; Sotiropoulos, K. Constrained by Aliens, Shifting Landscape, or Poor Water Quality? Factors Affecting the Persistence of Amphibians in an Urban Pond Network. Aquat. Conserv. 2020, 30, 1037–1049. [Google Scholar] [CrossRef]
- Kats, L.B.; Ferrer, R.P. Alien Predators and Amphibian Declines: Review of Two Decades of Science and the Transition to Conservation. Divers. Distrib. 2003, 9, 99–110. [Google Scholar] [CrossRef]
- Van Buskirk, J.; Schmidt, B.R. Predator-Induced Phenotypic Plasticity in Larval Newts: Trade-Offs, Selection, and Variation in Nature. Ecology 2000, 81, 3009. [Google Scholar] [CrossRef]
- Joly, P.; Miaud, C.; Lehmann, A.; Grolet, O. Habitat Matrix Effects on Pond Occupancy in Newts. Conserv. Biol. 2001, 15, 239–248. [Google Scholar] [CrossRef]
- Denoël, M.; Ficetola, G.F.; Ćirović, R.; Radović, D.; Džukić, G.; Kalezić, M.L.; Vukov, T.D. A Multi-Scale Approach to Facultative Paedomorphosis of European Newts (Salamandridae) in the Montenegrin Karst: Distribution Pattern, Environmental Variables, and Conservation. Biol. Conserv. 2009, 142, 509–517. [Google Scholar] [CrossRef]
- Covaciu-Marcov, S.-D.; Cicort-Lucaciu, A.-S.; Ferenczi, I.; Kovács, É.H.; Ferenti, S.; Sas, I. Which Aquatic Habitat Is Better for the Feeding of a Protected Newt Species (Triturus dobrogicus) in Carei Plain Natural Protected Area? Olten. Stud. Comun. Ştiinţele Nat. 2012, 28, 115–120. [Google Scholar]
- Tarkhnishvili, D.N. Amphibians of the Caucasus: Status of Natural Populations and Perspectives on Conservation. In Amphibian Populations in the Commonwealth of Independent States: Current Status and Declines; Kuzmin, S.L., Dodd, C.K., Jr., Pikulik, M.M., Eds.; Pensoft Publishers: Moscow, Russia, 1995; pp. 106–124. [Google Scholar]
- Phillips, R.A.; Patterson, D.; Shimmings, P. Increased Use of Ponds by Breeding Natterjack Toads, Bufo Calamita, Following Management. Herpetol. J. 2002, 12, 75–78. [Google Scholar]
- Smith, R.K.; Sutherland, W.J. Amphibian Conservation: Global Evidence for the Effects of Interventions; Pelagic Publishing: Exeter, UK, 2014. [Google Scholar]
- Smith, R.K.; Meredith, H.; Sutherland, W.J. Amphibian Conservation. In What Works in Conservation 2020; Sutherland, W.J., Dicks, L.V., Petrovan, S.O., Smith, R.K., Eds.; Open Book Publishers: Cambridge, UK, 2020; pp. 9–64. [Google Scholar]
- Mori, E.; Menchetti, M.; Cantini, M.; Bruni, G.; Santini, G.; Bertolino, S. Twenty Years’ Monitoring of a Population of Italian Crested Newts Triturus Carnifex: Strong Site Fidelity and Shifting Population Structure in Response to Restoration. Ethol. Ecol. Evol. 2017, 29, 460–473. [Google Scholar] [CrossRef]
- Staccione, A.; Broccoli, D.; Mazzoli, P.; Bagli, S.; Mysiak, J. Natural Water Retention Ponds for Water Management in Agriculture: A Potential Scenario in Northern Italy. J. Environ. Manag. 2021, 292, 112849. [Google Scholar] [CrossRef]
- Garner, T.W.J.; Pearman, P.B.; Angelone, S. Genetic Diversity across a Vertebrate Species’ Range: A Test of the Central-Peripheral Hypothesis. Mol. Ecol. 2004, 13, 1047–1053. [Google Scholar] [CrossRef]
- Channell, R.; Lomolino, M.V. Dynamic Biogeography and Conservation of Endangered Species. Nature 2000, 403, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Blank, L.; Sinai, I.; Bar-David, S.; Peleg, N.; Segev, O.; Sadeh, A.; Kopelman, N.M.; Templeton, A.R.; Merilä, J.; Blaustein, L. Genetic Population Structure of the Endangered Fire Salamander (Salamandra infraimmaculata) at the Southernmost Extreme of Its Distribution. Anim. Conserv. 2013, 16, 412–421. [Google Scholar] [CrossRef]
- Peterman, W.E.; Feist, S.M.; Semlitsch, R.D.; Eggert, L.S. Conservation and Management of Peripheral Populations: Spatial and Temporal Influences on the Genetic Structure of Wood Frog (Rana sylvatica) Populations. Biol. Conserv. 2013, 158, 351–358. [Google Scholar] [CrossRef]
- Lesica, P.; Allendorf, F.W. Are Small Populations of Plants Worth Preserving? Conserv. Biol. 1992, 6, 135–139. [Google Scholar] [CrossRef]
- Hunter, M.L.; Hutchinson, A. The Virtues and Shortcomings of Parochialism: Conserving Species That Are Locally Rare, but Globally Common. Conserv. Biol. 1994, 8, 1163–1165. [Google Scholar] [CrossRef]
- Hamilton, J.A.; Eckert, C.G. Population Genetic Consequences of Geographic Disjunction: A Prairie Plant Isolated on Great Lakes Alvars. Mol. Ecol. 2007, 16, 1649–1660. [Google Scholar] [CrossRef] [PubMed]
- Lambeck, R.J. Focal Species: A Multi-Species Umbrella for Nature Conservation. Especies Focales: Una Sombrilla Multiespecifica Para Conservar La Naturaleza. Conserv. Biol. 1997, 11, 849–856. [Google Scholar] [CrossRef]
- Fraser, D.J.; Bernatchez, L. Adaptive Evolutionary Conservation: Towards a Unified Concept for Defining Conservation Units. Mol. Ecol. 2001, 10, 2741–2752. [Google Scholar] [CrossRef]
- Coates, D.J.; Byrne, M.; Moritz, C. Genetic Diversity and Conservation Units: Dealing With the Species-Population Continuum in the Age of Genomics. Front. Ecol. Evol. 2018, 6, 165. [Google Scholar] [CrossRef]
- Wielstra, B.; Themudo, G.E.; Güçlü, Ö.; Olgun, K.; Poyarkov, N.A.; Arntzen, J.W. Cryptic Crested Newt Diversity at the Eurasian Transition: The Mitochondrial DNA Phylogeography of Near Eastern Triturus Newts. Mol. Phylogenet. Evol. 2010, 56, 888–896. [Google Scholar] [CrossRef]
- Arntzen, J.W.; Wielstra, B.; Wallis, G.P. The Modality of Nine Triturus Newt Hybrid Zones Assessed with Nuclear, Mitochondrial and Morphological Data. Biol. J. Linn. Soc. 2014, 113, 604–622. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clade 1 | Sub-Clade 2A | Sub-Clade 2Β | Clade 3 | T. carnifex | T. ivanbureschi | |
|---|---|---|---|---|---|---|
| Clade 1 | 0.6 (0.1) | 0.6 | 0.6 | 0.6 | 0.8 | 1.0 |
| Sub-Clade 2A | 2.8 | 0.0 (0.0) | 0.4 | 0.7 | 0.8 | 1.0 |
| Sub-Clade 2Β | 2.8 | 1.0 | 0.1 (0.0) | 0.7 | 0.8 | 1.0 |
| Clade 3 | 3.0 | 3.3 | 3.0 | 0.1 (0.1) | 0.9 | 1.1 |
| T. carnifex | 5.1 | 4.9 | 4.6 | 5.5 | - | 1.0 |
| T. ivanbureschi | 9.2 | 9.3 | 9.0 | 8.9 | 8.4 | - |
| Gene Pool | n | A | Ar | Ap | He | Ho | Ne |
|---|---|---|---|---|---|---|---|
| 1 | 71 | 4.9 | 3.0 | 14 | 0.512 | 0.464 | 290.2 |
| (E.P.) | (1.5) | (1.5) | (0.081) | (0.081) | 0.3–1456.7 | ||
| 2 | 231 | 4.4 | 1.9 | 11 | 0.568 | 0.508 | 37.9 |
| (W.P.) | (0.9) | (0.9) | (0.093) | (0.106) | (0.0–190.0) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danelis, T.; Theodoropoulos, A.; Toli, E.-A.; Bounas, A.; Korakis, A.; Sotiropoulos, K. Defining Evolutionary Conservation Units in the Macedonian Crested Newt, Triturus macedonicus (Amphibia; Salamandridae), in a Biodiversity Hotspot. Diversity 2023, 15, 671. https://doi.org/10.3390/d15050671
Danelis T, Theodoropoulos A, Toli E-A, Bounas A, Korakis A, Sotiropoulos K. Defining Evolutionary Conservation Units in the Macedonian Crested Newt, Triturus macedonicus (Amphibia; Salamandridae), in a Biodiversity Hotspot. Diversity. 2023; 15(5):671. https://doi.org/10.3390/d15050671
Chicago/Turabian StyleDanelis, Taxiarchis, Anagnostis Theodoropoulos, Elisavet-Aspasia Toli, Anastasios Bounas, Athanasios Korakis, and Konstantinos Sotiropoulos. 2023. "Defining Evolutionary Conservation Units in the Macedonian Crested Newt, Triturus macedonicus (Amphibia; Salamandridae), in a Biodiversity Hotspot" Diversity 15, no. 5: 671. https://doi.org/10.3390/d15050671
APA StyleDanelis, T., Theodoropoulos, A., Toli, E.-A., Bounas, A., Korakis, A., & Sotiropoulos, K. (2023). Defining Evolutionary Conservation Units in the Macedonian Crested Newt, Triturus macedonicus (Amphibia; Salamandridae), in a Biodiversity Hotspot. Diversity, 15(5), 671. https://doi.org/10.3390/d15050671