



Monitoring Eurasian Woodcock (Scolopax rusticola) with Pointing Dogs in Italy to Inform Evidence-Based Management of a Migratory Game Species
 ,
,
Abstract
1. Introduction
2. Methods
2.1. Field Survey
2.2. Statistical Analysis
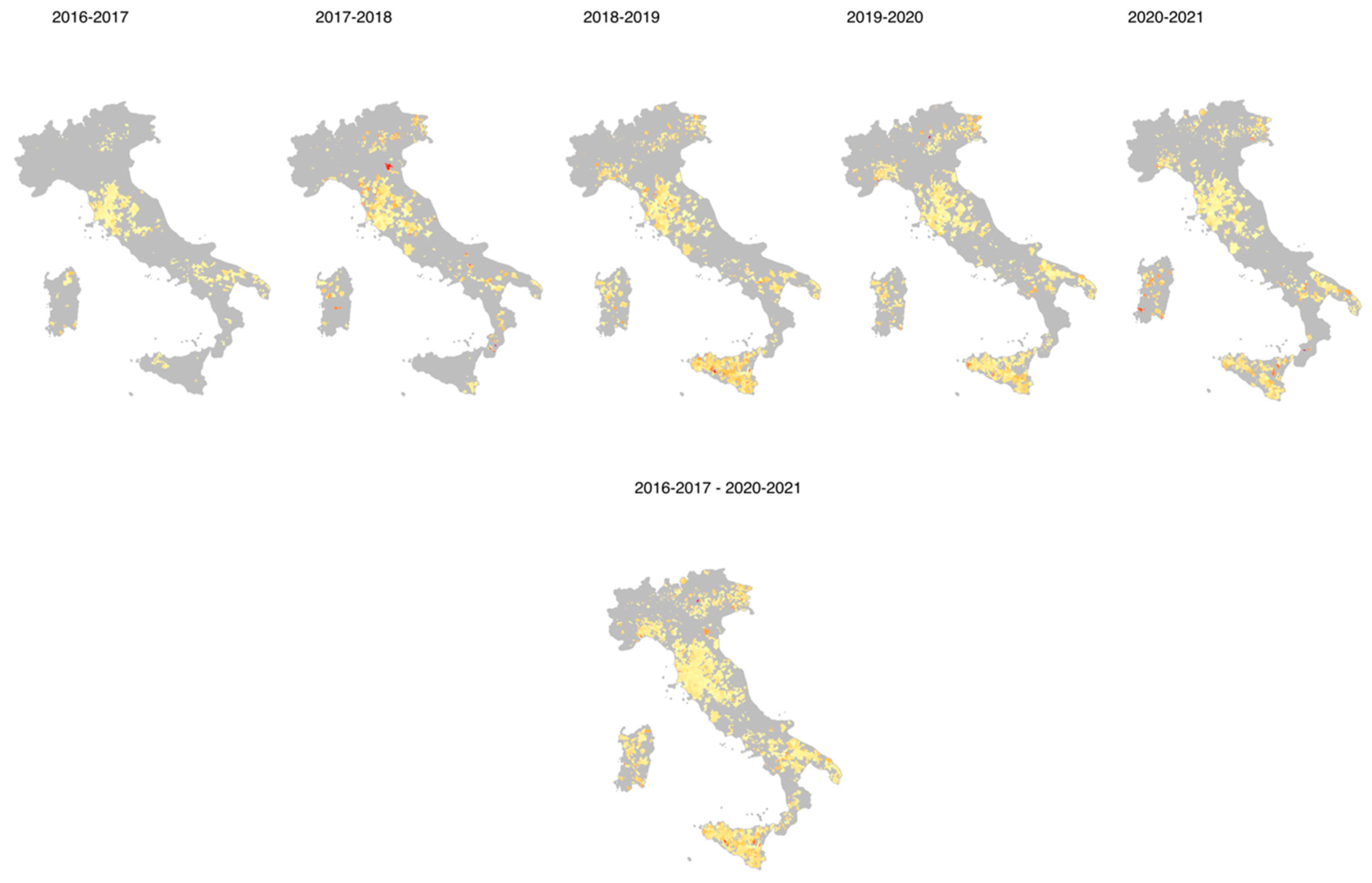
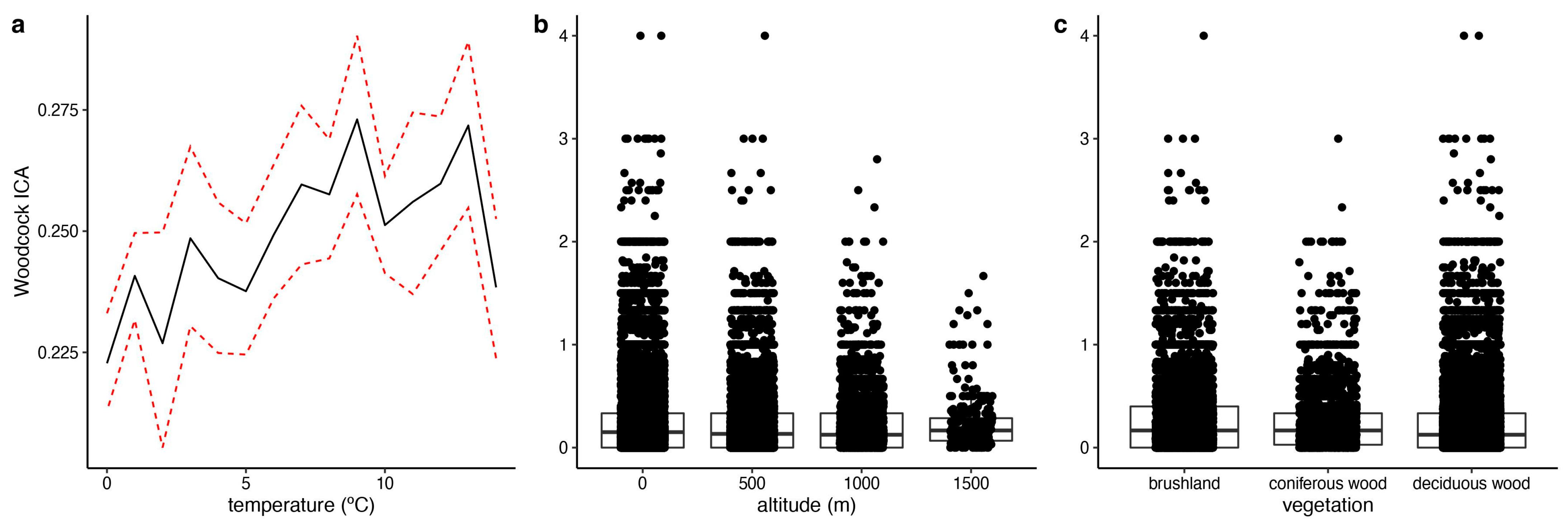
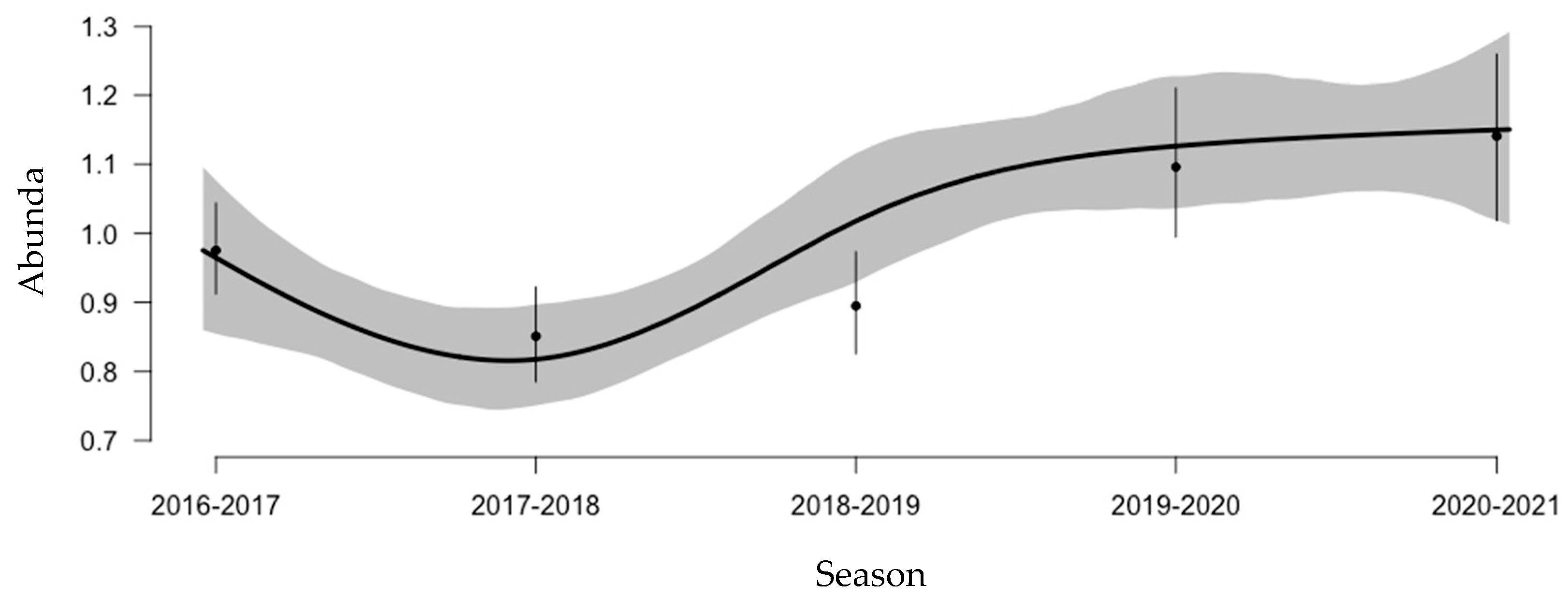
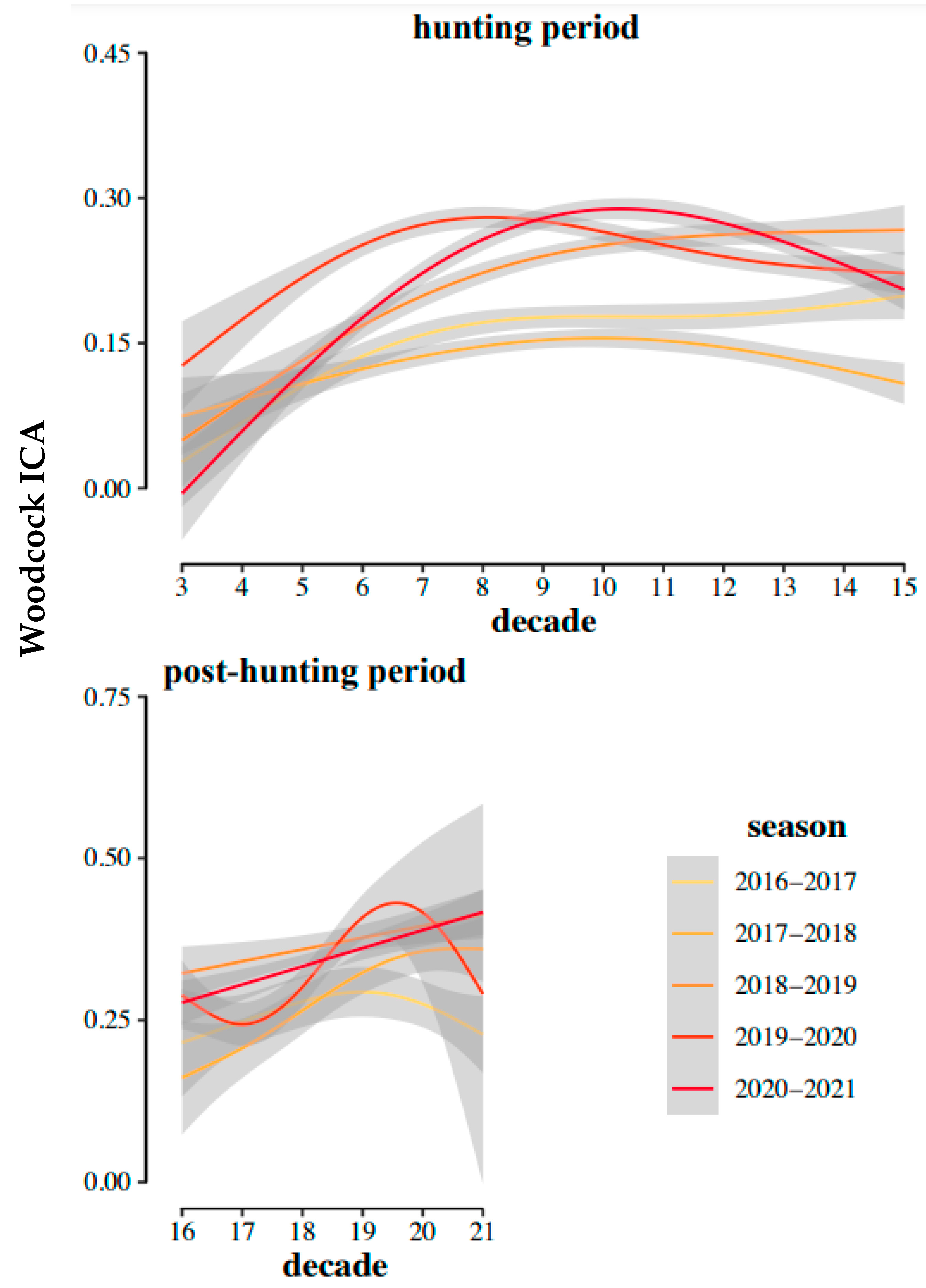
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferrand, Y.; Gossmann, F. Elements for a Woodcock (Scolopax rusticola) management plan. Game Wildl. Sci. 2001, 18, 115–139. [Google Scholar]
- Van Gils, J.; Wiersma, P.; Kirwan, G.M. Eurasian Woodcock (Scolopax rusticola), version 1.0. In Birds of the World; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar] [CrossRef]
- Hoodless, A.N.; Coulson, J.C. Survival rates and movements of British and continental Woodcock Scolopax rusticola in the British-Isles. Bird Study 1994, 41, 48–60. [Google Scholar] [CrossRef]
- Hagemeijer, W.; Blair, M. The EBBC atlas of European breeding birds. In Their Distribution and Abundance; EBCC and AD Poyser: London, UK, 1997; pp. 1–903. [Google Scholar]
- Gossmann, F.; Fokin, S.; Iljinski, I. Ringing of Woodcock in Russia from 1991 to 1997. In Fifth European Woodcock and Snipe Workshop, Proceedings of the an International Symposium of the Wetlands International Woodcock and Snipe Specialist Group. Wetlands International Global Series No. 4, and International Wader Studies 11, Czempin, Poland, 3–5 May 1998; Kalchreuter, H., Ed.; Smithsonian Libraries: Washington, DC, USA, 2000; pp. 10–14. [Google Scholar]
- Bauthian, I.; Grossmann, F.; Ferrand, Y.; Julliard, R. Quantifying the origin of Woodcock wintering in France. J. Wildl. Manag. 2007, 71, 701–705. [Google Scholar] [CrossRef]
- Ferrand, Y.; Gossmann, F. La Bécasse des Bois, Histoire Naturelle; EFFET de Lisière: Saint-Lucien, France, 2009; pp. 1–224. [Google Scholar]
- Guzmán, J.L.; Arroyo, B. Predicting winter abundance of Woodcock Scolopax rusticola using weather data: Implications for hunting management. Eur. J. Wildl. Res. 2015, 61, 467–474. [Google Scholar] [CrossRef]
- Guzmán, J.L. Factores que modulan la abundancia invernal de la becada (Scolopax rusticola): Implicaciones para su gestión y conservación. IREC Ph.D. Thesis, Universidad de Caltilla-La Mancha, Albacete, Spain, 2013. [Google Scholar]
- Ferrand, Y.; Gossmann, F.; Bastat, C.; Guénézan, M. Monitoring of the wintering and breeding Woodcock populations in France. Rev. Catalana. Ornitol. 2008, 24, 44–52. [Google Scholar]
- Cau, J.F.; Boidot, J.P. Assessment of the abundance of Woodcock over the last ten hunting seasons in France. Int. Wader Stud. 2006, 13, 24–26. [Google Scholar]
- Fadat, C. Estimation des variations relatives de densités de bécasses (Scolopax rusticola) par la méthodedes indices cynégétiques d’abondance (I.C.A.). Bull. Mens. Off. Nat. Chasse. No. Sp. Scien. Technol. Déc. 1979, 71–110. [Google Scholar]
- Gonçalves, D.; Rodrigues, T.M.; Pennacchini, P.; Lepetiti, J.P.; Taaffe, L.; Tuti, M.; Meunier, B.; Campana, J.P.; Gregori, G.; Pellegrini, A.; et al. Survey of Wintering Eurasian Woodcock in Western Europe. In Proceedings of the Eleventh American Woodcock Symposium; Krementz, D.G., Andersen, D.E., Cooper, T.R., Eds.; University of Minnesota Libraries: Minneapolis, MN, USA, 2019; pp. 240–251. [Google Scholar]
- Tuti, M.; Pennacchini, P.; Giannini, N.; Sargentini, C. Demographic structure of Woodcock (Scolopax rusticola) in Italy, ten years of observations 2010–2019. Atti Soc. Tosc. Sci. Nat. Mem. Ser. B 2021, 128, 29–37. (In Italian) [Google Scholar]
- Aradis, A.; Landucci, G.; Tagliavia, M.; Bultrini, M. Sex Determination of Eurasian Woodcock Scolopax rusticola: A molecular and morphological approach. Avocetta 2015, 39, 83–89. [Google Scholar]
- Spanò, S.; Ghelini, A. Some results of a five years study on woodcock in Italy. In Second European Woodcock and Snipe Workshop, Proceedings of International Waterfowl Research Bureau, Slimbridge, Glos, Fordingbridge, England, 30 March–1st April 1982; England, H., Kalchreuter, Herbert, Eds.; International Waterfowl Research Bureau (IWRB): London, UK, 1983; pp. 127–131. [Google Scholar]
- Aradis, A.; Miller, M.W.; Landucci, G.; Ruda, P.; Taddei, S.; Spina, F. Winter survival of Eurasian woodcock Scolopax rusticola in central Italy. Wildl. Biol. 2008, 14, 36–43. [Google Scholar] [CrossRef]
- Aradis, A.; Lo Verde, G.; Massa, B. Importance of millipedes (Diplopoda) in the autumn-winter diet of Scolopax rusticola. Eur. Zool. J. 2019, 86, 452–457. [Google Scholar] [CrossRef]
- Burlando, B.; Arillo, A.; Spanò, S. A study of the genetic variability in populations of the European woodcock (Scolopax rusticola) by random amplification of polymorphic DNA. Ital. J. Zool. 1996, 63, 31–36. [Google Scholar] [CrossRef][Green Version]
- Trucchi, E.; Allegrucci, G.; Riccarducci, G.; Aradis, A.; Spina, F.; Sbordoni, V. A genetic characterization of European Woodcock (Scolopax rusticola, Charadriidae, Charadriiformes) overwintering in Italy. Ital. J. Zool. 2011, 78, 146–156. [Google Scholar] [CrossRef][Green Version]
- Tedeschi, A.; Sorrenti, M.; Bottazzo, M.; Spagnesi, M.; Telletxea, I.; Ibàñez, R.; Tormenf, N.; De Pascalis, F.; Guidolin, L.; Rubolini, D. Interindividual variation and consistency of migratory behavior in the Eurasian Woodcock. Curr. Zool. 2020, 66, 155–163. [Google Scholar] [CrossRef] [PubMed]
- IUCN. International Union for Conservation of Nature: Annual Report 2019; IUCN: Gland, Switzerland, 2019; pp. 1–52. [Google Scholar]
- Wetlands International. Annual Reports and Accounts. 2011, pp. 1–67. Available online: www.wetlands.org (accessed on 23 April 2022).
- Wetlands International. Waterbird Population Estimates, 5th ed.; Summary Report; Wetlands International: Wageningen, The Netherlands, 2012; pp. 1–28. Available online: www.wetlands.org (accessed on 23 April 2022).
- BirdLife International. 2020. Species Factsheet: Ardea Insignis. Available online: http://datazone.birdlife.org/species/factsheet/White-bellied-Heron-Ardea-insignis (accessed on 28 April 2022).
- Bibby, C.J.; Burgess, N.D.; Hill, D.A.; Mustoe, S.H. Bird Census Techniques, 2nd ed.; Academic Press: Cambridge, MA, USA, 2000; p. 302. [Google Scholar]
- Ferrand, Y.; Aubry, P.; Gossmann, F.; Bastat, C.; Guénézan, M. Monitoring of the European woodcock populations, with special reference to France. In Proceedings of the Tenth American Woodcock Symposium, Michigan Department of Natural Resources and Environment; Stewart, C.A., Frawley, V.R., Eds.; ResearchGate: Roscommon, MI, USA, 2010; pp. 37–44. [Google Scholar]
- Lamb, E.G.; Bayne, E.; Holloway, G.; Schieck, J.; Boutin, S.; Herbers, J.; Haughland, D.L. Indices for monitoring biodiversity change: Are some more effective than others? Ecol. Indic. 2009, 9, 432–444. [Google Scholar] [CrossRef]
- Canterbury, E.G.; Martin, T.E.; Petit, D.R.; Petit, L.J.; Bradford, D.F. Bird communities and habitat as ecological indicators of forest condition in regional monitoring. Conserv. Biol. 2000, 14, 544–558. [Google Scholar] [CrossRef]
- Guetté, A.; Gaüzère, P.; Devictor, V.; Jiguet, F.; Godet, L. Measuring the synanthropy of species and communities to monitor the effects of urbanization on biodiversity. Ecol. Indic. 2017, 79, 139–154. [Google Scholar] [CrossRef]
- Padhye, A.D.; Pingankar, M.; Dahanukar, N.; Pande, S. Season and landscape element wise changes in the community structure of avifauna of Tamhini, Northern Western Ghats, India. Zoos 2007, 22, 2807–2815. [Google Scholar] [CrossRef]
- Madden, H.; Van Zanten, A. Monitoring of terrestrial avifauna in six habitats on St. Eustatius, Caribbean Netherlands, 2009–2017. Caribb. J. Sci. 2020, 50, 23–36. [Google Scholar] [CrossRef]
- Chambers, L.E.; Hughes, L.; Weston, M.A. Climate change and its impact on Australia’s avifauna. Emu 2005, 105, 1–20. [Google Scholar] [CrossRef]
- R Core Team. 2021 R: A Language and Environment for Statistical Computing. R Foundation for Statistica Computing, Vienna. Available online: https://www.R-project.org/ (accessed on 10 September 2022).
- Birtsas, P.; Sokos, C.; Papaspyropoulos, K.G.; Batselas, T.; Valiakos, G.; Billini, C. Abiotic factors and autumn migration phenology of Woodcock (Scolopax rusticola Linnaeus, 1758, Charadriiformes: Scolopacidae) in a Mediterranean area. Ital. J. Zool. 2013, 80, 392–401. [Google Scholar] [CrossRef][Green Version]
- Moreira, F.S.; Regos, A.; Gonçalves, J.F.; Rodrigues, T.M.; Verde, A.; Pagès, M.; Pérez, J.A.; Meunier, B.; Lepetit, J.-P.; Honrado, J.P.; et al. Combining citizen science data and satellite descriptors of ecosystem functioning to monitor the abundance of a migratory bird during the non-breeding season. Remote Sesns. 2022, 14, 463. [Google Scholar] [CrossRef]
- Newton, I.; Dale, L. Relationship between migration and latitude among west European birds. J. Anim. Ecol. 1996, 65, 137–146. [Google Scholar] [CrossRef]
- Forsman, J.T.; Mönkkönen, M. The role of climate in limiting European resident bird populations. J. Biogeog. 2003, 30, 55–70. [Google Scholar] [CrossRef]
- Spina, F.; Volponi, S. Atlas of Bird Migration in Italy. Vol. 1. Non-Passerines; Ministero dell’Ambiente, ISPRA, Tipografia SCR: Roma, Italy, 2008; pp. 1–32. (In Italian) [Google Scholar]
- Boidot, J.P. Mystérieuse et Fascinante Bécasse des Bois; Editions Imprimerie du Corrézien; Tome III: Naves, France, 2012; pp. 40–45. [Google Scholar]
- Tavecchia, G.; Pradel, R.; Gossman, F.; Bastat, C.; Ferrand, Y.; Lebreton, J.D. Temporal variation in annual survival probability of the Eurasian Woodcock Scolopax rusticola wintering in France. Wildl. Biol. 2002, 8, 21–30. [Google Scholar] [CrossRef]
- Doherty, K.E.; Andersen, D.E.; Meunier, J.; Oppelt, E.; Lutz, R.S.; Brug, J.C. Foraging location quality as a predictor of fidelity to a diurnal site for adult female American Woodcock Scolopax minor. Wildl. Biol. 2010, 16, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Duriéz, O.; Ferrand, Y.; Binet, F.; Corda, E.; Gossmann, F.; Fritz, H. Habitat selection of the Eurasian woodcock in winter in relation to earthworms availability. Biological. Conservation. 2005, 122, 479–490. [Google Scholar] [CrossRef]
- Fioravanti, G.; Fraschetti, P.; Lena, F.; Perconti, W.; Piervitali, E.; Pavan, V. Climate Indicators in Italy in 2020; ISPRA: Roma, Italy, 2021; pp. 1–76. ISBN 978-88-448-1062-7. (In Italian) [Google Scholar]
- Colombo, T.; Pelino, V.; Vergari, S.; Cristofanelli, P.; Bonasoni, P. Study of temperature and precipitation variations in Italy based on surface instrumental observations. Glob. Planet Chang. 2007, 57, 308–318. [Google Scholar] [CrossRef]
- Gasparini, P.; Floris, A.; Rizzo, M.; Patrone, A.; Credentino, L.; Papitto, G.; Di Martino, D. The Third Italian National Forest Inventory INFC 2015: Procedures, Tools and Applications. Available online: https://https://www.mediageo.it/ojs/index.php/GEOmedia/article/view/1767/1598 (accessed on 21 September 2022).
- Wilson, J. Wintering site fidelity of woodcock in Ireland. In Second European Woodcock and Snipe Workshop; Kalchreuter, H., Ed.; International Waterfowl Research Bureau: Slimbridge, UK, 1983; pp. 18–27. [Google Scholar]
- Imbert, G. Distribution spatio-temporelle des Bécasses (Scolopax rusticola) dans leur habitat diurne, en forêt domaniale de Boulogne-sur-mer (Pas-de-Calais) France. In Proceedings of the 3ème Symposium Européen sur la Bécasse et la Bécassine, Paris, France, 14–16 October 1988; Havet, P., Hirons, G., Eds.; pp. 53–59. [Google Scholar]
- Hirons, G.; Bickford-Smith, P. The diet and behaviour of Eurasian woodcock wintering in Cornwall. In Second European Woodcock and Snipe Workshop; Kalchreuter, H., Ed.; International Waterfowl Research Bureau: Slimbridge, UK, 1983; pp. 11–17. [Google Scholar]
- Binet, F. Dynamique des Peuplements et Fonctions des Lombriciens en Sols Cultivés Tempérés. Ph.D. Thesis, Université de Rennes 1, Rennes, France, 1993; p. 299. [Google Scholar]
- FAROW. FANPBO Annual Report on Woodcock 2015–2016 Hunting Season; Federation Associations Nationales Beccássiers Paléarctique Occidental, FANBPO, Ed.; University of Porto Press: Porto, Portugal, 2017; p. 44. [Google Scholar]
- Tuti, M.; Gambogi, R.; Galardini, A. The Eurasian Woodcock (Scolopax rusticola) At San Rossore Estate (Migliarino San Rossore Massaciuccoli Regional Park, Tuscany, Italy): A Four Years Study (In Italian). Atti Soc. Tosc. Sci. Nat. Mem. Ser. B 2017, 124, 109–119. [Google Scholar]
- Péron, G.; Ferrand, Y.; Gossmann, F.; Bastat, C.; Guénézan, M.; Gimenez, O. Escape migration decisions in Eurasian Woodcocks: Insights from survival analyses using large-scale recovery data. Behav. Ecol. Sociobiol. 2011, 65, 1949–1955. [Google Scholar] [CrossRef]
- Guzmán, J.L.; Caro, J.; Arroyo, B. Factors influencing mobility and survival of Eurasian Woodcock wintering in Spain. Avian Conserv. Ecol. 2017, 12, 21. [Google Scholar] [CrossRef]
- Bairlein, F.; Mattig, F.; Ambrosini, R. 2022. Analysis of the current migration seasons of hunted species as of Key Concepts of article 7(4) of Directive 79/409/EEC. In The Eurasian African Bird Migration Atlas; Spina, F., Baillie, S.R., Bairlein, F., Fiedler, W., Thorup, K., Eds.; EURING/CMS; 2022; Available online: https://migrationatlas.org (accessed on 12 January 2023).
- Casanova, P.; Memoli, A. Woodcock: A difficult migrant to manage [In Italian]. L’Italia For. E Mont. Anno LXIII 2008, 4, 351–363. [Google Scholar] [CrossRef]
- Leech, D.I.; Crick, H.Q.P. Influence of climate change on the abundance, distribution and phenology of woodland bird species in temperate regions. Ibis 2007, 149, 128–145. [Google Scholar] [CrossRef]
- Desiato, F.; Fioravanti, G.; Fraschetti, P.; Perconti, W.; Piervitali, E.; Pavan, V. Climate indicators in Italy in 2017; ISPRA: Roma, Italy, 2018; p. 75. ISBN 978-88-448-0904-1. (In Italian) [Google Scholar]
- Murphy, K.; Roberts, D.; Jensen, W.; Nielsen, S.; Johnson, S.; Hosek, B.; Stillings, B.; Kolar, J.; Boyce, M.; Ciuti, S. Mule deer fawn recruitment dynamics in an energy disturbed landscape. Ecol. Evol. 2023, 13, e9976. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | |||||||
|---|---|---|---|---|---|---|---|
| Sampling | Month | Decade | 2016–2017 | 2017–2018 | 2018–2019 | 2019–2020 | 2020–2021 |
| Hunting period | September | - | |||||
| 2 | 2 | 7 | 1 | 5 | 1 | ||
| 3 | 20 | 16 | 17 | 13 | 14 | ||
| October | 4 | 71 | 81 | 109 | 150 | 141 | |
| 5 | 117 | 144 | 272 | 448 | 342 | ||
| 6 | 278 | 436 | 438 | 729 | 766 | ||
| November | 7 | 291 | 555 | 746 | 983 | 932 | |
| 8 | 416 | 635 | 947 | 773 | 595 | ||
| 9 | 312 | 416 | 831 | 926 | 571 | ||
| December | 10 | 339 | 489 | 1016 | 964 | 580 | |
| 11 | 272 | 321 | 676 | 770 | 1093 | ||
| 12 | 281 | 381 | 833 | 894 | 399 | ||
| January | 13 | 166 | 220 | 482 | 665 | 478 | |
| 14 | 169 | 261 | 386 | 608 | 574 | ||
| 15 | 209 | 205 | 251 | 371 | 466 | ||
| Post-hunting period | February | 16 | 33 | 38 | 170 | 205 | 174 |
| 17 | 142 | 128 | 269 | 296 | 466 | ||
| 18 | 96 | 164 | 271 | 468 | 582 | ||
| March | 19 | 110 | 245 | 546 | 439 | 476 | |
| 20 | 170 | 243 | 230 | 13 | 433 | ||
| 21 | 89 | 176 | 225 | 4 | 174 | ||
| Edf | Ref.df | Chi-Square | p-Value | |
|---|---|---|---|---|
| Season | 3.921 | 3.994 | 240.6 | <0.001 |
| Day | 7.920 | 8.000 | 6593.7 | <0.001 |
| x,y | 16.805 | 18.284 | 496.5 | <0.001 |
| Hunter | 938.483 | 1263.000 | 13,217.0 | <0.001 |
| Hunting Period (37.7%) | Post-Hunting Period (31.1%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Edf | Ref.df | Chi sq. | p-Value | Edf | Ref.df | Chi sq. | p-Value | |
| Season | 3.97 | 3.99 | 554.8 | <0.001 | 3.54 | 3.86 | 14.3 | <0.001 |
| Decade | 2.99 | 3.00 | 598.4 | <0.001 | 1.58 | 1.82 | 63.1 | <0.001 |
| Commune | 401.62 | 1542.0 | 29,742.9 | <0.001 | 100.80 | 644.00 | 711.9 | 0.001 |
| Hunter | 769.51 | 1058.0 | 52,924.6 | <0.001 | 417.60 | 742.00 | 2749.9 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuti, M.; Rodrigues, T.M.; Bongi, P.; Murphy, K.J.; Pennacchini, P.; Mazzarone, V.; Sargentini, C. Monitoring Eurasian Woodcock (Scolopax rusticola) with Pointing Dogs in Italy to Inform Evidence-Based Management of a Migratory Game Species. Diversity 2023, 15, 598. https://doi.org/10.3390/d15050598
Tuti M, Rodrigues TM, Bongi P, Murphy KJ, Pennacchini P, Mazzarone V, Sargentini C. Monitoring Eurasian Woodcock (Scolopax rusticola) with Pointing Dogs in Italy to Inform Evidence-Based Management of a Migratory Game Species. Diversity. 2023; 15(5):598. https://doi.org/10.3390/d15050598
Chicago/Turabian StyleTuti, Marco, Tiago M. Rodrigues, Paolo Bongi, Kilian J. Murphy, Paolo Pennacchini, Vito Mazzarone, and Clara Sargentini. 2023. "Monitoring Eurasian Woodcock (Scolopax rusticola) with Pointing Dogs in Italy to Inform Evidence-Based Management of a Migratory Game Species" Diversity 15, no. 5: 598. https://doi.org/10.3390/d15050598
APA StyleTuti, M., Rodrigues, T. M., Bongi, P., Murphy, K. J., Pennacchini, P., Mazzarone, V., & Sargentini, C. (2023). Monitoring Eurasian Woodcock (Scolopax rusticola) with Pointing Dogs in Italy to Inform Evidence-Based Management of a Migratory Game Species. Diversity, 15(5), 598. https://doi.org/10.3390/d15050598