
Scale-Dependent Spatial Ecology of Paleotropical Leaf Litter Ants (Hymenoptera: Formicidae)

Abstract
1. Introduction
2. Materials and Methods
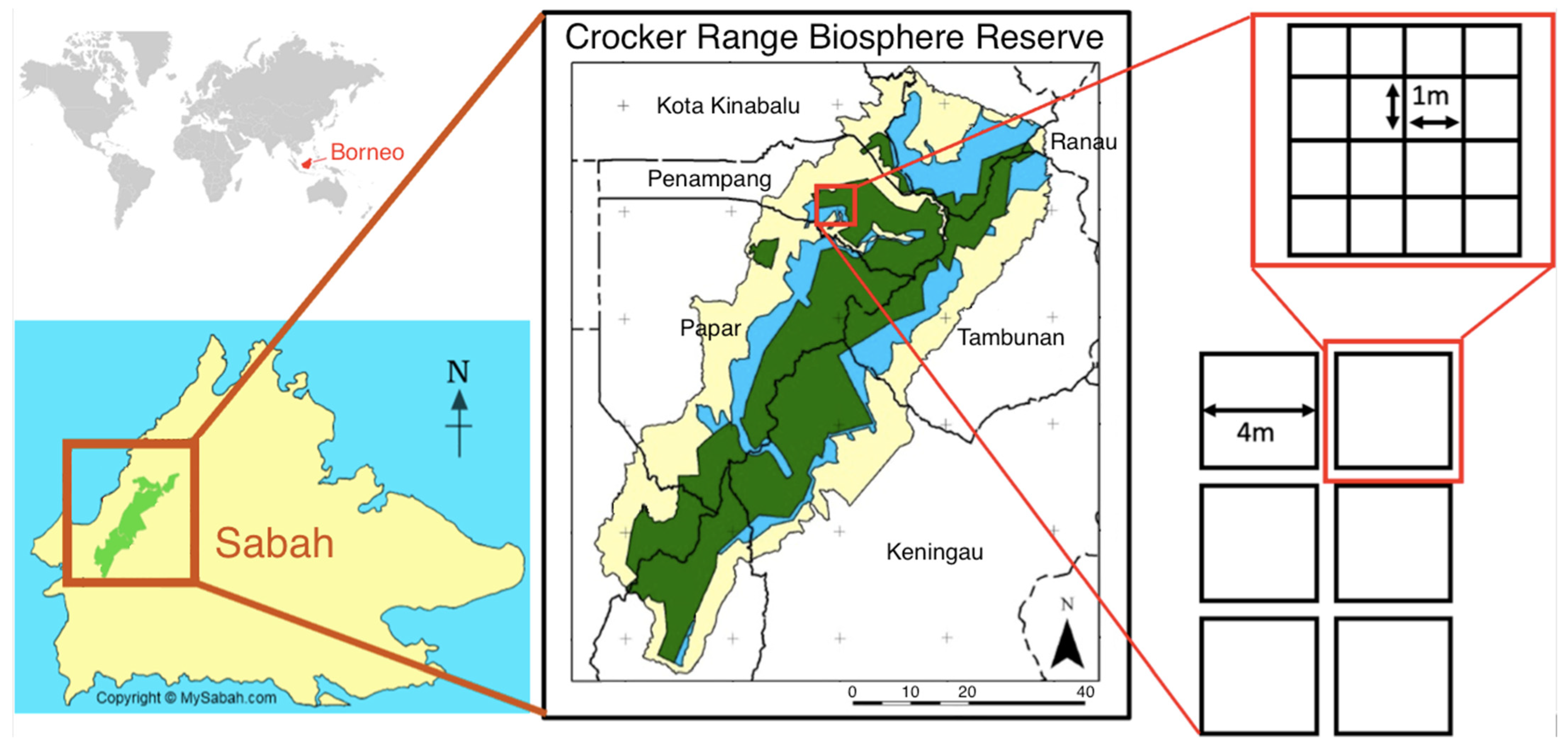
2.1. Study Site
2.2. Field Methods
2.3. Data Analysis
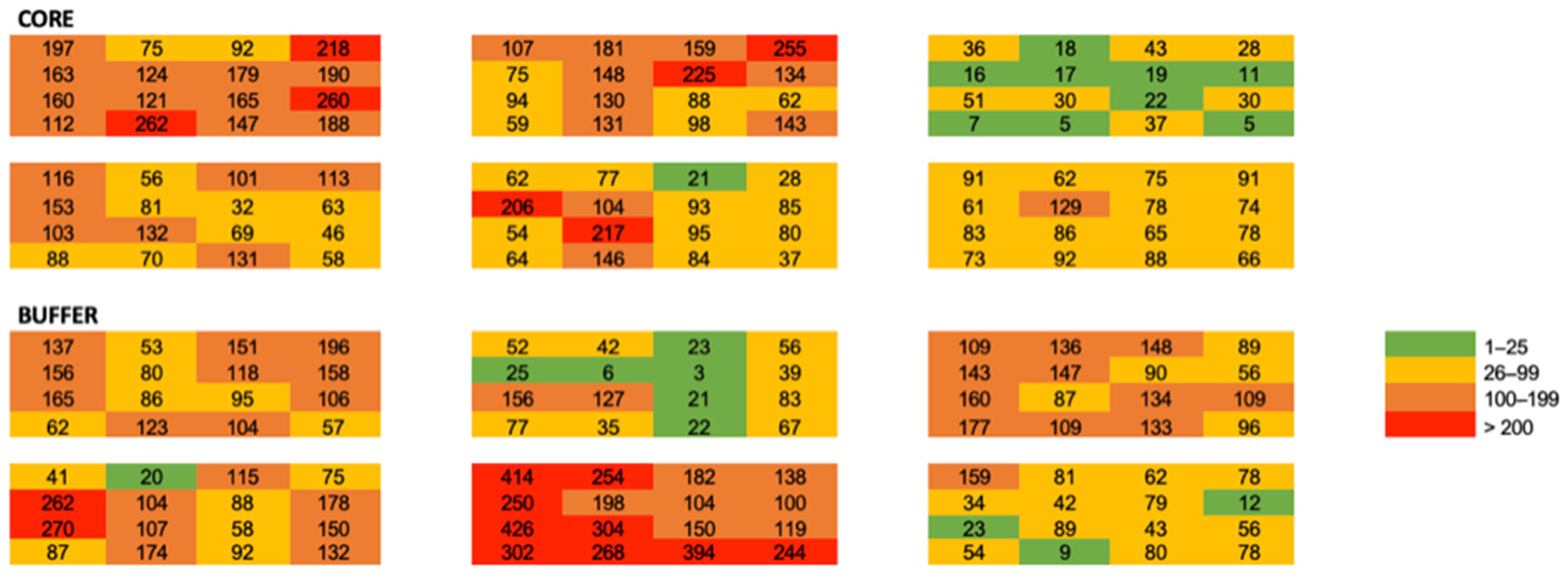
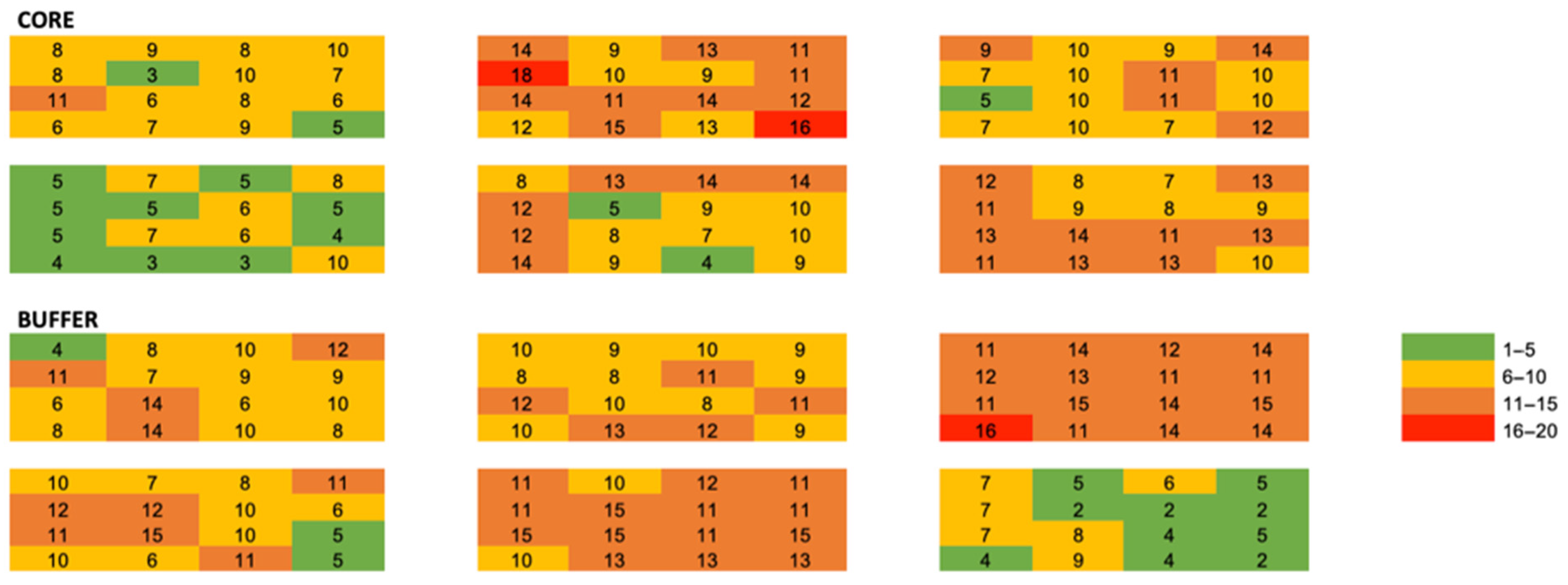
3. Results
Small Scale Spatial Ecology
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, C.; Dudley, K.L.; Xu, Z.-H.; Economo, E.P. Mountain metacommunities: Climate and spatial connectivity shape ant diversity in a complex landscape. Ecography 2018, 41, 101–112. [Google Scholar] [CrossRef]
- Andersen, A.N. Using ants as bioindicators: Multiscale issues in ant community ecology. Conserv. Ecol. 1997, 1, 8. [Google Scholar] [CrossRef]
- Sanders, N.J.; Lessard, J.P.; Fitzpatrick, M.C.; Dunn, R.R. Temperature, but not productivity or geometry, predicts elevation diversity gradients in ants across spatial grains. Glob. Ecol. Biogeogr. 2007, 16, 640–649. [Google Scholar] [CrossRef]
- Kaspari, M.; Yuan, M.; Alonso, L. Spatial grain and the causes of regional diversity gradients in ants. Am. Nat. 2003, 161, 459–477. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.M.; Kulig, A.W. Founding, foraging, and fighting: Colony size and the spatial distribution of harvester ant nests. Ecology 1996, 77, 2393–2409. [Google Scholar] [CrossRef]
- Verble, R.M.; Yanoviak, S.P. Short-term effects of prescribed burning on ant (Hymenoptera: Formicidae) assemblages in Ozark forests. Ann. Entomol. Soc. Am. 2013, 100, 198–203. [Google Scholar] [CrossRef]
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Spiesman, B.J.; Cumming, G.S. Communities in context: The influences of multiscale environmental variation on local ant community structure. Landsc. Ecol. 2008, 23, 313–325. [Google Scholar] [CrossRef]
- Kaspari, M. Testing resource-based models of patchiness in four neotropical litter ant assemblages. Oikos 1996, 76, 443–454. [Google Scholar] [CrossRef]
- Law, S.J.; Parr, C. Numerically dominant species drive patterns in resource use along a vertical gradient in tropical ant assemblages. Biotropica 2019, 52, 101–112. [Google Scholar] [CrossRef]
- Yusah, K.M.; Foster, W.A.; Reynolds, G.; Fayle, T.M. Ant mosaics in Bornean primary rain forest high canopy depend on spatial scale, time of day, and sampling method. PeerJ 2018, 6, e4231. [Google Scholar] [CrossRef] [PubMed]
- Thenuis, L.; Gilbert, M.; Roisin, Y.; Leponce, M. Spatial structure of litter-dwelling ant distribution in a subtropical dry forest. Insectes Sociaux 2005, 52, 366–377. [Google Scholar] [CrossRef]
- Kaspari, M. Litter ant patchiness at the 1-m2 scale: Disturbance dynamics in three Neotropical forests. Oecologia 1996, 107, 265–273. [Google Scholar] [CrossRef]
- Leponce, M.; Theunis, L.; Delabie, J.H.C.; Roisin, Y. Scale dependence of diversity measures in a leaf-litter ant assemblage. Ecography 2004, 27, 253–267. [Google Scholar] [CrossRef]
- Vasconcelos, H.L.; Macedo, A.C.C.; Vilhena, J.M.S. Influence of topography on the distribution of ground-dwelling ants in an Amazonian Forest. Stud. Neotrop. Fauna Environ. 2003, 38, 115–124. [Google Scholar] [CrossRef]
- Herbers, J.M. Community structure in north temperate ants: Temporal and spatial variation. Oecologia 1989, 81, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.O. Patchy distributions of ant species in New Guinea rain forests. Psyche 1958, 65, 26–38. [Google Scholar] [CrossRef]
- Bruhl, C.A. Leaf Litter Ant Communities in Tropical Lowland Rain Forests in Sabah, Malaysia: Effects of Forest Disturbance and Fragmentation. Ph.D. Thesis, Julius-Maximilians-Universität Würzburg, Würzburg, Germany, 2001. [Google Scholar]
- Holldobler, B.; Wilson, E.O. The Ants; Harvard University Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Karl, J.W.; Maurer, B.A. Spatial dependence of predictions from image segmentation: A variogram-based method to determine appropriate scales for producing land-management information. Ecol. Inform. 2010, 5, 194–202. [Google Scholar] [CrossRef]
- Yoh, N.; Azhar, I.; Fitzgerald, K.V.; Yu, R.; Smith-Butler, T.; Mahyudin, A.; Kingston, T. Bat ensembles differ in response to use zones in a tropical biosphere reserve. Diversity 2020, 12, 60. [Google Scholar] [CrossRef]
- Southwood, T.R.E.; Brown, V.K.; Reader, P.M. The relationships of plant and insect diversities in succession. Biol. J. Linn. Soc. 1978, 12, 327–348. [Google Scholar] [CrossRef]
- Bestelmeyer, B.T.; Agosti, D.; Alonso, L.E.; Brandão, C.R.F.; Brown, W.L.; Delabie, J.H.C.; Silvestre, R. Field techniques for the study of ground dwelling ants. An overview, description, and evaluation. In Ants: Standard Methods for Measuring and Monitoring Biodiversity; Agosti, D., Majer, J.D., Alonso, L.E., Schultz, T.R., Eds.; Smithsonian Institution Press: Washington, DC, USA; London, UK, 2000; pp. 122–144. [Google Scholar]
- Bolton, B. Identification Guide to the Ant Genera of the World; Harvard University Press: Cambridge, MA, USA, 1995. [Google Scholar]
- Hashimoto, Y. Identification guide to the ant genera of Borneo. In Inventory & Collection: Total Protocol for Understanding of Biodiversity; International Union for Conservation of Nature: Fontainebleau, France, 2003; Chapter 9; pp. 89–162. [Google Scholar]
- General, D.M.; Alpert, G.D. A synoptic review of the ant genera (Hymenoptera: Formicidae) of the Philippines. Zookeys 2012, 1, 1–111. [Google Scholar] [CrossRef] [PubMed]
- Souza, J.L.P.; Baccaro, F.B.; Landeiro, V.L.; Franklin, E.; Magnusson, W.E.; Pequeno, P.A.C.L.; Fernandes, I.O. Taxonomic sufficiency and indicator taxa reduce sampling costs and increase monitoring effectiveness for ants. Divers. Distrib. 2016, 22, 111–122. [Google Scholar] [CrossRef]
- Colwell, R.K.; Mao, C.X.; Chang, J. Interpolating, extrapolating, and comparing incidence-based species accumulation curves. Ecology 2004, 85, 2717–2727. [Google Scholar] [CrossRef]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples; Wiley: New York, NY, USA, 2013; Version 9. [Google Scholar]
- Eguchi, K.; Yamane, S. Species diversity of ants (Hymenoptera, Formicidae) in a lowland rainforest, Northwestern Borneo. New Entomol. 2003, 52, 2. [Google Scholar]
- Brühl, C.A.; Gunsalam, G.; Linsenmair, K.E. Stratification of ants (Hymenoptera, Formicidae) in a primary rain forest in Sabah, Borneo. J. Trop. Ecol. 1998, 14, 285–297. [Google Scholar] [CrossRef]
- Brühl, C.A.; Eltz, T. Fueling the biodiversity crisis: Species loss of ground-dwelling forest ants in oil palm plantations in Sabah, Malaysia (Borneo). Biodivers. Conserv. 2010, 19, 519–529. [Google Scholar] [CrossRef]
- Kamel, M.; Semida, F.; Moursy, M.; Kassem, H. Impact of hiking trail disturbance on ants’ diversity at Mount Sinai, St. Katherine protectorate, Egypt. J. Environ. Sci. 2019, 45, 37–56. [Google Scholar] [CrossRef]
- Barber, G.W. Ant (Hymenoptera: Formicidae) Assemblages in Three New York Pine Barrens and the Impacts of Hiking Trails. Master’s Thesis, University of Massachusetts Amherst, Amherst, MA, USA, 2015. [Google Scholar]
- Kwon, T.S. Ant assemblages along the Baekdudaegan Mountain Range in South Korea: Human roads and temperature niche. J. Asia-Pacific Biodivers. 2015, 8, 152–157. [Google Scholar] [CrossRef]
- Sanford, M.P.; Manley, P.N.; Murphy, D.D. Effects of urban development on ant communities: Implications for ecosystem services and management. Conserv. Biol. 2009, 23, 131–141. [Google Scholar] [CrossRef]
- Nakamura, A.; Catterall, C.P.; House, A.P.; Kitching, R.L.; Burwell, C.J. The use of ants and other soil and litter arthropods as bio-indicators of the impacts of rainforest clearing and subsequent land use. J. Insect Conserv. 2007, 11, 177–186. [Google Scholar] [CrossRef]
- Silva, P.S.; Bieber, A.G.; Corrêa, M.M.; Leal, I.R. Do leaf-litter attributes affect the richness of leaf-litter ants? Neotrop. Entomol. 2011, 40, 542–547. [Google Scholar] [PubMed]
- Levings, S.C. Seasonal, annual, and among-site variation in the ground ant community of a deciduous tropical forest: Some causes of patchy species distributions. Ecol. Monographs 1983, 53, 435–455. [Google Scholar] [CrossRef]
- Ribas, C.R.; Schoereder, J.H.; Pic, M.; Soares, S.M. Tree heterogeneity, resource availability, and larger scale processes regulating arboreal ant species richness. Austral Ecol. 2003, 28, 305–314. [Google Scholar] [CrossRef]
- Xing, S.; Hood, A.S.C.; Dial, R.J.; Fayle, T.M. Species turnover in ant assemblages is greater horizontally than vertically in the world’s tallest tropical forest. Ecol. Evol. 2022, 12, e9158. [Google Scholar] [CrossRef]
- Morlon, H.; Chuyong, G.; Condit, R.; Hubbell, S.; Kenfack, D.; Thomas, D.; Valencia, R.; Green, J.L. A general framework for the distance-decay of similarity in ecological communities. Ecol. Lett. 2008, 11, 904–917. [Google Scholar] [CrossRef]
- AntWiki: The Ants—Online. 2023. Available online: https://www.antwiki.org/wiki/Nylanderia#South_Pacific (accessed on 1 February 2023).
- Williams, J.L.; Lucky, A. Non-native and invasive Nylanderia crazy ants (Hymenoptera: Formicidae) of the world: Integrating genomics to enhance taxonomic preparedness. Ann. Entomol. Soc. Am. 2020, 113, 318–336. [Google Scholar] [CrossRef]
- Rubiana, R.; Rizali, A.; Denmead, L.H.; Alamsari, W.; Hidayat, P.; Pudjianto, H.D.; Clough, Y.; Tscharntke, T.; Buchori, D. Agricultural land use alters species composition but not species richness of ant communities. Asian Myrmecol. 2015, 7, 73–85. [Google Scholar]
- Hamer, M.T.; Marquis, A.D.; Guenard, B. Strumigenys perplexa (Smith, 1876) (Formicidae: Myrmicinae) a new exotic ant to Europe with establishment in Guernsey, Channel Islands. J. Hymenopt. Res. 2021, 83, 101–124. [Google Scholar] [CrossRef]
- Leitao, R.P.; Zuanon, J.; Villeger, S.; Williams, S.E.; Baraloto, C.; Fortunel, C.; Mendonca, F.P.; Mouillot, D. Rare species contribute disproportionately to the functional structure of species assemblages. Proc. R. Soc. B 2016, 283, 20160084. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subfamily | Genus | Total Abundance | Genus RA | Subfamily RA | Genus Total Abundance Buffer | Genus Total Abundance Core |
|---|---|---|---|---|---|---|
| Amblyoponinae | Mystrium | 3 | <0.001 | 0.002 | 3 | 0 |
| Stigmatomma | 41 | <0.001 | 26 | 15 | ||
| Dolichoderinae | Dolichoderus | 2 | <0.001 | 0.063 | 0 | 2 |
| Loweriella | 454 | 0.022 | 409 | 45 | ||
| Dorylinae | Aenictus | 69 | 0.003 | 0.006 | 35 | 34 |
| Cerapachys | 7 | <0.001 | 7 | 0 | ||
| Lioponera | 18 | <0.001 | 0 | 18 | ||
| Parasycia | 26 | 0.001 | 16 | 10 | ||
| Ectatomminae | Gnamptogenys | 14 | <0.001 | <0.001 | 4 | 10 |
| Formicinae | Anaplolepis | 2 | <0.001 | 0.140 | 1 | 1 |
| Camponotus | 28 | 0.001 | 15 | 13 | ||
| Colobopsis | 2 | <0.001 | 0 | 2 | ||
| Echinopla | 2 | <0.001 | 2 | 0 | ||
| Euprenolepis | 2 | <0.001 | 0 | 2 | ||
| Myrmoteras | 167 | 0.008 | 119 | 48 | ||
| Nylanderia | 2486 | 0.120 | 1939 | 547 | ||
| Paraparatrachina | 13 | <0.001 | 2 | 11 | ||
| Polyrachis | 6 | <0.001 | 1 | 5 | ||
| Prenolepis | 93 | 0.004 | 93 | 0 | ||
| Pseudolasius | 29 | 0.001 | 29 | 0 | ||
| Leptanillinae | Leptanilla | 2 | <0.001 | <0.001 | 0 | 2 |
| Myrmicinae | Acanthomyrmex | 60 | 0.003 | 0.695 | 25 | 35 |
| Calyptomyrmex | 58 | 0.003 | 29 | 29 | ||
| Cardiocondyla | 34 | 0.002 | 26 | 8 | ||
| Carebara | 3537 | 0.171 | 1683 | 1854 | ||
| Cataulacus | 2 | <0.001 | 2 | 0 | ||
| Crematogaster | 50 | 0.002 | 2 | 48 | ||
| Eurhopalothrix | 12 | <0.001 | 0 | 12 | ||
| Lophomyrmex | 2932 | 0.142 | 1347 | 1585 | ||
| Lordomyrma | 13 | <0.001 | 7 | 6 | ||
| Mayriella | 322 | 0.016 | 233 | 89 | ||
| Meranoplus | 29 | 0.001 | 25 | 4 | ||
| Monomorium | 37 | 0.002 | 37 | 0 | ||
| Myrmecina | 114 | 0.006 | 35 | 79 | ||
| Myrmicaria | 309 | 0.015 | 0 | 309 | ||
| Pheidole | 4284 | 0.207 | 2374 | 1910 | ||
| Pristomyrmex | 65 | 0.003 | 31 | 34 | ||
| Proatta | 73 | 0.004 | 73 | 0 | ||
| Solenopsis | 77 | 0.004 | 4 | 73 | ||
| Strumigenys | 1154 | 0.056 | 970 | 184 | ||
| Syllophopsis | 81 | 0.004 | 40 | 41 | ||
| Tetramorium | 1018 | 0.049 | 537 | 481 | ||
| Vollenhovia | 92 | 0.004 | 40 | 52 | ||
| Vombisidris | 2 | <0.001 | 0 | 2 | ||
| Ponerinae | Anochetus | 28 | 0.001 | 0.096 | 21 | 7 |
| Brachyponera | 1235 | 0.060 | 765 | 470 | ||
| Cryptopone | 16 | <0.001 | 12 | 4 | ||
| Diacamma | 1 | <0.001 | 1 | 0 | ||
| Ectomomyrmex | 7 | <0.001 | 3 | 4 | ||
| Emeryopone | 1 | <0.001 | 1 | 0 | ||
| Harpagnathos | 2 | <0.001 | 2 | 0 | ||
| Hypoponera | 424 | 0.021 | 246 | 178 | ||
| Leptogenys | 64 | 0.003 | 44 | 20 | ||
| Mesoponera | 1 | <0.001 | 1 | 0 | ||
| Myopias | 32 | 0.002 | 26 | 6 | ||
| Odontomachus | 51 | 0.002 | 7 | 44 | ||
| Odontoponera | 59 | 0.003 | 39 | 20 | ||
| Ponera | 52 | 0.003 | 19 | 33 | ||
| Proceratiinae | Discothyrea | 11 | <0.001 | <0.001 | 5 | 6 |
| Probolomyrmex | 3 | <0.001 | 1 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Germeroth, L.; Sumnicht, T.; Verble, R. Scale-Dependent Spatial Ecology of Paleotropical Leaf Litter Ants (Hymenoptera: Formicidae). Diversity 2023, 15, 494. https://doi.org/10.3390/d15040494
Germeroth L, Sumnicht T, Verble R. Scale-Dependent Spatial Ecology of Paleotropical Leaf Litter Ants (Hymenoptera: Formicidae). Diversity. 2023; 15(4):494. https://doi.org/10.3390/d15040494
Chicago/Turabian StyleGermeroth, Lillian, Theodore Sumnicht, and Robin Verble. 2023. "Scale-Dependent Spatial Ecology of Paleotropical Leaf Litter Ants (Hymenoptera: Formicidae)" Diversity 15, no. 4: 494. https://doi.org/10.3390/d15040494
APA StyleGermeroth, L., Sumnicht, T., & Verble, R. (2023). Scale-Dependent Spatial Ecology of Paleotropical Leaf Litter Ants (Hymenoptera: Formicidae). Diversity, 15(4), 494. https://doi.org/10.3390/d15040494