

Diverse Host Spectrum and the Parasitic Process in the Pantropical Hemiparasite Cassytha filiformis L. (Lauraceae) in China

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Growth Conditions of C. filiformis
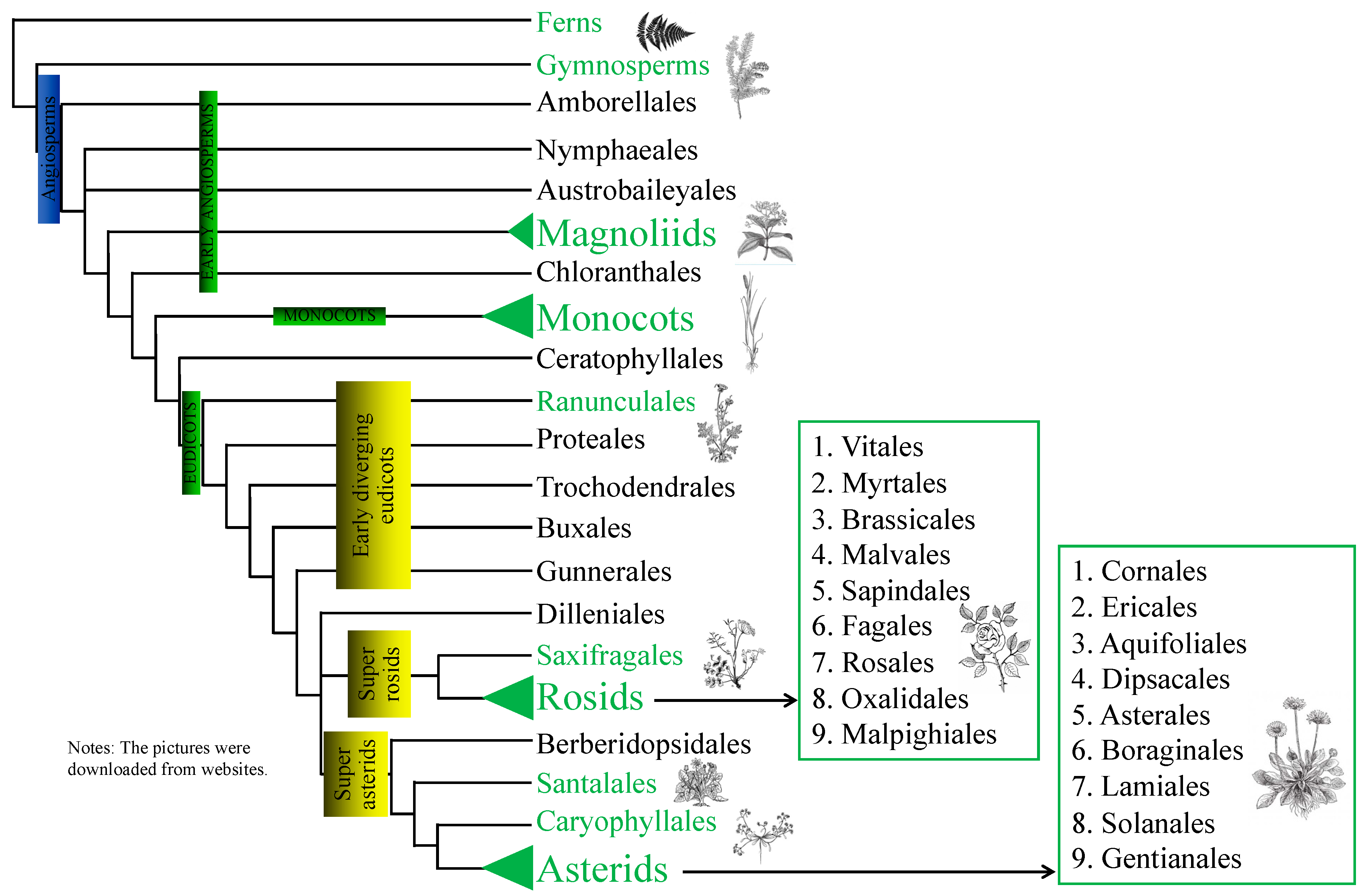
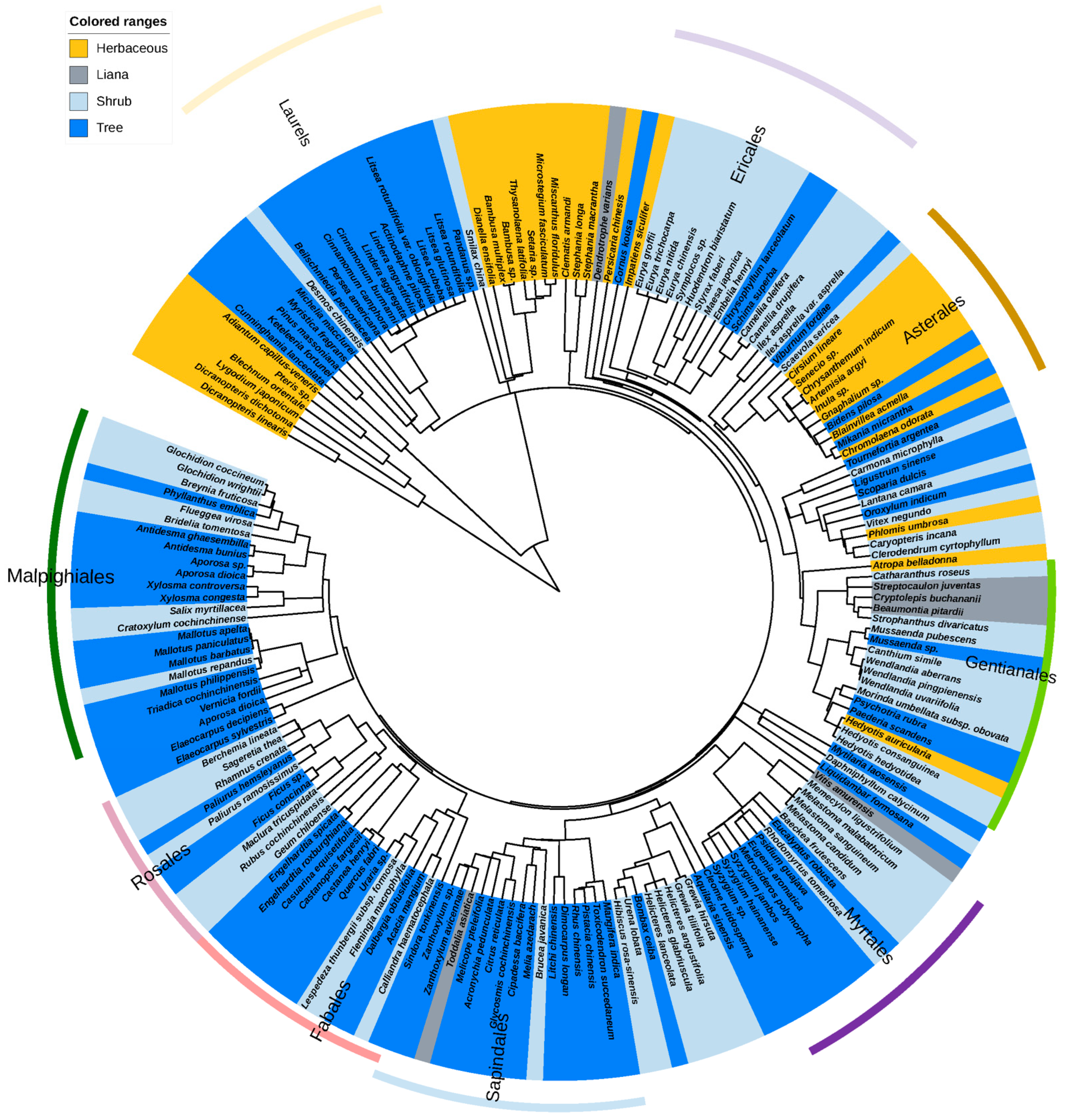
2.3. Phylogenic Analysis
3. Results
3.1. Biology of Laurel Dodder and Its Hosts
3.2. Host Range of C. filiformis
4. Discussion
4.1. Laurel Dodder Prefers Woody Hosts
4.2. The Host Phylogenetic Relationships of Laurel Dodder and Its Potential Application
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nickrent, D.L. Parasitic angiosperms: How often and how many? Taxon 2020, 69, 5–27. [Google Scholar] [CrossRef]
- Twyford, A.D. Parasitic plants. Curr. Biol. 2018, 28, R857–R859. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, Y.; Xie, J.; Zhuang, H.; Liu, H.; Shen, G.; Wu, J. Parasite dodder enables transfer of bidirectional systemic nitrogen signals between host plants. Plant Physiol. 2021, 185, 1395–1410. [Google Scholar] [CrossRef] [PubMed]
- Yoder, J.I.; Scholes, J.D. Host plant resistance to parasitic weeds; recent progress and bottlenecks. Curr. Opin. Plant Biol. 2010, 13, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Irving, L.J.; Cameron, D.D. You are what you eat: Interactions between root parasitic plants and their hosts. Adv. Bot. Res. 2009, 50, 87–138. [Google Scholar]
- Zhang, H.; Florentine, S.; Tennakoon, K.U. The angiosperm stem hemiparasitic genus Cassytha (Lauraceae) and its host interactions: A review. Front. Plant Sci. 2022, 13, 864110. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.R.; Moore, P.G.P.; Twyford, A.D. Performance of generalist hemiparasitic Euphrasia across a phylogenetically diverse host spectrum. New Phytol. 2021, 232, 2165–2174. [Google Scholar] [CrossRef]
- Matthies, D. Interactions between a root hemiparasite and 27 different hosts: Growth, biomass allocation and plant architecture. Perspect. Plant Ecol. 2017, 24, 118–137. [Google Scholar] [CrossRef]
- Tesitel, J.; Tesitelova, T.; Fisher, J.P.; Leps, J.; Cameron, D.D. Integrating ecology and physiology of root-hemiparasitic interaction: Interactive effects of abiotic resources shape the interplay between parasitism and autotrophy. New Phytol. 2015, 205, 350–360. [Google Scholar] [CrossRef]
- Bize, P.; Jeanneret, C.; Klopfenstein, A.; Roulin, A. What makes a host profitable? Parasites balance host nutritive resources against immunity. Am. Nat. 2008, 171, 107–118. [Google Scholar] [CrossRef]
- Rowntree, J.K.; Cameron, D.D.; Preziosi, R.F. Genetic variation changes the interactions between the parasitic plant-ecosystem engineer Rhinanthus and its hosts. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1380–1388. [Google Scholar] [CrossRef] [PubMed]
- Hautier, Y.; Hector, A.; Vojtech, E.; Purves, D.; Turnbull, L.A. Modelling the growth of parasitic plants. J. Ecol. 2010, 98, 857–866. [Google Scholar] [CrossRef]
- Bickford, C.P.; Kolb, T.E.; Geils, B.W. Host physiological condition regulates parasitic plant performance: Arceuthobium vaginatum subsp. cryptopodum on Pinus ponderosa. Oecologia 2005, 146, 179–189. [Google Scholar] [CrossRef]
- Adler, L.S. Alkaloid uptake increases fitness in a hemiparasitic plant via reduced herbivory and increased pollination. Am. Nat. 2000, 156, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Matthies, D. Interactions between the root hemiparasite Melampyrum arvense and mixtures of host plants: Heterotrophic benefit and parasite-mediated competition. Oikos 1996, 75, 118–124. [Google Scholar] [CrossRef]
- Garnier, E. Growth analysis of congeneric annual and perennial grass species. J. Ecol. 1992, 80, 665–675. [Google Scholar] [CrossRef]
- Phoenix, G.K.; Press, M.C. Linking physiological traits to impacts on community structure and function: The role of root hemiparasitic Orobanchaceae (ex-Scrophulariaceae). J. Ecol. 2005, 93, 67–78. [Google Scholar] [CrossRef]
- Tennakoon, K.U.; Pate, J.S.; Arthur, D. Ecophysiological aspects of the woody root hemiparasite Santalum acuminatum (R Br) A DC and its common hosts in south western Australia. Ann. Bot. 1997, 80, 245–256. [Google Scholar] [CrossRef]
- Le, Q.V.; Tennakoon, K.U.; Metali, F.; Lim, L.B.L.; Bolin, J.F. Ecophysiological responses of mistletoe Dendrophthoe curvata (Loranthaceae) to varying environmental parameters. J. Trop. For. Sci. 2016, 28, 59–67. [Google Scholar]
- Glatzel, G.; Geils, B.W. Mistletoe ecophysiology: Host–parasite interactions. Botany 2009, 87, 10–15. [Google Scholar] [CrossRef]
- Furuhashi, T.; Furuhashi, K.; Weckwerth, W. The parasitic mechanism of the holostemparasitic plant Cuscuta. J. Plant Interact. 2011, 6, 207–219. [Google Scholar] [CrossRef]
- Weber, J. A taxonomic revision of Cassytha (Lauraceae) in Australia. J. Adel. Bot. Gard. 1981, 3, 187–262. [Google Scholar]
- Press, M.C.; Phoenix, G.K. Impacts of parasitic plants on natural communities. New Phytol. 2005, 166, 737–751. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Smith, R.S.; Shiel, R.S.; Peacock, S.; Simkin, J.M.; Quirk, H.; Hobbs, P.J. Parasitic plants indirectly regulate below-ground properties in grassland ecosystems. Nature 2006, 439, 969–972. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, M.J.; Dick, J.T.A.; Dunn, A.M. Diverse effects of parasites in ecosystems: Linking interdependent processes. Front. Ecol. Environ. 2012, 10, 186–194. [Google Scholar] [CrossRef]
- Tesitel, J.; Li, A.R.; Knotkova, K.; McLellan, R.; Bandaranayake, P.C.G.; Watson, D.M. The bright side of parasitic plants: What are they good for? Plant Physiol. 2021, 185, 1309–1324. [Google Scholar] [CrossRef]
- Adamu, A.A.; Garba, F.N.; Ahmed, T.M.; Abubakar, A. Pharmacognostic studies and elemental analysis of Cassytha filiformis Linn. J. Pharmacogn. Phytother. 2017, 9, 131–137. [Google Scholar] [CrossRef]
- Nelson, S.C. Cassytha filiformis. In Plant Disease PD-42; University of Hawaii: Honolulu, HI, USA, 2008; pp. 1–10. [Google Scholar]
- Nickrent, D.L. Plantas parásitas en el mundo. Capitulo 2. In Plantas Parásitas de la Península Ibérica e Islas Baleares, López-Sáez, J.A., Catalán, P., Sáez, L., Eds.; Mundi-Prensa Libros, S.A.: Madrid, Spain, 2002; pp. 7–27. [Google Scholar]
- Werth, C.R.; Pusateri, W.P.; Eshbaugh, W.H.; Wilson, T.K. Field observations on the natural history of Cassytha filiformis L. (Lauraceae) in the Bahamas. In The 2nd International Symposium on Parasitic Weeds; Musselman, L.J., Worsham, A.D., Eplee, R.E., Eds.; North Carolina State University College: Raleigh, NC, USA, 1979; pp. 94–102. [Google Scholar]
- Thorogood, C.J.; Rumsey, F.J.; Hiscock, S.J. Host-specific races in the holoparasitic angiosperm Orobanche minor: Implications for speciation in parasitic plants. Ann. Bot. 2009, 103, 1005–1014. [Google Scholar] [CrossRef]
- Cirocco, R.M.; Facelli, J.M.; Watling, J.R. High water availability increases the negative impact of a native hemiparasite on its non-native host. J. Exp. Bot. 2016, 67, 1567–1575. [Google Scholar] [CrossRef]
- Cirocco, R.M.; Facelli, J.M.; Watling, J.R. Does nitrogen affect the interaction between a native hemiparasite and its native or introduced leguminous hosts? New Phytol. 2017, 213, 812–821. [Google Scholar] [CrossRef]
- Prider, J.; Watling, J.; Facelli, J.M. Impacts of a native parasitic plant on an introduced and a native host species: Implications for the control of an invasive weed. Ann. Bot. 2009, 103, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Furuhashi, K.; Iwase, K.; Furuhashi, T. Role of light and plant hormones in stem parasitic plant (Cuscuta and Cassytha) twining and haustoria induction. Photochem. Photobiol. 2021, 97, 1054–1062. [Google Scholar] [CrossRef]
- Li, H.W.; Li, J.; Huang, P.H.; Wei, F.N.; Cui, H.B.; van der Werff, H. Lauraceae. In Flora of China, Calycanthaceae–Schisandraceae; Wu, Z.Y., Raven, P.H., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MI, USA, 2008; Volume 7, pp. 102–254. [Google Scholar]
- Li, Q.; Yao, D.; Cai, J.; Huang, L. A field survey of Cassytha filiformis host range. Guangxi Plant Prot. 1992, 4, 21–24. [Google Scholar]
- Huang, Z.J. Some observations on the host of Cassytha filiformis and the exploration of its harmfulness. J. Sci. Res. Repert. Guangxi Agric. Coll. 1957, 54–60. [Google Scholar]
- Shao, S.Z. Records of the detriment of Cassytha filiformis on Pinus massoniana. For. Pest Commun. 1988, 4, 3. [Google Scholar]
- Mahadevan, N.; Jayasuriya, K. Water-impermeable fruits of the parasitic angiosperm Cassytha filiformis (Lauraceae): Confirmation of physical dormancy in Magnoliidae and evolutionary considerations. Aust. J. Bot. 2013, 61, 322–329. [Google Scholar] [CrossRef]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Qian, H.V. PhyloMaker2: An updated and enlarged R package that can generate very large phylogenies for vascular plants. Plant Divers. 2022, 44, 335–339. [Google Scholar] [CrossRef]
- The Angiosperm Phylogeny Group. An update of the angiosperm phylogeny group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 12385. [Google Scholar] [CrossRef]
- Parra-Tabla, V.; Tun-Garrido, J.; García-Franco, J.; Martínez, M. The recent expansion of the invasive hemiparasitic plant Cassytha filiformis and the reciprocal effect with its main hosts. Res. Square 2023. [Google Scholar] [CrossRef]
- Buriyo, A.S.; Kasuga, L.; Moshi, H.N.; Nene, W.A. Ecological distribution and abundance of the parasitic weed, Cassytha filiformis L. (Lauraceae) in major cashew, Anacardium occidentale L. growing regions in Tanzania. Int. J. Basic Appl. Sci. 2015, 5, 109–116. [Google Scholar]
- Zhang, C.; Ma, H.; Sanchez-Puerta, M.V.; Li, L.; Xiao, J.; Liu, Z.; Ci, X.; Li, J. Horizontal gene transfer has impacted cox1 gene evolution in Cassytha filiformis. J. Mol. Evol. 2020, 88, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Rajanna, L.; Shivamurthy, G.R. Occurrence of graniferous tracheary elements in the haustorium of Cassytha filiformis Linn., a stem parasite of Lauraceae. Taiwania 2001, 46, 40–48. [Google Scholar]
- Kokubugata, G.; Nakamura, K.; Forster, P.I.; Wilson, G.W.; Holland, A.E.; Hirayama, Y.; Yokota, M. Cassytha pubescens and C. glabella (Lauraceae) are not disjunctly distributed between Australia and the Ryukyu Archipelago of Japan—Evidence from morphological and molecular data. Aust. Syst. Bot. 2012, 25, 364–373. [Google Scholar] [CrossRef]
- Costea, M.; ElMiari, H.; Farag, R.; Fleet, C.; Stefanović, S. Cuscuta sect. Californicae (Convolvulaceae) revisited:‘cryptic’ speciation and host range differentiation. Syst. Bot. 2020, 45, 638–651. [Google Scholar] [CrossRef]
- Facelli, E.; Wynn, N.; Tsang, H.T.; Watling, J.R.; Facelli, J.M. Defence responses of native and invasive plants to the native generalist vine parasite Cassytha pubescens—Anatomical and functional studies. Aust. J. Bot. 2020, 68, 300. [Google Scholar] [CrossRef]
- Kelly, C.K. Resource choice in Cuscuta europaea. Proc. Natl. Acad. Sci. USA 1992, 89, 12194–12197. [Google Scholar] [CrossRef]
- Runyon, J.B.; Mescher, M.C.; De Moraes, C.M. Plant defenses against parasitic plants show similarities to those induced by herbivores and pathogens. Plant Signal. Behav. 2010, 5, 929–931. [Google Scholar] [CrossRef]
- Machado, M.A.; Zetsche, K. A structural, functional and molecular analysis of plastids of the holoparasites Cuscuta reflexa and Cuscuta europaea. Planta 1990, 181, 91–96. [Google Scholar] [CrossRef]
- Liu, Z.-F.; Ma, H.; Ci, X.-Q.; Li, L.; Song, Y.; Liu, B.; Li, H.-W.; Wang, S.-L.; Qu, X.-J.; Hu, J.-L.; et al. Can plastid genome sequencing be used for species identification in Lauraceae? Bot. J. Linn. Soc. 2021, 197, 1–14. [Google Scholar] [CrossRef]
- Schonenberger, J.; von Balthazar, M. Asterids. Bot. J. Linn. Soc. 2013, 173, 321–324. [Google Scholar] [CrossRef][Green Version]
- Fay, M.F. Rosids. Bot. J. Linn. Soc. 2013, 172, 399–403. [Google Scholar] [CrossRef][Green Version]
- Soltis, D.E.; Mort, M.E.; Latvis, M.; Mavrodiev, E.V.; O’Meara, B.C.; Soltis, P.S.; Burleigh, J.G.; Rubio de Casas, R. Phylogenetic relationships and character evolution analysis of Saxifragales using a supermatrix approach. Am. J. Bot. 2013, 100, 916–929. [Google Scholar] [CrossRef]
- Su, H.-J.; Hu, J.-M.; Anderson, F.E.; Der, J.P.; Nickrent, D.L. Phylogenetic relationships of Santalales with insights into the origins of holoparasitic Balanophoraceae. Taxon 2015, 64, 491–506. [Google Scholar] [CrossRef]
- Tesitel, J.; Fibich, P.; de Bello, F.; Chytry, M.; Leps, J. Habitats and ecological niches of root-hemiparasitic plants: An assessment based on a large database of vegetation plots. Preslia 2015, 87, 87–108. [Google Scholar]
- Birschwilks, M.; Haupt, S.; Hofius, D.; Neumann, S. Transfer of phloem-mobile substances from the host plants to the holoparasite Cuscuta sp. J. Exp. Bot. 2006, 57, 911–921. [Google Scholar] [CrossRef]
- Kaminska, M.; Korbin, M. Graft and dodder transmission of phytoplasma affecting lily to experimental hosts. Acta Physiol. Plant. 1999, 21, 21–26. [Google Scholar] [CrossRef]
- Lanini, W.T.; Kogan, M. Biology and management of Cuscuta in crops. Cienc. Investig. Agrar. 2005, 32, 127–141. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.-F.; Ci, X.-Q.; Zhang, S.-F.; Zhang, X.-Y.; Zhang, X.; Dong, L.-N.; Conran, J.G.; Li, J. Diverse Host Spectrum and the Parasitic Process in the Pantropical Hemiparasite Cassytha filiformis L. (Lauraceae) in China. Diversity 2023, 15, 492. https://doi.org/10.3390/d15040492
Liu Z-F, Ci X-Q, Zhang S-F, Zhang X-Y, Zhang X, Dong L-N, Conran JG, Li J. Diverse Host Spectrum and the Parasitic Process in the Pantropical Hemiparasite Cassytha filiformis L. (Lauraceae) in China. Diversity. 2023; 15(4):492. https://doi.org/10.3390/d15040492
Chicago/Turabian StyleLiu, Zhi-Fang, Xiu-Qin Ci, Shi-Fang Zhang, Xiao-Yan Zhang, Xue Zhang, Li-Na Dong, John G. Conran, and Jie Li. 2023. "Diverse Host Spectrum and the Parasitic Process in the Pantropical Hemiparasite Cassytha filiformis L. (Lauraceae) in China" Diversity 15, no. 4: 492. https://doi.org/10.3390/d15040492
APA StyleLiu, Z.-F., Ci, X.-Q., Zhang, S.-F., Zhang, X.-Y., Zhang, X., Dong, L.-N., Conran, J. G., & Li, J. (2023). Diverse Host Spectrum and the Parasitic Process in the Pantropical Hemiparasite Cassytha filiformis L. (Lauraceae) in China. Diversity, 15(4), 492. https://doi.org/10.3390/d15040492