

Climate Change Influence on the Potential Distribution of Some Cavity-Nesting Bees (Hymenoptera: Megachilidae)


Abstract
1. Introduction
2. Materials and Methods
2.1. Occurrence Data
2.2. Environmental Variables
2.3. Predictive Modeling
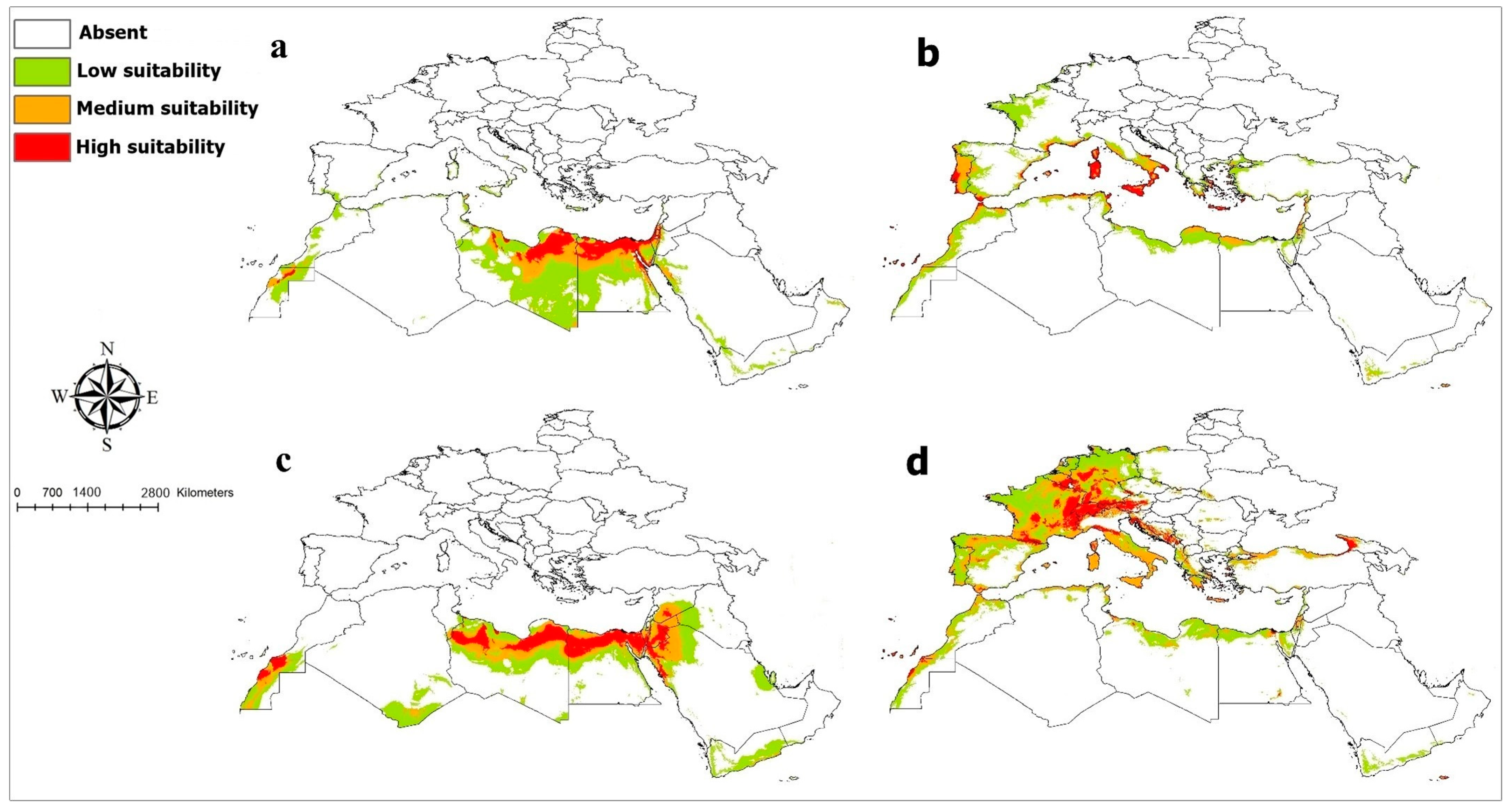
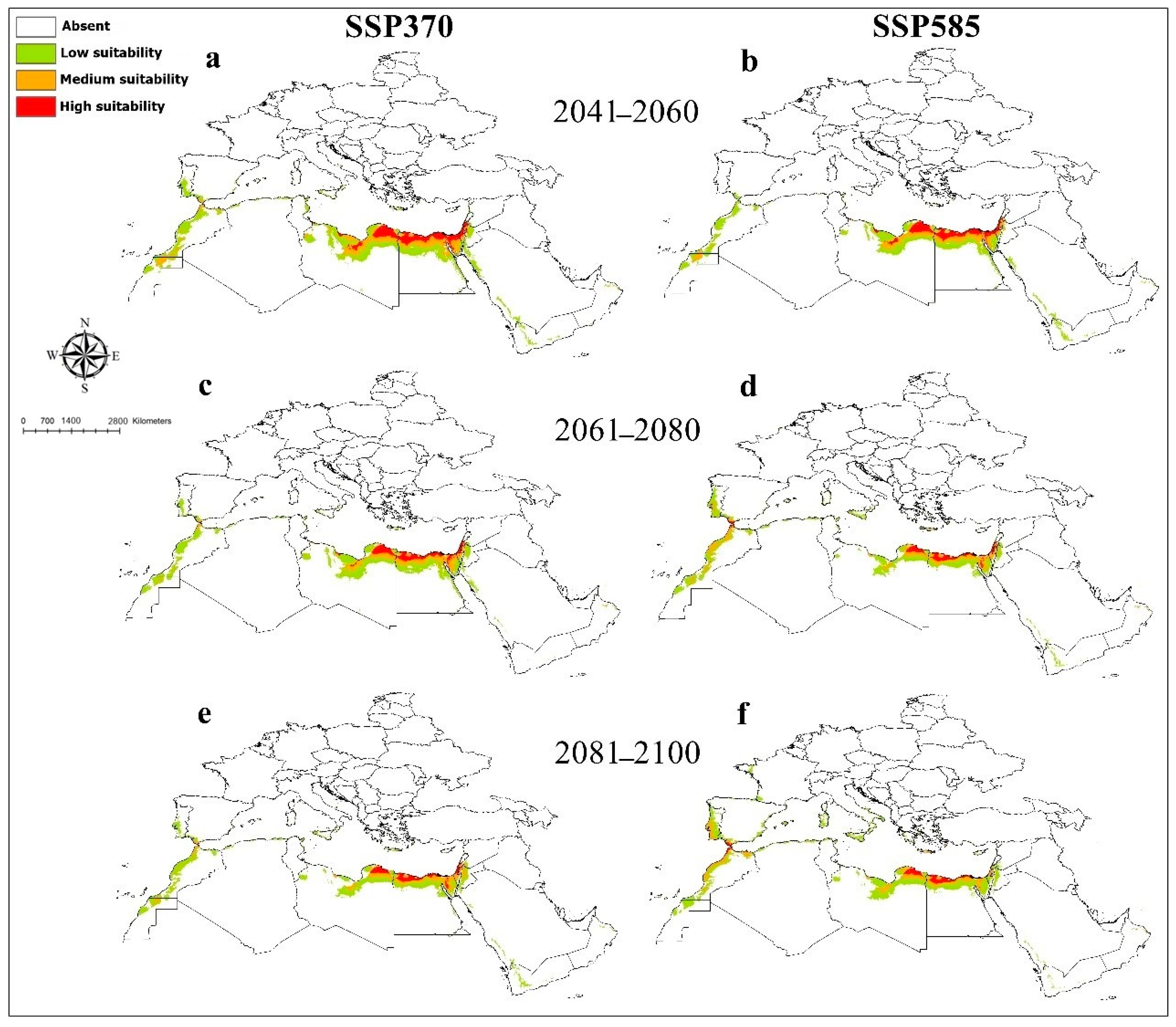
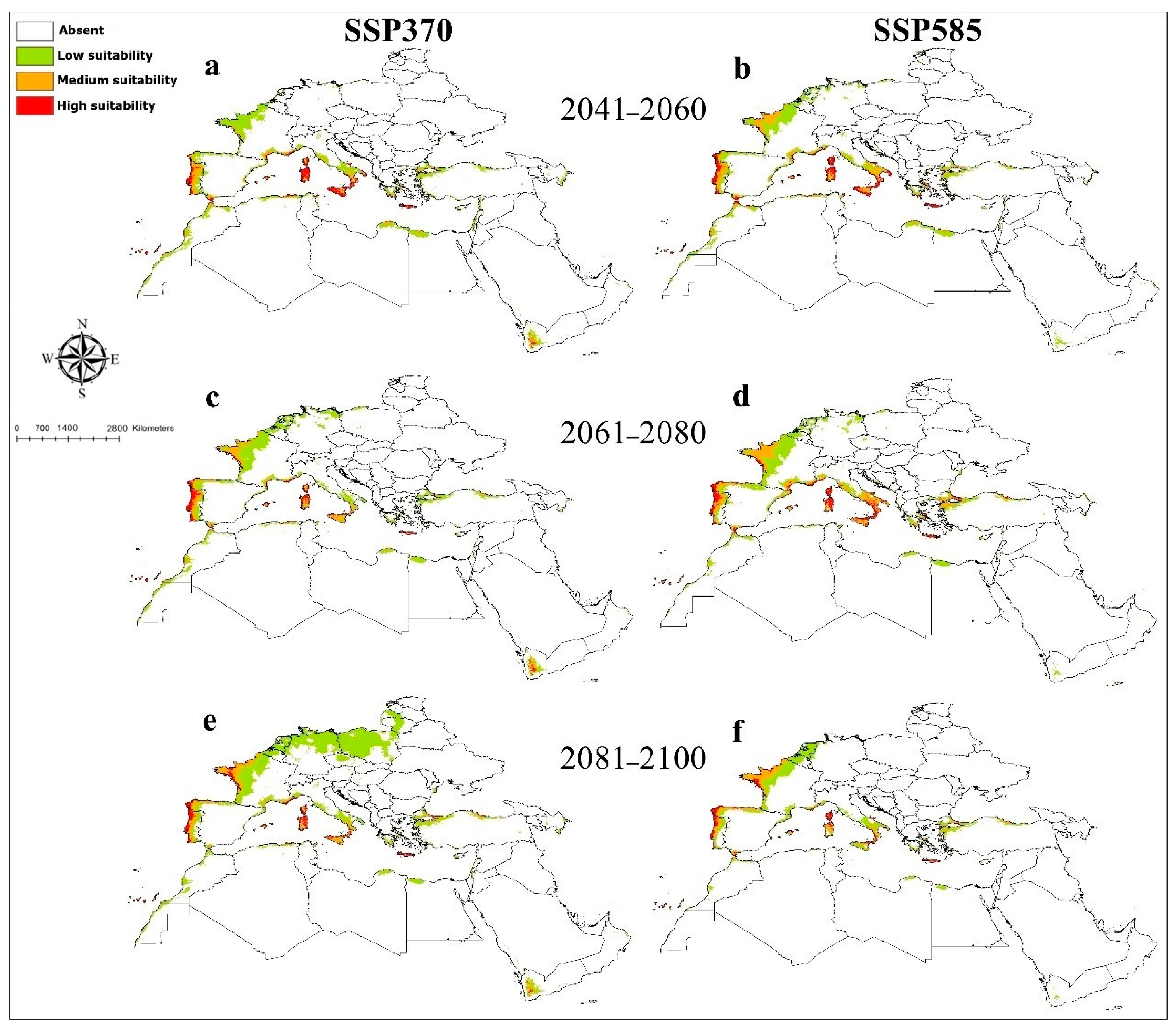
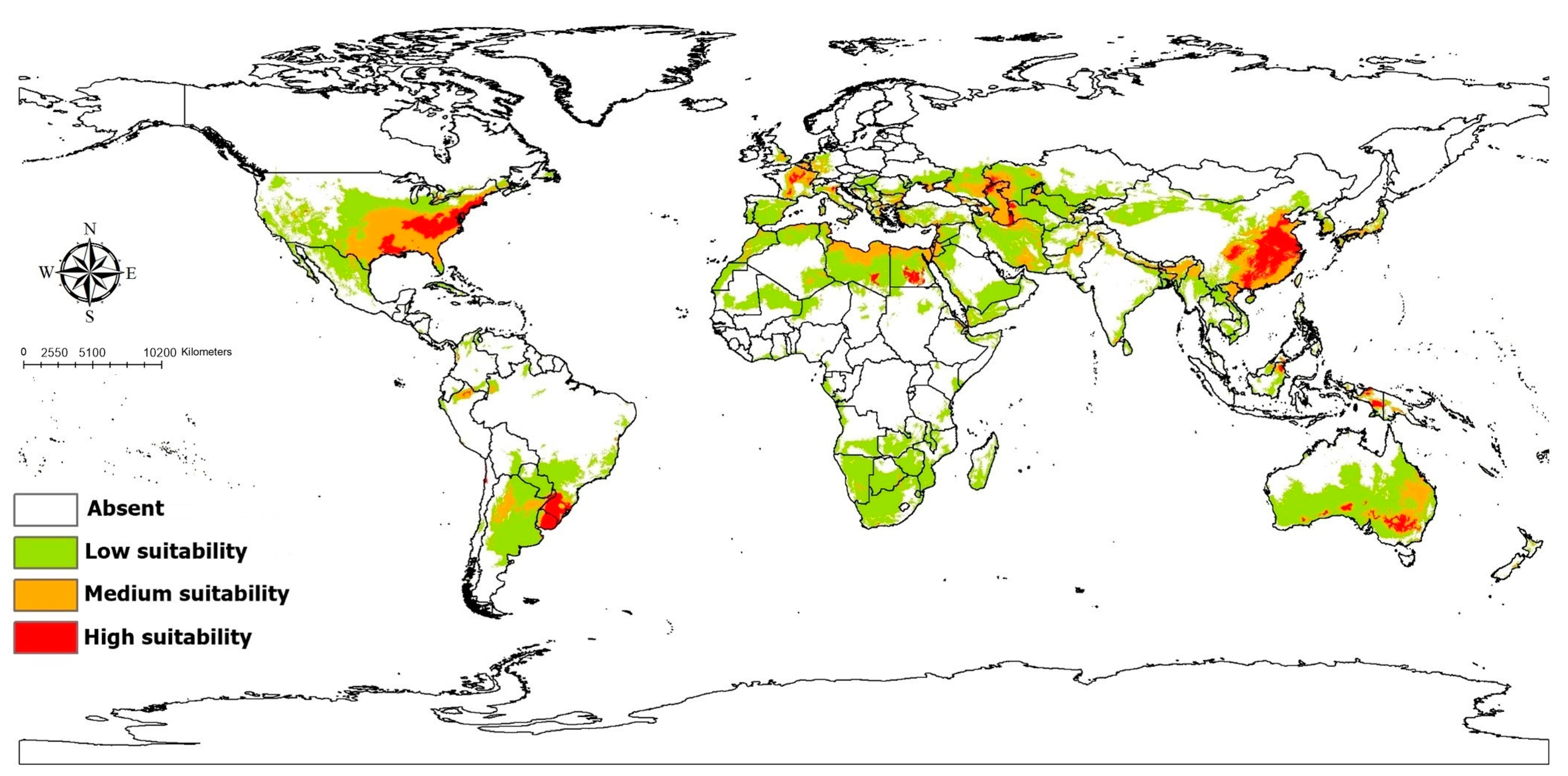
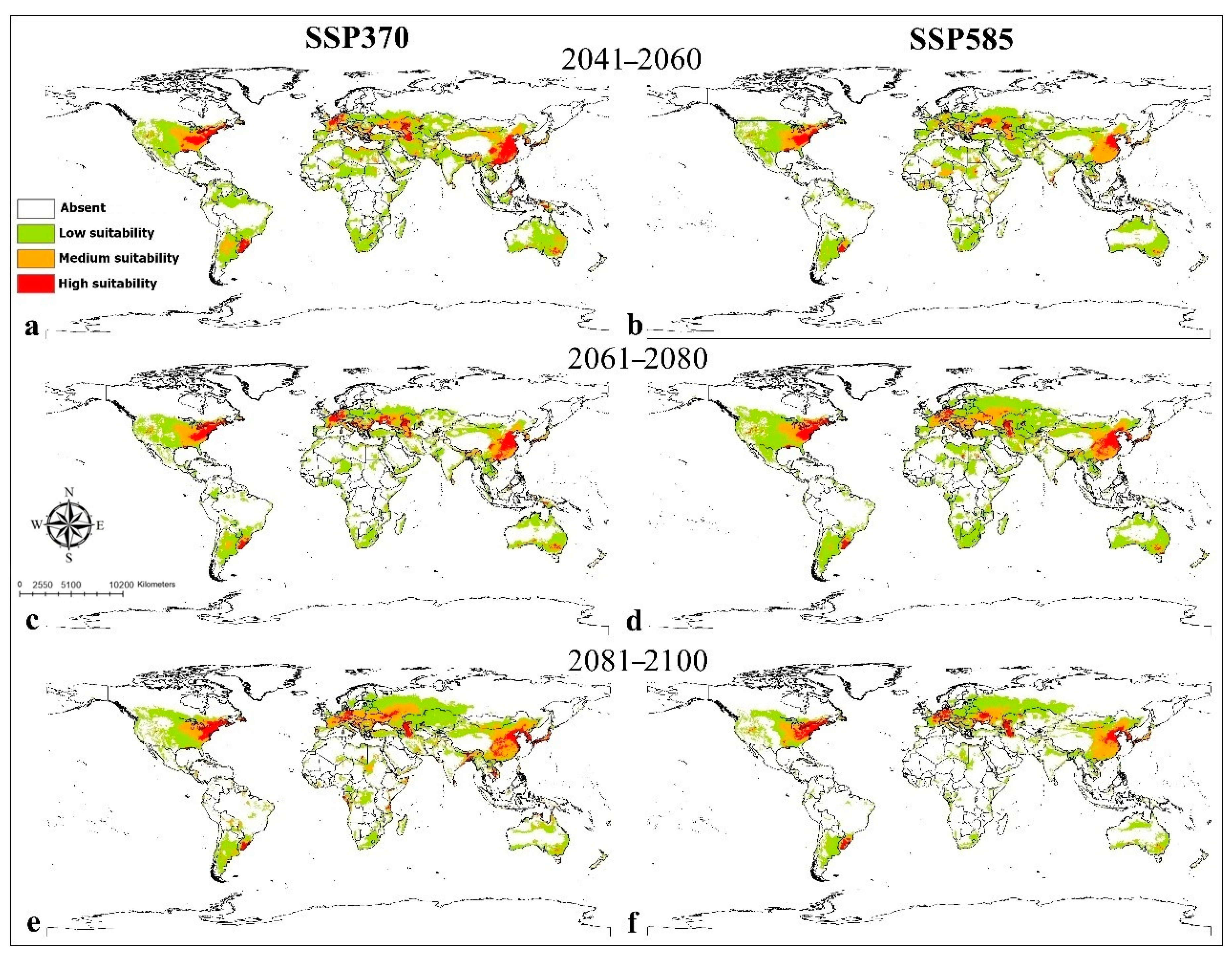
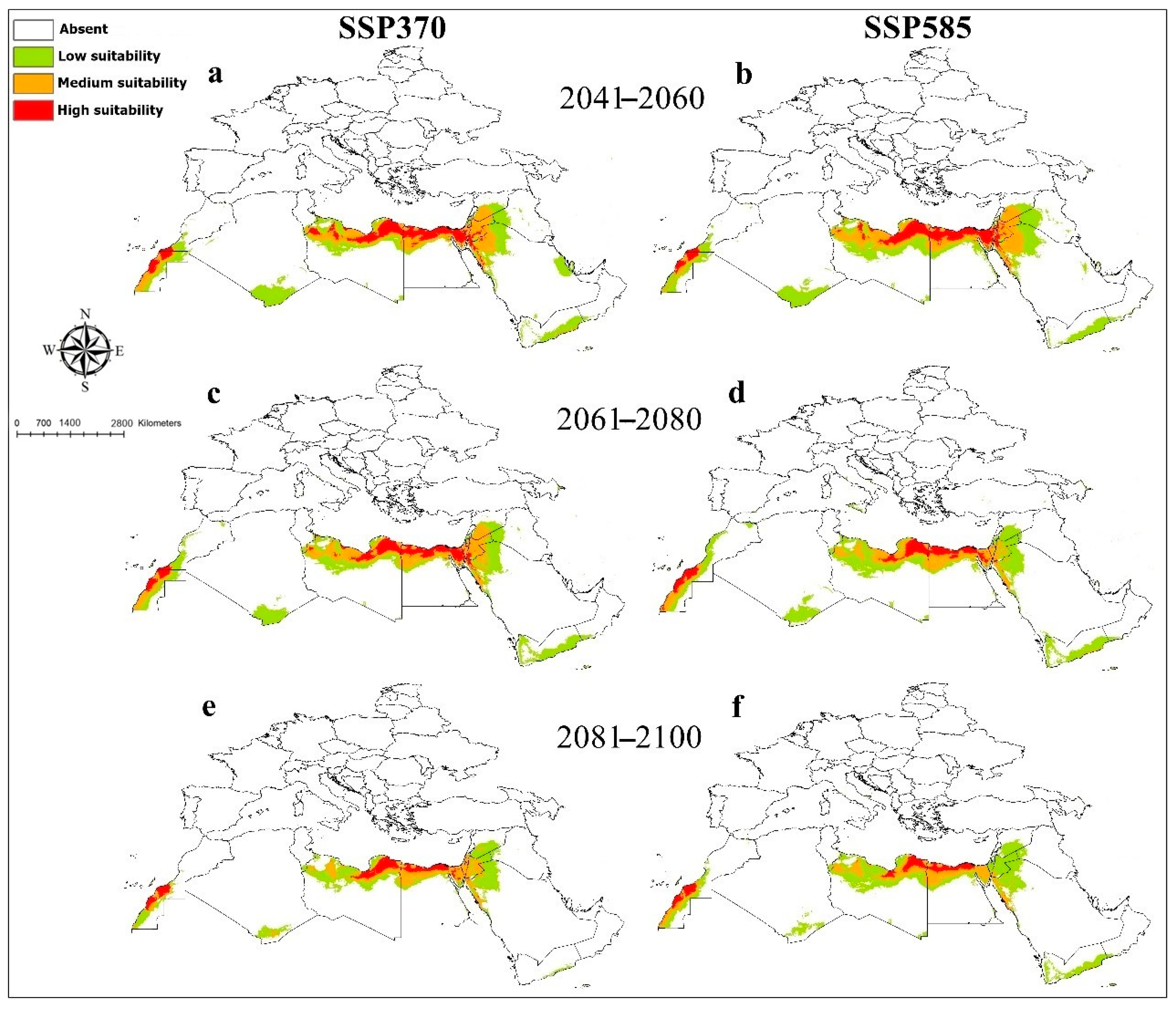
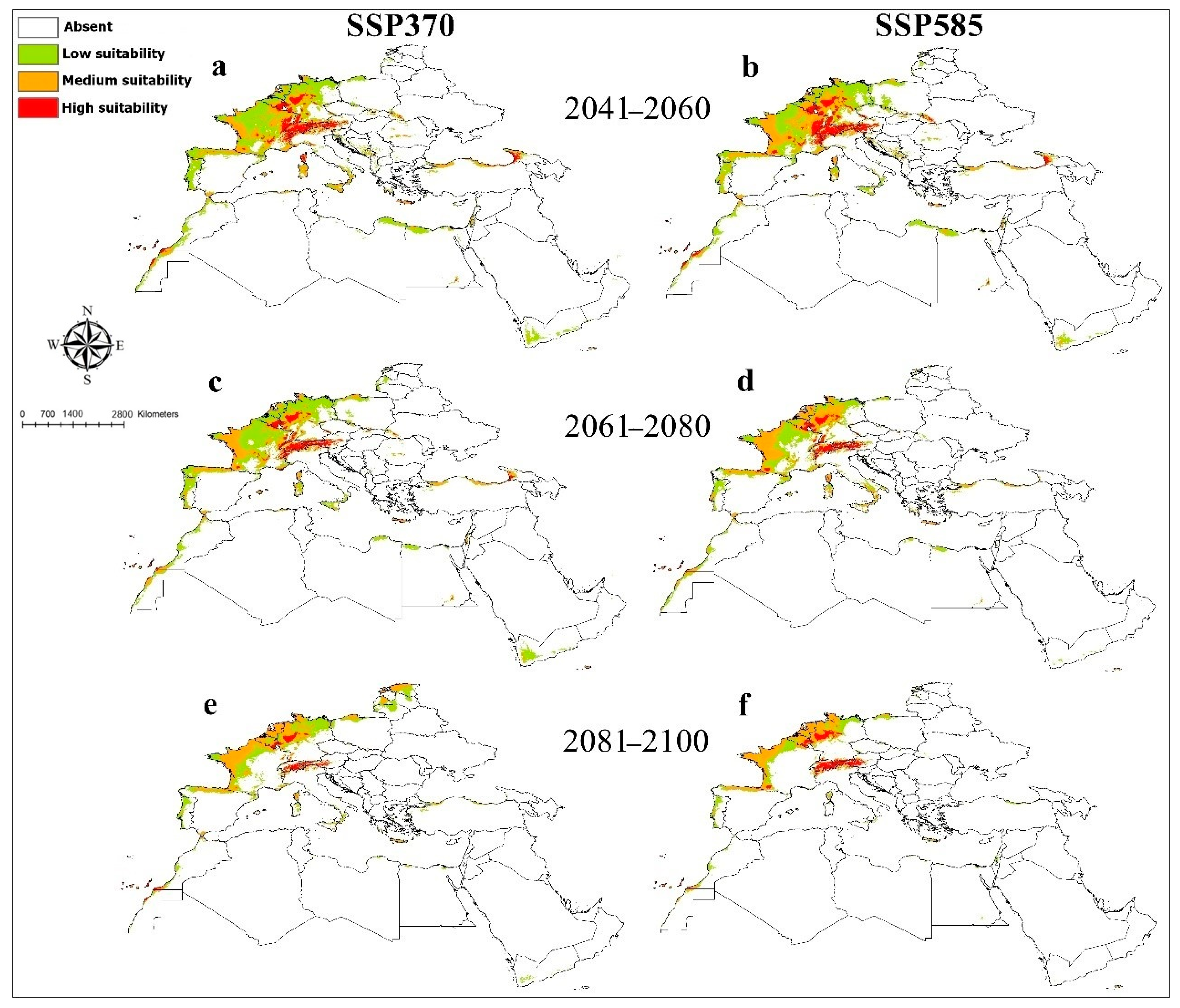
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Michener, C.D. The Bees of the World, 2nd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2007; p. 953. [Google Scholar]
- Freitas, B.M.; Paxton, R.J. A comparison of two pollinators: The introduced honey bee Apis mellifera and an indigenous bee Centris tarsata on cashew Anacardium occidentale in its native range of NE Brazil. J. Appl. Ecol. 1998, 35, 109–121. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef] [PubMed]
- Greenleaf, S.S.; Kremen, C. Wild bees enhance honey bees’ pollination of hybrid sunflower. Proc. Natl. Acad. Sci. USA 2006, 103, 13890–13895. [Google Scholar] [CrossRef] [PubMed]
- Vanbergen, A.J.; The Insect Pollinator Initiative. Threats to an ecosystem service: Pressures on pollinators. Front. Ecol. Environ. 2013, 11, 251–259. [Google Scholar] [CrossRef]
- Shebl, M.A.; Ben Abdelkader, F.; Bendifallah, L.; Benachour, K.; Bataw, A.A.; Bufliga, E.M.; Osman, M.A.; Kamel, S.M. The melittology research in northern Africa and the Middle East: Past and present situations. J. Basic Appl. Zool. 2021, 82, 18. [Google Scholar] [CrossRef]
- Cane, J.H.; Tepedino, V.J. Causes and extent of declines among native North American invertebrate pollinators: Detection, evidence, and consequences. Conserv. Ecol. 2001, 5, 1. [Google Scholar] [CrossRef]
- Kevan, P.G.; Phillips, T.P. The economic impacts of pollinator declines: An approach to assessing the consequences. Conserv. Ecol. 2001, 5, 8. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Ollerton, J. Pollinator diversity: Distribution, ecological function, and conservation. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 353–376. [Google Scholar] [CrossRef]
- Brading, P.; El-Gabbas, A.; Zalat, S.; Gilbert, F. Biodiversity economics: The value of pollination services to Egypt. Egypt. J. Biol. 2009, 11, 46–51. [Google Scholar]
- Shebl, M.; Kamel, S.; Mahfouz, H. Bee fauna (Apoidea: Hymenoptera) of the Suez Canal region, Egypt. J. Apic. Sci. 2013, 57, 33–44. [Google Scholar] [CrossRef]
- Osman, M.A.M.; Shebl, M.A. Vulnerability of crop pollination ecosystem services to climate change. In Climate Change Impacts on Agriculture and Food Security in Egypt: Land and Water Resources—Smart Farming—Livestock, Fishery, and Aquaculture; Ewis Omran, E.-S., Negm, A.M., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 223–247. [Google Scholar]
- Miranda, E.A.; Carvalho, A.F.; de Jesus Gomes-Miranda, J.; de Souza, C.R.; Costa, M.A. Priority areas for conservation of orchid bees (Apidae, Euglossini) in the Atlantic Forest. J. Insect Conserv. 2019, 23, 613–621. [Google Scholar] [CrossRef]
- Giannini, T.C.; Acosta, A.L.; da Silva, C.I.; de Oliveira, P.E.A.M.; Imperatriz-Fonseca, V.L.; Saraiva, A.M. Identifying the areas to preserve passion fruit pollination service in Brazilian Tropical Savannas under climate change. Agric. Ecosyst. Environ. 2013, 171, 39–46. [Google Scholar] [CrossRef]
- Carvalho, A.F.; Del Lama, M.A. Predicting priority areas for conservation from historical climate modelling: Stingless bees from Atlantic Forest hotspot as a case study. J. Insect Conserv. 2015, 19, 581–587. [Google Scholar] [CrossRef]
- Nasser, M.; El-Hawagry, M.; Okely, M. Environmental niche modeling for some species of the genus Anthrax Scopoli (Diptera: Bombyliidae) in Egypt, with special notes on St. Catherine protected area as a suitable habitat. J. Insect Conserv. 2019, 23, 831–841. [Google Scholar] [CrossRef]
- Hu, X.; Liu, J.; Ding, G.; Naeem, M.; Li, J.; Ma, F.; Huang, J.; An, J. An evaluation of habitat uses and their implications for the conservation of the Chinese bumblebee Bombus pyrosoma (Hymenoptera: Apidae). Front. Ecol. Evol. 2021, 9, 667949. [Google Scholar] [CrossRef]
- Nascimento, A.C.; Montalva, J.; Ascher, J.S.; Engel, M.S.; Silva, D.P. Current and future distributions of a native Andean bumble bee. J. Insect Conserv. 2022, 26, 559–569. [Google Scholar] [CrossRef]
- Nemésio, A.; Silva, D.P.; Nabout, J.C.; Varela, S. Effects of climate change and habitat loss on a forest-dependent bee species in a tropical fragmented landscape. Insect Conserv. Divers. 2016, 9, 149–160. [Google Scholar] [CrossRef]
- Gonzalez, V.H.; Cobos, M.E.; Jaramillo, J.; Ospina, R. Climate change will reduce the potential distribution ranges of Colombia’s most valuable pollinators. Perspect. Ecol. Conserv. 2021, 19, 195–206. [Google Scholar] [CrossRef]
- Rahimi, E.; Barghjelveh, S.; Dong, P. Estimating potential range shift of some wild bees in response to climate change scenarios in northwestern regions of Iran. J. Ecol. Environ. 2021, 45, 14. [Google Scholar] [CrossRef]
- Syfert, M.M.; Smith, M.J.; Coomes, D.A. The effects of sampling bias and model complexity on the predictive performance of MaxEnt species distribution models. PLoS ONE 2013, 8, e55158. [Google Scholar] [CrossRef]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef] [PubMed]
- Okely, M.; Al-Khalaf, A.A. Predicting the potential distribution of the cattle fever tick Rhipicephalus annulatus (Acari: Ixodidae) using ecological niche modeling. Parasitol. Res. 2022, 121, 3467–3476. [Google Scholar] [CrossRef] [PubMed]
- Beyer, H. Hawth’s Analysis Tools for ArcGIS. 2004. Available online: http://www.spatialecology.com/htools (accessed on 7 April 2019).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Hortal, J.; Roura-Pascual, N.; Sanders, N.J.; Rahbek, C. Understanding (insect) species distributions across spatial scales. Ecography 2010, 33, 51–53. [Google Scholar] [CrossRef]
- Escobar, L.E.; Lira-Noriega, A.; Medina-Vogel, G.; Peterson, A.T. Potential for spread of the white-nose fungus (Pseudogymnoascus destructans) in the Americas: Use of Maxent and NicheA to assure strict model transference. Geospat. Health 2014, 9, 221–229. [Google Scholar] [CrossRef]
- Datta, A.; Schweiger, O.; Kühn, I. Origin of Climatic Data Can Determine the Transferability of Species Distribution Models. Neobiota 2020, 59, 61–76. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Alahmed, A.M.; Naeem, M.; Kheir, S.M.; Sallam, M.F. Ecological distribution modeling of two malaria mosquito vectors using geographical information system in Al-Baha Province, Kingdom of Saudi Arabia. Pak. J. Zool. 2015, 47, 1797–1806. [Google Scholar]
- Di Febbraro, M.; Bosso, L.; Fasola, M.; Santicchia, F.; Aloise, G.; Lioy, S.; Tricarico, E.; Ruggieri, L.; Bovero, S.; Mori, E.; et al. Different facets of the same niche: Integrating citizen science and scientific survey data to predict biological invasion risk under multiple global change drivers. Glob. Change Biol. 2023, 29, 5509–5523. [Google Scholar] [CrossRef]
- Xu, Y.; Zhou, J.; Liu, T.; Liu, P.; Wu, Y.; Lai, Z.; Gu, J.; Chen, X.-G. Assessing the risk of spread of Zika virus under current and future climate scenarios. Biosaf. Health 2022, 4, 193–204. [Google Scholar] [CrossRef]
- Bosso, L.; Luchi, N.; Maresi, G.; Cristinzio, G.; Smeraldo, S.; Russo, D. Predicting current and future disease outbreaks of Diplodia sapinea shoot blight in Italy: Species distribution models as a tool for forest management planning. For. Ecol. Manag. 2017, 400, 655–664. [Google Scholar]
- Salvacion, A.R. Soil erosion modeling under future climate change: A case study on Marinduque Island, Philippines. In Water, Land, and Forest Susceptibility and Sustainability; Elsevier: Amsterdam, The Netherlands, 2023; pp. 381–398. [Google Scholar]
- Shao, M.; Wang, L.; Li, B.; Li, S.; Fan, J.; Li, C. Maxent Modeling for Identifying the Nature Reserve of Cistanche deserticola Ma under Effects of the Host (Haloxylon Bunge) Forest and Climate Changes in Xinjiang, China. Forests 2022, 13, 189. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar]
- Elith, J.H.; Graham, C.P.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Khanum, R.; Mumtaz, A.S.; Kumar, S. Predicting impacts of climate change on medicinal asclepiads of Pakistan using Maxent modeling. Acta Oecol. 2013, 49, 23–31. [Google Scholar] [CrossRef]
- Owens, H.L.; Campbell, L.P.; Dornak, L.L.; Saupe, E.E.; Barve, N.; Soberón, J.; Ingenloff, K.; Lira-Noriega, A.; Hensz, C.M.; Myers, C.E.; et al. Constraints on interpretation of ecological niche models by limited environmental ranges on calibration areas. Ecol. Model. 2013, 263, 10–18. [Google Scholar] [CrossRef]
- Okely, M.; Anan, R.; Gad-Allah, S.; Samy, A.M. Mapping the environmental suitability of etiological agent and tick vectors of Crimean-Congo hemorrhagic fever. Acta Trop. 2020, 203, 105319. [Google Scholar]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Nasser, M.; Okely, M.; Nasif, O.; Alharbi, S.; GadAllah, S.; Al-Obaid, S.; Enan, R.; Bala, M.; Al-Ashaal, S. Spatio-temporal analysis of Egyptian flower mantis Blepharopsis mendica (order: Mantodea), with notes of its future status under climate change. Saudi J. Biol. Sci. 2021, 28, 2049–2055. [Google Scholar] [CrossRef] [PubMed]
- Okely, M.; Nasser, M.; Enan, R.; GadAllah, S.; AlAshaal, S. Mantodea oasis of Palaearctic region: Biogeographical analysis of Mantodea in Egypt. Egypt. J. Biol. Pest. Control 2020, 30, 136. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jiménez-Valverde, A.; Real, R. AUC: A misleading measure of the performance of predictive distribution models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Peterson, A.T.; Papeş, M.; Soberón, J. Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecol. Model. 2008, 213, 63–72. [Google Scholar] [CrossRef]
- Martins, A.C.; Silva, D.P.; De Marco, P., Jr.; Melo, G.A.R. Species conservation under future climate change: The case of Bombus bellicosus, a potentially threatened South American bumblebee species. J. Insect Conserv. 2015, 19, 33–43. [Google Scholar] [CrossRef]
- Hinojosa-Díaz, I.A.; Alqarni, A.S.; Lira-Noriega, A.; Engel, M.S. Ecological niche modeling of the rare bee Promelitta alboclypeata reveals possible cryptic differentiation across northern Africa and Arabia (Hymenoptera: Melittidae). Apidologie 2016, 47, 509–514. [Google Scholar] [CrossRef]
- Yurrita, C.L.; Ortega-Huerta, M.A.; Ayala, R. Distributional analysis of Melipona stingless bees (Apidae: Meliponini) in Central America and Mexico: Setting baseline information for their conservation. Apidologie 2017, 48, 247–258. [Google Scholar] [CrossRef]
- Parichehreh, S.; Tahmasbi, G.; Sarafrazi, A.; Tajabadi, N.; Solhjouy-Fard, S. Distribution modeling of Apis florea Fabricius (Hymenoptera, Apidae) in different climates of Iran. J. Apic. Res. 2022, 61, 469–480. [Google Scholar] [CrossRef]
- Goulson, D. Conserving wild bees for crop pollination. J. Food Agric. Environ. 2003, 1, 142–144. [Google Scholar]
- Kasina, J.M.; Mburu, J.; Kraemer, M.; Holm-Mueller, K. Economic benefit of crop pollination by bees: A case of Kakamega small-holder farming in western Kenya. J. Econ. Entomol. 2009, 102, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, A.L.; Youngsteadt, E.; Frank, S.D. Wild bee abundance declines with urban warming, regardless of floral density. Urban Ecosyst. 2018, 21, 419–428. [Google Scholar] [CrossRef]
- Kerr, J.T.; Pindar, A.; Galpern, P.; Packer, L.; Potts, S.G.; Roberts, S.M.; Rasmont, P.; Schweiger, O.; Colla, S.R.; Richardson, L.L.; et al. Climate change impacts on bumblebees converge across continents. Science 2015, 349, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Winfree, R.; Bartomeus, I.; Cariveau, D.P. Native pollinators in anthropogenic habitats. Annu. Rev. Ecol. Evol. Syst. 2011, 42, 1–22. [Google Scholar] [CrossRef]
- Fliszkiewicz, M.; Giejdasz, K.; Wasielewski, O.; Krishnan, N. Influence of winter temperature and simulated climate change on body mass and fat body depletion during diapause in adults of the solitary bee, Osmia rufa (Hymenoptera: Megachilidae). Environ. Entomol. 2012, 41, 1621–1630. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.; Sgolastra, F.; Kemp, W.P. Timing of eclosion affects diapause development, fat body consumption and longevity in Osmia lignaria, a univoltine, adult-wintering solitary bee. J. Insect Physiol. 2010, 56, 1949–1957. [Google Scholar] [CrossRef] [PubMed]
- Rozen, J.G.; Kamel, S.M. Hospicidal behavior of the cleptoparasitic bee Coelioxys (Allocoelioxys) coturnix, including descriptions of its larval instars (Hymenoptera: Megachilidae). Am. Mus. Novit. 2008, 3636, 1–15. [Google Scholar] [CrossRef]
- Alqarni, A.S.; Hannan, M.A.; Gonzalez, V.H.; Engel, M.S. Nesting biology of the leafcutting bee Megachile minutissima (Hymenoptera: Megachilidae) in central Saudi Arabia. Ann. Entomol. Soc. Am. 2014, 107, 635–640. [Google Scholar] [CrossRef]
- Faria, L.R.R.; Gonçalves, R.B. Abiotic correlates of bee diversity and composition along eastern Neotropics. Apidologie 2013, 44, 547–562. [Google Scholar] [CrossRef]
- Shebl, M.A.; Kamel, S.M.; Abu Hashesh, T.A.; Osman, M.A. The impact of using leafcutting bees (Megachilidae hymenoptera) with different fertilization treatments on alfalfa seed production. Rev. Cienc. Suelo Nutr. Veg. 2009, 9, 134–141. [Google Scholar] [CrossRef]
- Shebl, M.A.; Hassan, H.A.; Kamel, S.M.; Osman, M.A.M.; Engel, M.S. Biology of the mason bee Osmia latreillei (Hymenoptera: Megachilidae) under artificial nesting conditions in Egypt. J. Asia Pac. Entomol. 2018, 21, 754–759. [Google Scholar] [CrossRef]
- Kamel, S.M.; Osman, M.A.M.; Mahmoud, M.F.; Haggag, E.-S.I.; Aziz, A.R.; Shebl, M.A. Influence of temperature on breaking diapause, development and emergence of Megachile minutissima (Hymenoptera, Megachilidae). Vestn. Zool. 2019, 53, 245–254. [Google Scholar] [CrossRef][Green Version]
- Blackmore, L.M.; Goulson, D. Evaluating the effectiveness of wildflower seed mixes for boosting floral diversity and bumblebee and hoverfly abundance in urban areas. Insect Conserv. Divers. 2014, 7, 480–484. [Google Scholar] [CrossRef]
- Fischer, L.K.; Eichfeld, J.; Kowarik, I.; Buchholz, S. Disentangling urban habitat and matrix effects on wild bee species. PeerJ 2016, 4, e2729. [Google Scholar] [CrossRef]
- Xie, Z.; Shebl, M.A.; Pan, D.; Wang, J. Synergistically positive effects of brick walls and farmlands on Anthophora waltoni populations. Agric. For. Entomol. 2020, 22, 328–337. [Google Scholar] [CrossRef]
- Buonincontri, M.P.; Bosso, L.; Smeraldo, S.; Chiusano, M.L.; Pasta, S.; Di Pasquale, G. Shedding light on the effects of climate and anthropogenic pressures on the disappearance of Fagus sylvatica in the Italian lowlands: Evidence from archaeo-anthracology and spatial analyses. Sci. Total Environ. 2023, 877, 162893. [Google Scholar] [CrossRef]
- Forister, M.L.; Black, S.H.; Elphick, C.S.; Grames, E.M.; Halsch, C.A.; Schultz, C.B.; Wagner, D.L. Missing the bigger picture: Why insect monitoring programs are limited in their ability to document the effects of habitat loss. Conserv. Lett. 2023, 16, e12951. [Google Scholar] [CrossRef]
- Lozada, A.; Day, C.C.; Landguth, E.L.; Bertin, A. Simulation-based insights into community uniqueness within fragmented landscapes. Landsc. Ecol. 2023, 38, 2533–2546. [Google Scholar] [CrossRef]
- Wilson, C.J.; Jamieson, M.A. The effects of urbanization on bee communities depends on floral resource availability and bee functional traits. PLoS ONE 2019, 14, e0225852. [Google Scholar] [CrossRef]
- Xie, Z.; Qiu, J.S.; Chen, X.M. Decline of nest site availability and nest density of underground bees along a distance gradient from human settlements. Entomol. Sci. 2013, 16, 170–178. [Google Scholar] [CrossRef]
- Shebl, M.A.; Al Aser, R.M.; Ibrahim, A. Nesting biology and seasonality of long-horned bee Eucera nigrilabris Lepeletier (Hymenoptera: Apidae). Sociobiology 2016, 63, 1031–1037. [Google Scholar] [CrossRef]
- Pereira, F.W.; Carneiro, L.; Gonçalves, R.B. More losses than gains in ground-nesting bees over 60 years of urbanization. Urban Ecosyst. 2021, 24, 233–242. [Google Scholar]
- Matteson, K.C.; Ascher, J.S.; Langellotto, G.A. Bee richness and abundance in New York City urban gardens. Ann. Entomol. Soc. Am. 2008, 101, 140–150. [Google Scholar]
- Everaars, J.; Strohbach, M.W.; Gruber, B.; Dormann, C.F. Microsite conditions dominate habitat selection of the red mason bee (Osmia bicornis, Hymenoptera: Megachilidae) in an urban environment: A case study from Leipzig, Germany. Landsc. Urban Plan. 2011, 103, 15–23. [Google Scholar]
- Ascher, J.S.; Pickering, J. Discover Life Bee Species Guide and World Checklist (Hymenoptera: Apoidea: Anthophila). 2020. Available online: http://www.discoverlife.org/mp/20q?guide=Apoidea_species (accessed on 13 April 2022).
- Da Rocha Filho, L.C.; Garófalo, C.A. Nesting Biology of Megachile (Chrysosarus) guaranitica and High Mortality Caused by Its Cleptoparasite Coelioxys bertonii (Hymenoptera: Megachilidae) in Brazil. Austral. Entomol. 2015, 55, 25–31. [Google Scholar]
- O’Neill, K.M.; O’Neill, J.F. Brood Parasitism of the Resin Bee Megachile campanulae (Robertson) by Coelioxys modesta Smith (Hymenoptera: Megachilidae). J. Kans. Entomol. Soc. 2016, 89, 117–127. [Google Scholar]
- Sabino, W.D.O.; Antonini, Y. Nest architecture, life cycle, and natural enemies of the neotropical leafcutting bee Megachile (Moureapis) maculata (Hymenoptera: Megachilidae) in a montane forest. Apidologie 2017, 48, 450–460. [Google Scholar]
- Parizotto, D.R. Natural enemies of the oil-collecting bee Centris analis (Fabricius, 1804) with notes on the behavior of the cleptoparasite Coelioxys nigrofimbriata Cockerell, 1919 (Hymenoptera, Apidae). J. Hymenopt. Res. 2019, 70, 1–16. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Description |
|---|---|
| Bio 1 | Annual mean temperature |
| Bio 2 | Mean diurnal range (mean of monthly max temp–min temp) |
| Bio 3 | Isothermality (bio2/bio7) × 100 |
| Bio 4 | Temperature seasonality (standard deviation × 100) |
| Bio 5 | Max temperature of the warmest month |
| Bio 6 | Min temperature of the coldest month |
| Bio 7 | Temperature annual range |
| Bio 8 | Mean temperature of the wettest quarter |
| Bio 9 | Mean temperature of the driest quarter |
| Bio 10 | Mean temperature of the warmest quarter |
| Bio 11 | Mean temperature of the coldest quarter |
| Bio 12 | Annual precipitation |
| Bio 13 | Precipitation of the wettest month |
| Bio 14 | Precipitation of the driest month |
| Bio 15 | Precipitation seasonality (coefficient of variation) |
| Bio 16 | Precipitation of the wettest quarter |
| Bio 17 | Precipitation of the driest quarter |
| Bio 18 | Precipitation of the warmest quarter |
| Bio 19 | Precipitation of the coldest quarter |
| Species | No. NS | pROC Ratio | ||
|---|---|---|---|---|
| Minimum | Maximum | Mean | ||
| Chalicodomaflavipes | 500 | 1.53 | 1.99 | 1.9 |
| Chalicodomasicula | 500 | 1.91 | 1.99 | 1.96 |
| Coelioxys coturnix | 500 | 1.34 | 1.87 | 1.67 |
| Megachileminutissima | 500 | 1.63 | 1.99 | 1.87 |
| Osmiasubmicans | 500 | 1.9 | 1.96 | 1.94 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Okely, M.; Engel, M.S.; Shebl, M.A. Climate Change Influence on the Potential Distribution of Some Cavity-Nesting Bees (Hymenoptera: Megachilidae). Diversity 2023, 15, 1172. https://doi.org/10.3390/d15121172
Okely M, Engel MS, Shebl MA. Climate Change Influence on the Potential Distribution of Some Cavity-Nesting Bees (Hymenoptera: Megachilidae). Diversity. 2023; 15(12):1172. https://doi.org/10.3390/d15121172
Chicago/Turabian StyleOkely, Mohammed, Michael S. Engel, and Mohamed A. Shebl. 2023. "Climate Change Influence on the Potential Distribution of Some Cavity-Nesting Bees (Hymenoptera: Megachilidae)" Diversity 15, no. 12: 1172. https://doi.org/10.3390/d15121172
APA StyleOkely, M., Engel, M. S., & Shebl, M. A. (2023). Climate Change Influence on the Potential Distribution of Some Cavity-Nesting Bees (Hymenoptera: Megachilidae). Diversity, 15(12), 1172. https://doi.org/10.3390/d15121172