Abstract
The natural valleys of large rivers provide suitable nesting and foraging sites for many bird species. In this study, we assess the effects of water fluctuations during the breeding season on Common Sandpipers’ nest survival on a semi-natural section of the Vistula River, and evaluate which tactics this species can adopt to minimise the effects on a long-term scale. Birds can avoid losses due to flooding by adjusting the timing of nesting to avoid the occurrence of extreme water conditions, and/or by choosing a safe nesting location. We found that nest location is a direct response to periodic high water, and the threat of flooding seems to be one of the important determinants of Common Sandpiper nest placement. The choice of nest location is not random, as 70% of pairs nested above a height that provides an 80% chance of avoiding flooding of the nest. For the lowest-lying nests found in this study, nesting success was reduced up to 92% because of inundation. Birds choose more elevated places, even during the second half of the season when the threat of flooding is lower. Their preference for nesting above a certain height does not change over the breeding season, which corresponds to the very small impact of the date of first egg laying on breeding success. Those who breed earlier in the season have a slightly higher risk of being flooded, but nesting in more elevated places reduces this risk much more than shifting the time of breeding.
1. Introduction
The riparian zones of large rivers are considered hot spots of biodiversity due to their continuous supply and accumulation of nutrient-rich sediments and high environmental heterogeneity in space and time, which makes floodplains among the world’s most biologically productive areas [1,2]. A natural flow regime is vital to the functioning and maintenance of a healthy river ecosystem [3], as proper nutrient exchange influences abiotic and biotic processes within the riverbed and enhances biological productivity [4,5,6]. Hence, the natural flow of the river is the primary factor shaping river habitat dynamics [7]. Undisturbed flow regimes, especially peak flows, are crucial for maintaining river habitats, and altering the natural fluvial characteristics of the river changes the habitats present in the river valley [5,8]. Many ground-nesting bird species form distinctive bird communities nesting in riverbeds, occupying both sandy fragments and areas of low vegetation [9,10,11]. The flow regime has a significant impact on the evolution of riparian species’ adaptations [12], the diversity of bird communities [9,13], and the timing of their breeding [14].
The Vistula is Poland’s longest river, whose catchment area covers approximately 62% of the country’s area and is the second largest catchment area draining into the Baltic Sea [15]. Its middle run is an example of a semi-natural braided middle European river with highly well preserved semi-natural conditions [16,17]. Its yearly transport of about 0.5 million m3 of sediment affects the ongoing creation of sandy bars and islands [18], which are important breeding sites for terns, gulls, and plovers. Approximately 50% of the Polish populations of Little Tern Sternula albifrons (VU on the Polish red list) and Common Ringed Plover Charadrius hiaticula (EN on Polish red list) breed on these sandy bars and islands [19,20].
The only species from the Scolopacidae family that nests in the Vistula River bed is the Common Sandpiper Actitis hypoleucos, a widespread riverine, migratory, wide-ranging species breeding from Western Europe to East Asia [21,22]. It breeds on various water bodies and rivers of all sizes, including small mountain and large lowland rivers, and prefers riverbeds with sand and gravel substrate and natural character with varied channel morphology and low levels of anthropogenic alteration [23,24,25]. In Europe, their population is estimated to be 794,000–1,460,000 pairs and is decreasing [26,27]. The number of breeding Common Sandpipers in Poland is 1600–2200 pairs, with a moderate decrease. [28,29]. The average density of this species in the middle Vistula River is 5.7–6.5 pairs/10 km of the river, but it reaches up to 35 pairs/10 km locally [30,31]. The breeding of Common Sandpiper populations on small rivers has been studied in detail [32,33,34]; however, in mountain rivers, spring floods result in the spontaneous restoration of natural habitats and may increase the number of breeding pairs due to the increase in breeding site availability and food abundance [35,36]. In large lowland rivers, seasonal spring floods are responsible for many breeding failures in birds nesting in the riverbed [37,38,39], which is also the case in the middle Vistula [40,41]. To date, no analyses have been presented on the effects of periodic high-water events on the breeding success of riverine species over a long period of time. Understanding the impact of flow regime on riverine birds might be important in quantifying the environmental impact of river regulations on birds’ populations, especially during a period of rapid climate change [42].
Birds can change their breeding timing due to yearly changes in water levels [43,44,45], and repeated disturbances can influence the long-time mean breeding date, as is observed in plovers breeding in tidal areas [46]. Moreover, there may also be adaptations related to the selective choice of nest placement that minimises the risk of flooding [45]. However, due to climate change and the increased frequency of extreme events, as well as the decreasing possibility of buffering atmospheric phenomena in the catchment, the frequency of extreme events on the river may increase [47,48,49], and the impact of flooding as a factor limiting the breeding success of the Common Sandpiper and other riverine species may increase. There are two options to avoid losses due to flooding: adjusting the timing of nesting to the occurrence of extreme water conditions and/or choosing a safe nesting location [50]. In this study, we assess the effects of natural spring floods on the Common Sandpiper’s nest survival on a semi-natural section of a large lowland river, and evaluate what tactics this species can adopt to minimise the effects.
2. Materials and Methods
2.1. Study Area
The studies were conducted in Central Poland in the 10 km long section of the Vistula River. The study area is within the Natura 2000 protected area (PLB 140004 Middle Vistula River) and the Important Birds Area (PL083) [20] (Figure 1). There are two nature reserves (Wyspy Świderskie Nature Reserve and Wyspy Zawadowskie Nature Reserve) in this river section, created to protect rare and endangered bird species (i.e., the Little Tern, the Common Tern Sterna hirundo, the Common Gull Larus canus, and the Common Ringed Plover) and their habitats, consisting of riverbeds, islands, and natural processes of sedimentation and erosion. In the studied area, there is still a continuous process of deposition and redeposition of sand and the formation of islands and sandy bars [51], with the most quasi-natural water regime on the Vistula River compared to other sections of this river [52]. There was no significant anthropogenic impact on natural fluvial processes; however, there are levees on both sides that significantly narrowed the floodplain to 1.0–1.8 km wide [51,53,54].
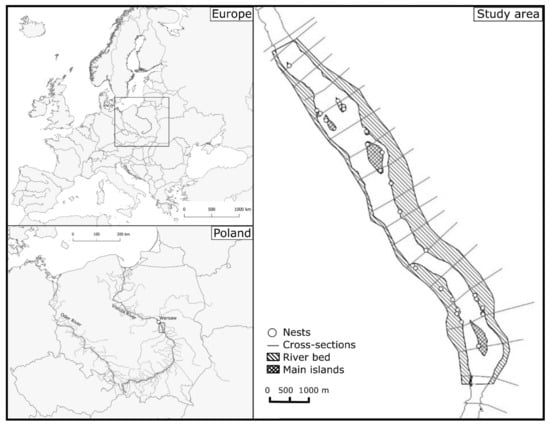
Figure 1.
Study area with cross-sections used to model water level.
2.2. Field Study
Nest searching was conducted between mid-April and mid-July in 2014 and 2015. During the surveys, we used a standardised territory mapping technique [55], adapted to the species and study area [56]. At the beginning of the season, we mapped the territory based on singing or fighting males or multiple observations of pairs of birds, including singing or copulations. Starting from the second half of May, single observations of birds in territories were also considered indicative of the presence of a breeding territory. We surveyed every known territory at least 2 times during the season, and we monitored nests every 3–7 days. In total, we found 43 nests by thoroughly searching during walking controls in already mapped territories. To monitor the incubation phenology, we put Thermochron® iButton data loggers (Maxim Integrated Products, Sunnyvale, CA, USA) in eight nests in 2014 and 21 nests in 2015 to collect temperature data every 5 to 20 min. The positions of all nests were marked using a Garmin GPS device (GPSMAP 64, Garmin Ltd., Olathe, KS, USA).
In the case of eight nests found during egg laying (18.7%), the first egg day (FED) was determined directly. In other cases, when the nest was found during incubation, the assessment of FED was based on back-calculation when the hatching date was known (N = 12; 27.9% cases), assuming that the incubation and laying period is 26 days [57], and using the floating test [58] to determine the number of days of incubation (N = 23; 53.5%). We determined the incubation start (IS), assuming that birds lay subsequent eggs at 1.5-day intervals [57]. Second clutches were distinguished when they were found in known territory after the identified loss of the first clutch in a given territory (after a flood or after predation) or found very late in the season (FED > 50), because no known first clutches were found after this date. However, females may lay one or, very rarely, two replacement clutches, if none of the previous clutches were successful [59,60]. We did not observe clutches being repeated more than once and we assumed that clutches were repeated only once.
To determine the nest height (in m above sea level), we used the Digital Terrain Model created in 2011, with a 10 cm grid, obtained from the Polish Main Office of Geodesy and Cartography. This model is the closest to the years of field study carried out. Due to the accuracy of GPS devices, the height was obtained not by determining the height of the exact point, but by taking the median heights from a 2 m buffer zone around each nest. In 11 cases (25.6%), when the nest was located on the slope and the 2 m buffer zone reached the base of the slope, lowering the median height, the buffer zone was manually moved on the map toward the top of the hill. The correction was also made based on the photographic documentation of each nest, which was taken during the fieldwork. After the correction, the nest was always in the buffer zone but out of the centre of the buffer. All work involving the maps and calculations of this part of the work was completed using QuantumGIS 3.4 software [61].
2.3. Water Level Assessment for Each Nest Site
To model river surface elevation in the riverbed and river valley, we used the one-dimensional model Mike 11 [62], based on flow data from 1980 to 2015 (Polish Institute of Meteorology and Water Management). The model took into account the width of the riverbed and the floodplain terrace, and we obtained the water surface level (in m above sea level) in 19 modelled cross-sections of the river. For each cross-section, the water level was known for every 6 h over a span of 36 years (4 records per day) (Figure A2).
Water levels at each nest were inferred from the water levels at sections directly upstream and downstream of each nest (Figure 2), assuming that the angle of decline was constant between two neighbouring sections. The water level at the nest n at time-stamp t (Wn,t) was calculated for a given nest using the formula in Equation (1):
where Wαn,t is the water level measured at the closest upstream section of the nest n at time-stamp t, Wβn,t is the measured water level at the closest downstream section of the nest n at time-stamp t, Dβn is the distance between the nest n and the closest downstream section, and Dαn is the distance between the nest n and the closest upstream section. Measurements were calculated by transforming each line section into a series of points with a distance of 1 m between them; then, the distance was measured between each nest and the section point closest to that nest for the closest downstream and upstream sections.
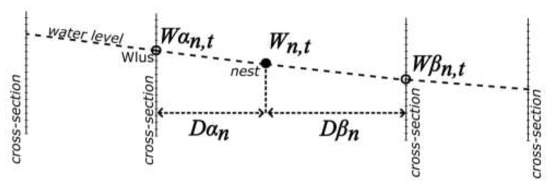
Figure 2.
The method of calculating the water level at the nest location (white dot) based on known water level in the following cross-sections. Wn,t: water level at the nest location, Wαn,t: water level at upstream cross-section, Wβn,t: water level at downstream cross-section, Dαn: distance between nest and upstream cross-section, and Dβn: distance between nest and downstream cross-section.
Within the studied section of the Vistula River, the slope, according to the Digital Terrain Model, is about 3.4%, and the mean water level difference between the beginning and the end of the study area is 4.2 m. Hence, the nests’ heights, expressed in metres above sea level, are not comparable and do not translate into actual height above the water level; therefore, we calculated the relative nest height (RNH), which was the difference between the nest height (in m above sea level) and the water level from one particular day for all nests. We chose the 31 May 2007, when the water level was the lowest in April–May for all years. In effect, we obtained the RNH (in m) to compare nests and for use in further analyses.
2.4. Estimating the Probability of Avoiding Flooding of the Nest
To determine the probability of avoiding flooding of the nest, data on whether nests might survive each day were simulated using the known location and heights of the nests found in two studied seasons. We modelled the survival of the nest locations for every possible FED starting from day 24 (24th April) to day 79 (18th June) for every 6-hour period over a span of 36 years (1980–2015). If the water level was higher than the nest location even once during the incubation period, the nest was assumed not to have survived due to flooding. The time needed for laying and incubation in the Common Sandpiper lasts 26 days [57]. The survival probability for a nest on a given FED was estimated as the percentage of the 36 years that a nest in that location would have survived if it had been built on that FED. To calculate this, we used the locations of nests found during 2014–2015. The water discharge data modelled to the water level were the only long-term data available to use to quantify the probability of avoiding flooding of the nest. There are neither data on nest location nor a Digital Terrain Model available for such a long period. Therefore, in this study, data on the height of nests found and incubation dates provide a background for the analysis of the impact of seasonal changes in the water level of the Vistula on the survival of Common Sandpiper nests over a 36-year period.
2.5. Assessing the Effect of the FED on the Probability of Avoiding Flooding of the Nest
To check if there was any significant difference in the probability of avoiding flooding of the nest between FEDs, we used a Friedman test with the Conover adjustment [63] usin the R package PMCMRplus [64]. To avoid a large number of subsequent pairwise tests, the FED values were clustered using hierarchical clustering [65] to find suitable continuous groups of FED values to compare. When clustering, each FED was represented by a vector of the probability of avoiding flooding of the nest for each nest if they had been built on that FED. The vectors of the probability of avoiding flooding of the nest were then averaged for each cluster, and the resulting mean vectors for each cluster were compared using pairwise Wilcoxon tests with Bonferroni adjustments for multiple testing [66,67]. We used linear and polynomial regressions to quantify the effects of FED on the probability of avoiding flooding of the nest. The polynomial regression was performed by adding additional predictors when calculating linear regression, calculated as FED to the power of 2, 3, 4, and 5.
2.6. Assessing the Effect of RNH on the Probability of Avoiding Flooding of the Nest
We used spline regression with linear splines, linear regression on the natural logarithm of relative height, linear regression on the natural logarithm of relative height plus the relative height, and polynomial regression of the second and third degree to assess the influence of RNH on the probability of avoiding flooding of the nest. The R package ‘splines’ [68] was used for linear spline regression where the location of the knot was obtained through optimization using the limited-memory Broyden–Fletcher–Goldfarb–Shanno algorithm (BFGS) [69]).
For all regression model evaluations, we used the Bayesian information criterion (BIC) as this is considered a more conservative information criterion, which gives higher penalties to models with more parameters [70]. The model with the lowest BIC score is considered the best fitting model [70]; hence, we calculated the differences between the model with the lowest BIC and subsequent candidate models (ΔBIC). Large absolute values of ΔBIC indicate that one approach performed substantially better than the other, whereas ΔBIC values less than 2 indicate that the two methods performed equally well [71]. In the case of two models with similar BICs and with ΔBIC < 2, we chose the model with the fewest parameters. The significance level was set at p < 0.05 in all analyses.
3. Results
3.1. Characteristics of Found Nests
In 2014–2015, Common Sandpipers started egg laying between 25th April and 18th June (24–79 FED); for most known first clutches, egg laying starts before day 39, and in known repeated clutches, egg laying starts after day 54 (N = 40; Figure 3). There is no relationship between nest height and FED (r = 0.09; N = 43; p = 0.550).
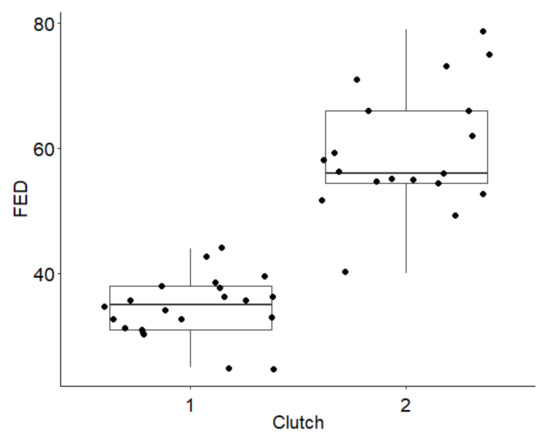
Figure 3.
Dates of (1) first and (2) repeated clutches of the Common Sandpiper during 2014–2015. The box-plot shows the median date (horizontal line), interquartile range (rectangle), and range (vertical line). FED—the date of first egg laying.
The nest height of the 43 nests found during 2014–2015 varied from 82.07 m to 88.20 m above sea level (mean = 85.07, range = 6.13), and RNH varied from 0.64 m to 4.58 m (mean = 2.80, range = 3.94). The distribution of nest height was negatively skewed (Sk = −0.42), and more nests were built above the mean height (Figure 4).
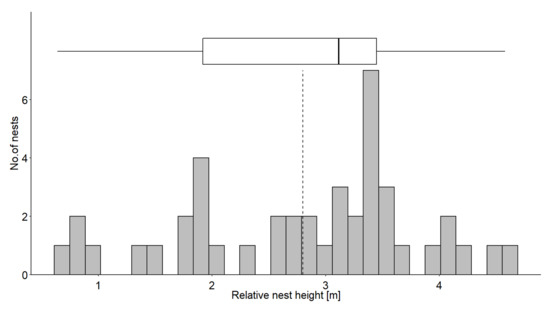
Figure 4.
Distribution of relative nest heights (RNH). The box plot shows median and interquartile range, and the dashed line represents the mean.
3.2. Assessment of the Probability of Avoiding Flooding of the Nest during a 36-Year Period
The mean survival of individual nests during the 36-year period varied from 16.6% to 98.5%, with a mean of 80.7% (Figure 5). The distribution of individual probability of avoiding flooding of the nest was negatively skewed (Sk = −1.93), and more nests had a higher probability of avoiding flooding of the nest than the mean (Figure 5).
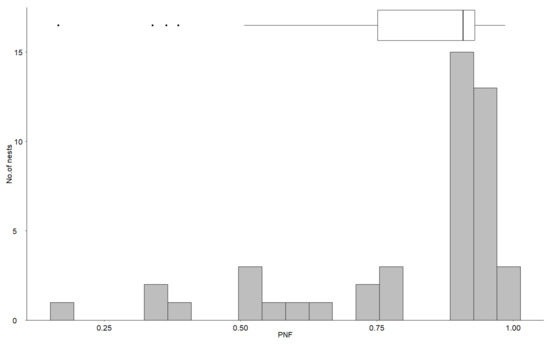
Figure 5.
Distribution of individual probability of avoiding flooding of the nest (PNF) during a 36-year period. The box plot shows the median and interquartile range.
The yearly mean probability of avoiding flooding of the nest during the analysed period varied from 7.8% (in 2010) to 100% (in 1993 and 2007), with a mean of 80.7% and no significant trend (linear regression coefficient = −0.004; p = 0.205; Figure 6). The exceptionally low nest survival rate in 2010 was due to flooding that covered much of central Europe [72].
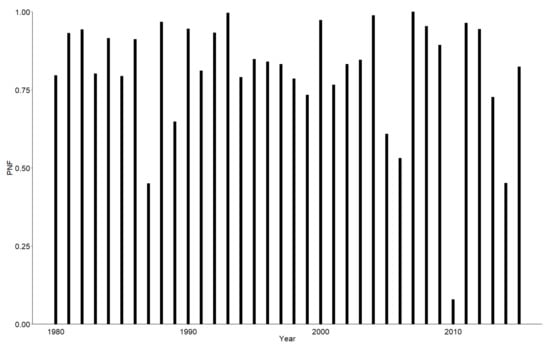
Figure 6.
Mean probability of avoiding flooding of the nest (PNF) in the Common Sandpiper from 1980 to 2015.
3.3. Effect of FED on Probability of Avoiding Flooding of the Nest
We could not find a model to describe the probability of a nest not being flooded as a function of FED. The best model found (Equation (2)) had a non-significant regression coefficient (t = 1.626; p = 0.104) and a very low adjusted coefficient of determination (R2 < 0.001), indicating that even though it was the best model, it was not a good fit for the data (Table A1). Moreover, there was no formula that could estimate the probability of a nest not being flooded significantly better than taking the average probability of the nest not being flooded.
p(not flooded) = 0.7856 + 0.0004∙FED,
The probability of avoiding flooding of the nest within the breeding season varied from 0.78 to 0.84, and there was a significant difference in the probability of nests not being flooded between different FEDs at the 95% confidence level (Friedman test with Conover adjustment, F = 15.15; p < 0.001). Hierarchical clustering based on the probability of avoiding flooding of the nest revealed three clusters with different survival rates: early FED (25–43), middle FED (44–71), and late FED (72–79) (Figure A1, Figure 7). The probability of nests not being flooded in the middle period was significantly lower than for nests in the late period (Wilcoxon test with Bonferroni adjustment, W = 56, adjusted p < 0.001). There were no significant differences in not being flooded between early nests and middle ones (Wilcoxon test with Bonferroni adjustment, W = 486, adjusted p = 1.0) or for early and late nests (Wilcoxon test with Bonferroni adjustment, W = 369, adjusted p = 0.09). In the middle cluster, there is a slight increase in the probability of avoiding flooding of the nest after an FED of 55. In fact, late first clutches had the smallest probability of avoiding flooding of the nest—even smaller than early first clutches and replaced ones (Figure 7).
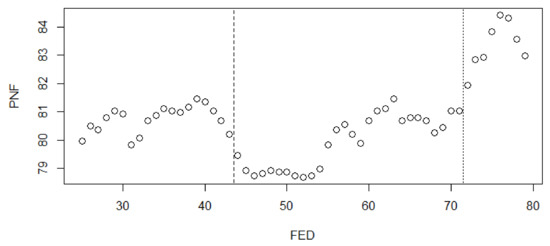
Figure 7.
Average probability of avoiding flooding of the nest (PNF) between 1980 and 2015 if placed on a particular FED. The dashed lines show the split between early, middle, and late clusters with different survival rates according to hierarchical clustering.
3.4. Effect of RNH on Probability of Avoiding Flooding of the Nest
The probability of avoiding flooding of the nest was strongly dependent on RNH. The two models were fitted similarly (Table A2). The model using both ln(RNH) and RNH (Equation (3)) and the spline regression with 1 knot (Equation (4)) (R2 = 0.95 in both cases) showed a significantly higher probability of avoiding flooding of the nest for nests built above the 2.25 m RNH (Figure 8).
p(n not flooded) = 0.7575∙ln(RNH) − 0.1600 + 0.5479,
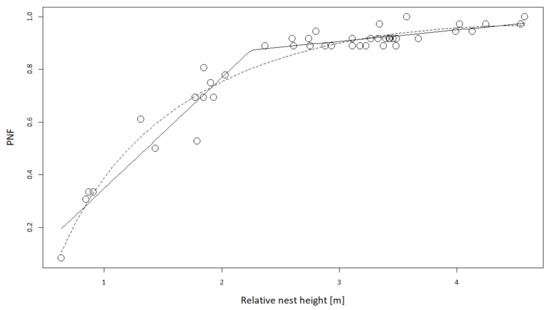
Figure 8.
Dependence of the probability of avoiding flooding of the nest (PNF) on relative nest height. The solid line represents the linear spline regression with one breakpoint at 2.25 m, and the dashed line represents the model with both ln(RNH) and RNH as independent variables. White dots show probability of not flooding for each nest with fixed RNH over the 36-year period.
For nests built below 2.25 m, every 10 cm increase in elevation provided a 4.3% higher survival probability, while for nests built above 2.25 m, there was only a 0.4% increase in survival probability. Hence, it was most beneficial for birds to build nests above the 2.25 m RNH, and indeed, 70% of found nests were placed above the 2.25 m RNH.
The effect of FED on the probability of avoiding flooding of the nest was much less important than the effect of RNH, as the coefficient for FED was not significant and very small. Even if the coefficient was significant, it would indicate that waiting 1 day would result in an increase in survival probability of 0.040%, which is roughly the effect obtained by 1 additional cm of nest height when the nest was already above 2.25 m (0.043%).
4. Discussion
The regulation of European rivers is very common, and it is estimated that more than one million dams on rivers of all sizes have been constructed, affecting entire river catchments [73,74] and riparian habitats [75]. Altering of the river flow, e.g., due to development and dam construction or climate change, has a significant impact on aquatic and riparian organisms [76]. Climate and anthropogenic changes in the entire river catchment manifested due to increased intensity of extreme events [76,77]. At the present time, the pressure from floods is generally higher than during thw time when the Common Sandpiper was adapting to living in a riparian environment [49,78]. Anthropogenic changes in hydrological conditions in the Vistula Valley began in the 13th century and resulted in increased debris flow, faster runoff, and shorter time between precipitation and runoff [53,54]. The narrowing of the riverbed that has occurred in the last 100 years concentrates the water in a narrow area between levees, not allowing flood waters to spill over into the wide river valley [16,40,54]. Hence, these changes may have important implications for the height of nest placement and the probability of avoiding flooding of the nest in the Common Sandpiper. On one hand, floods are an important cause of breeding losses, while on the other hand, they are responsible for the annual restoration of river habitats to their initial state, such as sandbars and sandy islets, which are among the most sensitive and crucial habitats for breeding birds and also the most vulnerable to flooding [20,36,79,80]. Hence, water conditions in a riverbed may strongly affect the species richness and distribution [36,81,82], breeding success [83], foraging efficiency [84], and survival rate [82] of riverine bird species.
In the Vistula River, the water conditions are very unstable during the breeding season, and the hydrological regime of the studied part of the Vistula depends more on hydrological and precipitation conditions in its tributaries than on atmospheric conditions in the studied area [52]. The difference between the minimum and maximum flow of the Vistula is more than 80 times [85], and the water height during floods is up to 6.5 m higher than the average water level [17]. Although the Common Sandpiper usually chose more elevated nest locations, nests were still exposed to floods, which, in some seasons, may significantly reduce the breeding success of this species. For the lowest-lying nests found in this study, the probability of nest inundation was up to 92%. Hence, flooding must be a considerable factor that affects not only breeding success but also reproductive strategy. Especially over the 36 years considered in our study, the water level of the river did not show a long-term trend of change. Hence, it seems that flooding is not responsible for the population declines in Common Sandpipers.
Nesting early in the season has the obvious advantages of being able to occupy the best available territory and attract the best mate [86,87]. Moreover, early breeding increases the time available for replacement clutches and can be the sole important factor in increasing breeding success [87]. The Common Sandpiper usually repeats breeding after failure [59], but in northern latitudes with shorter breeding seasons, only birds that start their first broods early can repeat them after loss [60]. The timing of breeding can also be adaptive to yearly water conditions on the river or seasonal tide [43,46], on a long-term scale. In the middle Vistula, early nesting certainly has not had a significant influence on lowering the probability of flooding. This is due to the unpredictable timing of floods, which occur on the Vistula most frequently between April and June [52], covering the entire breeding season of the Common Sandpiper. Moreover, the exact date of flood occurrence in a given year is unpredictable [52], and this is why it is difficult for birds to adjust their phenology to this event. Significantly higher nest survival probability occurs only at the very end of the breeding season, when the water level in the Vistula is usually low and stable. The highest probability of avoiding flooding of the nest applies to the latest, repeated broods, which have the lowest chance of being exposed to flooding. The mating system (social monogamy) of the Common Sandpiper should promote birds arriving early in breeding territories. Moreover, in early broods, the proportion of males, which are the more competitive sex, is higher than that in late broods [88]. Males hatched earlier in the season may start breeding for the first time at a younger age and achieve better conditions due to a longer growth period [59,88]. It seems that the benefits of early breeding are higher than the unpredictable costs of being flooded, especially since the changing of FED is not crucial to avoiding inundation. The second way to mitigate nesting losses due to flooding is ensuring proper nest location [43,50]. In the study area, the height at which the Common Sandpipers built their nests was not random. The majority of nests (70%) were built 2.25 m above the RNH, which allowed them to achieve a probability of avoiding flooding of the nest above 80% over a long period of time. It appears that while the benefits of early nesting are related to factors other than hydrological ones, nest location is a direct response to periodic high water levels, and the threat from flooding seems to be one of the important determinants of Common Sandpipers’ nesting locations within a riverbed. The preference for nesting above a certain height did not change over the breeding season, which corresponded with the small impact of FED on breeding success. The lack of such variation in nest height over time shows that Common Sandpipers are ‘prepared’ for flooding that may occur at any time during the season. Therefore, for Common Sandpipers, it is profitable to breed as early as possible due to the mating system, and as high as possible due to possibility of flooding; however, only proper nest height significantly increases the likelihood of the nest being unaffected by flooding.
Author Contributions
M.E. and W.M. developed the ideas and designed the study; M.E. carried out the fieldwork; E.R. statistically analysed the data, and M.E. and W.M. took the lead in writing the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research did not receive any specific grants from funding agencies in the public or commercial sectors. The fieldwork complies with the current laws of Poland on voluntary basis.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are available upon reasonable request to the corresponding author.
Acknowledgments
We would like to thank Radosław Pasiok for modelling the water level, and all the volunteers who carried out fieldwork, especially Łukasz Wardecki and Dawid Sikora. The fieldwork complies with the current laws of Poland.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A

Figure A1.
Heatmap of probability of nests not being flooded when built on a particular FED. Darker colours indicate a higher probability of not being flooded. FEDs are rearranged due to clustering.

Figure A2.
Mean (black line), maximum, and minimum water levels (above sea level) on one cross-section of the Vistula River over 36 years during breeding season (day 1 = 1 April).

Table A1.
Regression model comparison for modelling the probability of avoiding flooding of the nest as a function of first egg day (FED). Only models with ΔBIC lower than 10 are shown.
Table A1.
Regression model comparison for modelling the probability of avoiding flooding of the nest as a function of first egg day (FED). Only models with ΔBIC lower than 10 are shown.
| Model No. | Model Structure | BIC | ΔBIC |
|---|---|---|---|
| 1 | spline(h) + FED + FED2 | −7333.18 | 0.00 |
| 2 | spline(h) + FED + FED2 + FED3 | −7331.06 | 2.12 |
| 3 | spline(h) + FED + FED2 + FED3 + FED4 | −7329.69 | 3.48 |
| 3 | spline(h) + FED + FED2 + FED3 + FED4 | −7329.69 | 3.48 |
Table A2.
Regression model comparison for modelling probability of avoiding flooding of the nest as a function of relative nest height (RNH). Only models with ΔBIC lower than 10 are shown.
Table A2.
Regression model comparison for modelling probability of avoiding flooding of the nest as a function of relative nest height (RNH). Only models with ΔBIC lower than 10 are shown.
| Model No. | Model Structure | BIC | ΔBIC |
|---|---|---|---|
| 1 | Ln(RNH) + RNH | −129.55 | 0.00 |
| 2 | Linear splines, 1 knot | −129.25 | 0.30 |
| 3 | Ln(RNH) | −126.94 | 2.61 |
| 3 | 3rd degree polynomial | −126.29 | 3.26 |
References
- Ward, J.V.; Tockner, K.; Schiemer, F. Biodiversity of floodplain river ecosystems: Ecotones and connectivity. Regul. Rivers Res. Manag. 1999, 15, 125–139. [Google Scholar] [CrossRef]
- Tockner, K.; Malard, F.; Ward, J.V. An extension of the flood pulse concept. Hydrol. Process. 2000, 14, 2861–2883. [Google Scholar] [CrossRef]
- Sofi, M.S.; Bhat, S.U.; Rashid, I.; Kuniyal, J.C. The natural flow regime: A master variable for maintaining river ecosystem health. Ecohydrology 2020, 13, e2247. [Google Scholar] [CrossRef]
- Bayley, P. Understanding large river-floodplain ecosystems. BioScience 1995, 45, 153–158. [Google Scholar] [CrossRef]
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The natural flow regime: A paradigm for river conservation and restoration. BioScience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Ballinger, A.A.; Lake, P.S. Energy and nutrient fluxes from rivers and streams into terrestrial food webs. Mar. Freshw. Res. 2006, 57, 15–28. [Google Scholar] [CrossRef]
- Ward, J.V.; Tockner, K.; Arscott, D.; Claret, C. Riverine landscape diversity. Freshw. Biol. 2002, 47, 517–539. [Google Scholar] [CrossRef]
- Zeiringer, B.; Seliger, C.; Greimel, F.; Schmutz, S. River hydrology, flow alteration, and environmental flow. In Riverine Ecosystem Management; Schmutz, S., Sendzimir, J., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 67–89. [Google Scholar] [CrossRef]
- Figarski, T.; Kajtoch, Ł. Alterations of riverine ecosystems adversely affect bird assemblages. Hydrobiologia 2015, 744, 287–296. [Google Scholar] [CrossRef]
- Božič, L.; Denac, D. Population dynamics of five riverbed breeding bird species on the lower Drava River, NE Slovenia. Acrocephalus 2017, 38, 85–126. [Google Scholar] [CrossRef]
- Keller, M.; Kot, H.; Dombrowski, A.; Rowiński, P.; Chmielewski, S.; Bukaciński, D. Birds of the Middle Vistula River; Mazowiecko-Świętokrzyskie Towarzystwo Ornitologiczne: Pionki, Poland, 2017; (In Polish with English summary). [Google Scholar]
- Poff, N.L.R. Beyond the natural flow regime? Broadening the hydro-ecological foundation to meet environmental flows challenges in a non-stationary world. Freshw. Biol. 2018, 63, 1011–1021. [Google Scholar] [CrossRef]
- Belmar, O.; Ibáñez, C.; Forner, A.; Caiola, N. The influence of flow regime on ecological quality, bird diversity, and shellfish fisheries in a lowland Mediterranean river and its coastal area. Water 2019, 11, 918. [Google Scholar] [CrossRef]
- D’Amico, F.; Manel, S.; Mouchès, C.; Ormerod, S.J. River birds in regulated rivers: Cost or benefit? Verh. Internat. Verein. Limnol 2000, 27, 167–170. [Google Scholar] [CrossRef]
- Bolle, H.J.; Menenti, M.; Rasool, S.I. (Eds.) Second Assessment of Climatechange for the Baltic Sea Basin; Springer: Cham, Switzerland, 2015. [Google Scholar] [CrossRef]
- Starkel, L. Evolution of the Vistula River Valley since the Last Glaciation till Present; Polish Academy of Sciences: Warsaw, Poland, 2001; (In Polish with English summary). [Google Scholar]
- Romanowski, J.; Matuszkiewicz, J.; Kowalczyk, K.; Kowalska, A.; Kozlowska, A.; Solon, J.; Bouwma, I.M.; Middendorp, H.; Reijnen, R.; Roemeijer, R.; et al. Evaluation of Ecological Consequences of Development Scenarios for the Vistula River Valley; Vistula Econet Development and Implementation: Warsaw, Poland, 2005. [Google Scholar]
- Sosnowska, A. Dynamics of mid-channel bars in the Middle Vistula River in response to ferry crossing abutment construction. Open Geosci. 2020, 12, 290–298. [Google Scholar] [CrossRef]
- Wilk, T.; Chodkiewicz, T.; Sikora, A.; Chylarecki, P.; Kuczyński, L. Red List of the Birds of Poland; Polish Society for the Protection of Birds: Marki, Poland, 2020; (In Polish with English summary). [Google Scholar]
- Wilk, T.; Jujka, M.; Krogulec, J.; Chylarecki, P. (Eds.) Important Bird Areas of International Importance in Poland; Polish Society for the Protection of Birds: Marki, Poland, 2010. [Google Scholar]
- Cramp, S.; Simons, K.E.L. The Birds of the Western Palearctic; Waders to Gulls; Oxford University Press: Oxford, UK, 1983; Volume III. [Google Scholar]
- Van Gils, J.; Wiersma, P. Common Sandpiper (Actitis hypoleucos). In Handbook of the Birds of the World; Del Hoyo, J., Elliot, A., Sargatal, J., Eds.; Volume 3 (Hoatzin to Auks); Lynx Edicions: Barcelona, Spain, 1996; p. 513. [Google Scholar]
- Diez, F.; Peris, S.J. Habitat selection by the Common Sandpiper (Actitis hypoleucos) in west-central Spain. Ornis Fenn. 2001, 78, 127–134. [Google Scholar]
- Hammer, T.; Liker, A.; Szentirmai, I. Habitat preference of common sandpipers (Actitis hypoleucos) along the River Rába, Hungary. Ornis Hung. 2013, 21, 26–35. [Google Scholar] [CrossRef]
- Wilk, T.; Bobrek, R.; Pępkowska-Król, A.; Neubauer, G.; Kosicki, J.Z. (Eds.) The Birds of the Polish Carpathians—Status, Threats, Conservation; Polish Society for the Protection of Birds: Marki, Poland, 2016; (In Polish with English summary). [Google Scholar]
- BirdLife. European Bird Populations, Trends and National Responsibilities; BirdLife International: Cambridge, UK, 2017. [Google Scholar]
- PECMBS. Trends Common Birds in Europe, 2021 Update. Available online: https://pecbms.info/trends-and-indicators/species-trends/ (accessed on 12 September 2022).
- Chodkiewicz, T.; Kuczyński, L.; Sikora, A.; Chylarecki, P.; Neubauer, G.; Ławicki, Ł.; Stawarczyk, T. Population estimates of breeding birds in Poland in 2008–2012. Ornis Pol. 2015, 56, 149–189, (In Polish with English summary). [Google Scholar]
- Chodkiewicz, T.; Neubauer, G.; Bobrek, R.; Chylarecki, P. The report under Article 12 of the Birds Directive in Poland for the period 2013–2018: Status, trends, threats. Biul. Monit. Przyr. 2019, 20, 46, (In Polish with English summary). [Google Scholar] [CrossRef]
- Bukaciński, D.; Keller, M.; Buczyński, A.; Bukacińska, M. Breeding avifauna of the Middle Vistula riverbed in 2009: The variation in species number and distribution in the last 36 years. In Birds of the Middle Vistula River; Keller, M., Kot, H., Dombrowski, A., Rowiński, P., Chmielewski, S., Bukaciński, D., Eds.; Mazowiecko-Świętokrzyskie Towarzystwo Ornitologiczne: Pionki, Poland, 2017; pp. 671–682, (In Polish with English summary). [Google Scholar]
- Elas, M.; Meissner, W. High density of breeding common sandpipers Actitis hypoleucos in the middle Vistula river, Poland. Wader Study 2019, 126, 67–68. [Google Scholar] [CrossRef]
- Holland, P.K.; Robson, J.E.; Yalden, D.W. The breeding biology of the Common Sandpiper Actitis hypoleucos in the Peak District. Bird Study 1982, 29, 99–110. [Google Scholar] [CrossRef]
- Holland, P.K.; Yalden, D.W. Population dynamics of common sandpipers Actitis hypoleucos breeding along an upland river system. Bird Study 1991, 38, 151–159. [Google Scholar] [CrossRef]
- Mee, A.; Whitfield, D.P.; Thompson, D.B.A.; Burke, T. Extrapair paternity in the Common Sandpiper, Actitis hypoleucos, revealed by DNA fingerprinting. Anim. Behav. 2004, 67, 333–342. [Google Scholar] [CrossRef]
- Kajtoch, Ł.; Figarski, T. Bird Study Short-term restoration of riverine bird assemblages after a severe flood. Bird Study 2013, 60, 327–334. [Google Scholar] [CrossRef]
- Kajtoch, Ł.; Piestrzyńska-Kajtoch, A. An assessment of the impact of environmental changes on two riverine bird species. Acta Zoöl. Crac. 2017, 59, 163–175. [Google Scholar] [CrossRef]
- Claassen, A.H. Breeding Ecology and Conservation of Ground-Nesting Waterbirds in North America and Southeast Asia. Ph.D. Thesis, University of Minnesota, Minneapolis, MN, USA, 2016. [Google Scholar]
- Claassen, A.H.; Forester, J.D.; Arnold, T.W.; Cuthbert, F.J. Consequences of multiscale habitat selection on reproductive success of riverine sandbar-nesting birds in Cambodia. Avian Biol. Res. 2018, 11, 108–122. [Google Scholar] [CrossRef]
- Espie, R.H.M.; James, P.C.; Brigham, R.M. The effects of flooding on piping plover Charadrius melodus reproductive success at Lake Diefenbaker, Saskatchewan, Canada. Biol. Conserv. 1998, 86, 215–222. [Google Scholar] [CrossRef]
- Dombrowski, A.; Kot, H.; Bukaciński, D. Threats to birds of the Middle Vistula River and their habitats. In Birds of the Middle Vistula River; Keller, M., Kot, H., Dombrowski, A., Rowiński, P., Chmielewski, S., Bukaciński, D., Eds.; Mazowiecko-Świętokrzyskie Towarzystwo Ornitologiczne: Pionki, Poland, 2017; pp. 671–682, (In Polish with English summary). [Google Scholar]
- Bukaciński, D.; Bukacińska, M.; Buczyński, A. Threats and the active protection of birds in a riverbed: Postulates for the strategy of the preservation of the middle Vistula River avifauna. Stud. Ecol. Bioethicae 2018, 16, 5–23. [Google Scholar] [CrossRef]
- Royan, A.; Prudhomme, C.; Hannah, D.M.; Reynolds, S.J.; Noble, D.G.; Sadler, J.P. Climate-induced changes in river flow regimes will alter future bird distributions. Ecosphere 2015, 6, 50. [Google Scholar] [CrossRef]
- Finch, D.M. House wrens adjust lying dates and clutch size in relation to annual flooding. Wilson Bull. 1991, 103, 25–43. [Google Scholar]
- Zhang, W.; Liu, T.; Cheng, K.; Rummy, P. Declining water depth delayed the breeding time of Fulica atra, not human disturbance. PLoS ONE 2018, 13, e0202684. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, J.; Liu, J.; Yang, C.; Liang, W.; Møller, A.P. Adaptation or ecological trap? Altered nest-site selection by Reed Parrotbills after an extreme flood. Avian Res. 2019, 10, 4–11. [Google Scholar] [CrossRef]
- Plaschke, S.; Bulla, M.; Cruz-López, M.; del Ángel, S.G.; Küpper, C. Nest initiation and flooding in response to season and semi-lunar spring tides in a ground-nesting shorebird. Front. Zool. 2019, 16, 15. [Google Scholar] [CrossRef] [PubMed]
- Dankers, R.; Feyen, L. Climate change impact on flood hazard in Europe: An assessment based on high-resolution climate simulations. J. Geophys. Res. Atmos. 2008, 113, D19105. [Google Scholar] [CrossRef]
- UNDRR. The Human Cost of Disasters: An Overview of the Last 20 Years (2000–2019); UNDRR: Geneva, Switzerland, 2020. [Google Scholar]
- World Meteorological Organization. Atlas of Mortality and Economic Losses from Weather, Climate and Water Extremes (1970–2019); World Meteorological Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Bailey, L.D.; Ens, B.J.; Both, C.; Heg, D.; Oosterbeek, K.; van de Pol, M. No phenotypic plasticity in nest-site selection in response to extreme flooding events. Philos. Trans. R Soc. B Biol. Sci. 2017, 372, 20160139. [Google Scholar] [CrossRef] [PubMed]
- Kajak, Z. The Vistula river and its riparian zones. Hydrobiologia 1993, 251, 149–157. [Google Scholar] [CrossRef]
- Wrzesinski, D.; Sobkowiak, L. Transformation of the flow regime of a large allochthonous river in central Europe-an example of the Vistula River in Poland. Water 2020, 12, 507. [Google Scholar] [CrossRef]
- Gacka-Grześkiewicz, E. Vistula as an Ecological Corridor. State-Functioning-Threats; Fundacja IUCN Poland: Warsaw, Poland, 1995; (In Polish with English summary). [Google Scholar]
- Matuszkiewicz, J.; Roo-Zielińska, E. (Eds.) Inter-Embankment of Vistula River as a Peculiar Natural System: Section Pilica—Narew; Polish Academy of Sciences: Warsaw: Poland, 2000; (In Polish with English summary). [Google Scholar]
- Bibby, C.J.; Burgess, N.D.; Hill, D.A.; Mustoe, S.H. Bird Census Techniques; Academic Press: London, UK, 2000. [Google Scholar]
- Elas, M.; Kajzer, K.; Grzębkowski, M.; Koliński, A.; Różycki, A.; Sikora, D.; Wardecki, Ł.; Węgrzynowicz, A. Assessment of the number of breeding pairs of the Common Sandpiper Actitis hypoleucos depending of the survey method. Ornis Pol. 2015, 56, 212–219, (In Polish with English summary). [Google Scholar]
- Colwell, M.A. Egg-laying Intervals in Shorebirds. Wader Study Group Bull. 2006, 111, 50–59. [Google Scholar]
- Liebezeit, J.R.; Smith, P.A.; Lanctot, R.B.; Schekkerman, H.; Tulp, I.; Kendall, S.J.; Tracy, D.M.; Rodrigues, R.J.; Meltofte, H.; Robinson, J.A.; et al. Assessing the development of shorebird eggs using the flotation method: Species-specific and generalized regression models. Condor 2007, 109, 32–47. [Google Scholar] [CrossRef]
- Mee, A. Reproductive strategies in the common sandpiper Actitis hypoleucos. Ph.D. Thesis, Department of Animal & Plant Sciences, University of Sheffield, Sheffield, UK, 2001. [Google Scholar]
- Holland, P.K.; Yalden, D.W. Population dynamics of common sandpipers Actitis hypoleucos in the Peak District of Derbyshire—A different decade. Bird Study 2002, 49, 131–138. [Google Scholar] [CrossRef]
- Quantum GIS Development Team. QGIS Geographic Information System. QGIS Association. 2020. Available online: https://www.qgis.org (accessed on 1 May 2021).
- Kochanek, K.; Karamuz, E.; Osuch, M. Distributed modelling of flow in the middle reach of the river Vistula. GeoPlanet Earth Planet Sci. 2015, 16, 83–107. [Google Scholar] [CrossRef]
- Bewick, V.; Cheek, L.; Ball, J. Statistics review 10: Further nonparametric methods. Crit. Care 2004, 8, 196–199. [Google Scholar] [CrossRef]
- Pohlert, T. Package PMCMRplus. 2022. Available online: https://CRAN.R-project.org/package=PMCMRplus (accessed on 30 April 2022).
- Kotu, V.; Deshpande, B. Clustering. In Data Science; Kotu, V., Deshpande, B., Eds.; Morgan Kaufmann: Burlington, MA, USA, 2019; pp. 221–261. [Google Scholar] [CrossRef]
- Whitley, E.; Ball, J. Statistics review 6: Nonparametric methods. Crit. Care 2002, 6, 509–513. [Google Scholar] [CrossRef]
- Johnson, R.A.; Wichern, D.W. Applied Multivariate Statistical Analysis; Pearson Prentice Hall: Upper Saddle River, NJ, USA, 2014. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org (accessed on 30 April 2022).
- Byrd, R.H.; Lu, P.; Nocedal, J.; Zhu, C. A limited memory algorithm for bound constrained optimization. SIAM J. Sci. Comput. 1995, 16, 1190–1208. [Google Scholar] [CrossRef]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Burnham, K.; Anderson, D. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach; Springer: New York, NY, USA, 2002. [Google Scholar]
- Bissolli, P.; Friedrich, K.; Rapp, J.; Ziese, M. Flooding in eastern central Europe in May 2010—Reasons, evolution and climatological assessment. Weather 2011, 66, 147–153. [Google Scholar] [CrossRef]
- Nilsson, C.; Reidy, C.A.; Dynesius, M.; Revenga, C. Fragmentation and flow regulation of the world’s large river systems. Science 2005, 308, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Belletti, B.; Garcia de Leaniz, C.; Jones, J.; Bizzi, S.; Börger, L.; Segura, G.; Castelletti, A.; Van de Bund, W.; Aarestrup, K.; Barry, J.; et al. More than one million barriers fragment Europe’s rivers. Nature 2020, 588, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, C.; Berggren, K. Alterations of riparian ecosystems caused by river regulation. BioScience 2000, 50, 571–584. [Google Scholar] [CrossRef]
- Tonkin, J.D.; Merritt, D.M.; Olden, J.D.; Reynolds, L.V.; Lytle, D.A. Flow regime alteration degrades ecological networks in riparian ecosystems. Nat. Ecol. Evol. 2018, 2, 86–93. [Google Scholar] [CrossRef]
- Coumou, D.; Rahmstorf, S. A decade of weather extremes. Nat. Clim. Change 2012, 2, 491–496. [Google Scholar] [CrossRef]
- Magnuszewski, A.; Gutry-Korycka, M. Flood waters flow in the contemporary Vistula River channel. Pr. Stud. Geogr. 2009, 43, 153–162, (In Polish with English summary). [Google Scholar]
- Matuszkiewicz, J.; Chojnacki, J.; Kozłowska, A.; Plitt, J.; Roo-Zielińska, E. The Typological, Spatial and Dynamic Differentiation of the Area between the Floodbanks along the Warsaw Stretch of the Vistula In Inter-Embankment of Vistula River as a Peculiar Natural System: (Section Pilica—Narew); Matuszkiewicz, J., Roo-Zielińska, E., Eds.; Polish Academy of Sciences: Warsaw, Poland, 2000; pp. 31–78, (In Polish with English summary). [Google Scholar]
- Spurr, E.B.; Ledgard, N.J. Population trends of braided river birds on the Ashley River. Nothornis 2016, 63, 73–86. [Google Scholar]
- Knutson, M.G.; Klaas, E.E. Declines in abundance and species richness of birds following a major flood on the Upper Mississippi River. Auk 1997, 114, 367–380. [Google Scholar] [CrossRef]
- Reiley, B.M.; Benson, T.J.; Everitts, J.; Bednarz, J.C. Does flooding effect the apparent survival and body condition of a ground foraging migrant passerine. PLoS ONE 2017, 12, e0175179. [Google Scholar] [CrossRef]
- Royan, A.; Hannah, D.M.; Reynolds, S.J.; Noble, D.G.; Sadler, J.P. River birds’ response to hydrological extremes: New vulnerability index and conservation implications. Biol. Conserv. 2014, 177, 64–73. [Google Scholar] [CrossRef]
- Kozik, R.; Meissner, W.; Listewnik, B.; Nowicki, J.; Lasecki, R. Differences in foraging behaviour of a migrating shorebird at stopover sites on regulated and unregulated sections of a large European lowland river. J. Ornithol. 2022, 163, 791–802. [Google Scholar] [CrossRef]
- Kaznowska, E.; Hejduk, A.; Kempiński, C. The Vistula River low flows in Warsaw in the 21st century. Acta Sci. Pol. 2018, 17, 33–43. [Google Scholar] [CrossRef]
- Kokko, H. Competition for early arrival in migratory birds. J. Anim. Ecol. 1999, 68, 940–950. [Google Scholar] [CrossRef]
- Morrison, C.A.; Alves, J.A.; Gunnarsson, T.G.; Þórisson, B.; Gill, J.A. Why do earlier-arriving migratory birds have better breeding success? Ecol. Evol. 2019, 9, 8856–8864. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Wallander, J.; Oring, L.; Akst, E.; Reed, J.M.; Fleischer, R.C. Adaptive seasonal trend in brood sex ratio: Test in two sister species with contrasting breeding systems. J. Evol. Biol. 2003, 16, 510–515. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).