
Molecular and Morphological Characterization of Two New Species of Globisporangium from Southern China, G. pengfuense and G. tenuihyphum



Abstract
1. Introduction
2. Materials and Methods
2.1. Isolates
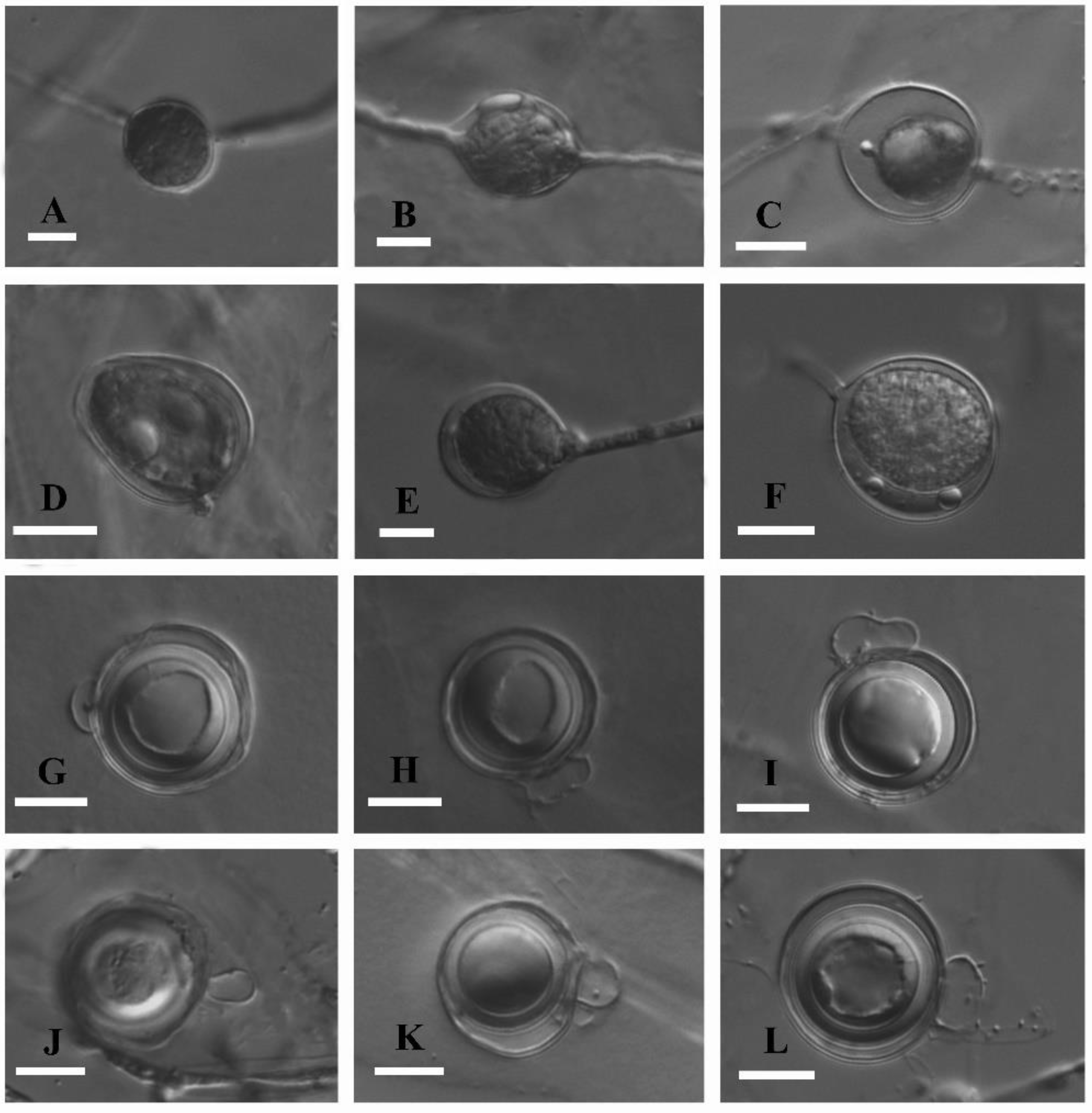
2.2. Morphology and Growth Rate
2.3. DNA Extraction, PCR and Sequencing
2.4. Phylogenetic Analyses
2.5. Pathogenicity
3. Results
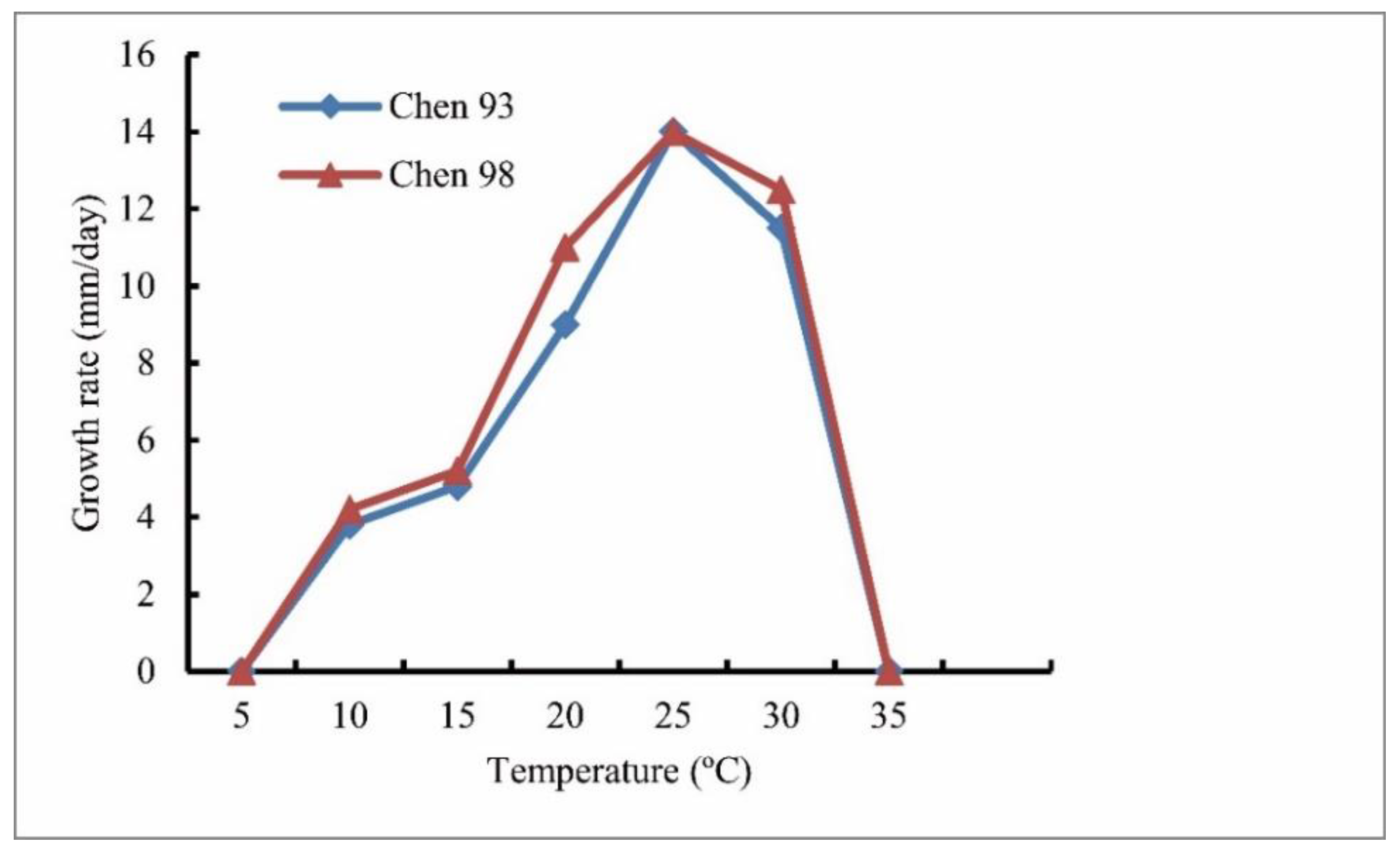
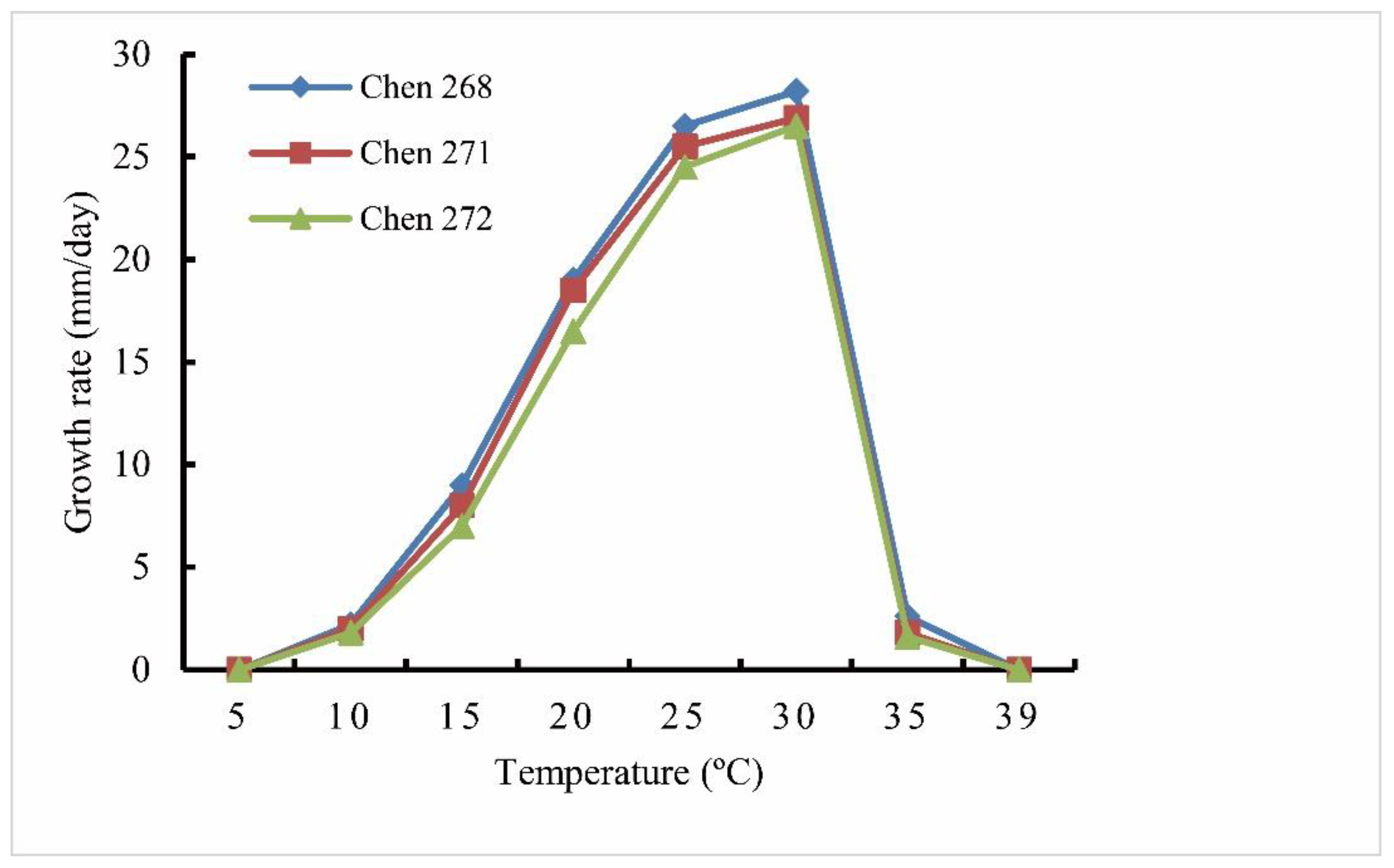
3.1. Isolates
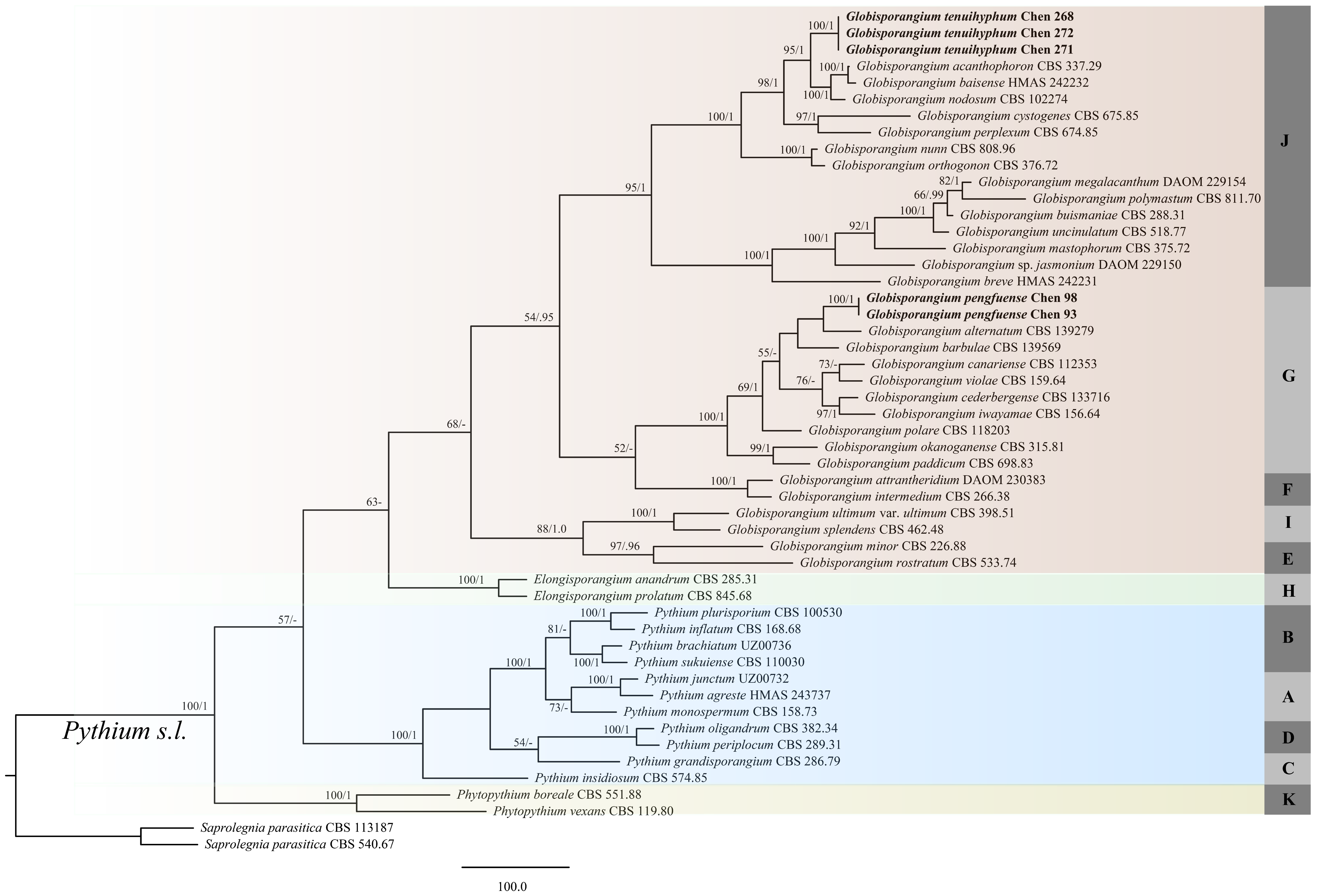
3.2. Phylogenetic Analyses
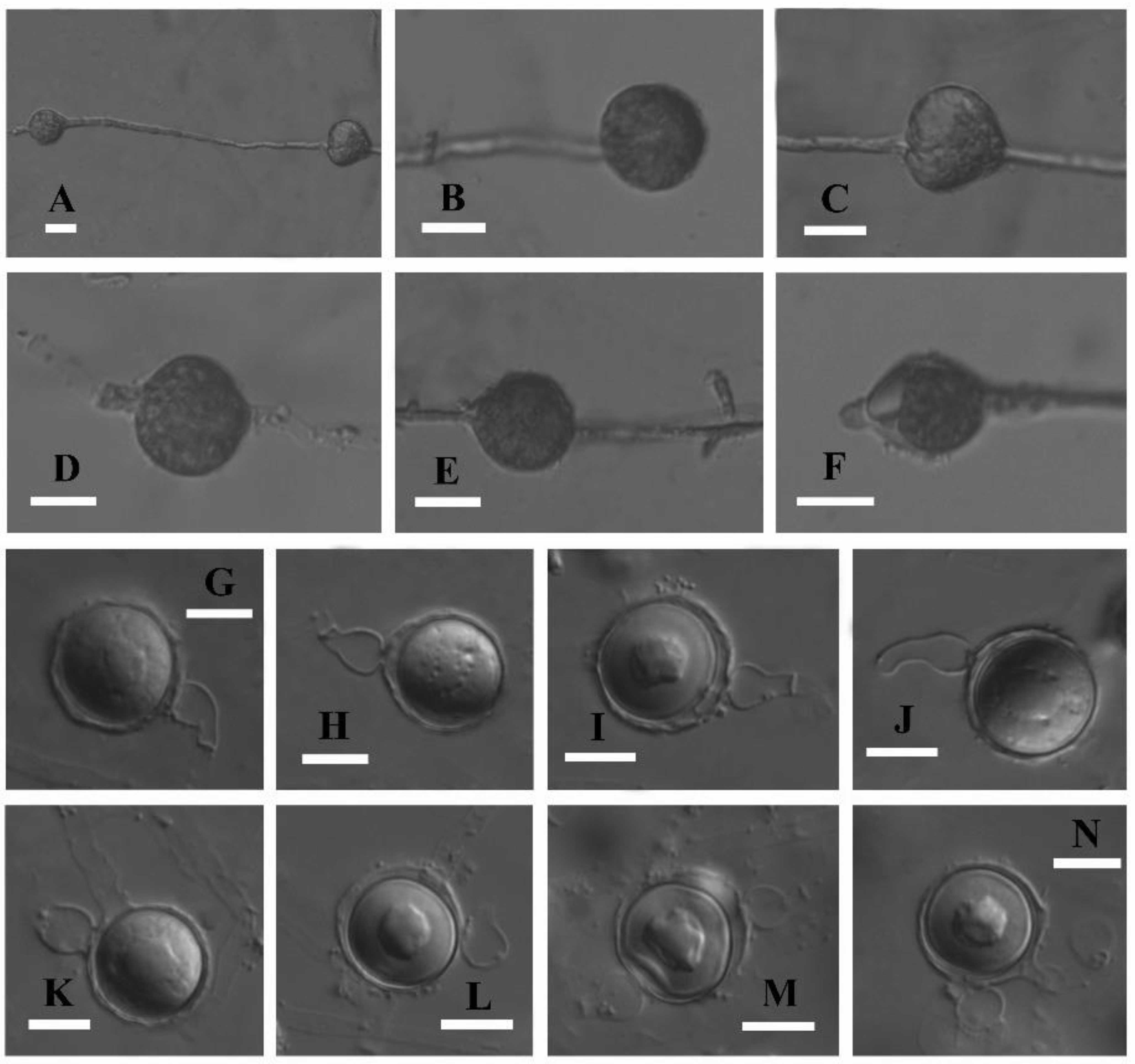
3.3. Taxonomy
3.4. Pathogenicity
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Pringsheim, N. Beiträge zur morphology and systematik der algen. 2. Die Saprolegníeen [Contributions to the morphology and systematics of algae]. Jb. Wiss. Bot. 1858, 1, 284–306. [Google Scholar]
- van der Plaäts-Niterink, A.J. Monograph of the genus Pythium. Stud. Mycol. 1981, 21, 242. [Google Scholar]
- Uzuhashi, S.; Okada, G.; Ohkuma, M. Four new Pythium species from aquatic environments in Japan. Anton. Leeuw. Int. J. G 2015, 107, 375–391. [Google Scholar] [CrossRef] [PubMed]
- Ueta, S.; Tojo, M. Pythium barbulae sp. nov. isolated from the moss, Barbula unguiculata; morphology, molecular phylogeny and pathogenicity. Mycoscience 2016, 57, 11–19. [Google Scholar] [CrossRef]
- Salmaninezhad, F.; Mostowfizadeh-Ghalamfarsa, R. Three new Pythium species from rice paddy fields. Mycologia 2019, 111, 274–290. [Google Scholar] [CrossRef]
- Chen, J.J.; Feng, H.; Yu, J.; Ye, W.W.; Zheng, X.B. Pythium huanghuaiense sp. nov. isolated from soybean: Morphology, molecular phylogeny and pathogenicity. Biodivers. Data J. 2021, 9, e65227. [Google Scholar] [CrossRef]
- Ali-Shtayeh, M.S.; Saleh, A.S.F. Isolation of Pythium acanthicum, P. oligandrum, and P. periplocum from soil and evaluation of their mycoparasitic activity and biocontrol efficacy against selected phytopathogenic Pythium species. Mycopathologia 1999, 145, 143–153. [Google Scholar] [CrossRef]
- Lévesque, C.A.; de Cock, A.W.A.M. Molecular phylogeny and taxonomy of the genus Pythium. Mycol. Res. 2004, 108, 1363–1383. [Google Scholar] [CrossRef]
- Nguyen, H.D.T.; Dodge, A.; Dadej, K.; Rintoul, T.L.; Ponomareva, E.; Martin, F.N.; de Cock, A.W.A.M.; Lévesque, C.A.; Redhead, S.A.; Spies, C.F.J. Whole genome sequencing and phylogenomic analysis show support for the splitting of genus Pythium. Mycologia 2022, 114, 501–515. [Google Scholar] [CrossRef]
- Miller, P.M. V-8 juice agar as a general purpose medium for fungi and bacteria. Phytopathology 1955, 145, 461–462. [Google Scholar]
- Cui, B.K.; Li, H.J.; Ji, X.; Zhou, J.L.; Song, J.; Si, J.; Yang, Z.L.; Dai, Y.C. Species diversity, taxonomy and phylogeny of Polyporaceae (Basidiomycota) in China. Fungal Divers. 2019, 97, 137–392. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gefand, D.H., Sninsky, J.J., White, J.T., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Robideau, G.P.; de Cock, A.W.; Coffey, M.D.; Voglmayr, H.; Brouwer, H.; Bala, K.; Chitty, D.W.; Désaulniers, N.; Eggertson, Q.A.; Gachon, C.M.; et al. DNA barcoding of oomycetes with cytochrome c oxidase subunit I and internal transcribed spacer. Mol. Ecol. Resour. 2011, 11, 1002–1011. [Google Scholar] [CrossRef]
- Rahman, M.Z.; Uematsu, S.; Coffey, M.D.; Uzuhashi, S.; Suga, H.; Kageyama, K. Re-evaluation of Japanese Phytophthora isolates based on molecular phylogenetic analyses. Mycoscience 2014, 55, 314–327. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The Clustal_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Hall, T.A. Bioedit: A user–friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Villa, N.O.; Kageyama, K.; Asano, T.; Suga, H. Phylogenetic relationships of Pythium and Phytophthora species based on ITS rDNA, cytochrome oxidase II and tubulin gene sequences. Mycologia 2006, 98, 410–422. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP: Phylogenetic Analysis Using Parsimony Version 4.0b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Felsenstein, J. Confidence intervals on phylogenetics: An approach using bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Mark, P.; Avres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes3.2: Efficient Bayesian phylogenetic inference and model choice, across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Dorrance, A.E.; Berry, S.A.; Anderson, T.R.; Meharg, C. Isolation, storage, pathotype characterization, and evaluation of resistance for Phytophthora sojae in soybean. Plant health Prog. 2008, 9, 35. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W. Basic local alignment search tool. J. Mol. Biol. 1990, 25, 403–410. [Google Scholar] [CrossRef]
- Long, Y.Y.; Wei, J.G.; Sun, X.; He, Y.Q.; Luo, J.T.; Guo, L.D. Two new Pythium species from China based on the morphology and DNA sequence data. Mycol. Prog. 2012, 11, 689–698. [Google Scholar] [CrossRef]
- Ho, H.H. The genus Pythium in mainland China. Mycosystema 2013, 32, 20–44. [Google Scholar]
- Rahman, M.H.; Abdelzaher, H.M.A.; Mingzhu, L.; Motohashi, K.; Suga, H.; Kageyama, K. Pythium rishiriense sp. nov. from water and P. alternatum sp. nov. from soil, two new species from Japan. FEMS Microbiol. Lett. 2015, 362, fnv086. [Google Scholar]
- Paul, B. ITS region of Pythium canariense sp. nov., its morphology and its interaction with Botrytis cinerea. FEMS Microbiol. Lett. 2002, 208, 135–141. [Google Scholar] [CrossRef][Green Version]
- Bahramisharif, A.; Lamprecht, S.C.; Spies, C.F.J.; Botha, W.J. Pythium cederbergense sp. nov. and related taxa from Pythium clade G associated with the South African indigenous plant Aspalathus linearis (rooibos). FEMS Microbiol. Lett. 2013, 105, 1174–1189. [Google Scholar]
- Paul, B.; Galland, D.; Bhatnagar, T.; Dulieu, H. A new species of Pythium isolated from the Burgundy region in France. FEMS Microbiol. Lett. 1998, 158, 207–213. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Sample No. | Locality | GenBank Accession No. | |
|---|---|---|---|---|
| ITS | Cox1 | |||
| Elongisporangium anandrum | CBS 285.31 | – | HQ643435 | HQ708482 |
| E. prolatum | CBS 845.68 | USA | HQ643754 | HQ708795 |
| Globisporangium acanthophoron | CBS 337.29 | USA | HQ643413 | HQ708460 |
| G. alternatum | CBS 139279 | Japan | AB998876 | AB998877 |
| G. attrantheridium | DAOM 230383 | Canada | HQ643477 | HQ708524 |
| G. baisense | HMAS 242232 | China | FR775440 | FR774198 |
| G. barbulae | CBS 139569 | Japan | LC028389 | LC028392 |
| G. buismaniae | CBS 288.31 | Netherlands | HQ643479 | HQ708526 |
| G. canariense | CBS 112353 | Spain | HQ643482 | HQ708528 |
| G. cederbergense | CBS 133716 | South Africa | JQ412768 | JQ412793 |
| G. cystogenes | CBS 675.85 | Netherlands | HQ643518 | HQ708564 |
| G. intermedium | CBS 266.38 | Netherlands | HQ643572 | HQ708616 |
| G. iwayamae | CBS 156.64 | Australia | HQ643669 | HQ708713 |
| G. mastophorum | CBS 375.72 | United Kingdom | HQ643691 | HQ708735 |
| G. megalacanthum | DAOM 229154 | Germany | HQ643693 | HQ708737 |
| G. minor | CBS 226.88 | United Kingdom | HQ643696 | HQ708740 |
| G. nodosum | CBS 102274 | France | HQ643709 | HQ708753 |
| G. nunn | CBS 808.96 | USA | HQ643711 | HQ708755 |
| G. okanoganense | CBS 315.81 | USA | HQ643714 | HQ708758 |
| G. orthogonon | CBS 376.72 | Lebanon | HQ643723 | HQ708764 |
| G. paddicum | CBS 698.83 | Japan | HQ643728 | HQ708769 |
| G. pengfuense | Chen 93 | China | MF984129 | MF984166 |
| G. pengfuense | Chen 98 | China | MF984130 | MF984167 |
| G. perplexum | CBS 674.85 | Netherlands | HQ643744 | HQ708785 |
| G. polare | CBS 118203 | Japan | KJ716859 | – |
| G. polymastum | CBS 811.70 | Netherlands | HQ643752 | HQ708793 |
| G. rostratum | CBS 533.74 | Netherlands | HQ643767 | HQ708808 |
| G. sp. jasmonium | DAOM 229150 | USA | HQ643670 | HQ708714 |
| G. splendens | CBS 462.48 | USA | HQ643795 | HQ708836 |
| G. tenuishyphum | Chen 268 | China | MF984123 | MF984160 |
| G. tenuishyphum | Chen 271 | China | MF984124 | MF984161 |
| G. tenuishyphum | Chen 272 | China | MF984125 | MF984162 |
| G. ultimum var. ultimum | CBS398.51 | Netherlands | HQ643865 | HQ708906 |
| G. uncinulatum | CBS 518.77 | Netherlands | HQ643944 | HQ708985 |
| G. violae | CBS 159.64 | Australia | HQ643958 | HQ708999 |
| Phytopythium boreale | CBS 551.88 | China | AB725879 | AB690647 |
| Ph. vexans | CBS 119.80 | Iran | HQ643400 | HQ708447 |
| Pythium agreste | HMAS 243737 | China | HE862395 | HE862399 |
| P. brachiatum | UZ00736 | Japan | KJ995581 | KJ995593 |
| P. breve | HMAS 242231 | China | FR751317 | FR774196 |
| P. grandisporangium | CBS 286.79 | USA | HQ643546 | HQ708590 |
| P. inflatum | CBS 168.68 | USA | HQ643566 | HQ708610 |
| P. insidiosum | CBS 574.85 | Costa Rica | HQ643570 | HQ708614 |
| P. junctum | UZ00732 | Japan | KJ995576 | KJ995595 |
| P. monospermum | CBS 158.73 | United Kingdom | HQ643697 | HQ708741 |
| P. oligandrum | CBS 382.34 | United Kingdom | HQ643715 | HQ708759 |
| P. plurisporium | CBS 100530 | USA | HQ643749 | HQ708790 |
| P. periplocum | CBS 289.31 | USA | HQ643743 | HQ708784 |
| P. sukuiense | CBS 110030 | Taiwan | HQ643836 | HQ708877 |
| Saprolegnia parasitica | CBS 113187 | Russia | HQ644005 | HQ709046 |
| S. parasitica | CBS 540.67 | United Kingdom | HQ644000 | HQ709041 |
| G.pengfuense (Chen 93) | G. alternatum | G. barbulae | G. canariense | G. cederbergense | G. iwayamae | G. violae | |
|---|---|---|---|---|---|---|---|
| Width of Hyphae (μm) | Up to 5 | Up to 4.5 | Up to 7 | Up to 7 | Unknown | Up to 6.6 | Up to 6 |
| Sporangia/hyphal swellings | Globose to sub-globose, catenulate, terminal, occasionally with apical papillae or intercalary | Globose to sub-globose, intercalary or terminal | Globose or subglobose, terminal or intercalary | Spherical, globose, subglobose, pyriform, pea-nut-shaped, dumb-bell-shaped, and elongated; some with papilla-like projections, usually intercalary, catenulate and at times terminal | Globose, terminal | Globose, ellipsoidal, ovoid or limoniform, terminal | Terminal and intercalary, |
| Oogonia (μm) | 12.5–22.5 (av. 17.5), terminal or intercalary | 13–22 (av. 18), terminal or intercalary | 12.5–18 (av. 15.3), terminal and rarely intercalary | 19–30 (av. 25.9), terminal, rarely intercalary | 17–29 (av. 23), mostly terminal, sometimes intercalary | 23–29 (av. 27), terminal or intercalary, | 25–38 (av. 29.5), terminal or intercalary |
| Oogonium ornamentation | Absent | Absent | Absent | Absent | Mostly smooth, some ornamented with short, conical papillae | Mostly smooth, some ornamented with short, conical papillae | Absent |
| Antheridia | Mostly monoclinous, occasionally diclinous | Monoclinous or diclinous, occasionally on short pedicel or sometimes hypogynous | Mostly monoclinous, sometimes diclinous | Mostly monoclinous, sometimes diclinous | Monoclinous | Monoclinous or diclinous | Mostly monoclinous |
| Oospores (μm) | Plerotic or nearly plerotic, 10.5–21.5 (av. 16.5) | Mostly aplerotic, 10–18 (av. 15) | Plerotic or nearly plerotic, 11.1–16.5 (av. 13.9) | Plerotic, 17–29 (av. 20.7) | Mainly aplerotic to almost plerotic, 15–27 (av. 21) | Plerotic or aplerotic, 19–24 (av. 22) | Aplerotic, 22–32 (av. 27) |
| Oospore wall thickness (μm) | 0.5–1.0 | Unknown | 0.3– 1.3 | 2–4 | 2–4 | Unknown | Up to 3 |
| Cardinal temperature | Min 5 °C, optimum 25 °C, and max 35 °C | Min 5 °C, optimum 25 °C, and max 30 °C | Min 0 °C, optimum 28 °C, and max 34 °C | Unknown | Min 3–5 °C, optimum 27–30 °C, and max 37 °C | Min 5 °C, optimum 25 °C, and max 30 °C | Min 5 °C, optimum 25 °C, and max 35 °C |
| Daily growth rates on PCA at 25 °C | 14 | 8 | 13.4 | 14–15 | 24 | 14 | 15 |
| Reference | This study | [26] | [4] | [27] | [28] | [2] | [2] |
| G.tenuihyphum (Chen 268) | G. acanthophoron (Sideris) Uzuhashi, Tojo and Kakish. | G. baisense (Y.Y. Long, J.G. Wei and L.D. Guo) H.D.T. Nguyen and C.F.J. Spies | G. cystogenes (De Cock and Lévesque) Uzuhashi, Tojo and Kakish. | G. nodosum (B. Paul, D. Galland, T. Bhatn. and Dulieu) Uzuhashi, Tojo and Kakish. | G. perplexum (H. Kouyeas and Theoh.) Uzuhashi, Tojo and Kakish. | |
|---|---|---|---|---|---|---|
| Width of Hyphae (μm) | Up to 4 | Up to 7 | Up to 6.3 | Up to 7 | Up to 7 | Unknown |
| Sporangia | Mostly sub-globose to globose, occasionally ovoid-obpyriform, terminal or intercalary | Unknown | Globose, subglobose to elongate, at times with a papilla, terminal or intercalary | Globose, subglobose, ovoid, often provided with a more or less distinct papilla, terminal, occasionally intercalary | Globose, subglobose, ovoid, pyriform and at times peanut-shaped, mostly intercalary and catenulate, at times terminal | Globose, elongated, terminal or intercalary, occasionally in series |
| Oogonia (μm) | 20–25 (av. 23.5), terminal | 20–30 | 13.8–28.8 (av. 21.2), terminal or sessile, occasionally lateral | 28–35 (av. 30.7), terminal or laterally stalked, occasionally intercalary | 11–27 (av. 20.8), terminal and intercalary, | 16–27 (av. 20), terminal on lateral branches |
| Oogonium ornamentation | Absent | Present | Absent | Absent | Absent | Absent |
| Antheridia | Monoclinous | Monoclinous or diclinous | Monoclinous and diclinous | Diclinous | Unknown | Mostly monoclinous |
| Oospores (μm) | Aplerotic, 15–21.5 (av. 18.5) | 15–20 | Mostly aplerotic, sometimes nearly plerotic, 11.3–23.8 (av. 18.5) | Aplerotic, 21–26 (av. 23.3) | Aplerotic, 10–21 (av. 16.9) | Aplerotic, 12–20 (av. 16) |
| Oospore wall thickness (μm) | 1.5–2.5 | Up to 1.5 | Unknown | Up to 3 | Unknown | 1.5–1.7 |
| Double oospores | Absent | Unknown | Present | Absent | Present | Absent |
| Cardinal temperature | Min 5 °C, optimum 30 °C, and max 39 °C | Min 5 °C, optimum 30 °C, and max 37 °C | Min 8 °C, optimum 30 °C, and max 39 °C | Min 3–6 °C, optimum 24 °C, and max 27–30 °C | Unknown | Min 4 °C, optimum 22 °C, and max 32 °C |
| Daily growth rates on PCA at 25 °C | 25.5/per day | 20/per day | 30/per day | 17/per day | 17/per day | 12–13/per day |
| Reference | This study | [2] | [24] | [17] | [29] | [2] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, J.-J.; Zong, S.-B.; Huang, X.-X.; Yu, J.; Yang, H.-J.; Ye, W.-W. Molecular and Morphological Characterization of Two New Species of Globisporangium from Southern China, G. pengfuense and G. tenuihyphum. Diversity 2022, 14, 528. https://doi.org/10.3390/d14070528
Chen J-J, Zong S-B, Huang X-X, Yu J, Yang H-J, Ye W-W. Molecular and Morphological Characterization of Two New Species of Globisporangium from Southern China, G. pengfuense and G. tenuihyphum. Diversity. 2022; 14(7):528. https://doi.org/10.3390/d14070528
Chicago/Turabian StyleChen, Jia-Jia, Shu-Bin Zong, Xiao-Xiao Huang, Jian Yu, Hong-Jun Yang, and Wen-Wu Ye. 2022. "Molecular and Morphological Characterization of Two New Species of Globisporangium from Southern China, G. pengfuense and G. tenuihyphum" Diversity 14, no. 7: 528. https://doi.org/10.3390/d14070528
APA StyleChen, J.-J., Zong, S.-B., Huang, X.-X., Yu, J., Yang, H.-J., & Ye, W.-W. (2022). Molecular and Morphological Characterization of Two New Species of Globisporangium from Southern China, G. pengfuense and G. tenuihyphum. Diversity, 14(7), 528. https://doi.org/10.3390/d14070528