
Reproductive Ecology of Distylous Shoreside Polygonum criopolitanum Hance

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Materials
2.2. Study Methods
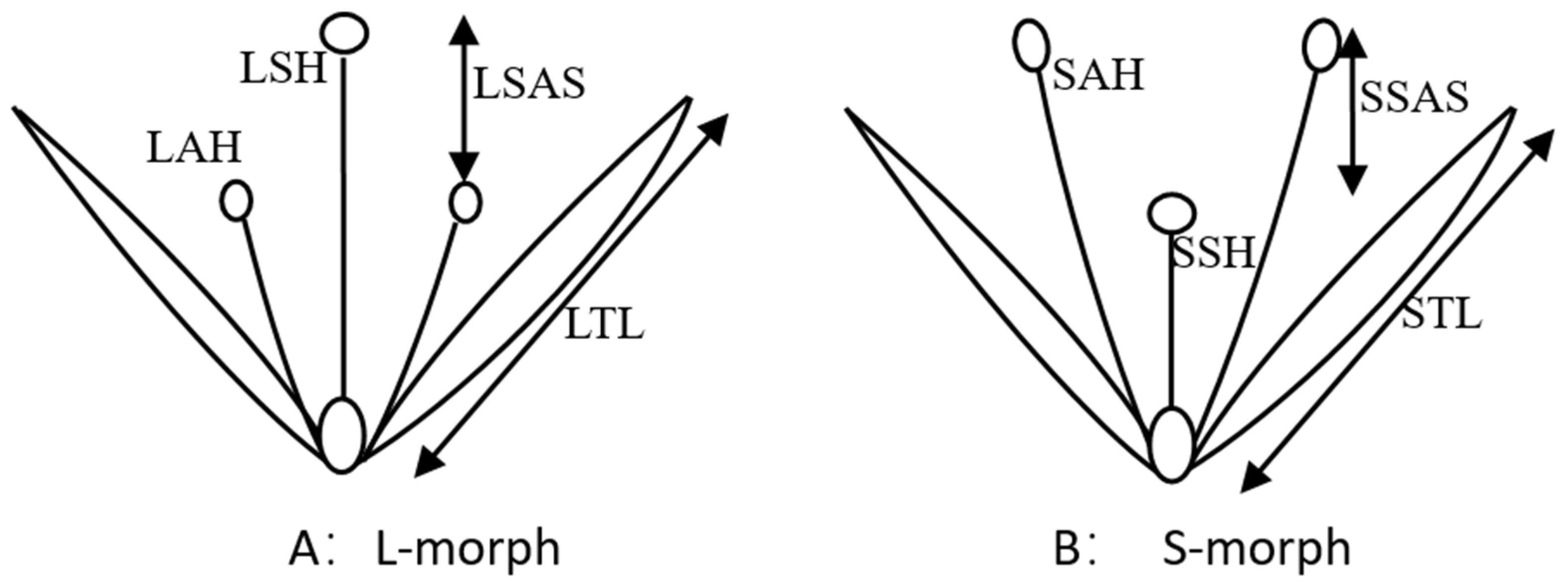
2.2.1. Floral Characters
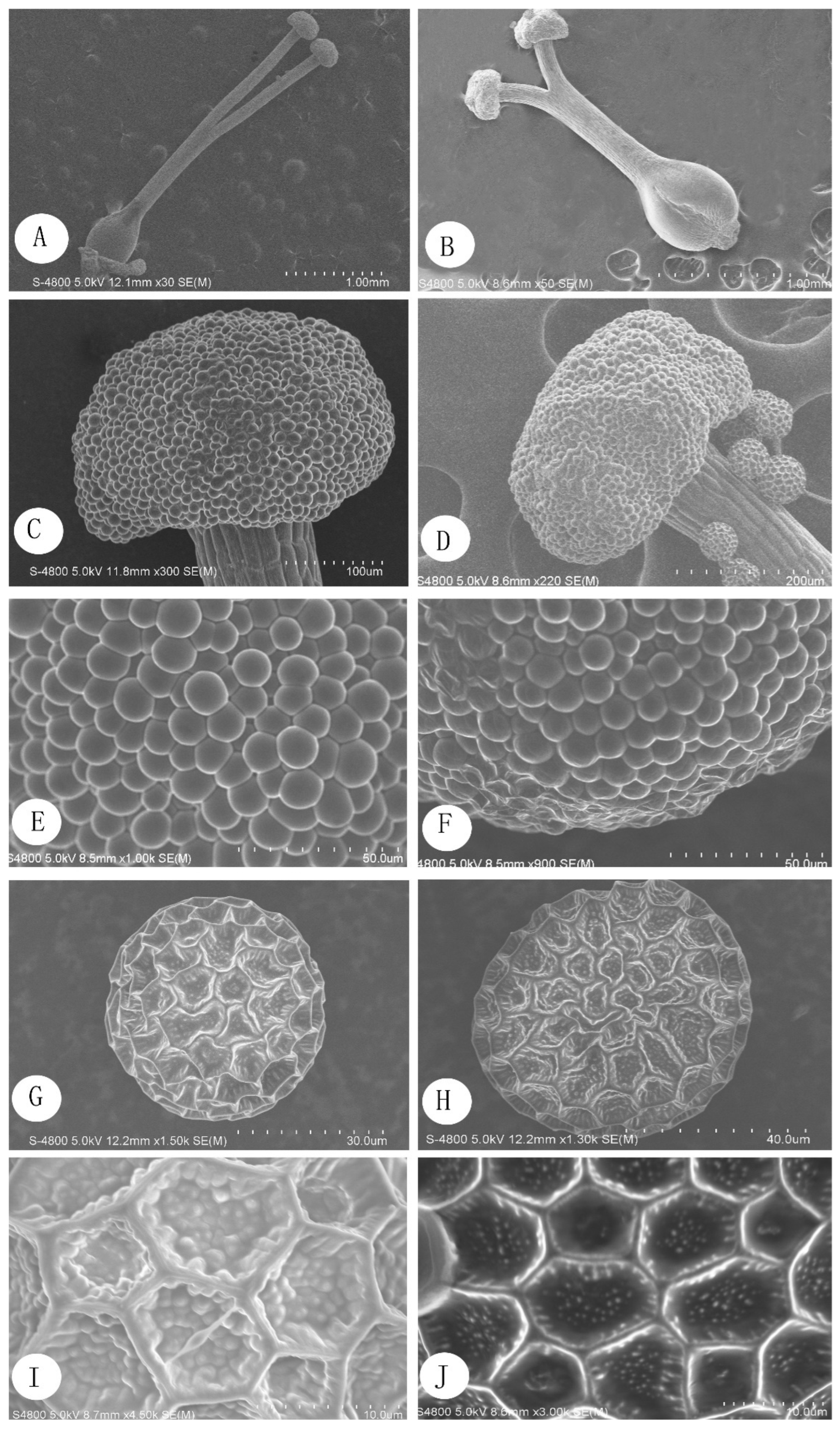
2.2.2. Scanning Electron Microscopy
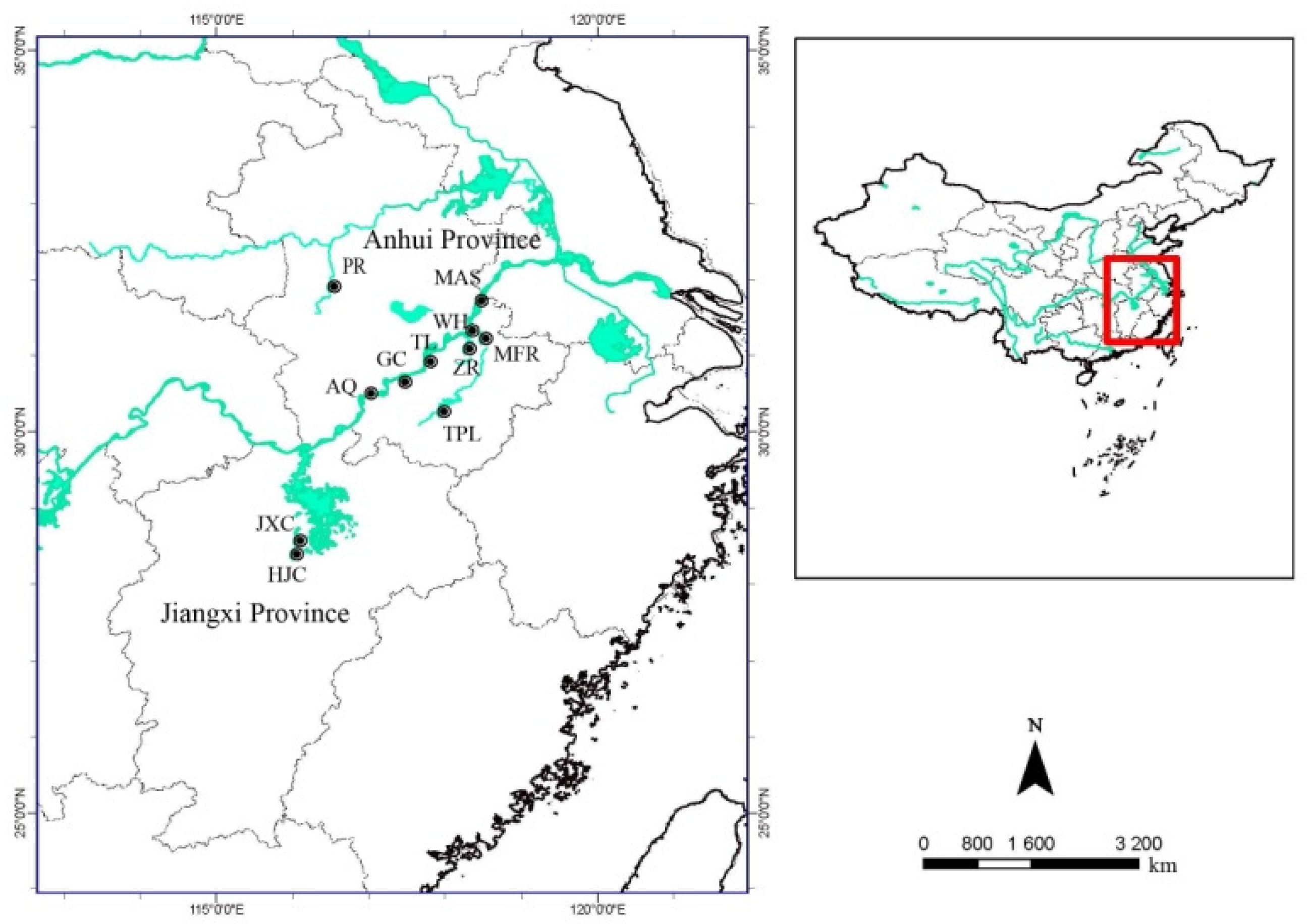
2.2.3. Population Structure Survey
2.2.4. Environmental and Ecological Factors
2.2.5. CCA
2.2.6. Fluorescent Microscopy
2.2.7. Seed Set Experiments
2.2.8. Seed Germination Experiments
3. Results
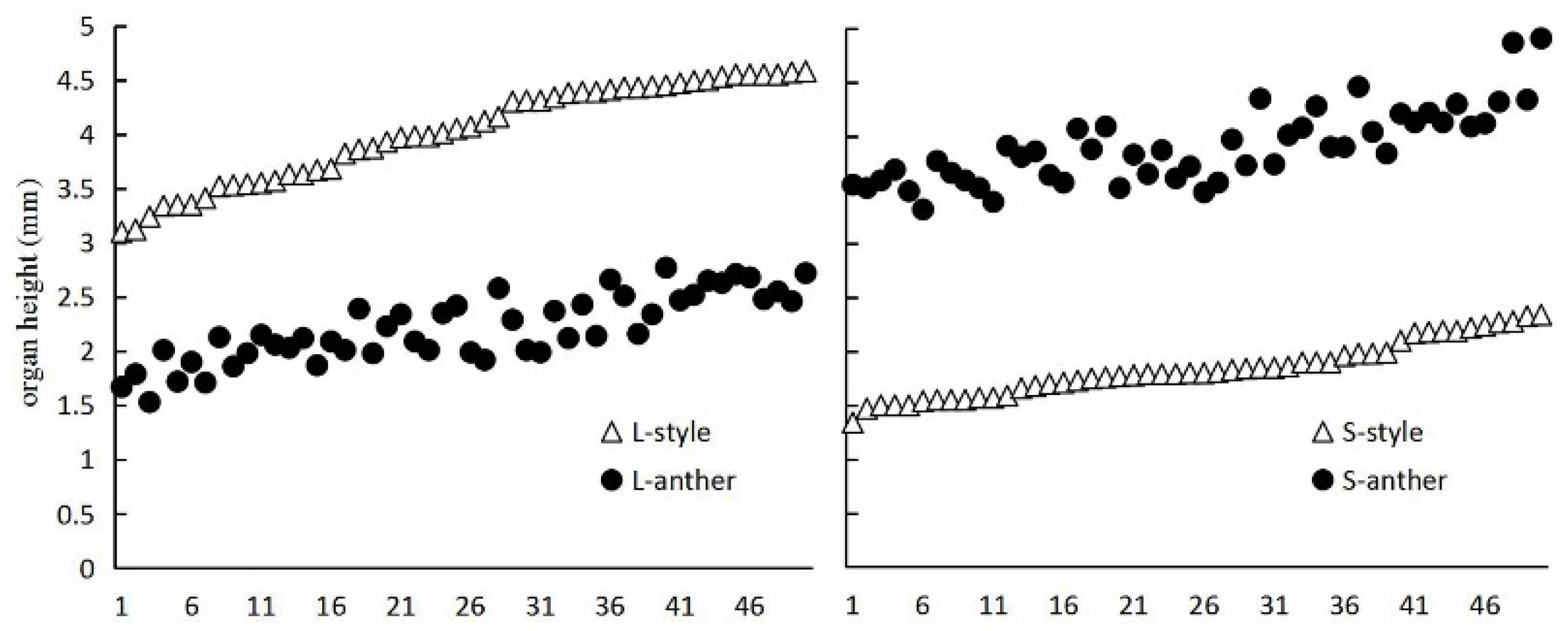
3.1. Floral Biology
3.2. Style-Morph Ratios
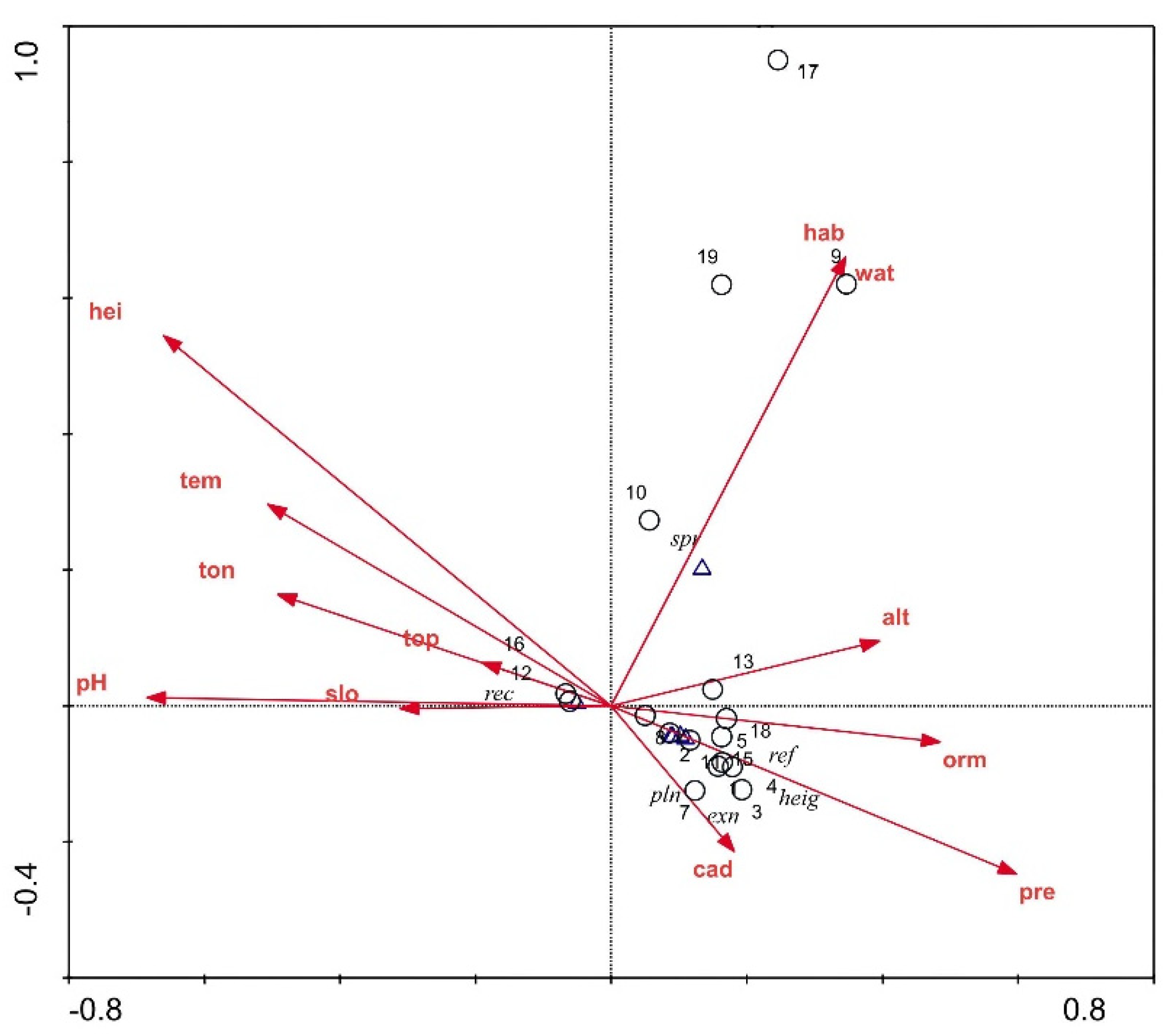
3.3. CCA Results
3.3.1. Characteristics of the Environmental Factors
3.3.2. Characteristics of Ecological Factors
3.3.3. CCA of P. criopolitanum Samples
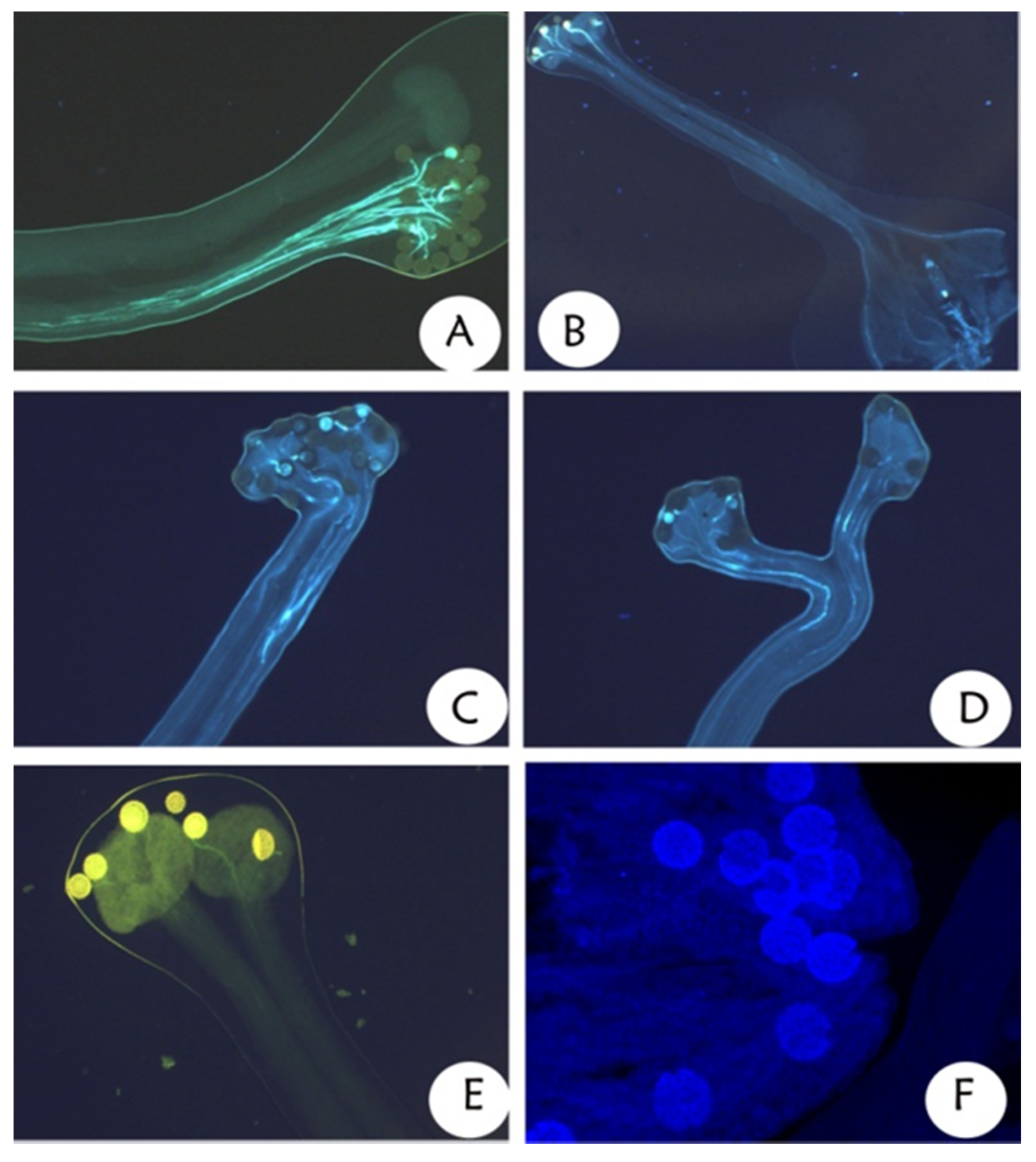
3.4. Mating System Relationships
3.5. Seed Sets
3.6. Seed Germination
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ganders, E. The biology of heterostyly. N. Z. J. Bot. 1979, 17, 607–635. [Google Scholar] [CrossRef]
- Lloyd, D.G.; Webb, C.J. The evolution of heterostyly. In Evolution and Function of Heterostyly; Barrett, S.C.H., Ed.; Springer: Berlin, Germany, 1992; pp. 151–178. [Google Scholar]
- Barrett, S.C.H.; Jesson, L.K.; Baker, A.M. The evolution and function of stylar polymorphisms in flowering plants. Ann. Bot. 2000, 85 (Suppl. A), 253–265. [Google Scholar] [CrossRef]
- Chen, M.L.; You, Y.L.; Zhang, X.P. Advances in the research of heterostyly. Acta Pratacult. Sin. 2013, 19, 226–239. [Google Scholar]
- Moon, H.K. Confirmation of distyly in Perovskia and floral dimorphism of P. abrotanoides (Salviinae: Lamiaceae). Flora 2021, 283, 151905. [Google Scholar] [CrossRef]
- Darwin, C. The Different Forms of Flowers on Plants of the Same Species; Appleton & Co.: New York, NY, USA, 1877. [Google Scholar]
- Graham, R.A. Polygonaceae. In Flora of Tropical East; Turril, W.B., Redhead, E.M., Eds.; Africa Crown Agents: London, UK, 1958; pp. 1–40. [Google Scholar]
- Reddy, N.P.; Bahadur, B.; Kumar, P.V. Heterostyly in Polygonum chinense. J. Genet. 1977, 63, 79–82. [Google Scholar] [CrossRef]
- Hong, S.P. The dimorphic heterostyly in Aconogonon campanulatum (Polygonaceae). Plant Syst. Evol. 1991, 176, 125–131. [Google Scholar] [CrossRef]
- Hong, S.P. Pollen dimorphism in heterostylous species of Oxygonum. Syst. Geogr. Plants 1999, 68, 245–252. [Google Scholar] [CrossRef]
- Li, A.J.; Bao, B.J. Polygonaceae. In Flora of China; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2003; Volume 5, pp. 277–350. [Google Scholar]
- Chen, M.L.; Zhang, X.P. Distyly in Polygonum jucundum Meisn. Plant Syst. Evol. 2010, 288, 139–148. [Google Scholar] [CrossRef]
- Chen, M.L. Floral morphology and breeding system in Polygonum hastato-sagittatum Mak. (Polygonaceae). Flora 2012, 207, 365–371. [Google Scholar] [CrossRef]
- Lv, F.J.; Cui, M.C.; Chen, M.L. Reproductive biology of Polygonum japonicum. Acta Pratacult. Sin. 2013, 22, 196–203. [Google Scholar]
- Wen, H.H.; Chen, M.L.; Zhang, Z.S. Reproductive biology in the distylous species of Polygonum orientale. Acta Pratacult. Sin. 2015, 24, 155–162. [Google Scholar]
- Wang, C.H.; Du, W.; Wang, X.F. Reproductive investment in a cleistogamous morph of Polygonum jucundum (Polygonaceae). PlantSyst. Evol. 2017, 303, 559–563. [Google Scholar] [CrossRef]
- Guo, Y.N.; Chen, S.F.; Chen, M.L.; Li, B. Persicariajucunda var. rotunda (Polygonaceae, Persicarieae), a distinct distylous taxa raised to specific rank. PhytoKeys 2019, 126, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.W. Reproductive Ecology of Polygonum criopolitanum; Jiangxi Agricultural University: Nanchang, China, 2019. [Google Scholar]
- Fang, Y.B.; Shao, J.W.; Wei, Y.; Zhang, X.P. Pilot study on the breeding system of seven species of Polygonum. J. Biol. 2009, 26, 38–40. [Google Scholar]
- Xu, S.Q. Studies on Polygonum resources and utilization in Cixi. Chin. Wild Plant Resour. 2013, 32, 32–36. [Google Scholar]
- Karise, R.; Mänd, M.; Ivask, M.; Koskor, E.; Bender, A. The effect of pollen amount and its caloric value in hybrid lucerne (Medicago xvaria) on its attractiveness to bumble bees (Bombus terrestris). Agron. Res. 2006, 4, 211–216. [Google Scholar]
- Erdtman, G. The acetolysis technique: A revised description. Sven. Bot. Tisdkr. 1960, 54, 561–564. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing, Vienna. 2019. Available online: https://www.R-Project.org (accessed on 15 October 2021).
- Bao, S.D. Soil Agrochemical Analysis, 3rd ed.; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Hermann, B.P.; Mal, T.K.; Williams, R.J.; Dollahon, N.R. Quantitative evaluation of stigma polymorphism in a tristylous weed, Lythrumsalicaria (Lythraceae). Am. J. Bot. 1999, 86, 1121–1129. [Google Scholar] [CrossRef]
- Endels, P.; Jacquemyn, H.; Brys, R.; Hermy, M.; De Blust, G. Temporal changes (1986–1999) in populations of primrose (Primula vulgaris Huds.) in an agricultural landscape and implications for conservation. Biol. Conserv. 2002, 105, 11–25. [Google Scholar] [CrossRef]
- Goodwillie, C. Inbreeding depression and mating systems in two species of Linanthus (Polemoniaceae). Heredity 2000, 84, 283–293. [Google Scholar] [CrossRef]
- Byers, D.L.; Meagher, T.R. Mate availability in small populations of plant species with homomorphic sporophitic self-incompatibility. Heredity 1992, 68, 353–359. [Google Scholar] [CrossRef]
- Kéry, M.; Matthies, D.; Spillmann, H.H. Reduced fecundity and offspring performance in small populations of the declining grassland plants Primula veris and Gentiana lutea. J. Ecol. 2000, 88, 17–30. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Brys, R.; Hermy, M. Patch occupancy, population size and reproductive success of a forest herb (Primula elatior) in a fragmented landscape. Oecologia 2002, 130, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Hartley, M.L.; Tshamekeng, E.; Thomas, S.M. Functional heterostyly in Tylosema esculentum (Caesalpinioideae). Ann. Bot. 2002, 89, 67–76. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, C.; Wang, L.L.; Lan, D.; Yang, Y.P.; Duan, Y.W. Pollination ecology of Arnebiaszechenyi(Boraginaceae), a Chinese endemic perennial characterized by distyly and heteromorphic self-incompatibility. Ann. Bot. Fenn. 2014, 51, 297–304. [Google Scholar] [CrossRef]
- Arnold, E.S.; Richards, A.J. On the occurrence of unilateral imcompatibility in Primula section Aleuritia Duby and the origin of Primula scotica Hook. Bot. J. Linn. Soc. 1998, 128, 359–368. [Google Scholar]
- Chen, M.L. Comparative reproductive biology of Primula merrilliana Schltr. and P. cicutariifolia Pax. Plant Syst. Evol. 2009, 278, 23–32. [Google Scholar] [CrossRef]
- Brys, R.; Jacquemyn, H.; Beeckman, T. Morph-ratio variation, population size and female reproductive success in distylous Pulmonaria officinalis (Boraginaceae). J. Evol. Biol. 2008, 21, 1281–1289. [Google Scholar] [CrossRef]
- Gao, S.P.; Li, W.J.; Hong, M.T.; Lei, T.; Shen, P.; Li, J.N.; Jiang, M.Y.; Duan, Y.F.; Shi, L.S. The nonreciprocal heterostyly and heterotypic self-incompatibility of Ceratostigma willmottianum. J. Plant Res. 2021, 134, 543–557. [Google Scholar] [CrossRef]
- Yuan, S.; Barrett, S.C.H.; Zhang, D.X. Genetics of distyly and homostyly in a self-compatible Primula. Heredity 2019, 122, 110–119. [Google Scholar] [CrossRef]
- Faria, R.R.; Ferrero, V.; Navarro, L.; Araujo, A.C. Flexible mating system in distylous populations of Psychotria carthagenensis Jacq. (Rubiaceae) in Brazilian Cerrado. Plant Syst. Evol. 2012, 298, 619–627. [Google Scholar] [CrossRef]
- Rodrigues, E.B.; Consolaro, H. Atypical distyly in Psychotria goyazensis Mull. Arg. (Rubiaceae), an intramorph self-compatible species. Acta Bot. Bras. 2013, 27, 155–161. [Google Scholar] [CrossRef]
- Baker, H.G. Self-compatibility and establishment after “long-distance” dispersal. Evolution 1955, 9, 347–348. [Google Scholar]
- Ågren, J. Population size, pollinator limitation, and seed set in the self-incompatible herb Lythrumsalicaria. Ecology 1996, 77, 1779–1790. [Google Scholar] [CrossRef]
- Smith, J.R.; Hough-Goldstein, J.; Lake, E.C. Variable Seed Viability of Mile-a-Minute Weed (Devil’s Tearthumb, Persicaria perfoliata). Invasive Plant Sci. Manag. 2014, 7, 107–112. [Google Scholar] [CrossRef]
- Momose, K.; Inoue, T. Pollination and factors limiting fruit set of chasmogamous flowers of an amphicarpic annual, Polygonum thunbergii (Polygonaceae). Res. Popul. Ecol. 1993, 35, 79–93. [Google Scholar] [CrossRef]
- Diggle, P.K.; Meixner, M.A.; Carroll, A.B.; Aschwanden, C.F. Barriers to sexual reproduction in Polygonum viviparum: A comparative developmental analysis of P. viviparum and P. bistortoides. Ann. Bot. 2002, 89, 145–156. [Google Scholar] [CrossRef]
- Barrett, S.C.H. The reproductive biology and genetics of island plants. Philos. Trans. R. Soc. Lond. B 1996, 351, 725–733. [Google Scholar]
- Sculthorpe, C.D. The Biology of Aquatic Vascular Plants; Edward Arnold: London, UK, 1967. [Google Scholar]
- Philbrick, C.T.; Les, D.H. Evolution of aquatic angiosperm reproductive system. BioScience 1996, 46, 815–826. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flower Characteristic | L-Morph | S-Morph | p-Value | Sample Number |
|---|---|---|---|---|
| Length of tepal(mm) | 2.52 ± 0.44 | 2.47 ± 0.29 | >0.05 | 50 |
| Flower diameter(mm) | 6.58 ± 0.47 | 6.42 ± 0.25 | >0.05 | 50 |
| Height of stigma(mm) | 4.02 ± 0.66 | 1.84 ± 0.26 | <0.001 | 50 |
| Height of anther(mm) | 2.19 ± 0.42 | 3.90 ± 0.42 | <0.001 | 50 |
| Stigma-anther separation(mm) | 2.06 ± 0.39 | 1.83 ± 0.55 | <0.05 | 100 |
| Pollen number | 647 ± 40 | 526 ± 38 | <0.01 | 100 |
| Pollen diameter(μm) | 51 ± 1.92 | 62 ± 2.51 | <0.001 | 100 |
| Geographic Position | Sample Sites | Sample Size (m2) | Flower No. of L-Morph | Flower No. of S-Morph | L-Morph:S-Morph | χ2 | p-Value | Deviate from 1:1 |
|---|---|---|---|---|---|---|---|---|
| Anqing City | ZhengfengTower | 1 | 211 | 0 | - | 211 | 2.2 × 10−16 | Y |
| Zongyang County | 1 | 185 | 0 | - | 185 | 2.2 × 10−16 | Y | |
| Guichi City | Huamiao | 1 | 12 | 0 | - | 12 | 5.32 × 10−4 | Y |
| Shibasuo | 1 | 0 | 97 | - | 97 | 2.2 × 10−16 | Y | |
| Tongling City | Shizishan 1 | 1 | 0 | 37 | - | 37 | 1.18 × 10−9 | Y |
| Shizishan 2 | 1 | 0 | 285 | - | 285 | 2.2 × 10−16 | Y | |
| Wuhu City | MafengRiver | 100 | 785 | 721 | 1.08 | 2.72 | 9.91 × 10−2 | N |
| MafengRiver 1 | 1 | 0 | 63 | - | 63 | 2.07 × 10−15 | Y | |
| MafengRiver 2 | 1 | 82 | 0 | - | 82 | 2.2 × 10−16 | Y | |
| MafengRiver 3 | 1 | 46 | 0 | - | 46 | 1.18 × 10−11 | Y | |
| QingyiRiver | 1 | 0 | 81 | - | 81 | 2.2 × 10−16 | Y | |
| Zhang River 1 | 100 | 102 | 0 | - | 102 | 2.2 × 10−16 | Y | |
| Zhang River 2 | 1 | 150 | 162 | 0.93 | 0.46 | 0.50 | Y | |
| ZhaojiaRiver | 1 | 72 | 0 | - | 72 | 2.2 × 10−16 | Y | |
| LongwoLake | 1 | 0 | 95 | - | 95 | 2.2 × 10−16 | Y | |
| Wanzhi | 1 | 0 | 89 | - | 89 | 2.2 × 10−16 | Y | |
| Manshan City | Yushanqu 1 | 1 | 19 | 0 | 1 | 19 | 1.31 × 10−5 | Y |
| Yushanqu 2 | 1 | 36 | 0 | 1 | 36 | 1.97 × 10−9 | Y | |
| Yushanqu 3 | 1 | 22 | 16 | 1.38 | 0.95 | 0.33 | N | |
| Yushanqu 4 | 1 | 21 | 0 | 1 | 21 | 4.59 × 10−6 | Y | |
| Yushanqu 5 | 1 | 18 | 92 | 0.20 | 49.78 | 1.72 × 10−12 | Y | |
| Tunxi City | Taiping Lake | 100 | 340 | 316 | 1.08 | 0.88 | 0.35 | N |
| Taiping Lake 1 | 100 | 54 | 0 | - | 54 | 2 × 10−13 | Y | |
| Taiping Lake 2 | 1 | 31 | 0 | - | 31 | 2.58 × 10−8 | Y | |
| Taiping Lake 3 | 1 | 0 | 49 | - | 49 | 2.56 × 10−12 | Y | |
| Taiping Lake 4 | 1 | 11 | 0 | - | 11 | 9.11 × 10−4 | Y | |
| Luan City | Luan Pi River | 100 | 345 | 67 | 5.15 | 187.58 | 2.2 × 10−16 | Y |
| Luan Pi River1 | 1 | 121 | 0 | - | 121 | 2.2 × 10−16 | Y | |
| Luan Pi River2 | 1 | 56 | 0 | - | 56 | 2.2 × 10−16 | Y | |
| Luan Pi River3 | 1 | 0 | 160 | - | 160 | 2.2 × 10−16 | Y | |
| Luan 1 | 1 | 42 | 24 | 1.75 | 4.91 | 0.03 | Y | |
| Luan 2 | 1 | 35 | 0 | 1 | 35 | 3.30 × 10−9 | Y | |
| Luan 3 | 1 | 33 | 18 | 1.83 | 4.41 | 0.04 | Y | |
| Nanchang City | Jinxian County | 100 | 895 | 842 | 1.06 | 1.62 | 0.20 | N |
| Jinxian County 1 | 100 | 112 | 0 | - | 112 | 2.2 × 10−16 | Y | |
| Jinxian County 2 | 1 | 56 | 12 | 4.67 | 28.47 | 9.51 × 10−9 | Y | |
| Jinxian County 3 | 1 | 0 | 90 | - | 90 | 2.2 × 10−16 | Y | |
| Huangjiacun 1 | 1 | 34 | 11 | 3.09 | 11.76 | 6.1 × 10−4 | Y | |
| Huangjiacun 2 | 1 | 0 | 34 | 1 | 34 | 2.2 × 10−16 | Y | |
| Huangjiacun 3 | 1 | 26 | 0 | 1 | 26 | 3.41e−7 | Y | |
| Huangjiacun 4 | 1 | 43 | 0 | 1 | 43 | 5.47 × 10−11 | Y | |
| Huangjiacun 5 | 1 | 27 | 30 | 0.90 | 0.16 | 0.69 | N | |
| Huangjiacun 6 | 1 | 0 | 30 | 1 | 30 | 4.32 × 10−8 | Y | |
| Huangjiacun 7 | 1 | 29 | 27 | 1.07 | 0.07 | 0.79 | N |
| Sample | Population No. | Altitude (m) | Height Gap to the Adjacent Water (m) | Soil Water Content | Habitat (1: Aquatic, 2: Hygrocolous, 3: Xeromorphic) | Annual Precipitation (mm) | Annual Average Temperature (℃) | Slope (℃) | Canopy Density | pH | Organic Matter (g/kg) | Total Nitrogen (g/kg) | Total Phosphorus (g/kg) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| alt | heig | wat | hab | pre | tem | slo | cad | pH | orm | ton | top | ||
| WushiTown | 1 | 18 | 0.5 | 0.39 | 2 | 1564.5 | 15.4 | 6 | 0.7 | 6.56 | 38.5 | 0.409 | 0.208 |
| Taiping Lake | 2 | 17 | 0.8 | 0.38 | 2 | 1564.5 | 15.4 | 12 | 0.56 | 6 | 50.4 | 0.254 | 0.45 |
| AnqingYingjiangTower | 3 | 21 | 1 | 0.32 | 2 | 1400 | 15.5 | 49 | 0.1 | 5.6 | 48.6 | 0.102 | 0.345 |
| Guichishibasuo | 4 | 17 | 1.2 | 0.25 | 2 | 1578.3 | 15.9 | 8 | 0.32 | 5.72 | 19.5 | 0.393 | 0.41 |
| Tonglingshizishan | 5 | 156 | 2.1 | 0.34 | 2 | 1589.1 | 15.8 | 32 | 0.38 | 7 | 22.7 | 0.327 | 0.208 |
| Yangzi River 1 | 6 | 6.5 | 0 | 1 | 1 | 1054.8 | 16.2 | 12 | 0 | 7.32 | 33.5 | 0.256 | 0.467 |
| Yangzi River 2 | 7 | 7.5 | 1 | 0.38 | 2 | 1054.8 | 16.2 | 25 | 0.75 | 6.33 | 21.9 | 0.278 | 0.285 |
| Yangzi River 3 | 8 | 9.5 | 3 | 0.32 | 2 | 1054.8 | 16.2 | 25 | 0.76 | 6.38 | 34.6 | 0.346 | 0.156 |
| Yangzi River 4 | 9 | 11.5 | 5 | 0.09 | 3 | 1054.8 | 16.2 | 25 | 0.56 | 6.36 | 34.9 | 0.165 | 0.234 |
| Zhang River 1 | 10 | 9 | 0 | 1 | 1 | 1054.8 | 16.2 | 6 | 0 | 7.39 | 44.6 | 0.287 | 0.35 |
| Zhang River 2 | 11 | 11 | 2 | 0.519 | 2 | 1054.8 | 16.2 | 23 | 0.63 | 7.17 | 29.3 | 0.158 | 0.4 |
| Zhang River 3 | 12 | 12 | 3 | 0.485 | 2 | 1054.8 | 16.2 | 25 | 0.42 | 7.14 | 28.6 | 0.402 | 0.321 |
| Zhang River 4 | 13 | 13 | 4 | 0.123 | 3 | 1054.8 | 16.2 | 26 | 0.35 | 7.52 | 44.7 | 0.483 | 0.235 |
| LongwoLake 1 | 14 | 10 | 0 | 1 | 1 | 1054.8 | 16.2 | 42 | 0 | 7.23 | 45.6 | 0.306 | 0.408 |
| LongwoLake 2 | 15 | 11 | 1 | 0.34 | 2 | 1054.8 | 16.2 | 12 | 0.23 | 7.26 | 48.9 | 0.415 | 0.296 |
| LongwoLake 3 | 16 | 13 | 3 | 0.26 | 2 | 1054.8 | 16.2 | 28 | 0.25 | 7.42 | 21.3 | 0.439 | 0.318 |
| LongwoLake 4 | 17 | 15 | 5 | 0.45 | 3 | 1054.8 | 16.2 | 12 | 0.27 | 7.05 | 20.5 | 0.308 | 0.375 |
| Pi River 1 | 18 | 56 | 1.5 | 0.29 | 2 | 1100 | 16.3 | 21 | 0.32 | 5.67 | 42.6 | 0.446 | 0.284 |
| Pi River 2 | 19 | 62 | 5 | 0.03 | 3 | 1100 | 16.3 | 19 | 0.29 | 5.62 | 38.6 | 0.479 | 0.368 |
| Samples | Sample Number | Plants Number | Relative Coverage | Height(cm) | Relative Frequency | Species Richness | Exist or Not |
|---|---|---|---|---|---|---|---|
| Wushi Town | 1 | 118 | 0.85 | 16 | 0.76 | 3 | 1 |
| Taiping Lake | 2 | 12 | 0.34 | 15 | 0.54 | 4 | 1 |
| AnqingYingjiangTower | 3 | 4 | 0.16 | 14 | 0.12 | 1 | 1 |
| GuichiShibasuo | 4 | 3 | 0.12 | 11 | 0.21 | 2 | 1 |
| TonglingShizishan | 5 | 4 | 0.15 | 13 | 0.18 | 5 | 1 |
| Yangzi River 1 | 6 | 0 | 0 | 0 | 0 | 0 | 0 |
| Yangzi River 2 | 7 | 28 | 0.8 | 15 | 0.76 | 2 | 1 |
| Yangzi River 3 | 8 | 39 | 0.82 | 14 | 0.82 | 5 | 1 |
| Yangzi River 4 | 9 | 0 | 0 | 0 | 0 | 8 | 0 |
| Zhang River 1 | 10 | 0 | 0 | 0 | 0 | 1 | 0 |
| Zhang River 2 | 11 | 105 | 0.82 | 17 | 0.76 | 4 | 1 |
| Zhang River 3 | 12 | 90 | 78 | 16 | 0.74 | 8 | 1 |
| Zhang River 4 | 13 | 0 | 0 | 0 | 0 | 1 | 0 |
| Longwo Lake 1 | 14 | 0 | 0 | 0 | 0 | 0 | 0 |
| Longwo Lake 2 | 15 | 7 | 0.13 | 15 | 0.26 | 5 | 1 |
| Longwo Lake 3 | 16 | 4 | 0.08 | 15 | 0.21 | 4 | 1 |
| Longwo Lake 4 | 17 | 0 | 0 | 0 | 0 | 4 | 0 |
| Pi River 1 | 18 | 29 | 0.76 | 16 | 0.35 | 6 | 1 |
| Pi River 2 | 19 | 0 | 0 | 0 | 0 | 12 | 0 |
| Population | Population | Fruit Sets | |
|---|---|---|---|
| L-Morph | S-Morph | ||
| Taipinghu 1 | 156 | 32 | 4.17 |
| Taipinghu2 | 58 | 0 | 0 |
| Luan Pi River | 145 | 0 | 0 |
| Guichi Shibasuo | 0 | 45 | 0 |
| Anqing Zhengfeng Tower | 32 | 0 | 0 |
| Tongling | 49 | 0 | 0 |
| Wuhu MafengRiver | 176 | 152 | 3.16 |
| Wuhu Qingyijiang | 65 | 0 | 0 |
| Wuhu Longwo Lake | 0 | 47 | 0 |
| Wuhu Zhang River 1 | 89 | 0 | 0 |
| Wuhu Zhang River 2 | 80 | 11 | 0 |
| Jinxian County 1 | 215 | 208 | 6.67 |
| Jinxian County 2 | 0 | 73 | 0 |
| Jinxian County 3 | 87 | 0 | 0 |
| Process Mode | Pin | Thrum | ||||
|---|---|---|---|---|---|---|
| Flowers | Seeds | Seed Sets (%) | Flowers | Seeds | Seed Sets (%) | |
| Emasculation, bagged | 100 | 0 | 0 | 100 | 0 | 0 |
| Selfing | 100 | 0 | 0 | 100 | 0 | 0 |
| Illegitimate pollination (intramorph) | 100 | 0 | 0 | 100 | 0 | 0 |
| Legitimate pollination | 100 | 3 | 3% | 100 | 5 | 5% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, M.-L.; Qi, M.-Y.; Bai, B.-B.; Han, X. Reproductive Ecology of Distylous Shoreside Polygonum criopolitanum Hance. Diversity 2022, 14, 222. https://doi.org/10.3390/d14030222
Chen M-L, Qi M-Y, Bai B-B, Han X. Reproductive Ecology of Distylous Shoreside Polygonum criopolitanum Hance. Diversity. 2022; 14(3):222. https://doi.org/10.3390/d14030222
Chicago/Turabian StyleChen, Ming-Lin, Meng-Ying Qi, Bei-Bei Bai, and Xue Han. 2022. "Reproductive Ecology of Distylous Shoreside Polygonum criopolitanum Hance" Diversity 14, no. 3: 222. https://doi.org/10.3390/d14030222
APA StyleChen, M.-L., Qi, M.-Y., Bai, B.-B., & Han, X. (2022). Reproductive Ecology of Distylous Shoreside Polygonum criopolitanum Hance. Diversity, 14(3), 222. https://doi.org/10.3390/d14030222