
Basaltic Outcrops as Centers of Diversity for Xerothermic Plants in the Sudetes Mountains (Central Europe)


Abstract
1. Introduction
2. Materials and Methods
2.1. Basaltic Outcrops
2.2. Plant Species
2.3. Statistical Analysis
3. Results
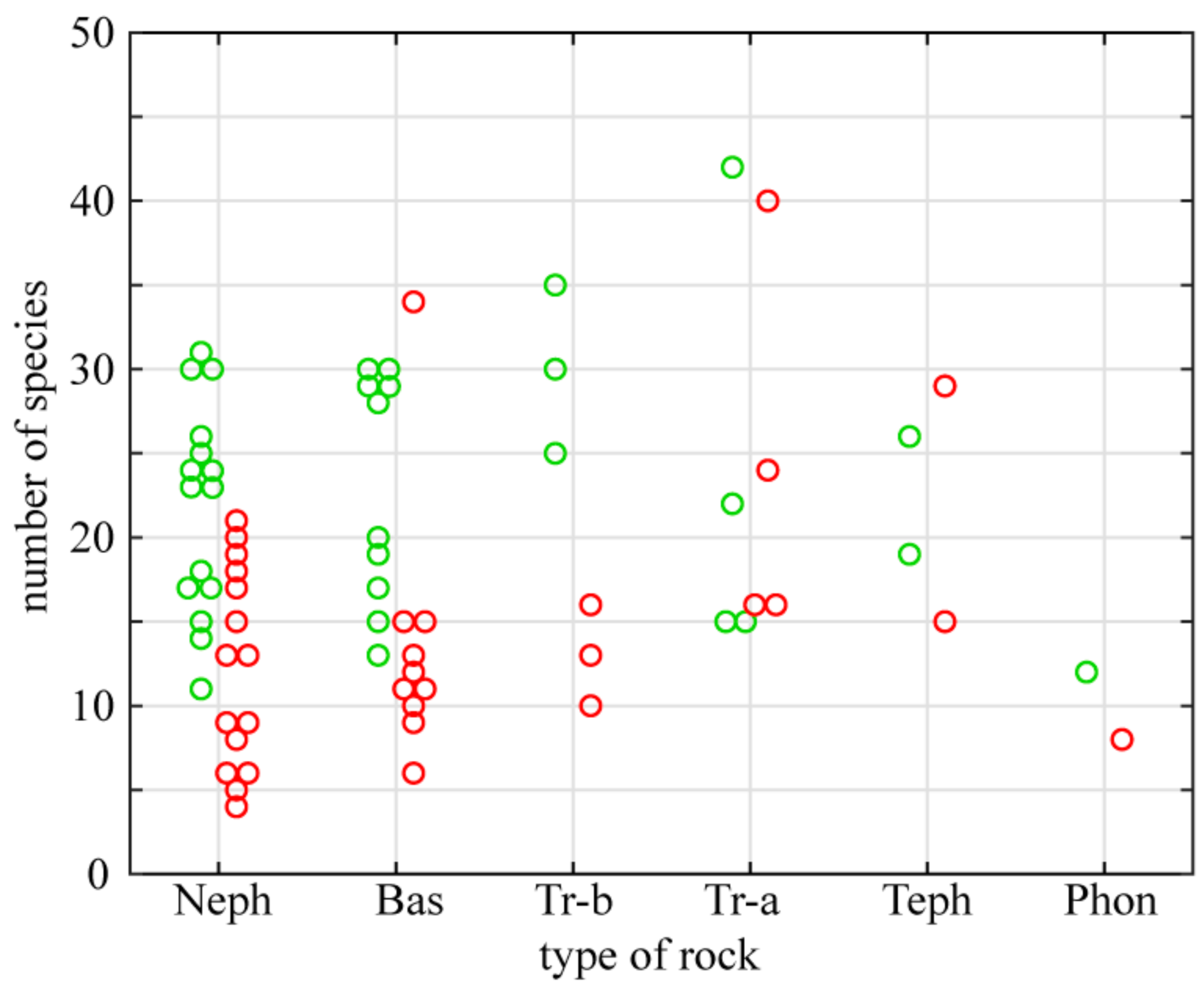
3.1. Phytogeographical Elements, Distribution of Xerothermic Species and Floristic Richness of Outcrops
3.2. Relationships between Xerothermic Species Richness and Outcrop Parameters
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Species | Family | Ellenberg Indicator Value | ||
|---|---|---|---|---|
| L | T | M | ||
| * Achillea pannonica Scheele | Asteraceae | 7 | 7 | 3 |
| * Acinos arvensis (Lam.) Dandy | Lamiaceae | 9 | 6 | 2 |
| Agrimonia eupatoria L. | Rosaceae | 7 | 6 | 4 |
| * Ajuga genevensis L. | Lamiaceae | 8 | 6 | 3 |
| * Allium lusitanicum Lam. | Alliaceae | 9 | 6 | 2 |
| * Allium oleraceum L. | Alliaceae | 7 | 6 | 3 |
| * Allyssum alyssoides (L.) L. | Brassicaceae | 9 | 6 | 3 |
| * Anthemis tinctoria L. | Asteraceae | 8 | 6 | 3 |
| * Anthericum ramosum L. | Anthericaceae | 7 | 5 | 3 |
| * Anthyllis vulneraria L. subsp. vulneraria | Fabaceae | 8 | 6 | 3 |
| * Arabis hirsuta (L.) Scop. | Brassicaceae | 7 | 5 | 4 |
| * Artemisia campestris L. subsp. campestris | Asteraceae | 9 | 6 | 2 |
| Astragalus glycyphyllos L. | Fabaceae | 6 | 6 | 4 |
| * Brachypodium pinnatum (L.) P. Beauv. | Poaceae | 6 | 5 | 4 |
| * Bromus erectus Huds. | Poaceae | 8 | 5 | 3 |
| Bupleurum falcatum L. | Apiaceae | 6 | 6 | 3 |
| * Camelina microcarpa DC. | Brassicaceae | 7 | 6 | 4 |
| Campanula cervicaria L. | Campanulaceae | 6 | 6 | 5 |
| * Campanula glomerata L. subsp. glomerata | Campanulaceae | 7 | 6 | 4 |
| Campanula persicifolia L. | Campanulaceae | 5 | 5 | 4 |
| * Carex caryophyllea Latourr. | Cyperaceae | 8 | 5 | 4 |
| Carex montana L. | Cyperaceae | 5 | 6 | 4 |
| * Carlina vulgaris L. | Asteraceae | 7 | 5 | 4 |
| * Centaurea scabiosa L. subsp. scabiosa | Asteraceae | 7 | 6 | 3 |
| * Centaurea stoebe L. | Asteraceae | 8 | 7 | 3 |
| Cephalanthera longifolia (L.) Fritsch | Orchidaceae | 5 | 5 | 4 |
| * Cerastium brachypetalum Pers. subsp. brachypetalum | Caryophyllaceae | 9 | 7 | 3 |
| * Cerastium glutinosum Fr. | Caryophyllaceae | 9 | 7 | 3 |
| * Cerastium pumilum Curtis | Caryophyllaceae | 8 | 7 | 2 |
| Cervaria rivini Gaertn. | Apiaceae | 8 | 7 | 5 |
| * Cirsium acaule Scop. | Asteraceae | 9 | 5 | 3 |
| Clinopodium vulgare L. | Lamiaceae | 7 | 5 | 4 |
| Cotoneaster integerrimus Medik. | Rosaceae | 8 | 6 | 3 |
| * Crepis praemorsa (L.) Walther | Asteraceae | 6 | 7 | 3 |
| Crataegus rhipidophylla Gand. s.str. | Rosaceae | 7 | 6 | 5 |
| Dactylorhiza sambucina (L.) Soó | Orchidaceae | 7 | 5 | 4 |
| * Dianthus carthusianorum L. | Caryophyllaceae | 8 | 5 | 3 |
| Digitalis grandiflora Mill. | Plantaginaceae | 7 | 4 | 5 |
| Drymocallis rupestris (L.) Soják | Rosaceae | 7 | 7 | 4 |
| Festuca brevipila (L.) R. Tracey | Poaceae | 8 | 6 | 3 |
| * Festuca pallens Host | Poaceae | 9 | 7 | 2 |
| * Filipendula vulgaris Moench | Rosaceae | 7 | 6 | 3 |
| Fragaria moschata Weston | Rosaceae | 6 | 6 | 5 |
| Fragaria viridis Weston | Rosaceae | 7 | 5 | 3 |
| Gagea villosa (M. Bieb) Sweet | Liliaceae | 6 | 7 | 4 |
| Galium verum L. s.str. | Rubiaceae | 7 | 6 | 4 |
| Genista germanica L. | Fabaceae | 7 | 5 | 4 |
| Genista tinctoria L. subsp. tinctoria | Fabaceae | 8 | 6 | 6 |
| * Gentianopsis ciliata (L.) Ma | Gentianaceae | 7 | 6 | 3 |
| Geranium sanguineum L. | Geraniaceae | 7 | 6 | 3 |
| * Helianthemum nummularium (L.) Mill. subsp. obscurum (Wahlenb.) Holub | Cistaceae | 8 | 5 | 3 |
| Hieracium diapahanoides Lindeb. | Asteraceae | 5 | 5 | 4 |
| Hieracium schmidtii Tausch | Asteraceae | 8 | 6 | 4 |
| * Holosteum umbellatum L. | Caryophyllaceae | 8 | 6 | 3 |
| Hypericum montanum L. | Hypericaceae | 5 | 6 | 4 |
| * Hypochaeris maculata L. | Asteraceae | 7 | 6 | 4 |
| Inula conyzae (Griess.) DC. | Asteraceae | 6 | 6 | 4 |
| * Inula hirta L. | Asteraceae | 7 | 6 | 3 |
| * Jovibarba globifera (L.) J. Parn. subsp. globifera | Crassulaceae | 9 | 6 | 2 |
| * Koeleria macrantha (Ledeb.) Schult. | Poaceae | 7 | 6 | 3 |
| Lathyrus niger (L.) Bernh. | Fabaceae | 5 | 6 | 3 |
| Lathyrus sylvestris L. subsp. sylvestris | Fabaceae | 7 | 6 | 4 |
| Lychnis viscaria L. | Caryophyllaceae | 7 | 6 | 3 |
| Medicago falcata L. | Fabaceae | 8 | 6 | 3 |
| * Medicago minima (L.) L. | Fabaceae | 9 | 7 | 3 |
| * Melica transsilvanica Schur | Poaceae | 7 | 8 | 3 |
| Melittis melissophyllum L. | Lamiaceae | 5 | 7 | 4 |
| Orchis mascula (L.) L. subsp. speciosa (W. D. J. Koch) Hegi | Orchidaceae | 6 | 6 | 4 |
| Origanum vulgare L. subsp. vulgare | Lamiaceae | 7 | 6 | 3 |
| * Ornithogalum angustifolium Boreau | Hyacinthaceae | 9 | 8 | 2 |
| * Petrorhagia prolifera (L.) P. W. Ball et Heywood | Caryophyllaceae | 8 | 7 | 3 |
| Peucedanum oreoselinum (L.) Moench | Apiaceae | 6 | 6 | 3 |
| * Phleum phleoides (L.) H. Karst. | Poaceae | 8 | 6 | 3 |
| * Pilosella bauhinii (Schult.) Arv.-Touv. subsp. bauhinii | Asteraceae | 9 | 7 | 3 |
| * Poa angustifolia L. | Poaceae | 7 | 6 | 5 |
| * Poa bulbosa L. | Poaceae | 8 | 7 | 3 |
| * Polygala comosa Schkuhr | Polygalaceae | 8 | 6 | 3 |
| Polygonatum odoratum (Mill.) Druce | Ruscaceae | 7 | 5 | 3 |
| Potentilla alba L. | Rosaceae | 6 | 6 | 4 |
| * Potentilla inclinata Vill. | Rosaceae | 9 | 7 | 2 |
| * Potentilla leucopolitana P. J. Müll | Rosaceae | 9 | 6 | 2 |
| * Potentilla neumanniana Rchb. | Rosaceae | 8 | 6 | 3 |
| Potentilla recta L. | Rosaceae | 9 | 7 | 3 |
| * Prunella grandiflora (L.) Scholler | Lamiaceae | 9 | 7 | 3 |
| Pulmonaria angustifolia L. | Boraginaceae | 5 | 7 | 5 |
| Ranunculus polyanthemos L. subsp. polyanthemos | Ranunculaceae | 6 | 6 | 4 |
| * Salvia pratensis L. | Lamiaceae | 8 | 6 | 3 |
| * Sanguisorba minor Scop. subsp. minor | Rosaceae | 7 | 6 | 3 |
| * Saxifraga tridactylites L. | Saxifragaceae | 8 | 6 | 2 |
| * Scabiosa columbaria L. | Dipsacaceae | 8 | 5 | 3 |
| * Scabiosa ochroleuca L. | Dipsacaceae | 8 | 7 | 3 |
| Securigera varia (L.) Lassen | Fabaceae | 7 | 6 | 4 |
| * Seseli annuum L. | Apiaceae | 8 | 7 | 3 |
| Silene nutans L. | Caryophyllaceae | 7 | 6 | 3 |
| Sorbus torminalis (L.) Crantz | Rosaceae | 4 | 7 | 4 |
| * Stachys germanica L. | Lamiaceae | 7 | 7 | 3 |
| Staphyllea pinnata L. | Staphyleaceae | 7 | 7 | 5 |
| Tanacetum corymbosum (L.) Sch. Bip. | Asteraceae | 6 | 7 | 4 |
| * Taraxacum sect. Erythrosperma (H. Lindb.) Dahlst. | Asteraceae | 8 | 6 | 3 |
| * Teucrium botrys L. | Lamiaceae | 9 | 6 | 2 |
| Thalictrum minus L. subsp. minus | Ranunculaceae | 6 | 6 | 3 |
| Trifolium alpestre L. | Fabaceae | 7 | 6 | 3 |
| Trifolium medium L. | Fabaceae | 7 | 6 | 4 |
| * Trifolium montanum L. | Fabaceae | 8 | 6 | 3 |
| Trifolium rubens L. | Fabaceae | 7 | 6 | 3 |
| * Trifolium striatum L. | Fabaceae | 8 | 7 | 3 |
| * Turritis glabra L. | Brassicaceae | 6 | 6 | 3 |
| Valeriana pratensis Dierb. subsp. angustifolia (Soó) Kirschner et al. | Valerianaceae | 7 | 6 | 4 |
| Verbascum lychnitis L. | Scrophulariaceae | 7 | 6 | 3 |
| * Veronica prostrata L. s.str. | Plantaginaceae | 9 | 7 | 2 |
| Veronica teucrium L. | Plantaginaceae | 7 | 6 | 3 |
| Vicia dumetorum L. | Fabaceae | 6 | 6 | 5 |
| Vicia pisiformis L. | Fabaceae | 6 | 7 | 4 |
| Vicia sylvatica L. | Fabaceae | 7 | 6 | 4 |
| Vicia tenuifolia Roth s.str. | Fabaceae | 7 | 8 | 4 |
| Vincetoxicum hirundinaria Medik. | Apocynaceae | 6 | 5 | 3 |
| * Viola collina Besser | Violaceae | 6 | 5 | 3 |
| Viola hirta L. | Violaceae | 6 | 5 | 3 |
| * Viola rupestris F. W. Schmidt | Violaceae | 6 | 5 | 3 |
| Viola scabra F. Braun | Violaceae | 6 | 5 | 3 |
References
- Honnay, O.; Jacquemyn, H.; Bossuyt, B.; Hermy, M. Forest fragmentation effects on patch occupancy and population variability of herbaceous plant species (Tansley review). New Phytol. 2005, 166, 723–736. [Google Scholar] [CrossRef] [PubMed]
- Lundholm, J.T. Plant species diversity and environmental heterogeneity: Spatial scale and competing hypotheses. J. Veg. Sci. 2009, 20, 377–391. [Google Scholar] [CrossRef]
- Tamme, R.; Hiiesalu, I.; Laanisto, L.; Szava-Kovats, R.; Pärtel, M. Environmental heterogeneity, species diversity and co-existence at different spatial scales. J. Veg. Sci. 2010, 21, 796–801. [Google Scholar] [CrossRef]
- Stein, A.; Gerstner, K.; Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 2014, 17, 866–880. [Google Scholar] [CrossRef]
- Fahrig, L. Ecological responses to habitat fragmentation per se. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 1–23. [Google Scholar] [CrossRef]
- Fahrig, L.; Arroyo-Rodriguez, V.; Bennett, J.R.; Boucher-Lalonde, V.; Cazetta, E.; Currie, D.J.; Eigenbrod, F.; Ford, A.T.; Harrison, S.P.; Jaeger, J.A.G.; et al. Is habitat fragmentation bad for biodiversity? Biol. Conserv. 2019, 230, 179–186. [Google Scholar] [CrossRef]
- Deák, B.; Kovács, B.; Rádai, Z.; Apostolova, I.; Kelemen, A.; Kiss, R.; Lukács, K.; Palpurina, S.; Sopotlieva, D.; Báthori, F.; et al. Linking environmental heterogeneity and plant diversity: The ecological role of small natural features in homogeneous landscapes. Sci. Total Environ. 2021, 763, 144199. [Google Scholar] [CrossRef]
- Szarzyński, J. Xeric Islands: Environmental Conditions on Inselbergs. In Inselbergs Biotic Diversity of Isolated Rock Outcrops in Tropical and Temperate Regions; Porembski, S., Barthlott, W., Eds.; Springer: Berlin/Heildelberg, Germany; New York, NY, USA, 2000; pp. 37–48. [Google Scholar]
- Oertli, B.; Joye, D.A.; Castella, E.; Juge, R.; Cambin, D.; Lachavanne, J.-B. Does size matter? The relationship between pond area and biodiversity. Biol. Conserv. 2002, 104, 59–70. [Google Scholar] [CrossRef]
- Rejmánková, R.; Rejmánek, M. Biogeography of artificial islands: Effects of age, area elevation, and isolation of plant species richness. Preslia 2002, 74, 307–314. [Google Scholar]
- Edvardsen, A.; Økland, R.H. Variation in plant species richness in and adjacent to 64 ponds in SE Norwegian agricultural landscapes. Aquat. Bot. 2006, 85, 79–91. [Google Scholar] [CrossRef]
- Vondrák, J.; Prach, K. Occurrence of heliophilous species on isolated rocky outcrops in a forested landscape: Relict species or recent arrivals ? Preslia 2006, 78, 115–121. [Google Scholar]
- Whittaker, R.J.; Fernández-Palacios, J.M. Island Biogeography. Ecology, Evolution, and Conservation; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Richardson, S.J.; Clayton, R.; Rance, B.D.; Broadbent, H.; McGlone, M.S.; Wilmshurst, J.M. Small wetlands are critical for safeguarding rare and threatened plant species. Appl. Veg. Sci. 2015, 18, 230–241. [Google Scholar] [CrossRef]
- Burns, K.C.; McHardy, R.P.; Pledger, S. The small-island effect: Fact or artefact? Ecography 2009, 32, 269–276. [Google Scholar] [CrossRef]
- Dengler, J. Robust methods for detecting a small island effect. Div. Distrib. 2010, 16, 256–266. [Google Scholar] [CrossRef]
- Triantis, K.A.; Sfenthourakis, S. Island biogeography is not a single-variable discipline: The small island effect debate. Div. Distrib. 2012, 18, 92–96. [Google Scholar] [CrossRef]
- Speziale, K.L.; Ezcurra, C. Rock outcrops as potential biodiversity refugia under climate change in North Patagonia. Plant Ecol. Divers. 2014, 8, 353–361. [Google Scholar] [CrossRef]
- Fitzsimons, J.A.; Michael, D.R. Rocky outcrops: A hard road in the conservation of critical habitats. Biol. Conserv. 2017, 211, 36–44. [Google Scholar] [CrossRef]
- Hunter, M.L., Jr.; Acuña, V.; Bauer, D.M.; Bell, K.P.; Calhoun, A.J.K.; Felipe-Lucia, M.R.; Fitzsimons, J.A.; González, E.; Kinnison, M.; Lindenmayer, D.; et al. Conserving small natural features with large ecological roles: A synthetic overview. Biol. Conserv. 2017, 211, 88–95. [Google Scholar] [CrossRef]
- Poschlod, P.; Braun-Reichert, R. Small natural features with large ecological roles in ancient agricultural landscapes of Central Europe—History, value, status, and conservation. Biol. Conserv. 2017, 211, 60–68. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, C.; Millien, V. A global synthesis of the small-island effect in habitat islands. Proc. R. Soc. B 2018, 285, 20181868. [Google Scholar] [CrossRef]
- Danin, A. Sandstone outcrops—A major refugium of Mediterranean flora in the xeric part of Jordan. Isr. J. Plant Sci. 1999, 47, 179–187. [Google Scholar] [CrossRef]
- Porembski, S.; Barthlott, W. Granitic and gneissic outcrop (inselbergs) as centers of diversity for desiccation-tolerant vascular plants. Plant Ecol. 2000, 151, 19–28. [Google Scholar] [CrossRef]
- Harrison, S.; Inouye, B.D. High β diversity in the flora of Californian serpentine ‘islands’. Biodiv. Conserv. 2002, 11, 1869–1876. [Google Scholar] [CrossRef]
- Burke, A. Inselbergs in a changing world—Global trends. Div. Distrib. 2003, 9, 375–383. [Google Scholar] [CrossRef]
- Larson, D.W.; Matthes, U.; Kelly, P.E. Cliff Ecology: Pattern and Process in Cliff Ecosystems; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Jacobi, C.M.; do Carmo, F.F.; Vincent, R.C.; Stehmann, J.R. Plant communities on ironstone outcrops: A diverse and endangered Brazilian ecosystem. Biodiv. Conserv. 2007, 16, 2185–2200. [Google Scholar] [CrossRef]
- Partzsch, M. Flora, Vegetation und historische Entwicklung der Porphyrkuppenlandschaft zwischen Halle und Wettin (Sachsen-Anhalt). Schlechtendalia 2007, 15, 1–91. [Google Scholar]
- Pope, N.; Harris, T.B.; Rajakaruna, N. Vascular Plants of Adjacent Serpentine and Granite Outcrops on the Deer Isles, Maine, U.S.A. Rhodora 2010, 112, 105–141. [Google Scholar] [CrossRef]
- Speziale, K.L.; Ezcurra, C. The role of outcrops in the diversity of Patagonian vegetation: Relicts of glacial palaeofloras? Flora Morphol. Distrib. Funct. Ecol. Plants 2012, 207, 141–149. [Google Scholar] [CrossRef]
- Djordjević, V.; Tsiftsis, S. Patterns of orchid species richness and composition in relation to geological substrates. Wulfenia 2019, 26, 1–21. [Google Scholar]
- Yates, C.J.; Robinson, T.; Wardell-Johnson, G.W.; Keppel, G.; Hopper, S.D.; Schut, A.G.T.; Byrne, M. High species diversity and turnover in granite inselberg floras highlight the need for a conservation strategy protecting many outcrops. Ecol. Evol. 2019, 9, 7660–7675. [Google Scholar] [CrossRef]
- Andrino, C.O.; Barbosa-Silva, R.G.; Lovo, J.; Viana, P.L.; Moro, M.F.; Zappi, D.C. Iron islands in the Amazon: Investigating plant beta diversity of canga outcrops. PhytoKeys 2020, 165, 1–25. [Google Scholar] [CrossRef]
- Taberlet, P.; Fumagalli, L.; Wust-Saucy, A.G.; Cossons, J.-F. Comparative phylogeography and post-glacial colonization routes in Europe. Molec. Ecol. 1998, 7, 453–464. [Google Scholar] [CrossRef]
- Kirschner, P.; Záveská, E.; Gamisch, A.; Hilpold, A.; Trucchi, E.; Paun, O.; Sanmartín, I.; Schlick-Steiner, B.C.; Frajman, B.; Arthofer, W.; et al. Long-term isolation of European steppe outposts boosts the biome’s conservation value. Nat. Commun. 2020, 11, 1968. [Google Scholar] [CrossRef]
- Pärtel, M. Local plant diversity patterns and evolutionary history at the regional scale. Ecology 2002, 83, 2361–2366. [Google Scholar] [CrossRef]
- Hejcman, M.; Hejcmanová, P.; Pavlů, V.; Beneš, J. Origin and history of grasslands in Central Europe—A review. Grass Forage Sci. 2013, 68, 345–363. [Google Scholar] [CrossRef]
- Leuschner, C.; Ellenberg, H. Ecology of Central European Non-Forest Vegetation: Coastal to Alpine, Natural to Man-Made Habitats. Vegetation Ecology of Central Europe Volume II; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Slavík, B. Přispěvek k fytogeografické charakteristice středočeského kraje. Stud. ČSAV 1980, 1, 45–107. [Google Scholar]
- Pott, R. Biototypen Schützenswerte Lebensräume Deutschlands und angrezender Regionen; Ulmer: Stuttgart, Germany, 1996. [Google Scholar]
- Hempel, W. Offenlandrelikte im Oberlausitzer Bergland und im angrezenden Nordböhmen. Ber. Naturforsch. Ges. Oberlausitz 2010, 18, 43–48. [Google Scholar]
- Ludwig, O. Das Pontische und Aquilonare Element in der Flora Schlesiens. Beibl. Bot. Jahrb. 1923, 130, 11–38. [Google Scholar]
- Schalow, E. Was lehrt die heutige Pflanzenverbreitung über die schlesische Urlandschaft? Mitt. Beuthen. Ges. Musver. 1931, 13/14, 250–259. [Google Scholar]
- Szeląg, Z. Rośliny naczyniowe Masywu Śnieżnika i Gór Bialskich; Instytut Botaniki im. W. Szafera, Polska Akademia Nauk: Kraków, Poland, 2000. [Google Scholar]
- Kwiatkowski, P. Current State, Separateness and Dynamics of Vascular Flora of the Góry Kaczawskie (Kaczawa Mountains) and Pogórze Kaczawskie (Kaczawa Plateau). II. Phytogeographical Analysis; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2007. [Google Scholar]
- Żołnierz, L. Zbiorowiska trawiaste występujące na Dolnośląskich serpentynitach—Wybrane aspekty ekologii. Zesz. Nauk. Uniw. Przyr. Wroc. Rozpr. 2007, 247, 1–231. [Google Scholar]
- Ewald, J. The calcareous riddle: Why are there so many calciphilous species in the Central European flora? Folia Geobot. 2003, 38, 357–366. [Google Scholar] [CrossRef]
- Lustrino, M.; Wilson, M. The circum-Mediterranean anorogenic Cenozoic igneous province. Earth Sci. Rev. 2007, 81, 1–65. [Google Scholar] [CrossRef]
- Ulrych, J.; Dostal, J.; Adamovič, J.; Jelínek, E.; Špaček, P.; Hegner, E.; Balogh, K. Recurrent Cenozoic volcanic activity in the Bohemian Massif (Czech Republic). Lithos 2011, 123, 133–144. [Google Scholar] [CrossRef]
- Janoška, M. Sopky a Sopečné Vrchy České Republiky; Academia: Praha, Czech Republic, 2013. [Google Scholar]
- Badura, J.; Pécskay, Z.; Koszowska, E.; Wolska, A.; Zuchiewicz, W.; Przybylski, B. New age and petrological constraints on Lower Silesian basaltoids, SW Poland. Acta Geodyn. Geomater. 2005, 2, 7–15. [Google Scholar]
- Tietz, O.; Büchner, J.; Suhr, P.; Abratis, M.; Goth, K. Die Geologie des Baruther Schafberges und der Dubrauker Horken—Aufbau und Entwicklung eines känozoischen Vulkankomplexes in Ostsachsen. Ber. Naturforsch. Ges. Oberlausitz Suppl. 2011, 18, 15–48. [Google Scholar]
- Buchner, J.; Tietz, O.; Viereck, L.; Suhr, P.; Abratis, M. Volcanology, geochemistry and age of the Lausitz Volcanic Field. J. Earth Sci. 2015, 104, 2057–2083. [Google Scholar]
- Anioł-Kwiatkowska, J.; Świerkosz, K. Flora i roślinność rezerwatu “Ostrzyca Proboszczowicka” oraz jego otoczenia. Acta Univ. Wrat. Prace Bot. 1992, 48, 45–115. [Google Scholar]
- Kwiatkowski, P. Zbiorowiska roślinne projektowanego rezerwatu “Rataj” koło Jawora. Parki Nar. Rez. Przyr. 1995, 14, 95–108. [Google Scholar]
- Kwiatkowski, P. Szata roślinna Bazaltowej Góry i jej otoczenia. Acta Univ. Wrat. Prace Bot. 1996, 70, 73–110. [Google Scholar]
- Kwiatkowski, P. Zmiany we florze roślin naczyniowych Masywu Grodźca (Pogórze Kaczawskie). Przyr. Sudet. 2013, 16, 45–66. [Google Scholar]
- Szczęśniak, E. Szata roślinna projektowanego rezerwatu “Krzyżowa Góra” koło Strzegomia (Dolny Śląsk). Ochr. Przyr. 1998, 55, 61–75. [Google Scholar]
- Ritz, C.; Wünsche, A. Basaltkuppen in der östlichen Oberlausitz. Tuexenia 2017, 10, 49–65. [Google Scholar]
- Jenik, J. Large-scale pattern of biodiversity in Hercynian massifs. In Spatial Processes in Plant Communities; Krahulec, F., Agnew, A.D.Q., Agnew, S., Willems, J.H., Eds.; SPB Academic Publishing and Academia: Praha, Czech Republic, 1990; pp. 251–259. [Google Scholar]
- Mazurski, K.R. Environmental problems in the Sudetes, Poland. GeoJournal 1999, 46, 271–277. [Google Scholar] [CrossRef]
- Sawicki, L. Mapa Geologiczna Regionu Dolnośląskiego z Przyległymi Obszarami Czech i Niemiec 1: 100,000. Podstawy Litostratygraficzne i Kodyfikacja Wydzieleń; Państwowy Instytut Geologiczny: Warszawa, Poland, 1997. [Google Scholar]
- Birkenmajer, K.; Jerzmański, J.; Nairn, A.E.M. Palaeomagnetic Studies of Polish Rocks. IV. Cenozoic Basalts of Lower Silesia. Ann. Soc. Geol. Pol. 1970, 40, 31–61. [Google Scholar]
- Birkenmajer, K.; Pécskay, Z.; Grabowski, J.; Lorenc, M.W.; Zagożdżon, P.P. Radiometric Dating of the Tertiary Volcanics in Lower Silesia, Poland. III. K-Ar and Palaeomagnetic Data from Early Miocene Basaltic Rocks near Jawor, Fore-Sudetic Block. Ann. Soc. Geol. Pol. 2002, 72, 241–253. [Google Scholar]
- Birkenmajer, K.; Pécskay, Z.; Grabowski, J.; Lorenc, M.W.; Zagożdżon, P.P. Radiometric Dating of the Tertiary Volcanics in Lower Silesia, Poland. IV. Further K-Ar and Paleomagnetic Data from Late Oligocene to Early Miocene Basaltic Rocks of the Fore-Sudetic Block. Ann. Soc. Geol. Pol. 2004, 74, 1–19. [Google Scholar]
- Birkenmajer, K.; Pécskay, Z.; Grabowski, J.; Lorenc, M.W.; Zagożdżon, P.P. Radiometric Dating of the Tertiary Volcanics in Lower Silesia, Poland. V. K-Ar and Paleomagnetic Data from Late Oligocene to Early Miocene Basaltic Rocks of the North-Sudetic Depression. Ann. Soc. Geol. Pol. 2007, 77, 1–16. [Google Scholar]
- Birkenmajer, K.; Pécskay, Z.; Grabowski, J.; Lorenc, M.W.; Zagożdżon, P.P. Radiometric Dating of the Tertiary Volcanics in Lower Silesia, Poland. VI. K-Ar and Paleomagnetic Data from Basaltic Rocks of the West Sudety Mountains and their Northern Foreland. Ann. Soc. Geol. Pol. 2011, 81, 115–131. [Google Scholar]
- Le Maitre, R.W.; Streckeisen, A.; Zanettin, B.; Le Bas, M.J.; Bonin, B.; Bateman, P.; Bellieni, G.; Dudek, A.; Efremova, S.; Keller, J. Igneous Rocks: A Classification and Glossary of Terms Recommendations of the International Union of Geological Sciences Subcommission on the Systematics of Igneous Rocks; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Jäger, E.J. (Ed.) Rothmaler-Exkursionsflora von Deutschland. Gefässpflanzen: Grundband, 21. durchgesehene Auflage; Spinger Spektrum: Berlin, Germany, 2017. [Google Scholar]
- Mucina, L. Conspectus of Classes of European Vegetation. Folia Geobot. Phytotax. 1997, 32, 117–172. [Google Scholar] [CrossRef]
- Matuszkiewicz, W. Przewodnik do Oznaczania Zbiorowisk Roślinnych Polski; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2001. [Google Scholar]
- Dengler, J.; Eisenberg, M.; Schröder, J. Die grundwasserfernen Saumgesellschaften Nordostniedersachsens im europäischen Kontext—Teil I: Säume magerer Standorte (Trifolio-Geranietea sanguinei). Tuexenia 2006, 26, 51–91. [Google Scholar]
- Roleček, J. Teplomilné doubravy (Quercetea pubescentis). In Vegetace České Republiky. 4. Lesní a Křovinná Vegetace; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2013; pp. 296–346. [Google Scholar]
- Meusel, H.; Jäger, E.; Weinert, E. Vergleichende Chorologie der Zentraleuropäischen Flora. 1; Veb Gustav Fischer Verlag: Jena, Germany, 1965. [Google Scholar]
- Meusel, H.; Jäger, E.; Rauschert, S.; Weinert, E. Vergleichende Chorologie der Zentraleuropäischen Flora. 2; Veb Gustav Fischer Verlag: Jena, Germany, 1978. [Google Scholar]
- Meusel, H.; Jäger, E. Vergleichende Chorologie der Zentraleuropäischen Flora. 3; Veb Gustav Fischer Verlag: Jena, Germany, 1992. [Google Scholar]
- Finnie, T.J.R.; Preston, C.D.; Hill, M.O.; Uotila, P.; Crawley, M.J. Floristic elements in European plants: An analysis based on Atlas Florae Europaeae. J. Biogeogr. 2007, 34, 1848–1872. [Google Scholar] [CrossRef]
- Ellenberg, H.; Weber, H.E.; Düll, R.; Wirth, V.; Werner, W.; Paulissen, D. Zeigerwerte von Pflanzen in Mitteleuropa. 3. Durchgesehene Auflage. Scr. Geobot. 2001, 18, 1–262. [Google Scholar]
- Kwiatkowski, P. Chorological and phytogeographical diversity of tree and shrubs as a mean to regionalization: Kaczawa Mountains, Sudetes, Poland. Willdenowia 2014, 44, 363–376. [Google Scholar] [CrossRef]
- Wagensommer, R.P.; Perrino, E.V.; Silletti, G.N. Carex phyllostachys A. Mey. (Cyperaceae) new for Italy and phytogeographical considerations. Phyton 2014, 54, 215–222. [Google Scholar]
- Meindl, C.; Brune, V.; Listl, D.; Poschlod, P.; Reisch, C. Survival and postglacial immigration of the steppe plant Scorzonera purpurea to Central Europe. Plant Syst. Evol. 2016, 302, 971–984. [Google Scholar] [CrossRef]
- Kwiatkowski, P. Podgórska ciepłolubna dąbrowa brekiniowa Sorbo torminalis-Quercetum na Pogórzu Złotoryjskim. Fragm. Flor. Geobot. Polon. 2003, 10, 175–193. [Google Scholar]
- Krauss, J.; Klein, A.-M.; Steffan-Dewenter, I.; Tscharntke, T. Effects of habitat area, isolation, and landscape diversity on plant species richness of calcareous grasslands. Biodiv. Conserv. 2004, 13, 1427–1439. [Google Scholar] [CrossRef]
- Helm, A.; Hanski, I.; Partel, M. Slow response of plant species richness to habitat loss and fragmentation. Ecol. Lett. 2006, 9, 72–77. [Google Scholar] [CrossRef]
- Fletcher, R.J., Jr.; Didham, R.K.; Banks-Leite, C.; Barlow, J.; Ewers, R.M.; Rosindell, J.; Holt, R.D.; Gonzalez, A.; Pardini, R.; Damschen, E.I.; et al. Is habitat fragmentation good for biodiversity? Biol. Conserv. 2018, 226, 9–15. [Google Scholar] [CrossRef]
- Tsiftsis, S. The complex effect of heterogeneity and isolation in determining alpha and beta orchid diversity on islands in the Aegean archipelago. Syst. Biodivers. 2020, 18, 281–294. [Google Scholar] [CrossRef]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography. With a New Preface by Edward O. Wilson; Princeton University Press: Princeton, NJ, USA, 2001. [Google Scholar]
- Watling, J.I.; Arroyo-Rodríguez, V.; Pfeifer, M.; Baetenm, L.; Banks-Leite, C.; Cisneros, L.M.; Fang, R.; Hamel-Leigue, A.C.; Lachat, T.; Leal, I.R.; et al. Support for the habitat amount hypothesis from a global synthesis of species density studies. Ecol. Lett. 2020, 23, 674–681. [Google Scholar] [CrossRef]
- Triantis, K.A.; Mylonas, M.; Lika, K.; Vardinoyannis, K. A model for the species–area–habitat relationship. J. Biogeogr. 2003, 30, 19–27. [Google Scholar] [CrossRef]
- Cramer, M.J.; Willig, M.R. Habitat heterogeneity, species diversity and null models. Oikos 2005, 108, 209–218. [Google Scholar] [CrossRef]
- MacDonald, Z.G.; Anderson, I.D.; Acorn, J.H.; Nielsen, S.E. The theory of island biogeography, the sample-area effect, and the habitat diversity hypothesis: Complementarity in a naturally fragmented landscape of lake islands. J. Biogeogr. 2018, 45, 2730–2743. [Google Scholar] [CrossRef]
- Quinn, J.F.; Harrison, S.P. Effects of habitat fragmentation and isolation on species richness: Evidence from biogeographic patterns. Oecologia 1988, 75, 132–140. [Google Scholar] [CrossRef]
- Schrader, J.; Moeljono, S.; Keppel, G.; Krett, H. Plants on small islands revisited: The effects of spatial scale and habitat quality on the species-area relationship. Ecography 2019, 42, 1405–1414. [Google Scholar] [CrossRef]
- Huston, M. A general hypothesis of species diversity. Am. Nat. 1979, 113, 81–101. [Google Scholar] [CrossRef]
- Murphy, G.E.P.; Romanuk, T.N. A meta-analysis of declines in local species richness from human disturbances. Ecol. Evol. 2014, 4, 91–103. [Google Scholar] [CrossRef]
- Fabiszewski, J.; Kwiatkowski, P. Threatened vascular plants of the Sudeten Mountains. Acta Soc. Bot. Pol. 2002, 71, 339–350. [Google Scholar] [CrossRef][Green Version]
- Bräutigam, S.; Otto, H.-W. Rote Liste der Farn- und Samenpflanzen der Oberlauitz—Aktualisierte Fassung. Ber. Naturforsch. Ges. Oberlausitz 2012, 20, 99–116. [Google Scholar]
- Davis, B.N.K. Ecology of Quarries: The Importance of Natural Vegetation; Institute of Terrestrial Ecology: Cambridge, UK, 1982. [Google Scholar]
- Kwiatkowski, P. Kamieniołomy wapienia w Górach Kaczawskich ostoją rzadkich i ginących gatunków flory naczyniowej Sudetów. Gór. Odkryw. 1998, 40, 156–163. [Google Scholar]
- Mückschel, C. Floristische Beobachtungen in aufgelassenen Steinbrücken des Rheinischen Westerwaldes. Decheniana 1999, 153, 59–67. [Google Scholar]
- Novák, J.; Prach, K. Artificial sowing of endangered dry grassland species into disused basalt quarries. Flora Morphol. Distrib. Funct. Ecol. Plants 2010, 205, 179–183. [Google Scholar] [CrossRef]
- Tropek, R.; Kadlec, T.; Karesova, P.; Spitzer, L.; Kocarek, P.; Malenovsky, I.; Banar, P.; Tuf, I.H.; Hejda, M.; Konvicka, M. Spontaneous succession in limestone quarries as an effective restoration tool for endangered arthropods and plants. J. Appl. Ecol. 2010, 47, 139–147. [Google Scholar] [CrossRef]
- Raška, P.; Riezner, J.; Pokorný, R.; Holec, M.; Raška, M. Relations between Biotic and Abiotic Diversity in Abandoned Basalt Quarry and Its Relevance for Ecologic Restoration (Radobyl Hill, Northern Czechia). Acta Univ. Agric. Silvc. Mendel. Brun. 2017, 65, 151–166. [Google Scholar] [CrossRef]
- Soons, M.B.; Heil, G.W. Reduced colonization capacity in fragmented populations of wind-dispersed grassland forbs. J. Ecol. 2002, 90, 1033–1043. [Google Scholar] [CrossRef]
- Murphy, H.T.; Lovett-Doust, J. Context and connectivity in plant metapopulations and landscape mosaics: Does the matrix matter? Oikos 2004, 105, 3–14. [Google Scholar] [CrossRef]
- Hannus, J.-J.; von Numers, M. Vascular plant species richness in relation to habitat diversity and island area in the Finnish Archipelago. J. Biogeogr. 2007, 35, 1077–1086. [Google Scholar] [CrossRef]
- Lombini, A.; Ferrari, C.; Carperiè, B. The ecology of ophiolitic scree vegetation: A survey on the northern Apennine outcrops (Italy). Bocconea 2001, 13, 561–571. [Google Scholar]
- Kruckerberg, A. Geology and Plant Life. The Effects of Landforms and Rock Types on Plants; Washington University Press: Seattle, WA, USA, 2004. [Google Scholar]
- Korneck, D. Xerothermvegetation von Rheinland-Pfalz und Nachbargebieten. Schriftenr. Vegetationskd. 1974, 7, 1–196. [Google Scholar]
- Royer, J.M. Synthèse eurosibérienne, phytosociologique et phytogéographique de la classe des Festuco-Brometea. Diss. Bot. 1991, 178, 1–296. [Google Scholar]
- Di Pietro, R.; Wagensommer, R.P. A new Sesleria juncifolia association from south-eastern Italy and its position in the amphi-Adriatic biogeographical context. Acta Bot. Croat. 2014, 73, 171–207. [Google Scholar]
- Michalik, S. Charakterystyka ekologiczna kserotermicznej i górskiej flory naczyniowej Ojcowskiego Parku narodowego. Stud. Nat. 1979, 19, 1–95. [Google Scholar]
- Slavíková, J.; Molíková, M.; Rejmánek, M.; Rydlo, J.; Studnič, M.; Studničková, I.; Suchara, I.; Štolcová-Březinová, J. Ecological and Vegetational Differentiation of a Solitary Conic Hill (Oblik in České Středohoři Mts.); Academia: Praha, Czech Republic, 1983. [Google Scholar]
- McCune, B.; Keon, D. Equations for potential annual direct incident radiation and heat load. J. Veg. Sci. 2002, 13, 603–606. [Google Scholar] [CrossRef]
- Cook, M.W.; Lane, K.T.; Foster, B.L.; Holt, R.D. Island theory, matrix effects and species richness patterns in habitat fragments. Ecol. Lett. 2002, 5, 619–623. [Google Scholar] [CrossRef]
- Löbel, S.; Dengler, J.; Hobohm, C. Species richness of vascular plants, bryophytes and lichens in dry grasslands: The effects of environment, landscape structure and competition. Folia Geobot. 2006, 41, 377–393. [Google Scholar] [CrossRef]
- Cox, C.B.; Moore, P.D.; Ladle, R.J. Biogeography. An Ecological Approach, 9th ed.; Wiley-Blackwell: Oxford, UK, 2016. [Google Scholar]
- Diekmann, M. Species indicator values as important tool in applied plant ecology—A review. Bas. Appl. Ecol. 2003, 4, 493–506. [Google Scholar] [CrossRef]
- Berg, C.; Welk, E.; Jäger, E.J. Revising Ellenberg’s indicator values for continentality based on global vascular plants species distribution. Appl. Veg. Sci. 2017, 20, 482–493. [Google Scholar] [CrossRef]
- Chytrý, M.; Tichý, L.; Dřevojan, P.; Sádlo, J.; Zelený, D. Ellenberg-type indicator values for the Czech Flora. Preslia 2018, 90, 83–103. [Google Scholar] [CrossRef]
- Hedwall, P.-O.; Brunet, J.; Diekmann, M. With Ellenberg indicator values towards the north: Does the indicate power-decrease with distance from Central Europe? J. Biogeogr. 2019, 46, 1041–1053. [Google Scholar] [CrossRef]
- Tyler, T.; Herbertsson, L.; Olofsson, J.; Olsson, P.A. Ecological indicator and traits values for Swedish vascular plants. Ecol. Ind. 2021, 120, 106923. [Google Scholar] [CrossRef]
- Heywood, V.H.; Dulloo, M.E. In Situ Conservation of Wild Plant Species: A Critical Global Review of Best Practices; International Plant Genetic Resources Institute (IPGRI): Technical Bulletin No 11; Bioversity International: Rome, Italy, 2005. [Google Scholar]
- Heywood, V.H. In situ conservation of plant species—An unattainable goal? Israel Plant Sci. 2015, 63, 211–231. [Google Scholar] [CrossRef]
- Perrino, E.V.; Wagensommer, R.P. Crop Wild Relatives (CWR) Priority in Italy: Distribution, Ecology, In Situ and Ex Situ Conservation and Expected Actions. Sustainability 2021, 13, 1682. [Google Scholar] [CrossRef]








| Number of Locality | Locality | Summit Coordinates | Elevation (m) | Area (ha) | Rocks | Quarry | No. of Strictly Xerothermic Taxa | No. of Thermophilous Taxa |
|---|---|---|---|---|---|---|---|---|
| 1 | Rottstein a | 51°6′19.619” 14°45′53.518” | 455 | 100 | Nephelenite | 15 | 30 | |
| 2 | Knorrberg a | 51°0′50.784” 14°52′59.693” | 378 | 10 | Tephrite | × | 15 | 19 |
| 3 | Hutberg a | 51°3′19.284” 14°53′12.452” | 310 | 9 | Nephelenite | × | 18 | 26 |
| 4 | Kleiner Hutberg a | 51°3′4.231” 14°53′15.220” | 298 | 7 | Nephelenite | 17 | 23 | |
| 5 | Landeskrone a | 51°7′46.156” 14°55′58.394” | 419 | 60 | Nephelenite | 20 | 24 | |
| 6 | Koło Obserwatora b | 50°53′44.419” 14°59′24.850” | 341 | 4 | Phonolite | × | 8 | 12 |
| 7 | Kodešův vrch b | 50°53′52.703” 15°0′12.634” | 342 | 9 | Nephelenite | × | 9 | 14 |
| 8 | Borowa b | 51°2′46.166” 14°59′26.163” | 280 | 5 | Nephelenite | 9 | 18 | |
| 9 | Czubatka b | 51°4′24.142” 15°10′20.605” | 353 | 4 | Basanite | × | 6 | 13 |
| 10 | Stożek Perkuna b | 51°1′3.395” 15°14′1.726” | 385 | 2 | Nephelenite | × | 5 | 17 |
| 11 | Krzyżowa Góra b | 51°6′23.644” 15°19′12.764” | 250 | 4 | Nephelenite | × | 4 | 15 |
| 12 | Zamkowa c | 50°57′59.604” 15°38′39.203” | 311 | 13 | Basanite | × | 9 | 29 |
| 13 | Grodziec c | 51°10′37.178” 15°45′34.052” | 389 | 40 | Nephelenite | 19 | 31 | |
| 14 | Świątek c | 51°6′6.037” 15°44′7.583” | 330 | 2 | Nephelenite | × | 6 | 11 |
| 15 | Ostrzyca c | 51°3′20.838” 15°45′46.460” | 501 | 23 | Basanite | 10 | 28 | |
| 16 | Kamienna Góra c | 51°6′16.514” 15°49′49.944” | 350 | 32 | Nephelenite | × | 8 | 23 |
| 17 | Gruchacz c | 51°8′4.352” 15°53′36.743” | 260 | 5 | Nephelenite | × | 13 | 17 |
| 18 | Jeziorna c | 51°5′39.453” 15°51′55.703” | 293 | 11 | Basanite | × | 15 | 20 |
| 19 | Czerwony Kamień d | 51°5′27.615” 15°52′44.524” | 325 | 15 | Basanite | × | 15 | 29 |
| 20 | Wilcza Góra d | 51°6′18.260” 15°54′44.721” | 367 | 24 | Basanite | × | 34 | 30 |
| 21 | Kostrza d | 51°6′43.674” 15°56′1.351” | 313 | 2 | Basanite | 11 | 17 | |
| 22 | Kozia Góra d | 51°5′39.259” 15°56′12.938” | 373 | 17 | Basanite | × | 12 | 20 |
| 23 | Łysanka d | 51°3′40.113” 15°56′5.445” | 444 | 15 | Nephelenite | × | 13 | 24 |
| 24 | Krzyżowa Góra d | 51°5′28.003” 16°2′35.622” | 258 | 3 | Tephrite | × | 29 | 26 |
| 25 | Winnik d | 51°4′48.408” 16°5′23.713” | 251 | 2 | Trachyandesite | × | 24 | 22 |
| 26 | Srebrnik d | 51°5′41.301” 16°6′50.722” | 205 | 1 | Trachyandesite | 16 | 15 | |
| 27 | Kopista d | 51°5′11.895” 16°6′24.275” | 264 | 30 | Trachyandesite | × | 40 | 42 |
| 28 | Czartki d | 51°3′44.191” 16°4′50.883” | 273 | 1 | Trachyandesite | 16 | 15 | |
| 29 | Górzec d | 51°3′23.438” 16°4′30.961” | 442 | 29 | Nephelenite | 6 | 30 | |
| 30 | Czartowska Skała d | 51°2′16.150” 16°1′43.094” | 468 | 6 | Nephelenite | × | 21 | 25 |
| 31 | Rataj d | 51°1′17.339” 16°7′30.777” | 350 | 10 | Trachybasalt | × | 13 | 30 |
| 32 | Bazaltowa Góra d | 51°0′48.041” 16°7′59.992” | 367 | 80 | Trachybasalt | × | 16 | 35 |
| 33 | Radogost d | 50°59′50.690” 16°7′45.402” | 398 | 14 | Trachybasalt | 5 | 25 | |
| 34 | Góra Św. Jerzego e | 50°58′25.105” 16°20′0.496” | 354 | 9 | Basanite | × | 11 | 19 |
| 35 | Krzyżowa Góra e | 50°58′24.923” 16°20′11.138” | 358 | 8 | Basanite | × | 13 | 15 |
| [w/w%] | Knorrberg | Hutberg | Kleiner Hutberg | Landeskrone | Koło Obserwatora | Stożek Perkuna | Krzyżowa Góra | Zamkowa | Wilcza Góra | Łysanka | Krzyżowa Góra | Kopista | Góra Św. Jerzego | Krzyżowa Góra |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SiO2 | 47.2 | 39.3 | 40.4 | 41 | 42.06 | 39.8 | 42.87 | 40.49 | 40.66 | 43.66 | 40.09 | 47.15 | 45.35 | 45.88 |
| TiO2 | 2.9 | 3.06 | 2.99 | 3.18 | 2.59 | 2.61 | 3.29 | 3.2 | 2.64 | 2.44 | 2.6 | 2.29 | 2.38 | 2.1 |
| Al2O3 | 14.7 | 12 | 11.5 | 11.7 | 12.69 | 10.72 | 14.08 | 12.59 | 13.31 | 14.22 | 12.73 | 14.56 | 15.89 | 15.43 |
| Fe2O3 | 13.8 | 13.7 | 13.4 | 13.9 | 11.15 | 12.14 | 12.86 | 12.23 | 12 | 12.03 | 12.25 | 11.29 | 11.52 | 11.46 |
| MnO | 0.29 | 0.27 | 0.26 | 0.25 | 0.24 | 0.2 | 0.18 | 0.18 | 0.21 | 0.18 | 0.19 | 0.17 | 0.16 | 0.17 |
| MgO | 4.4 | 11.2 | 11.4 | 11.4 | 9.92 | 15.87 | 8.92 | 12.86 | 11.6 | 11.56 | 11.75 | 9.38 | 8.7 | 9.9 |
| CaO | 9.6 | 13.6 | 13.4 | 13.1 | 10.99 | 12.38 | 10.37 | 11.2 | 12.25 | 9.77 | 12.46 | 9.15 | 10.13 | 9.74 |
| Na2O | 4.16 | 4.61 | 4.2 | 3.7 | 3.8 | 3.04 | 3.42 | 3.4 | 3.32 | 3.67 | 3.31 | 2.89 | 3.16 | 3.23 |
| K2O | 1.72 | 1.12 | 0.96 | 0.84 | 0.96 | 1.14 | 1.31 | 1.11 | 1.01 | 1.12 | 0.9 | 0.82 | 0.83 | 0.88 |
| P2O5 | 1.14 | 1.05 | 1.07 | 0.87 | 0.98 | 0.96 | 0.73 | 0.71 | 0.66 | 0.66 | 1.07 | 0.4 | 0.42 | 0.35 |
| others | 0.09 | 0.36 | 1.38 | 1.77 | 4.62 | 1.14 | 1.96 | 1.93 | 2.34 | 0.69 | 2.65 | 1.9 | 2.09 | 0.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwiatkowski, P. Basaltic Outcrops as Centers of Diversity for Xerothermic Plants in the Sudetes Mountains (Central Europe). Diversity 2021, 13, 164. https://doi.org/10.3390/d13040164
Kwiatkowski P. Basaltic Outcrops as Centers of Diversity for Xerothermic Plants in the Sudetes Mountains (Central Europe). Diversity. 2021; 13(4):164. https://doi.org/10.3390/d13040164
Chicago/Turabian StyleKwiatkowski, Paweł. 2021. "Basaltic Outcrops as Centers of Diversity for Xerothermic Plants in the Sudetes Mountains (Central Europe)" Diversity 13, no. 4: 164. https://doi.org/10.3390/d13040164
APA StyleKwiatkowski, P. (2021). Basaltic Outcrops as Centers of Diversity for Xerothermic Plants in the Sudetes Mountains (Central Europe). Diversity, 13(4), 164. https://doi.org/10.3390/d13040164