
Inter-Individual Differences in Ornamental Colouration in a Mediterranean Lizard in Relation to Altitude, Season, Sex, Age, and Body Traits





Abstract
1. Introduction
- (1)
- Morphometric traits (SVL, body mass, and head size). Lizard morphometric traits strongly correlate with fitness. Body size is positively related to reproductive success in males [35,37,41] and females [42]. Head size is typically related to bite force, representing fighting ability, so it is related to social dominance in P. algirus [37]. While it is well known that head breeding colouration (orange) indicates body size and fighting ability in the western clade [35,37,41], intervening in communication during agonist encounters, whether that is also the case of colour patterns in the eastern clade remains poorly understood.
- (2)
- Sex. Albeit well reported in the western clade [35,36], sexual dichromatism is understudied in eastern populations (but see [39]). Some degree of dichromatism is expected given that sexual selection is typically stronger in males, especially in polygynous species [2]. While the mating system in the eastern clade is unknown, lizards from the western clade are polygynous; a male’s territory overlaps with those of several females [35,37].
- (3)
- Age. In the western clade, adult males are well differentiated from immature males by head colouration, but how colouration patterns vary with the advances of years is still unknown. In fact, in the western clade, it is well-determined that male head colouration varies with body size [35]. However, given that lizards are indeterminate growers, body size increases with age, so it is unclear whether colour signals indicate body size or age in this lizard. Males indicating old age may be preferred by females as their signals would indicate longevity and hence individual quality [43].
- (4)
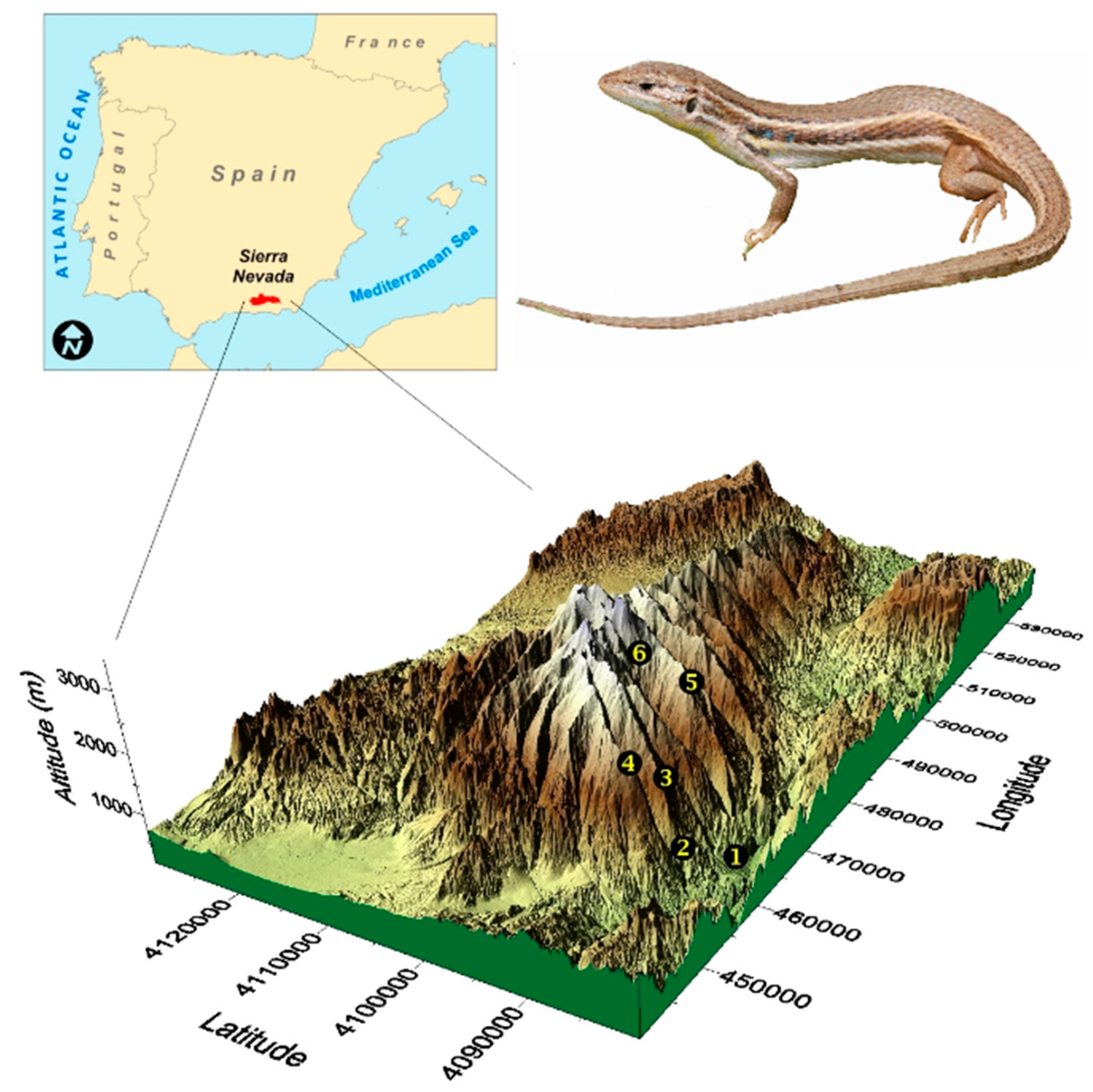
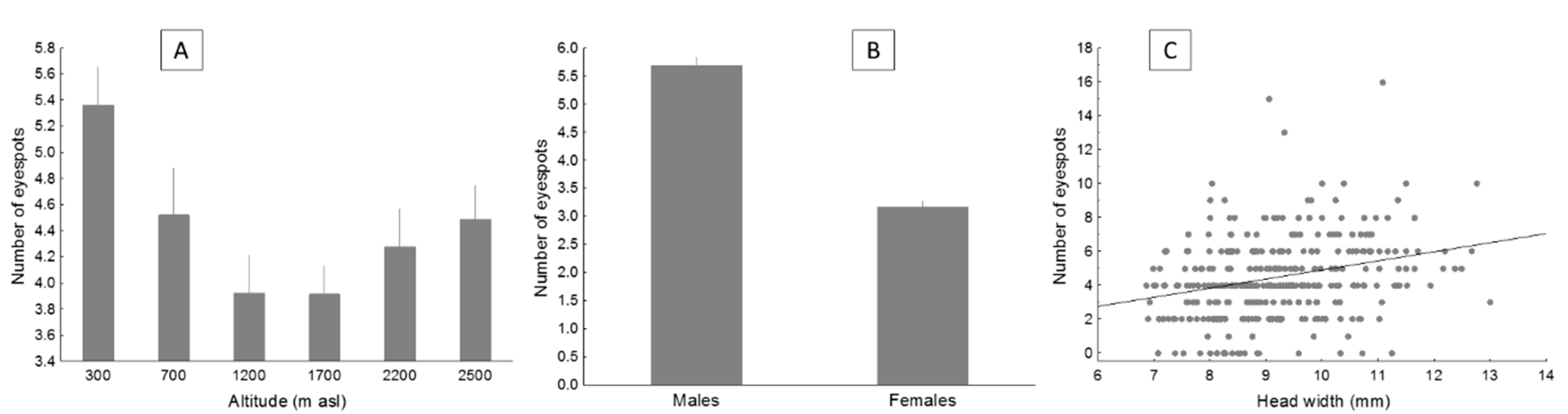
- Altitude. We examine how social colouration varies with altitude along a 2200-m elevational gradient. In a population of the western clade, lizards from localities separated 650 m in altitude differed in colouration, low-elevation individuals having more saturated colour in throats and more eyespots than high-elevation conspecifics [29,30]. However, whether colouration in populations from the eastern clade similarly varies with altitude remains unknown.
- (5)
2. Material and Methods
2.1. Sampling and General Procedures
2.2. Quantification of Colour Patches
2.3. Statistical Analyses
3. Results
3.1. Relationship among Colour Parameters
3.2. Correlates of Colouration
3.3. The Effect of Age
4. Discussion
4.1. Colouration as Indicators of Fighting Ability, Sex, and Age
4.2. Altitudinal Variation in Colouration
- (1)
- Population density might affect the investment in social communication. In more densely populated zones, social encounters should be more frequent, conducing to increased contests. Given that colour signals in P. algirus apparently inform about fighting ability, one could expect more investment in social signals in zones with denser populations. However, this hypothesis is not supported, as density was maximal at mid-elevation, and minimal at lowlands [44], where the number of eyespots was the highest. Therefore, altitudinal variation in population density did not covary with either variation in the number of eyespots or throat saturation.
- (2)
- We could also expect a trade-off between investment in signals and in self-maintenance [2]. Therefore, in zones where lizards invest more in longevity, social signals are expected to be less expressed (i.e., the pattern of longevity and signal intensity should be inverse). This hypothesis was not supported either, as longevity followed a U-shape with altitude [72], hence showing a pattern not consistent with a trade-off between self-maintenance and investment in colouration. Eyespots also followed a U-shaped pattern and throat colouration increased with elevation.
- (3)
- Temperature is the main environmental factor that varies with elevation. Colder temperatures at higher elevations may limit activity, especially for ectotherms [44,73]. Moreover, elevated temperatures may favour sexual selection [74]. Although this could partially explain the highest number of eyespots at low elevations, hardly could it explain why investment in throat colouration increases with ascending altitude.
- (4)
- Several colour traits in lizards are sensitive to parasites [36,75,76,77,78,79,80]. However, parasites follow a complex pattern with altitude in our study system, mite abundance decreasing, while haemoparasites prevalence increases, with ascending altitude [81]. Still, different types of colouration may be related to different types of parasites [82].
- (5)
- Food availability increases with elevation in our study population [83]. Chromatic properties of lizard skin correlate with pigment density, highly saturated colour patches having more pigment density than paler and duller ones [84]. Therefore, if throat pigment concentration depends on food (e.g., if they are carotenoid-dependent), the increase in food availability could explain the highest investment in throat colouration with altitude [85,86]. The altitudinal pattern for eyespots would be different as structural colour are presumed to be less affected by food availability.
- (6)
- Lizards at higher elevations suffer less oxidative damage than low-elevation ones [87,88]. Although we do not know if the yellow colouration in the throat is mediated by carotenoids or by pterins, both pigments may have antioxidant properties mainly mediated throughout a regulatory effect on the immune system [89]. Therefore, lizards from a higher elevation, exposed to a less oxidant ambient, could invest more in pigment-mediated social signals [90]. Meanwhile, the structural colouration of blue eyespots could be unaffected by oxidative stress.
- (7)
- (8)
- The efficiency of colour signals depends on the context where the visual stimulus is produced, as environmental conditions also affect the dispersion of the signal [94]. Blue-ultraviolet colours (short wavelengths) are more effective in partially covered habitats, while yellow-orange colourations are more effective in open areas [94]. Low elevation sites, where eyespots are more numerous, are composed of Mediterranean forests, with low arboreal cover, loose bushes, and a matrix of open and forestry zones. In this type of habitat, short-length colours would be favoured. Meanwhile, high elevation sites are above the treeline, with a habitat composed of short and compact scrubs. In these sites, by contrast, yellow colouration might be favoured.
4.3. Comparison with the Western Clade
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 300 | 700 | 1200 | 1700 | 2200 | 2500 | Total | |
|---|---|---|---|---|---|---|---|
| SVL | 61|45 | 20|36 | 19|30 | 37|36 | 42|50 | 60|46 | 482 |
| Body Mass | 59|44 | 20|34 | 18|30 | 36|36 | 42|50 | 60|46 | 475 |
| Head Width | 60|43 | 19|29 | 16|29 | 37|33 | 39|48 | 57|45 | 460 |
| Age | 11|12 | 8|11 | 10|10 | 10|9 | 9|7 | 11|10 | 118 |
| Eyespots | 44|28 | 19|29 | 16|23 | 37|32 | 32|37 | 43|31 | 371 |
| Throat Colour (LCH) | 59|45 | 19|35 | 17|30 | 36|35 | 42|49 | 60|46 | 473 |
| Commissure Patch Colour | 33|22 | 11|15 | 14|19 | 32|25 | 27|24 | 35|24 | 281 |
| Commissure Patch Size | 16|1 | 5|2 | 5|2 | 20|6 | 18|5 | 25|9 | 114 |
Appendix B

References
- Bradbury, J.W.; Vehrencamp, S.L. Principles of Animal Communication; Sinauer: Sunderland, MA, USA, 2011. [Google Scholar]
- Andersson, M. Sexual Selection; Princeton University Press: Princeton, NJ, USA, 1994. [Google Scholar]
- Stevens, M.; Merilaita, S. (Eds.) Animal Camouflage: Mechanisms and Function; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Pérez i de Lanuza, G.; Font, E. The evolution of colour pattern complexity: Selection for conspicuousness favours contrasting within-body colour combinations in lizards. J. Evol. Biol. 2016, 29, 942–951. [Google Scholar] [CrossRef]
- Andersson, S.; Pryke, S.R.; Örnborg, J.; Lawes, M.J.; Andersson, M. Multiple receivers, multiple ornaments, and a trade-off between agonistic and epigamic signaling in a widowbird. Am. Nat. 2002, 160, 683–691. [Google Scholar] [CrossRef]
- Candolin, U. The use of multiple cues in mate choice. Biol. Rev. 2003, 78, 575–595. [Google Scholar] [CrossRef] [PubMed]
- Martín, J.; López, P. Multiple color signals may reveal multiple messages in male Schreiber’s green lizards, Lacerta schreiberi. Behav. Ecol. Sociobiol. 2009, 63, 1743–1755. [Google Scholar] [CrossRef]
- Plasman, M.; Reynoso, V.H.; Nicolás, L.; Torres, R. Multiple colour traits signal performance and immune response in the Dickerson’s collared lizard Crotaphytus dickersonae. Behav. Ecol. Sociobiol. 2015, 69, 765–775. [Google Scholar] [CrossRef]
- Zamora-Camacho, F.J.; Comas, M. Beyond sexual dimorphism and habitat boundaries: Coloration correlates with morphology, age, and locomotor performance in a toad. Evol. Biol. 2019, 46, 60–70. [Google Scholar] [CrossRef]
- Møller, A.P.; Pomiankowski, A. Why birds got multiple ornaments? Behav. Ecol. Sociobiol. 1993, 32, 167–176. [Google Scholar] [CrossRef]
- Johnstone, R.A. Multiple displays in animal communication: “Backup signals” and “multiple messages”. Philos. Trans. R. Soc. B 1996, 351, 329–338. [Google Scholar]
- Grether, G.F.; Kolluru, G.R.; Nersissian, K. Individual colour patches as multicomponent signals. Biol. Rev. 2004, 79, 583–610. [Google Scholar] [CrossRef]
- Olsson, M.; Stuart-Fox, D.; Ballen, C. Genetics and evolution of colour patterns in reptiles. Semin. Cell Dev. Biol. 2013, 24, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Olsson, M. Nuptial colouration in the sand lizard, Lacerta agilis: An intra-sexually selected cue to fighting ability. Anim. Behav. 1994, 48, 607–613. [Google Scholar] [CrossRef]
- Whiting, M.J.; Stuart-Fox, D.M.; O’Connor, D.; Firth, D.; Bennett, N.C.; Blomberg, S.P. Ultraviolet signals ultra-aggression in a lizard. Anim. Behav. 2006, 72, 353–363. [Google Scholar] [CrossRef]
- Bajer, K.; Molnár, O.; Török, J.; Herczeg, G. Ultraviolet nuptial colour determines fight success in male European green lizards (Lacerta viridis). Biol. Lett. 2011, 7, 866–868. [Google Scholar] [CrossRef] [PubMed]
- Baird, T.A. Reproductive coloration in female collared lizards, Crotophytus collaris, stimulates courtship by males. Herpetologica 2004, 60, 337–348. [Google Scholar] [CrossRef]
- Salica, M.J.; Halloy, M. Nuptial coloration in female Liolaemus quilmes (Iguania: Liolaemidae): Relation to reproductive state. Rev. Esp. Herpetol. 2009, 23, 141–149. [Google Scholar]
- Stuart-Fox, D.; Goode, J.L. Female ornamentation influences male courtship investment in a lizard. Front. Ecol. Evol. 2014, 2, 2. [Google Scholar] [CrossRef]
- Díaz, J.A.; Alonso-Gomez, A.L.; Delgado, M.J. Seasonal variation of gonadal development, sexual steroids, and lipid reserves in a population of the lizard Psammodromus algirus. J. Herpetol. 1994, 28, 199–205. [Google Scholar] [CrossRef]
- Hews, D.K.; Moore, M.C. Influence of androgens on differentiation of secondary sex characters in tree lizards, Urosaurus ornatus. Gen. Comp. Endocrinol. 1995, 97, 86–102. [Google Scholar] [CrossRef]
- Cox, R.M.; Zilberman, V.; John-Alder, H.B. Testosterone stimulates the expression of a social color signal in Yarrow’s Spiny Lizard, Sceloporus jarrovii. J. Exp. Zool. A 2008, 309, 505–514. [Google Scholar] [CrossRef]
- Macedonia, J.M. Habitat light, colour variation, and ultraviolet reflectance in the Grand Cayman anole, Anolis conspersus. Biol. J. Linn. Soc. 2001, 73, 299–320. [Google Scholar] [CrossRef]
- Pérez i de Lanuza, G.; Carretero, M.A.; Font, E. Intensity of male-male competition predicts morph diversity in a color polymorphic lizard. Evolution 2017, 71, 1832–1840. [Google Scholar] [CrossRef]
- Moreno-Rueda, G.; González-Granda, L.G.; Reguera, S.; Zamora-Camacho, F.J.; Melero, E. Crypsis decreases with elevation in a lizard. Diversity 2019, 11, 236. [Google Scholar] [CrossRef]
- Keller, I.; Alexander, J.M.; Holderegger, R.; Edwards, P.J. Widespread phenotypic and genetic divergence along altitudinal gradients in animals. J. Evol. Biol. 2013, 26, 2527–2543. [Google Scholar] [CrossRef] [PubMed]
- Badyaev, A.V. Altitudinal variation in sexual dimorphism: A new pattern and alternative hypotheses. Behav. Ecol. 1997, 8, 675–690. [Google Scholar] [CrossRef]
- Leaché, A.D.; Helmer, D.-S.; Moritz, C. Phenotypic evolution in high-elevation populations of western fencce lizards (Sceloporus occidentalis) in the Sierra Nevada Mountains. Biol. J. Linn. Soc. 2010, 100, 630–641. [Google Scholar] [CrossRef]
- Iraeta, P.; Monasterio, C.; Salvador, A.; Díaz, J.A. Sexual dimorphism and interpopulation differences in lizard hind limb length: Locomotor performance or chemical signalling? Biol. J. Linn. Soc. 2011, 104, 318–329. [Google Scholar] [CrossRef]
- Llanos-Garrido, A.; Díaz, J.A.; Pérez-Rodríguez, A.; Arriero, E. Variation in male ornaments in two lizard populations with contrasting parasite loads. J. Zool. 2017, 303, 218–225. [Google Scholar] [CrossRef]
- Martín, J.; Zamora-Camacho, F.J.; Reguera, S.; López, P.; Moreno-Rueda, G. Variations in chemical sexual signals of Psammodromus algirus lizards along an elevation gradient may reflect altitudinal variation in microclimatic conditions. Sci. Nat. 2017, 104, 16. [Google Scholar] [CrossRef]
- Ortega, J.; Martín, J.; Crochet, P.-A.; López, P.; Clobert, J. Seasonal and interpopulational phenotypic variation in morphology and sexual signals of Podarcis liolepis lizards. PLoS ONE 2019, 14, e0211686. [Google Scholar] [CrossRef]
- Salvador, A. Lagartija colilarga—Psammodromus algirus (Linnaeus, 1758). In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Marco, A., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2015. [Google Scholar]
- Carranza, S.; Harris, D.J.; Arnold, E.N.; Batista, V.; Gonzalez De La Vega, J.P. Phylogeography of the lacertid lizard, Psammodromus algirus, in Iberia and across the Strait of Gibraltar. J. Biogeogr. 2006, 33, 1279–1288. [Google Scholar] [CrossRef]
- Díaz, J.A. Breeding coloration, mating opportunities, activity, and survival in the lacertid lizard Psammodromus algirus. Can. J. Zool. 1993, 71, 1104–1110. [Google Scholar] [CrossRef]
- Salvador, A.; Veiga, J.P.; Martín, J.; López, P.; Abelenda, M.; Puerta, M. The cost of producing a sexual signal: Testosterone increases the susceptibitily of male lizards to ectoparasitic infestation. Behav. Ecol. 1996, 7, 145–150. [Google Scholar] [CrossRef]
- Salvador, A.; Veiga, J.P. Male traits and pairing success in the lizard Psammodromus algirus. Herpetologica 2001, 57, 77–86. [Google Scholar]
- Salvador, A.; Veiga, J.P.; Martín, J.; López, P. Testosterone supplementation in subordinate, small male lizards: Consequences for aggressiveness, color development, and parasite load. Behav. Ecol. 1997, 8, 135–139. [Google Scholar] [CrossRef]
- Carretero, M.A. Sources of colour pattern variation in Mediterranean Psammodromus algirus. Neth. J. Zool. 2002, 52, 43–60. [Google Scholar] [CrossRef]
- Salvador, A.; Veiga, J.P. A permanent signal related to male pairing success and survival in the lizard Psammodromus algirus. Amphibia Reptilia 2008, 29, 117–120. [Google Scholar]
- Martín, J.; Forsman, A. Social costs and development of nuptial coloration in male Psammodromus algirus lizards: An experiment. Behav. Ecol. 1999, 10, 396–400. [Google Scholar] [CrossRef]
- Díaz, J.A.; Iraeta, P.; Verdú-Ricoy, J.; Siliceo, I.; Salvador, A. Intraspecific variation of reproduction traits in a Mediterranean lizard: Clutch, population, and lineage effects. Evol. Biol. 2012, 39, 106–115. [Google Scholar] [CrossRef]
- Brooks, R.C.; Kemp, D.J. Can older males deliver the good genes? Trends Ecol. Evol. 2001, 16, 308–313. [Google Scholar] [CrossRef]
- Zamora-Camacho, F.J.; Reguera, S.; Moreno-Rueda, G.; Pleguezuelos, J.M. Patterns of seasonal activity in a Mediterranean lizard along a 2200m altitudinal gradient. J. Thermal Biol. 2013, 38, 64–69. [Google Scholar] [CrossRef]
- Zamora-Camacho, F.J.; Reguera, S.; Moreno-Rueda, G. Bergmann’s Rule rules body size in an ectotherm: Heat conservation in a lizard along a 2200-metre elevational gradient. J. Evol. Biol. 2014, 27, 2820–2828. [Google Scholar] [CrossRef] [PubMed]
- Perry, G.; Wallace, M.C.; Perry, D.; Curzer, H.; Muhlberger, P. Toe clipping of amphibians and reptiles: Science, ethics, and the law. J. Herpetol. 2011, 45, 547–555. [Google Scholar] [CrossRef]
- Comas, M.; Reguera, S.; Zamora-Camacho, F.J.; Salvadó, H.; Moreno-Rueda, G. Comparison of the effectiveness of phalanges vs. humeri and femurs to estimate lizard age with skeletochronology. Anim. Biodiv. Conserv. 2016, 39, 237–240. [Google Scholar] [CrossRef]
- Zhao, M.; Klaassen, C.A.J.; Lisovski, S.; Klaassen, M. The adequacy of aging techniques in vertebrates for rapid estimation of population mortality rates from age distributions. Ecol. Evol. 2019, 9, 1394–1402. [Google Scholar] [CrossRef]
- Andersson, S.; Prager, M. Quantifying colors. In Bird Coloration. Vol. II: Function and Evolution; Hill, G.E., McGraw, K.J., Eds.; Harvard University Press: Boston, MA, USA, 2006; pp. 41–89. [Google Scholar]
- Montgomerie, R. Analazing colors. In Bird Coloration. Vol. I: Mechanisms and Measurements; Hill, G.E., McGraw, K.J., Eds.; Harvard University Press: Boston, MA, USA, 2006; pp. 90–147. [Google Scholar]
- Endler, J.A. On the measurement and classification of colour in studies of animal colour patterns. Biol. J. Linn. Soc. 1990, 41, 315–352. [Google Scholar] [CrossRef]
- Rasband, W.S. ImageJ; US National Institutes of Health: Bethesda, MD, USA, 2008. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Packard, G.C. Is logarithmic transformation necessary in allometry? Biol. J. Linn. Soc. 2013, 109, 476–486. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Ripley, B.; Venables, W. nnet: Feed-Forward Neural Networks and Multinomial Log-Linear Models; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Bartoń, K. MuMIn: Multi-Model Inference; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Reguera, S.; Zamora-Camacho, F.J.; Moreno-Rueda, G. The lizard Psammodromus algirus (Squamata: Lacertidae) is darker at high altitudes. Biol. J. Linn. Soc. 2014, 112, 132–141. [Google Scholar] [CrossRef]
- Huyghe, K.; Vanhooydonck, B.; Scheers, H.; Molina-Borja, M.; van Damme, R. Morphology, performance and fighting capacity in male lizards, Gallotia galloti. Funct. Ecol. 2005, 19, 800–807. [Google Scholar] [CrossRef]
- Stapley, J.; Whiting, M.J. Ultraviolet signals fighting ability in a lizard. Biol. Lett. 2006, 2, 169–172. [Google Scholar] [CrossRef]
- Pérez i de Lanuza, G.; Carazo, P.; Font, E. Colours of quality: Structural (but not pigment) coloration informs about male quality in a polychromatic lizard. Anim. Behav. 2014, 90, 73–81. [Google Scholar] [CrossRef]
- Hamilton, D.G.; Whiting, M.J.; Pryke, S.R. Fiery frills: Carotenoid-based coloration predicts contest success in frillneck lizards. Behav. Ecol. 2013, 24, 1138–1149. [Google Scholar] [CrossRef]
- Møller, A.P. Social control of deception among status signalling house sparrows Passer domesticus. Behav. Ecol. Sociobiol. 1987, 20, 307–311. [Google Scholar] [CrossRef]
- Lappin, A.K.; Brandt, Y.; Husak, J.F.; Macedonia, J.M.; Kemp, D.J. Gaping displays reveal and amplify a mechanically based index of weapon performance. Am. Nat. 2006, 168, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.L. Female-specific color is a signal of quality in the striped plateau lizard (Sceloporus virgatus). Behav. Ecol. 2006, 17, 726–732. [Google Scholar] [CrossRef]
- Lande, R. Sexual dimorphism, sexual selection, and adaptation in polygenic characters. Evolution 1980, 34, 292–305. [Google Scholar] [CrossRef]
- López, P.; Martín, J.; Cuadrado, M. Chemosensory cues allow male lizards Psammodromus algirus to override visual concealment of sexual identity by satellite males. Behav. Ecol. Sociobiol. 2003, 54, 218–224. [Google Scholar] [CrossRef]
- Searcy, W.A.; Nowicki, S. The Evolution of Animal Communication; Princeton University Press: Princeton, NJ, USA, 2005. [Google Scholar]
- Martín, J.; López, P. Nuptial coloration and mate guarding affect escape decision of male lizards Psammodromus algirus. Ethology 1999, 105, 439–447. [Google Scholar] [CrossRef]
- Dunn, P.O.; Armenta, J.K.; Whittingham, L.A. Natural and sexual selection act on different axes of variation in avian plumage color. Sci. Adv. 2015, 1, e1400155. [Google Scholar] [CrossRef] [PubMed]
- Comas, M.; Reguera, S.; Zamora-Camacho, F.J.; Moreno-Rueda, G. Age structure of a lizard along an elevational gradient reveals nonlinear lifespan patterns with altitude. Curr. Zool. 2020, 142, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Camacho, F.J.; Reguera, S.; Moreno-Rueda, G. Thermoregulation in the lizard Psammodromus algirus along a 2200-m elevational gradient in Sierra Nevada (Spain). Int. J. Biometeorol. 2016, 60, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Olsson, M.; Wapstra, E.; Schwartz, T.; Madsen, T.; Ujvari, B.; Uller, T. In hot pursuit: Fluctuating mating system and sexual selection in san lizards. Evolution 2011, 65, 574–583. [Google Scholar] [CrossRef] [PubMed]
- Václav, R.; Prokop, P.; Fekiač, V. Expression of breeding coloration in European Green Lizards (Lacerta viridis): Variation with morphology and tick infestation. Can. J. Zool. 2007, 85, 1199–1206. [Google Scholar] [CrossRef]
- Calisi, R.M.; Malone, J.H.; Hews, D.K. Female secondary coloration in the Mexican boulder spiny lizard is associated with nematode load. J. Zool. 2008, 276, 358–367. [Google Scholar] [CrossRef]
- Martín, J.; Amo, L.; López, P. Parasites and health affect multiple sexual signals in male common wall lizards, Podarcis muralis. Naturwissenschaften 2008, 95, 293–300. [Google Scholar] [CrossRef]
- Megía-Palma, R.; Martínez, J.; Merino, S. A structural colour ornament correlates positively with parasite load and body condition in an insular lizard species. Sci. Nat. 2016, 103, 52. [Google Scholar] [CrossRef]
- Megía-Palma, R.; Martínez, J.; Merino, S. Manipulation of parasite load induces significant changes in the structural-based throat color of male Iberian green lizards. Curr. Zool. 2018, 64, 293–302. [Google Scholar] [CrossRef]
- Megía-Palma, R.; Paranjpe, D.; Reguera, S.; Martínez, J.; Cooper, R.D.; Blaimont, P.; Merino, S.; Sinervo, B. Multiple color patches and parasites in Sceloporus occidentalis: Differential relationships by sex and infection. Curr. Zool. 2018, 64, 703–711. [Google Scholar] [CrossRef]
- Álvarez-Ruiz, L.; Megía-Palma, R.; Reguera, S.; Ruiz, S.; Zamora-Camacho, F.J.; Figuerola, J.; Moreno-Rueda, G. Opposed elevational variation in prevalence and intensity of endoparasites and their vectors in a lizard. Curr. Zool. 2018, 64, 197–204. [Google Scholar] [CrossRef]
- Megía-Palma, R.; Martínez, J.; Merino, S. Structural- and carotenoid-based throat colour patches in males of Lacerta schreiberi reflect different parasitic diseases. Behav. Ecol. Sociobiol. 2016, 70, 2017–2025. [Google Scholar]
- Moreno-Rueda, G.; Melero, E.; Reguera, S.; Zamora-Camacho, F.J.; Álvarez-Benito, I. Prey availability, prey selection, and trophic niche width in the lizard Psammodromus algirus along an elevational gradient. Curr. Zool. 2018, 64, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Cuervo, J.J.; Belliure, J.; Negro, J.J. Coloration reflects skin pterin concentration in a red-tailed lizard. Comp. Biochem. Physiol. B 2016, 193, 17–24. [Google Scholar] [CrossRef]
- Hill, G.E. Geographic variation in the carotenoid plumage pigmentation of male house finches (Carpodacus mexicanus). Biol. J. Linn. Soc. 1993, 49, 63–86. [Google Scholar] [CrossRef]
- Grether, G.F.; Hudon, J.; Millie, D.F. Carotenoid limitation of sexual coloration along an environmental gradient in guppies. Proc. R. Soc. B 1999, 266, 1317–1322. [Google Scholar] [CrossRef]
- Reguera, S.; Zamora-Camacho, F.J.; Trenzado, C.E.; Sanz, A.; Moreno-Rueda, G. Oxidative stress decreases with elevation in the lizard Psammodromus algirus. Comp. Biochem. Physiol. A 2014, 172, 52–56. [Google Scholar] [CrossRef]
- Reguera, S.; Zamora-Camacho, F.J.; Melero, E.; García-Mesa, S.; Trenzado, C.E.; Cabrerizo, M.J.; Sanz, A.; Moreno-Rueda, G. Ultraviolet radiation does not increase oxidative stress in the lizard Psammodromus algirus along an elevational gradient. Comp. Biochem. Physiol. A 2015, 183, 20–26. [Google Scholar] [CrossRef]
- McGraw, K.J. The antioxidant function of many animal pigments: Are there consistent health benefits of sexually selected colourants? Anim. Behav. 2005, 69, 757–764. [Google Scholar] [CrossRef]
- Olsson, M.; Tobler, M.; Healey, M.; Perrin, C.; Wilson, M. A significant component of ageing (DNA damage) is reflected in fading breeding colors: An experimental test using innate antioxidant mimetics in painted dragon lizards. Evolution 2012, 66, 2475–2483. [Google Scholar] [CrossRef]
- Jennions, M.D.; Petrie, M. Variation in mate choice and mating preferences: A review of causes and consequences. Biol. Rev. 1997, 72, 283–327. [Google Scholar] [CrossRef]
- Brooks, R.C.; Couldridge, V. Multiple sexual ornaments coevolve with multiple mating preferences. Am. Nat. 1999, 154, 37–45. [Google Scholar] [CrossRef]
- Kwiatkowski, M.A.; Sullivan, B.K. Geographic variation in sexual selection among populations of an iguanid lizard, Sauromalus obesus (=ater). Evolution 2002, 56, 2039–2051. [Google Scholar] [CrossRef] [PubMed]
- Endler, J.A. The color of light in forests and its implications. Ecol. Monogr. 1993, 63, 1–27. [Google Scholar] [CrossRef]
- Díaz, J.A.; Verdú-Ricoy, J.; Iraeta, P.; Llanos-Garrido, A.; Pérez-Rodríguez, A.; Salvador, A. There is more to the picture than meets the eye: Adaptation for crypsis blurs phylogeographical structure in a lizard. J. Biogeogr. 2017, 44, 397–408. [Google Scholar] [CrossRef]
- Stuart-Fox, D.M.; Moussalli, A.; Marshall, N.J.; Owens, I.P.F. Conspicuous males suffer higher predation risk: Visual modelling and experimental evidence from lizards. Anim. Behav. 2003, 66, 541–550. [Google Scholar] [CrossRef]








| Correlations | Commissure Colour | ||||||
|---|---|---|---|---|---|---|---|
| Throat Lightness | Throat Chroma | Throat Hue | Commissure Area | No (138) | Yellow (33) | Orange (106) | |
| Eyespots | r362 = 0.02 p = 0.71 | r362 = 0.03 p = 0.51 | r362 = 0.08 p = 0.12 | r110 = 0.04 p = 0.71 | 3.64 a ± 0.18 | 3.85 a ± 0.44 | 5.79 b ± 0.20 |
| Throat Lightness | r473 = −0.51 p < 0.001 | r473 = 0.21 p < 0.001 | r114 = −0.30 p < 0.001 | 81.28 a ± 0.37 | 76.09 b ± 1.12 | 79.57 c ± 0.44 | |
| Throat Chroma | r473 = −0.13 p = 0.006 | r114 = 0.46 p < 0.001 | 12.20 a ± 0.36 | 20.10 b ± 1.58 | 17.19 b ± 0.92 | ||
| Throat Hue | r114 = −0.18 p = 0.06 | 88.86 ± 1.09 | 88.87 ± 2.10 | 87.70 ± 1.08 | |||
| Throat | Commissure | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Eyespots | Lightness | Chroma | Hue | Colour | Area | |||||
| Df | F-Value | df | F-Value | F-Value | F-Value | df | χ2 | df | F-Value | |
| Altitude | 5, 333 | 3.88 ** | 5, 430 | 8.48 *** | 3.25 ** | 6.12 *** | 10 | 4.08 | 5, 97 | 0.75 |
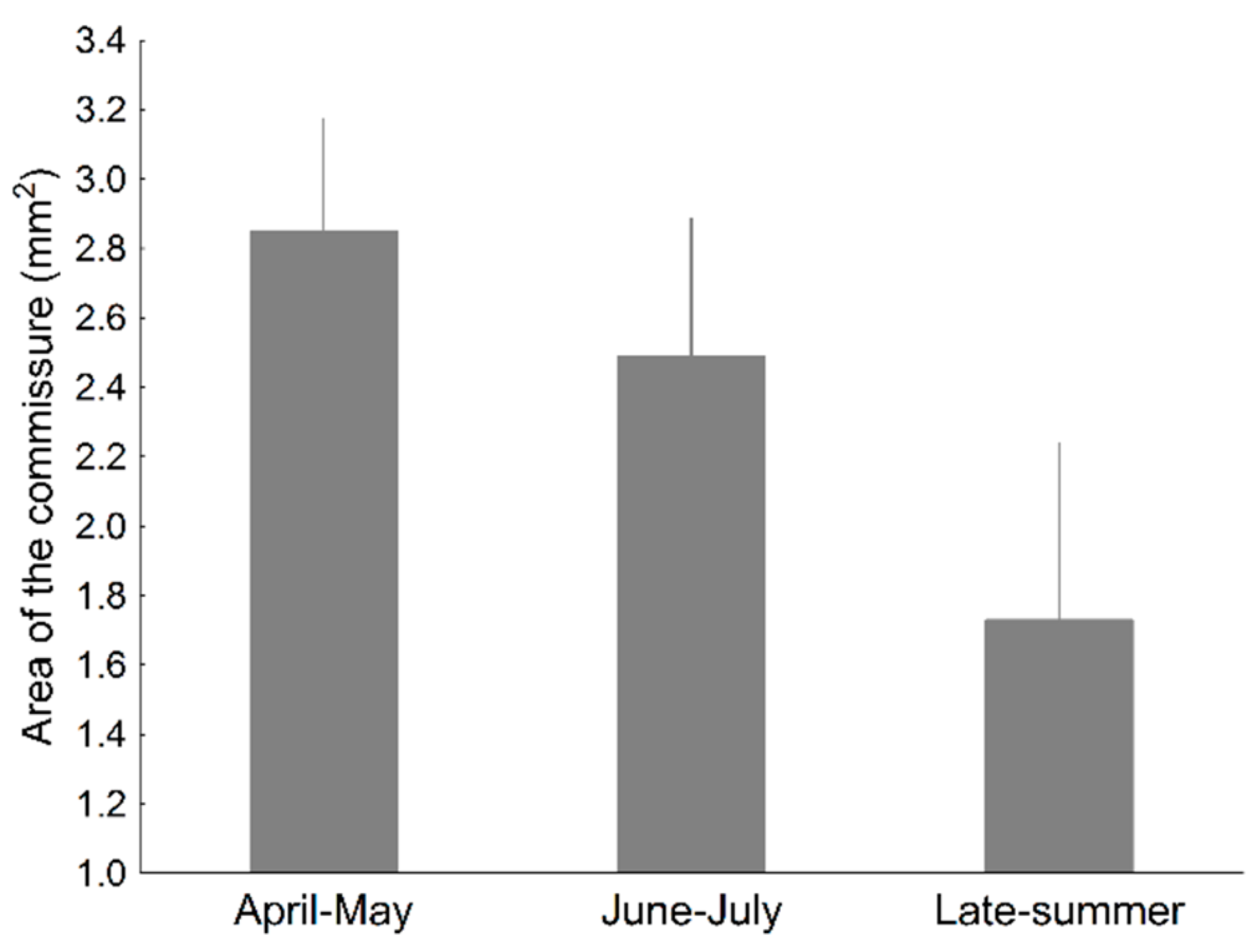
| Season | 2, 333 | 0.08 | 2, 430 | 12.79 *** | 10.93 *** | 8.16 *** | 4 | 10.38 * | 2, 97 | 4.96 ** |
| Year | 2, 333 | 1.18 | 3, 430 | 6.48 *** | 5.93 *** | 2.98 * | 4 | 1.92 | 2, 97 | 6.58 ** |
| Sex | 1, 333 | 57.62 *** | 1, 430 | 2.98 | 0.73 | 1.77 | 2 | 90.72 *** | 1, 97 | 2.23 |
| Mass | 1, 333 | 0.71 | 1, 430 | 0.99 | 2.83 | 0.31 | 2 | 0.99 | 1, 97 | 2.67 |
| SVL | 1, 333 | 1.44 | 1, 430 | 4.58 * | 8.50 ** | 1.12 | 2 | 0.73 | 1, 97 | 1.66 |
| Head | 1, 333 | 6.78 * | 1, 430 | 8.94 ** | 4.86 * | 1.12 | 2 | 8.99 * | 1, 97 | 0.15 |
| Dependent Variable | Predictors | df | AICc | ΔAIC | Weight |
|---|---|---|---|---|---|
| Eyespots | Altitude + Sex + Head | 9 | 1437.01 | 0 | 0.29 |
| Altitude + Sex + Head + SVL | 10 | 1438.80 | 1.79 | 0.12 | |
| Throat Lightness | Altitude + Season + Year + Head + SVL + Sex | 15 | −1161.73 | 0 | 0.31 |
| Altitude + Season + Year + Head + SVL + Sex + Mass | 16 | −1160.61 | 1.13 | 0.18 | |
| Altitude + Season + Year + Head + SVL | 14 | −1160.28 | 1.45 | 0.15 | |
| Altitude + Season + Year + Head + Sex | 14 | –1159.82 | 1.91 | 0.12 | |
| Throat Chroma | Altitude + Season + Year + Head + SVL + Mass | 15 | 2863.91 | 0 | 0.31 |
| Altitude + Season + Year + Head + SVL | 14 | 2864.35 | 0.45 | 0.25 | |
| Altitude + Season + Year + Head + SVL + Mass + Sex | 16 | 2865.30 | 1.40 | 0.15 | |
| Throat Hue | Altitude + Season + Year + Sex | 13 | –648.44 | 0 | 0.25 |
| Altitude + Season + Year + Sex + Head | 14 | –646.98 | 1.46 | 0.12 | |
| Altitude + Season + Year + Sex + Mass | 14 | –646.49 | 1.95 | 0.10 | |
| Commissure Colour | Season + Sex + Head + Mass | 12 | 272.67 | 0 | 0.41 |
| Season + Sex + Head + SVL | 12 | 273.09 | 0.42 | 0.33 | |
| Commissure Area | Year + Season + Mass + Head | 8 | 198.57 | 0 | 0.15 |
| Year + Season + Sex + Mass | 8 | 198.74 | 0.17 | 0.14 | |
| Year + Season + Sex + SVL + Mass | 9 | 198.87 | 0.30 | 0.13 | |
| Year + Season + SVL + Head | 8 | 199.25 | 0.68 | 0.11 | |
| Year + Season + Sex + SVL | 8 | 199.75 | 1.18 | 0.08 | |
| Year + Season + Sex + Mass + Head | 9 | 199.81 | 1.24 | 0.08 | |
| Year + Season + SVL + Mass + Head | 9 | 200.13 | 1.55 | 0.07 | |
| Year + Season + Sex + SVL + Head | 9 | 200.22 | 1.65 | 0.06 |
| Altitude | Season | Year | Sex | SVL | Mass | Head | |
|---|---|---|---|---|---|---|---|
| Eyespots | U-shaped | No | No | More in males | No | No | Increase |
| Throat Lightness | Darker at the highest altitudes | Lighter at the end of the season | Yes | No | Decrease | No | Decrease |
| Throat Chroma | Linear increase | Linear decrease | Yes | No | Increase (with age) | No | Increase |
| Throat Hue | Decrease | Linear increase | Yes | Yes | No | No | No |
| Commissure Colour | No | Less orange lizards with advanced season | No | Orange, males; yellow or colourless, females | No | No | Larger in coloured lizards |
| Commissure Area | No | Linear decrease | Yes | Larger in males | No | No | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Rueda, G.; Reguera, S.; Zamora-Camacho, F.J.; Comas, M. Inter-Individual Differences in Ornamental Colouration in a Mediterranean Lizard in Relation to Altitude, Season, Sex, Age, and Body Traits. Diversity 2021, 13, 158. https://doi.org/10.3390/d13040158
Moreno-Rueda G, Reguera S, Zamora-Camacho FJ, Comas M. Inter-Individual Differences in Ornamental Colouration in a Mediterranean Lizard in Relation to Altitude, Season, Sex, Age, and Body Traits. Diversity. 2021; 13(4):158. https://doi.org/10.3390/d13040158
Chicago/Turabian StyleMoreno-Rueda, Gregorio, Senda Reguera, Francisco J. Zamora-Camacho, and Mar Comas. 2021. "Inter-Individual Differences in Ornamental Colouration in a Mediterranean Lizard in Relation to Altitude, Season, Sex, Age, and Body Traits" Diversity 13, no. 4: 158. https://doi.org/10.3390/d13040158
APA StyleMoreno-Rueda, G., Reguera, S., Zamora-Camacho, F. J., & Comas, M. (2021). Inter-Individual Differences in Ornamental Colouration in a Mediterranean Lizard in Relation to Altitude, Season, Sex, Age, and Body Traits. Diversity, 13(4), 158. https://doi.org/10.3390/d13040158