Lichen Distribution Patterns in the Ecoregions of Italy

 ,
,  ,
,
Abstract
1. Introduction
2. Data and Methods
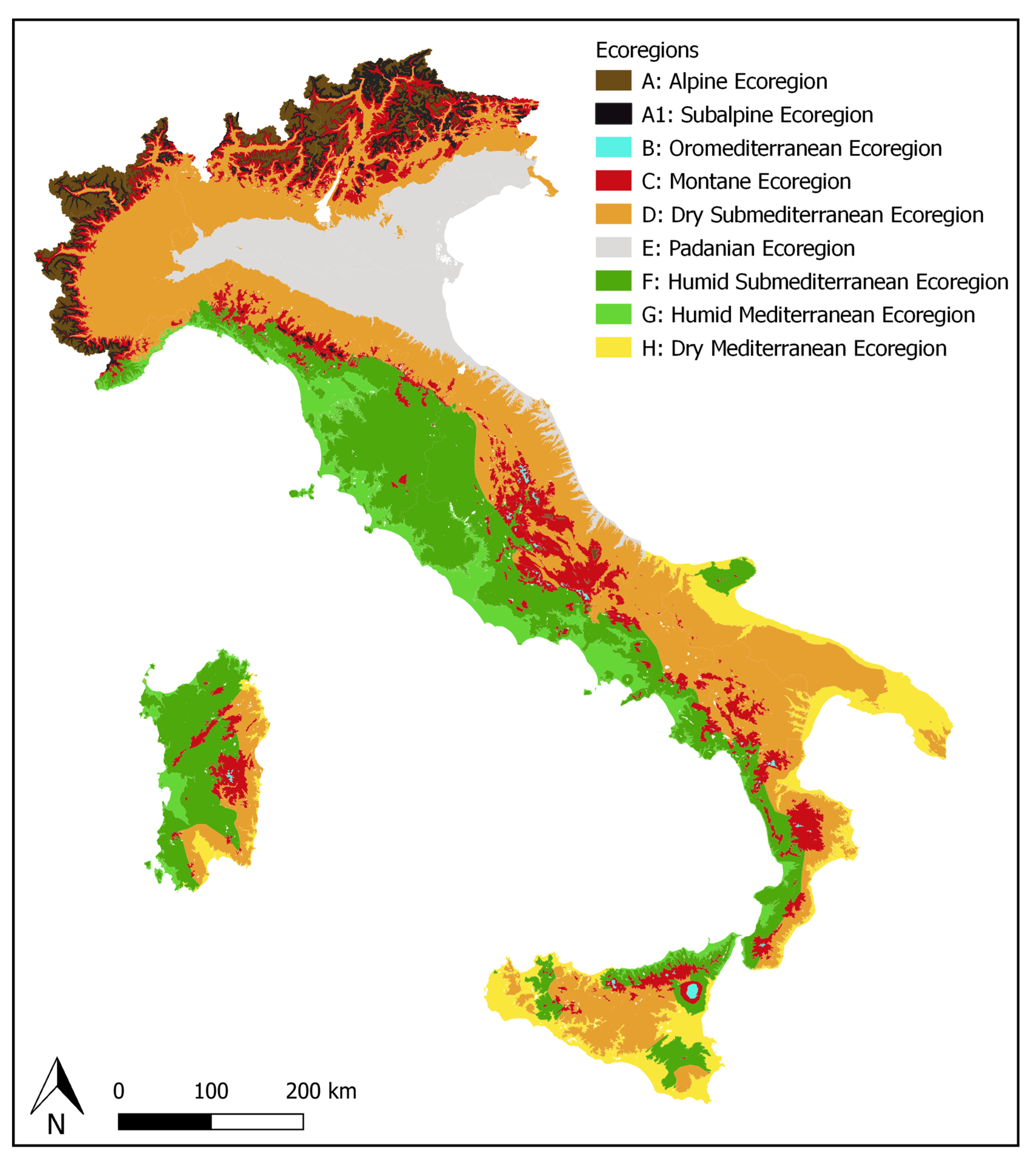
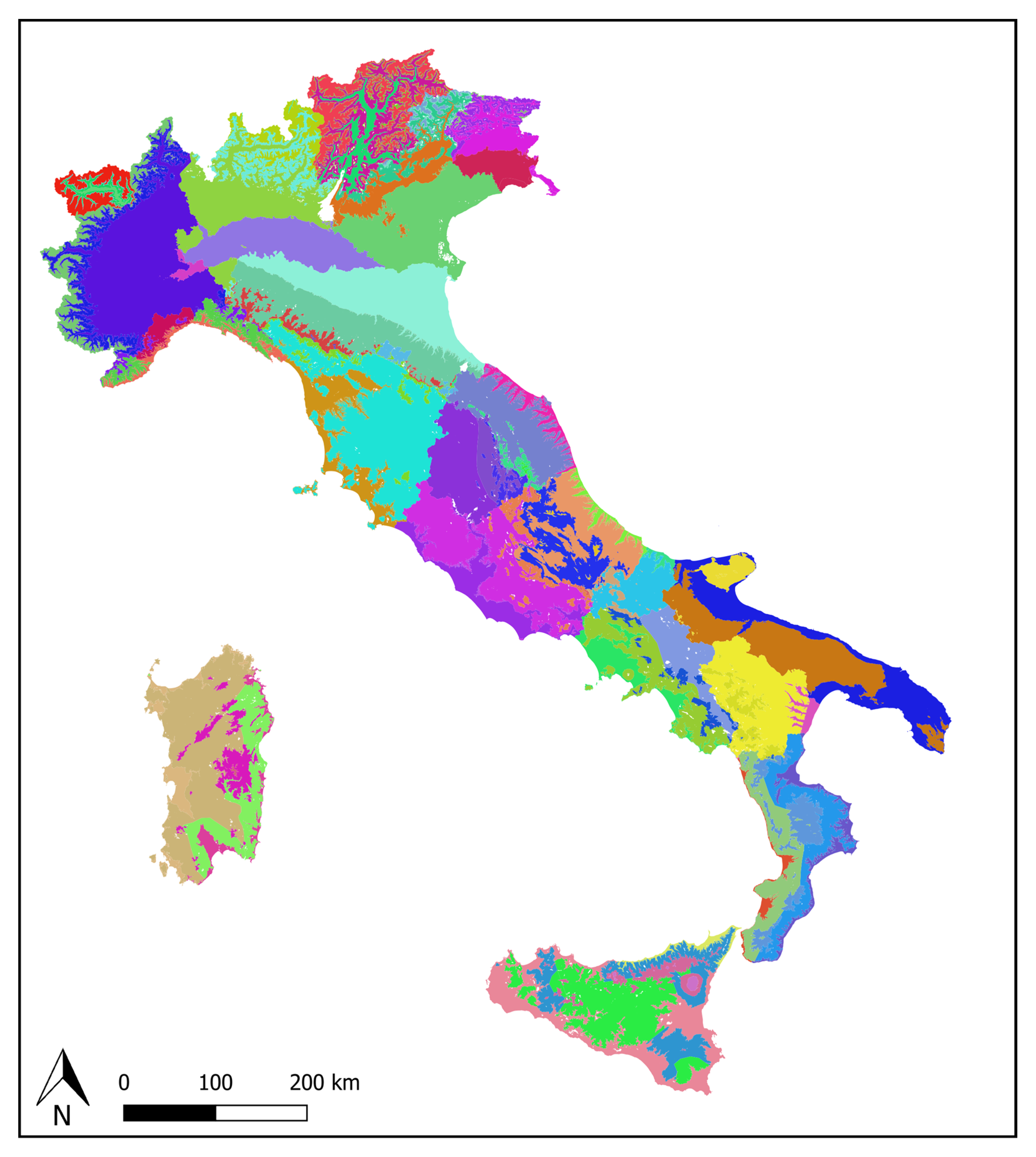
2.1. Ecoregions and OGUs
2.2. Functional Traits and Ecological Requirements
2.3. Environmental Data
2.4. Data Analysis
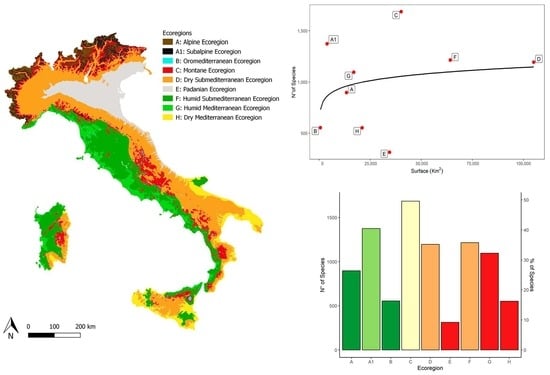
3. Results
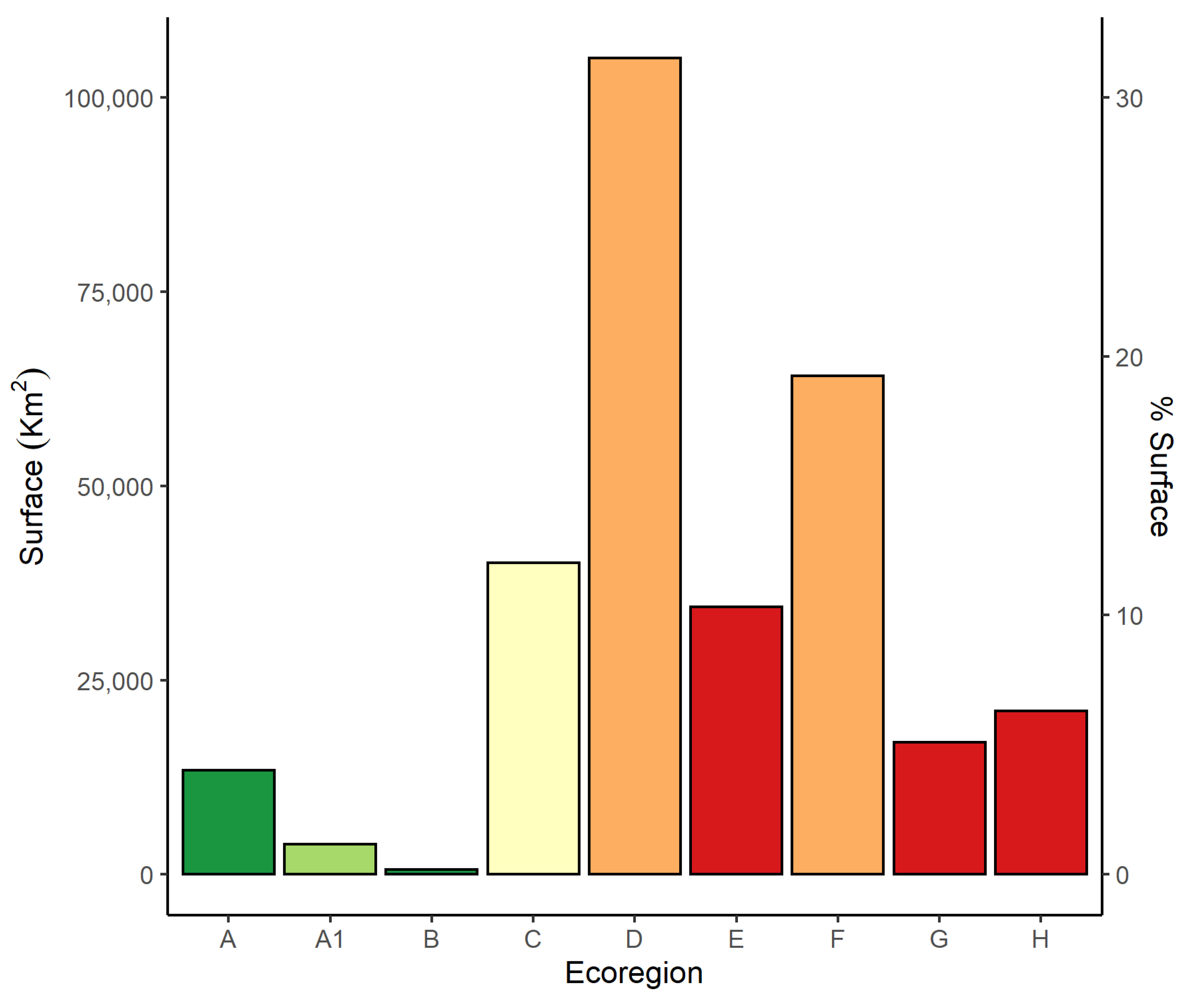
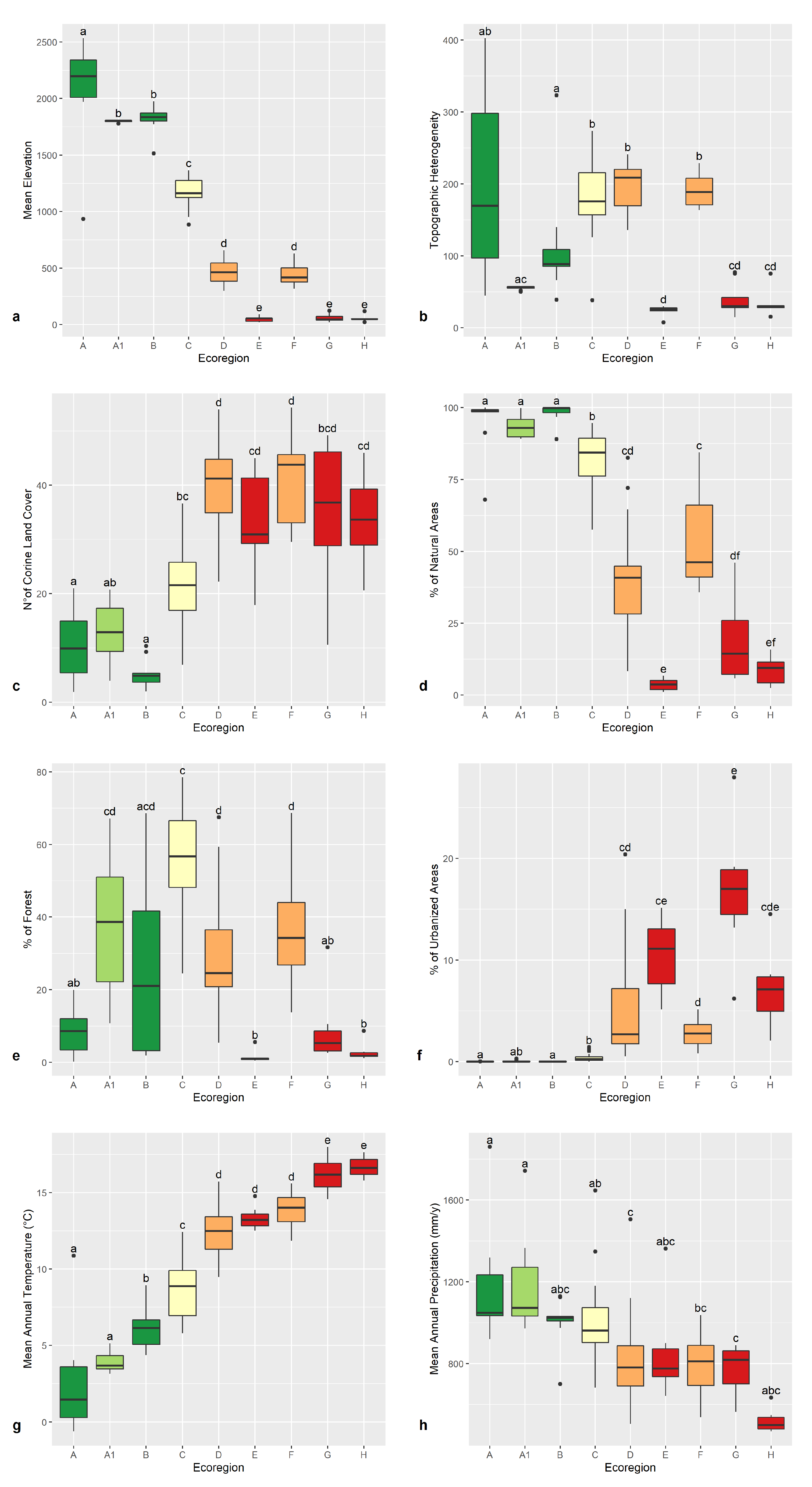
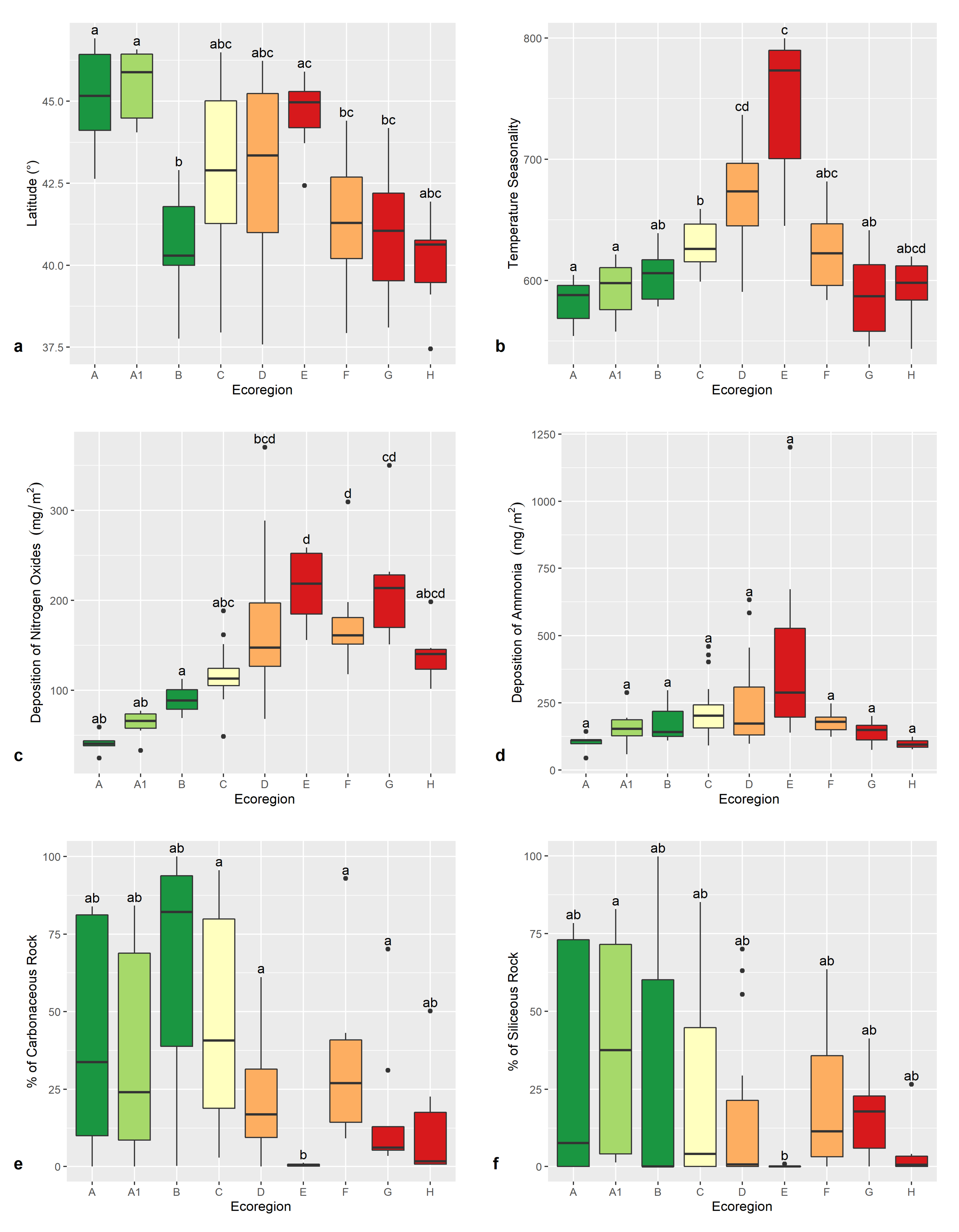
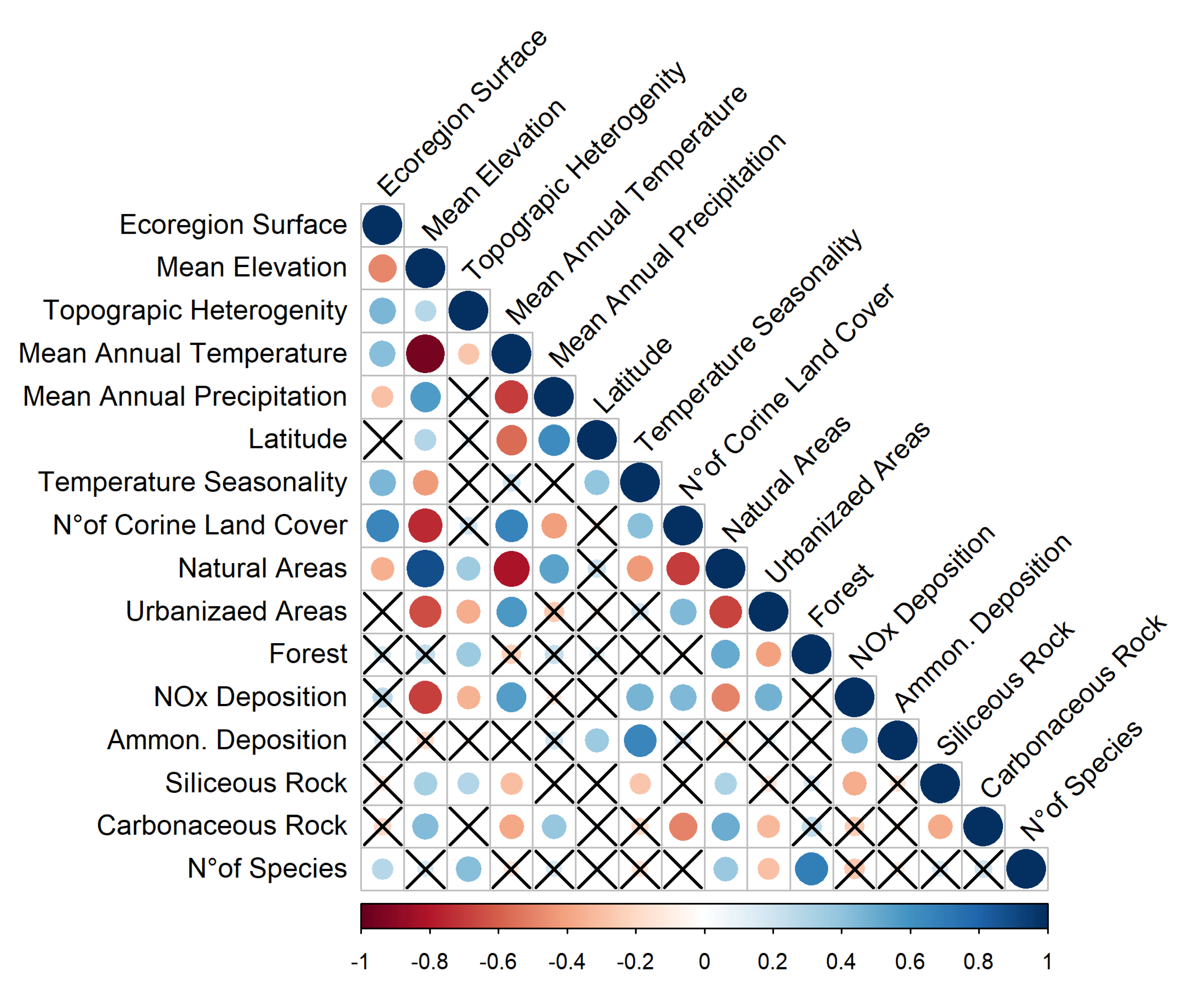
3.1. Environmental Features of the Ecoregions
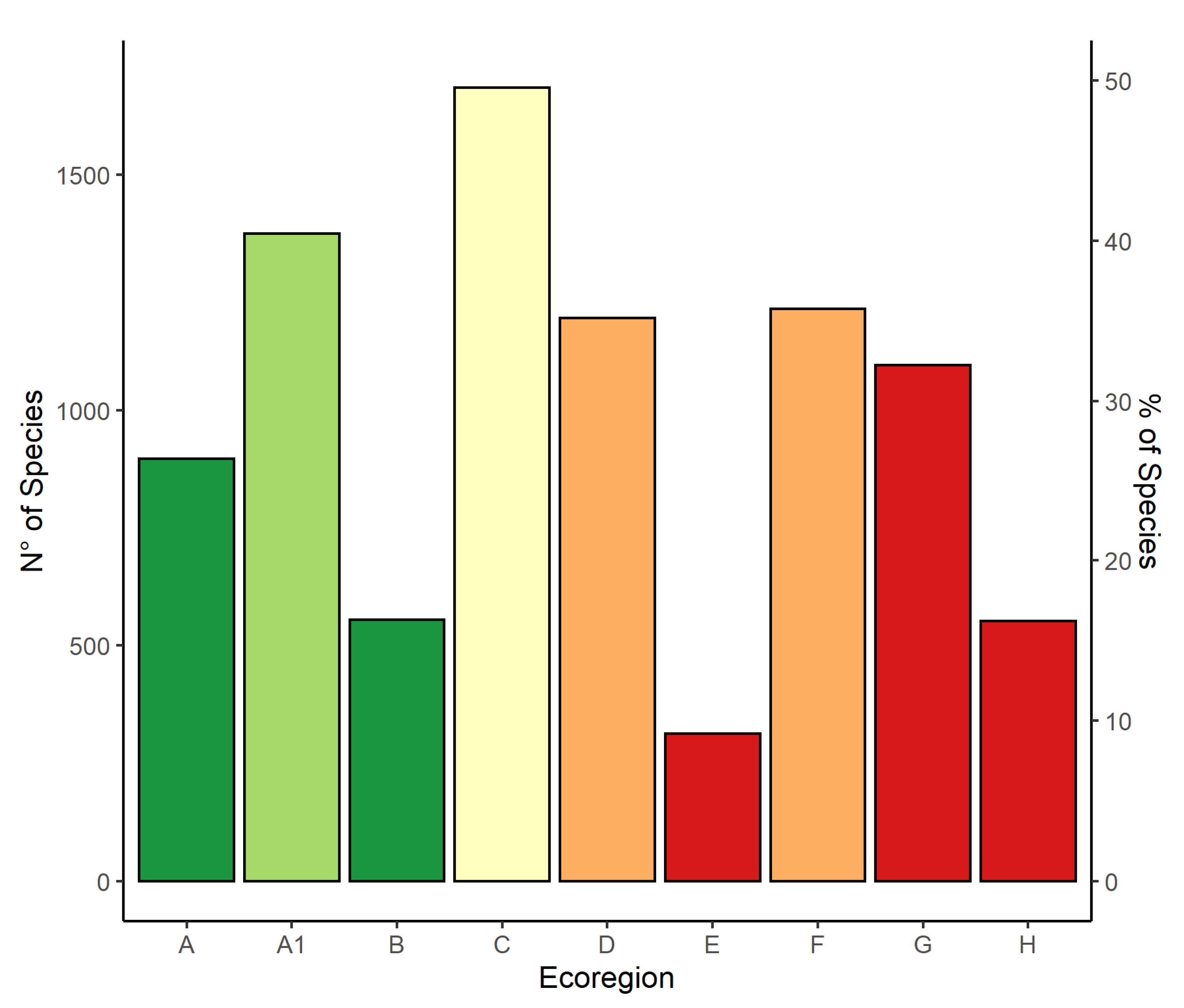
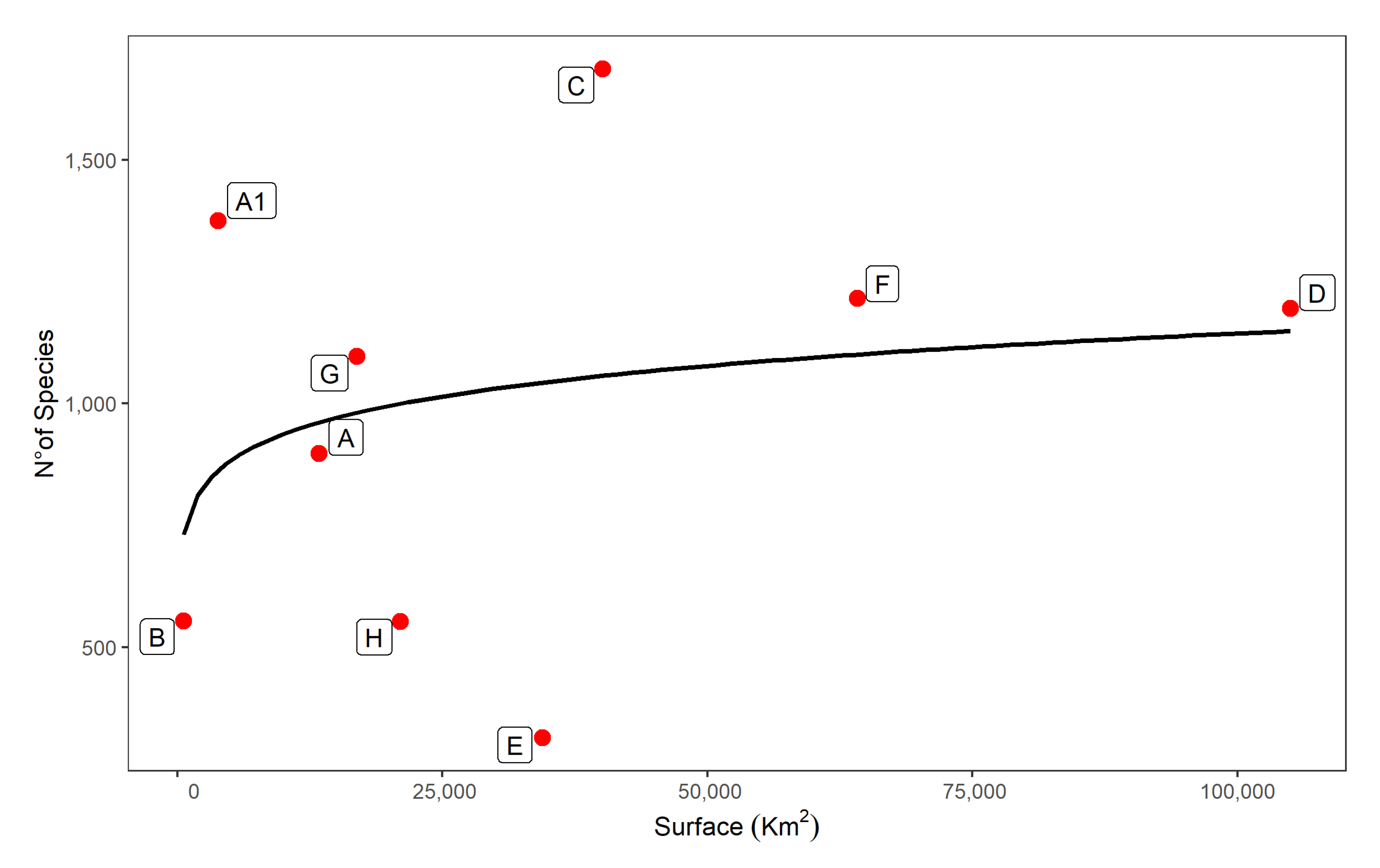
3.2. Lichen Richness in the Ecoregions
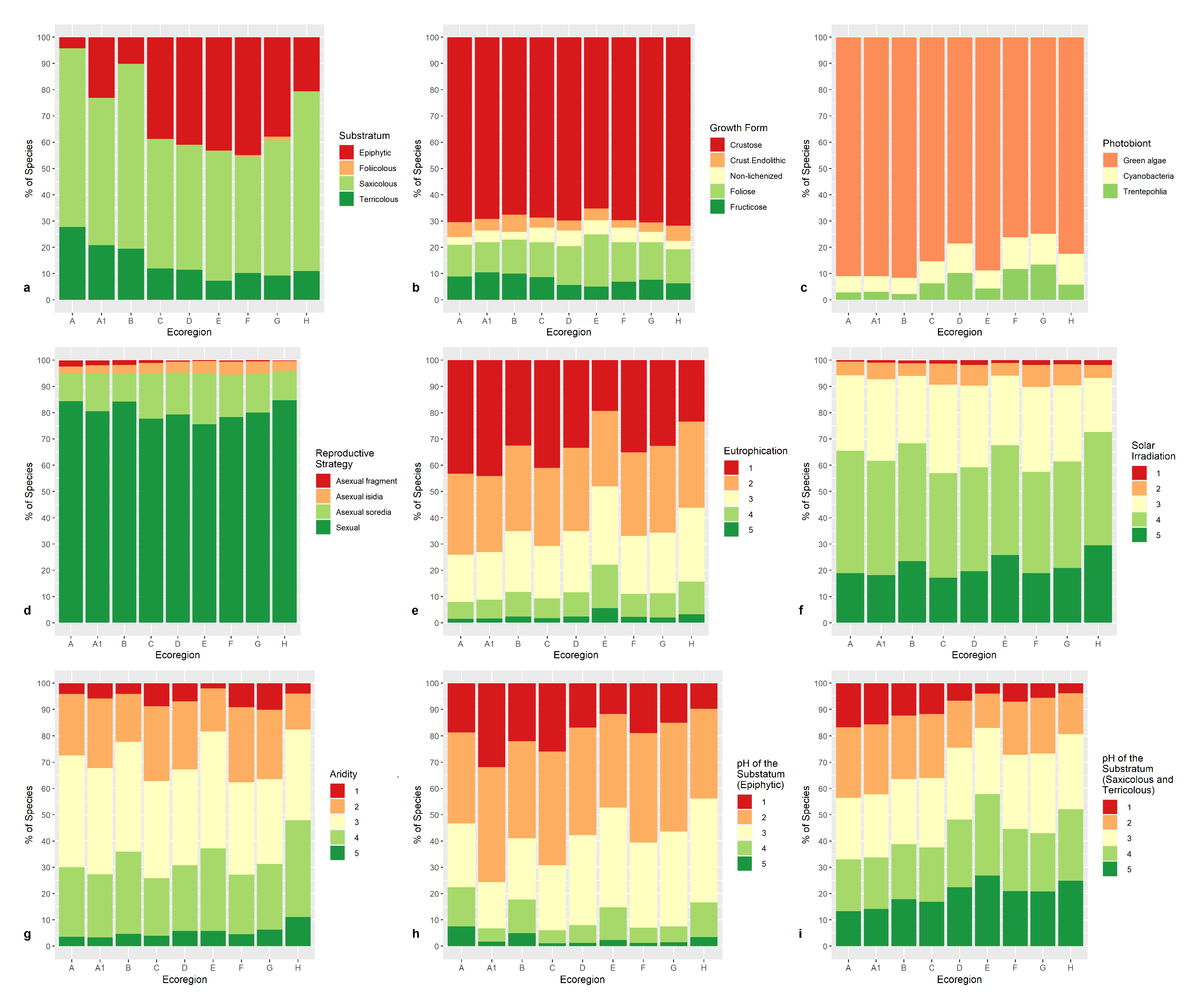
3.3. Lichen Traits and Ecological Requirements in the Ecoregions
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nascimbene, J.; Benesperi, R.; Brunialti, G.; Catalano, I.; Dalle Vedove, M.; Grillo, M.; Isocrono, D.; Matteucci, E.; Potenza, G.; Puntillo, D.; et al. Patterns and drivers of β-diversity and similarity of Lobaria pulmonaria communities in Italian forests. J. Ecol. 2013, 101, 493–505. [Google Scholar] [CrossRef]
- Nimis, P.L.; Tretiach, M. Delimiting Tyrrhenian Italy. A lichen foray in the SW part of the Peninsula. Bibl. Lichenol. 2004, 88, 465–478. [Google Scholar]
- Nimis, P.L. The Lichens of Italy: An Annotated Catalogue; Monografie XII; Museo Regionale Scienze Naturali di Torino: Torino, Italy, 1993; p. 897. [Google Scholar]
- Nimis, P.L.; Martellos, S. ITALIC, a database on Italian Lichens. Bibl. Lichenol. 2002, 82, 271–282. [Google Scholar]
- Nimis, P.L.; Hafellner, J.; Roux, C.; Clerc, P.; Mayrhofer, H.; Martellos, S.; Bilovitz, P.O. The lichens of the Alps—An annotated checklist. MycoKeys 2018, 31, 1–634. [Google Scholar] [CrossRef]
- Martellos, S. From a textual checklist to an information system: The case study of ITALIC, the Information System on Italian Lichens. Plant. Biosyst. 2012, 146, 764–770. [Google Scholar] [CrossRef]
- Nimis, P.L. The Lichens of Italy: A Second Annotated Catalogue; EUT: Trieste, Italy, 2016; p. 739. [Google Scholar]
- Nimis, P.L.; Martellos, S.; Chiarucci, A.; Ongaro, S.; Peplis, M.; Pittao, E.; Nascimbene, J. Exploring the relationships between ecology and species traits in cyanolichens: A case study in Italy. Fungal Ecol. 2020, 47, 100950. [Google Scholar] [CrossRef]
- Jørgensens, P.M. Distribution Patterns of Lichens in the Pacific Region. Aust. J. Bot. Suppl. Ser. 1983, 13, 43–66. [Google Scholar]
- Rubio-Salcedo, M.; Psomas, A.; Prieto, M.; Zimmermann, N.E.; Martìnez, I. Case study of the implications of climate change for lichen diversity and distributions. Biodivers. Conserv. 2017, 26, 1121–1141. [Google Scholar] [CrossRef]
- Kruskal, W.H.; Wallis, W.A. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Fotopoulos, S. Introduction to Modern Nonparametric Statistics. Technometrics 2004, 46, 488–489. [Google Scholar] [CrossRef]
- Arrhenius, O. Species and area. J. Ecol. 1921, 9, 95–99. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, B.; Simpson, G.; Solymos, P.; Stevens, H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.2-1, R Foundation for Statistical Computing: Vienna, Austria, 2015.
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Gleason, H.A. On the Relation between Species and Area. Ecology 1922, 3, 158–162. [Google Scholar] [CrossRef]
- Lawrey, J. The Species-Area Curve as an Index of Disturbance in Saxicolous Lichen Communities. Bryologist 1991, 94, 377–382. [Google Scholar] [CrossRef]
- Scheiner, S.; Chiarucci, A.; Fox, G.; Helmus, M.; McGlinn, D.; Willig, M. The underpinnings of the relationship of species richness with space and time. Ecol. Monogr. 2011, 81, 195–213. [Google Scholar] [CrossRef]
- Wilson, B.; Chiarucci, A. Do plant communities exist? Evidence from scaling-up local species-area relations to the regional level. J. Veg. Sci. 2009, 11, 773–775. [Google Scholar]
- Nascimbene, J.; Martellos, S.; Nimis, P.L. Epiphytic lichens of tree-line forests in the Central-Eastern Italian Alps and their importance for conservation. Lichenologist 2006, 38, 373–382. [Google Scholar] [CrossRef]
- Nascimbene, J.; Marini, L.; Motta, R.; Nimis, P.L. Lichen diversity of coarse woody habitats in a Pinus-Larix stand in the Italian Alps. Lichenologist 2008, 40, 153–163. [Google Scholar] [CrossRef]
- Nascimbene, J.; Thor, G.; Nimis, P.L. Habitat-types and lichen conservation in the Alps—Perspectives from a case study in the Stelvio National Park (Italy). Plant. Biosyst. 2012, 146, 428–442. [Google Scholar] [CrossRef]
- Chiarucci, C.; Nascimbene, J.; Campetella, G.; Chelli, S.; Dainese, M.; Giorgini, D.; Landi, S.; Lelli, C.; Canullo, R. Exploring patterns of beta-diversity to test the consistency of biogeographical boundaries: A case study across forest plant communities of Italy. Ecol. Evol. 2019, 9, 11716–11723. [Google Scholar] [CrossRef]
- Guttova, A.; Fackovcova, Z.; Martellos, S.; Paoli, L.; Munzi, S.; Pittao, E.; Ongaro, S. Ecological specialization of lichen congeners with a strong link to Mediterranean-type climate: A case study of the genus Solenopsora in the Apennine Peninsula. Lichenologist 2019, 51, 75–88. [Google Scholar] [CrossRef]
- Cislaghi, C.; Nimis, P.L. Lichens, air pollution and lung cancer. Nature 1997, 387, 463–464. [Google Scholar] [CrossRef] [PubMed]
- Giordani, P.; Calatayud, V.; Stofer, S.; Seidling, W.; Granke, O.; Fischer, R. Detecting the nitrogen critical loads on European forests by means of epiphytic lichens. A signal-to-noise evaluation. For. Ecol. Manag. 2014, 311, 29–40. [Google Scholar] [CrossRef]
- Contardo, T.; Giordani, P.; Paoli, L.; Vannini, A.; Loppi, S. May lichen biomonitoring of air pollution be used for environmental justice assessment? A case study from an area of N Italy with a municipal solid waste incinerator. Environ. Forensics 2018, 19, 265–276. [Google Scholar] [CrossRef]
- Nascimbene, J.; Marini, L. Oak forest exploitation and black locust invasion caused severe shifts in epiphytic lichen communities. Sci. Total Environ. 2010, 408, 5506–5512. [Google Scholar] [CrossRef]
- Nascimbene, J.; Lazzaro, L.; Benesperi, R. Patterns of beta-diversity and similarity reveal biotic homogenization of epiphytic lichen communities associated with the spread of Black locust forests. Fungal. Ecol. 2015, 14, 1–7. [Google Scholar] [CrossRef]
- van Herk, C.M.; Aptroot, A.; van Dobben, H.F. Long-term monitoring in the Netherlands suggests that lichens respond to global warming. Lichenologist 2002, 34, 141–154. [Google Scholar] [CrossRef]
- Aptroot, A.; van Herk, C.M. Further evidence of the effects of global warming on lichens, particularly those with Trentepohlia phycobionts. Environ. Pollut. 2007, 146, 293–298. [Google Scholar] [CrossRef]
- Marini, L.; Nascimbene, J.; Nimis, P.L. Large-scale patterns of epiphytic lichen species richness: Photobiont-dependent response to climate and forest structure. Sci. Total Environ. 2011, 409, 4381–4386. [Google Scholar] [CrossRef]
- Nascimbene, J.; Marini, L. Epiphytic lichen diversity along elevational gradients: Biological traits reveal a complex response to water and energy. J. Biogeogr. 2015, 42, 1222–1232. [Google Scholar] [CrossRef]
- Kappen, L. Plant activity under snow and ice, with particular reference to lichens. Arctic 1993, 46, 297–302. [Google Scholar] [CrossRef]
- Jahns, H.M. Felci, Muschi, Licheni d’Europa; Franco Muzzio Editore: Padova, Italy, 1992. [Google Scholar]
- Bässler, C.; Cadotte, M.W.; Beudert, B.; Heibl, C.; Blaschke, M.; Bradtka, J.H.; Langbehn, T.; Werth, S.; Müller, J. Contrasting patterns of lichen functional diversity and species richness across an elevation gradient. Ecography 2015, 38, 689–698. [Google Scholar] [CrossRef]
- Nash, T.H. Lichen Biology; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Grodzińska, K. Acidity of tree bark as a bioindicator of forest pollution in Southern Poland. Water Air Soil Pollut 1977, 8, 3–7. [Google Scholar] [CrossRef]
- Nimis, P.L.; Schiavon, L. The epiphytic lichen vegetation of the Tyrrhenian coasts in Central Italy. Ann. di Bot. 1986, 14, 39–67. [Google Scholar]
- Ravera, S.; Puglisi, M.; Vizzini, A.; Totti, C.; Arosio, G.; Benesperi, R.; Bianchi, E.; Boccardo, F.; Briozzo, I.; Dagnino, D.; et al. Notulae to the Italian flora of algae, bryophytes, fungi and lichens: 8. Ital. Bot. 2019, 8, 47–62. [Google Scholar] [CrossRef][Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OGU | Administrative Region | Ecoregion | Altitudinal Range |
|---|---|---|---|
| 1 | Abruzzo | E | 0–100 |
| 2 | Abruzzo | D | 101–900 |
| 3 | Abruzzo | A | 2001–4000 |
| 4 | Abruzzo | C | 901–2000 |
| 5 | Basilicata | H | 0–100 |
| 6 | Basilicata | G | 0–100 |
| 7 | Basilicata | D | 101–900 |
| 8 | Basilicata | F | 101–900 |
| 9 | Basilicata | B | 1701–4000 |
| 10 | Basilicata | C | 901–1700 |
| 11 | Calabria | H | 0–100 |
| 12 | Calabria | G | 0–100 |
| 13 | Calabria | D | 101–900 |
| 14 | Calabria | F | 101–900 |
| 15 | Calabria | B | 1701–4000 |
| 16 | Calabria | C | 901–1700 |
| 17 | Campania | G | 0–100 |
| 18 | Campania | D | 101–900 |
| 19 | Campania | F | 101–900 |
| 20 | Campania | B | 1701–4000 |
| 21 | Campania | C | 901–1700 |
| 22 | Emilia Romagna | E | 0–100 |
| 23 | Emilia Romagna | D | 101–900 |
| 24 | Emilia Romagna | A1 | 1701–1900 |
| 25 | Emilia Romagna | A | 1901–4000 |
| 26 | Emilia Romagna | C | 901–1700 |
| 27 | Friuli Venezia Giulia | E | 0–100 |
| 28 | Friuli Venezia Giulia | D | 101–900 |
| 29 | Friuli Venezia Giulia | A1 | 1701–1900 |
| 30 | Friuli Venezia Giulia | A | 1901–4000 |
| 31 | Friuli Venezia Giulia | C | 901–1700 |
| 32 | Lazio | G | 0–100 |
| 33 | Lazio | B | 1701–4000 |
| 34 | Lazio | F | 101–900 |
| 35 | Lazio | C | 901–1700 |
| 36 | Liguria | G | 0–250 |
| 37 | Liguria | A1 | 1701–1900 |
| 38 | Liguria | A | 1901–4000 |
| 39 | Liguria | D | 251–900 |
| 40 | Liguria | F | 251–900 |
| 41 | Liguria | C | 901–1700 |
| 42 | Lombardia | E | 0–100 |
| 43 | Lombardia | D | 101–900 |
| 44 | Lombardia | A1 | 1701–1900 |
| 45 | Lombardia | A | 1901–4000 |
| 46 | Lombardia | C | 901–1700 |
| 47 | Marche | E | 0–100 |
| 48 | Marche | D | 101–900 |
| 49 | Marche | B | 1701–4000 |
| 50 | Marche | C | 901–1700 |
| 51 | Molise | H | 0–100 |
| 52 | Molise | D | 101–900 |
| 53 | Molise | B | 1701–4000 |
| 54 | Molise | C | 901–1700 |
| 55 | Piemonte | E | 0–100 |
| 56 | Piemonte | D | 101–900 |
| 57 | Piemonte | A1 | 1701–1900 |
| 58 | Piemonte | A | 1901–4000 |
| 59 | Piemonte | C | 901–1700 |
| 60 | Puglia | H | 0–100 |
| 61 | Puglia | D | 101–900 |
| 62 | Puglia | F | 101–900 |
| 63 | Puglia | C | 901–1700 |
| 64 | Sardegna | H | 0–50 |
| 65 | Sardegna | G | 0–50 |
| 66 | Sardegna | B | 1401–4000 |
| 67 | Sardegna | D | 51–700 |
| 68 | Sardegna | F | 51–700 |
| 69 | Sardegna | C | 701–1400 |
| 70 | Sicilia | H | 0–250 |
| 71 | Sicilia | G | 0–250 |
| 72 | Sicilia | B | 1601–4000 |
| 73 | Sicilia | F | 251–900 |
| 74 | Sicilia | D | 251–900 |
| 75 | Sicilia | C | 901–1600 |
| 76 | Toscana | G | 0–100 |
| 77 | Toscana | D | 101–900 |
| 78 | Toscana | F | 101–900 |
| 79 | Toscana | A | 1701–4000 |
| 80 | Toscana | C | 901–1700 |
| 81 | Trentino Alto Adige | D | 101–900 |
| 82 | Trentino Alto Adige | A1 | 1701–1900 |
| 83 | Trentino Alto Adige | A | 1901–4000 |
| 84 | Trentino Alto Adige | C | 901–1700 |
| 85 | Umbria | D | 101–900 |
| 86 | Umbria | F | 101–900 |
| 87 | Umbria | B | 1701–4000 |
| 88 | Umbria | C | 901–1700 |
| 89 | Val d’Aosta | D | 101–900 |
| 90 | Val d’Aosta | A1 | 1701–1900 |
| 91 | Val d’Aosta | A | 1901–4000 |
| 92 | Val d’Aosta | C | 901–1700 |
| 93 | Veneto | E | 0–100 |
| 94 | Veneto | D | 101–900 |
| 95 | Veneto | A1 | 1701–1900 |
| 96 | Veneto | A | 1901–4000 |
| 97 | Veneto | C | 901–1700 |
| Administrative Region | % A | % A1 | % B | % C | % D | % E | % F | % G | % H | % Italy Surface | n° of Species | % of Species |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Piemonte | 12.6 | 3.64 | 16.65 | 65.22 | 1.85 | 1.48 | 1293 | 46.51 | ||||
| Valle D’Aosta | 63.5 | 8.44 | 20.97 | 6.85 | 0 | 8.55 | 945 | 33.99 | ||||
| Lombardia | 10.38 | 2.68 | 12.75 | 40.7 | 32.46 | 7.61 | 1295 | 46.58 | ||||
| Veneto | 3.87 | 2.55 | 13.97 | 20.87 | 57.89 | 7.98 | 1175 | 42.27 | ||||
| Trentino A.A. | 33.66 | 10.04 | 38.06 | 17.3 | 5.04 | 1579 | 56.8 | |||||
| Liguria | 0.07 | 0.25 | 14.18 | 22.09 | 38.58 | 23.96 | 7.43 | 1089 | 39.17 | |||
| Friuli V.G. | 1.6 | 2.33 | 22.56 | 39.91 | 32.8 | 1.79 | 1049 | 37.73 | ||||
| Emilia-Romagna | 0.02 | 0.14 | 7.81 | 42.68 | 49.18 | 4.51 | 775 | 27.88 | ||||
| Toscana | 0.21 | 6.22 | 2.82 | 68.35 | 21.63 | 4.52 | 1205 | 43.35 | ||||
| Umbria | 0.16 | 9.51 | 21.58 | 67.85 | 3.34 | 562 | 20.22 | |||||
| Marche | 0.69 | 8.02 | 74.79 | 16.15 | 2.8 | 673 | 24.21 | |||||
| Molise | 0.32 | 13.8 | 78.7 | 7.03 | 3.11 | 520 | 18.71 | |||||
| Lazio | 0.87 | 10.71 | 62.47 | 25.27 | 2.6 | 878 | 31.58 | |||||
| Abruzzo | 1.8 | 38.69 | 52.41 | 7.39 | 8.41 | 705 | 25.36 | |||||
| Puglia | 0.3 | 50.36 | 7 | 42.28 | 5.7 | 629 | 22.63 | |||||
| Campania | 0.05 | 9.36 | 28.08 | 40.23 | 21.3 | 6.09 | 844 | 30.36 | ||||
| Calabria | 0.37 | 20.34 | 32.59 | 28.65 | 4.66 | 13.13 | 7.9 | 975 | 35.07 | |||
| Basilicata | 0.25 | 16.64 | 71.74 | 3.67 | 0.05 | 7.2 | 1.08 | 639 | 22.99 | |||
| Sicilia | 0.75 | 7.25 | 33.96 | 21.68 | 2.79 | 31.53 | 6.47 | 1168 | 42.01 | |||
| Sardegna | 0.17 | 10.72 | 21.46 | 52.61 | 8.32 | 5.76 | 3.59 | 1252 | 45.04 |
| Oceanic | Suboceanic | Subcontinental | |
|---|---|---|---|
| Alpine | 0 | 23 (2.6%) | 26 (2.9%) |
| Subalpine | 1 (0.1%) | 69 (5.0%) | 46 (3.4%) |
| Oromediterranean | 0 | 15 (2.7%) | 21 (3.8%) |
| Montane | 19 (1.1%) | 262 (15.5%) | 72 (4.3%) |
| Humid Submediterranean | 22 (1.8%) | 317 (26.0%) | 49 (3.3%) |
| Dry Submediterranean | 9 (0.7%) | 239 (19.9%) | 65 (5.4%) |
| Humid Mediterranean | 40 (3.6%) | 313 (28.4%) | 33 (3.0%) |
| Dry Mediterranean | 0 | 58 (10.4%) | 37 (6.6%) |
| Padane | 0 | 22 (7.0%) | 4 (1.3%) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martellos, S.; d’Agostino, M.; Chiarucci, A.; Nimis, P.L.; Nascimbene, J. Lichen Distribution Patterns in the Ecoregions of Italy. Diversity 2020, 12, 294. https://doi.org/10.3390/d12080294
Martellos S, d’Agostino M, Chiarucci A, Nimis PL, Nascimbene J. Lichen Distribution Patterns in the Ecoregions of Italy. Diversity. 2020; 12(8):294. https://doi.org/10.3390/d12080294
Chicago/Turabian StyleMartellos, Stefano, Marco d’Agostino, Alessandro Chiarucci, Pier Luigi Nimis, and Juri Nascimbene. 2020. "Lichen Distribution Patterns in the Ecoregions of Italy" Diversity 12, no. 8: 294. https://doi.org/10.3390/d12080294
APA StyleMartellos, S., d’Agostino, M., Chiarucci, A., Nimis, P. L., & Nascimbene, J. (2020). Lichen Distribution Patterns in the Ecoregions of Italy. Diversity, 12(8), 294. https://doi.org/10.3390/d12080294