Patterns of Distribution of Bivalve Populations in a Mediterranean Temporary River

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Environmental Conditions
2.2. Qualitative Sampling
2.3. Quantitative Sampling
2.4. Data Analysis
3. Results
3.1. Sites Description
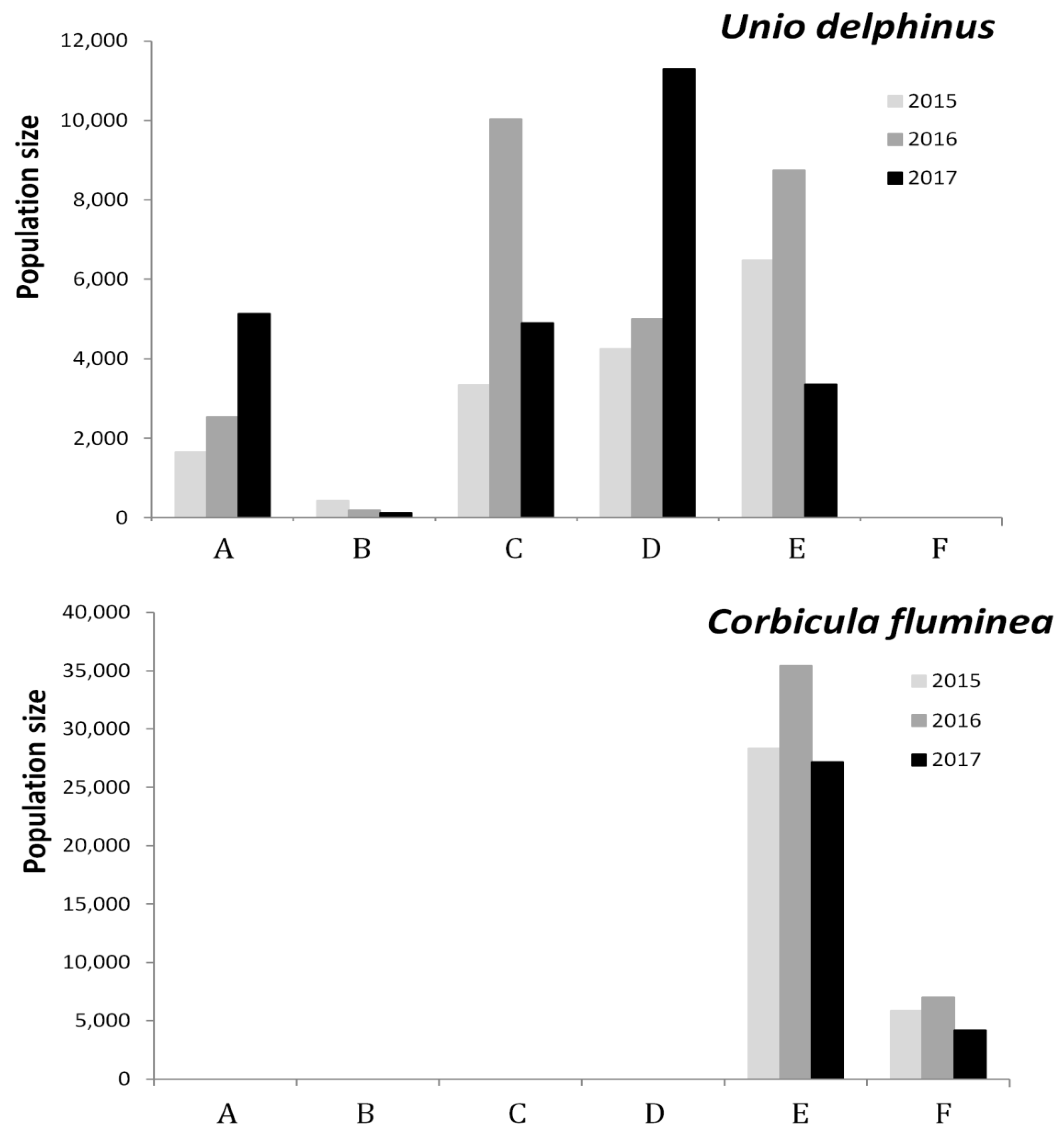
3.2. Bivalve Abundance and Distribution
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Species | Sediment Type | |||||
|---|---|---|---|---|---|---|---|
| Boulders | Cobblers | Coarse Gravel | Fine Gravel | Sand | Silt/Clay | ||
| 2015 | U. delphinus | 8 | 15 | 42 | 25 | 1 | 8 |
| C. fluminea | 4 | 16 | 41 | 8 | 0 | 31 | |
| 2016 | U. delphinus | 29 | 16 | 24 | 10 | 4 | 18 |
| C. fluminea | 13 | 22 | 30 | 22 | 8 | 5 | |
| 2017 | U. delphinus | 23 | 11 | 11 | 43 | 0 | 12 |
| C. fluminea | 19 | 13 | 10 | 44 | 0 | 13 | |
| % of occurrence | |||||||
| 2015 | 29 | 14 | 7 | 0 | 0 | 10 | |
| 2016 | 13 | 10 | 8 | 7 | 1 | 10 | |
| 2017 | 14 | 10 | 8 | 12 | 0 | 6 | |
References
- Vaughn, C.C.; Hakenkamp, C.C. The functional role of burrowing bivalves in freshwater ecosystems. Freshw. Biol. 2001, 46, 1431–1446. [Google Scholar] [CrossRef]
- Sala, O.E.; Chapin, F.S.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- Bogan, A.E. Global diversity of freshwater mussels (Mollusca, Bivalvia) in freshwater. Hydrobiol 2008, 595, 139–147. [Google Scholar] [CrossRef]
- Osterling, M.E.; Arvidsson, B.L.; Greenberg, L.A. Habitat degradation and the decline of the threatened mussel Margaritifera margaritifera: Influence of turbidity and sedimentation on the mussel and its host. J. Appl. Ecol. 2010, 47, 759–768. [Google Scholar] [CrossRef]
- Lopes-Lima, M.; Burlakova, L.E.; Karatayev, A.; Mehler, K.; Seddon, M.; Sousa, R. Conservation of freshwater bivalves at the global scale: Diversity, threats and research needs. Hydrobiologia 2018, 810, 1–14. [Google Scholar] [CrossRef]
- Lydeard, C.; Cowie, R.H.; Ponder, W.F.; Bogan, A.E.; Bouchet, P.; Clark, S.A.; Cummings, K.S.; Frest, T.J.; Gargominy, O.; Herbert, D.G.; et al. The global decline of nonmarine mollusks. BioScience 2004, 54, 321–330. [Google Scholar] [CrossRef]
- Aldridge, D.C.; Elliot, P.; Moggridge, G.D. The recent and rapid spread of the zebra mussel (Dreissena polymorpha) in Great Britain. Biol. Conserv. 2004, 119, 253–261. [Google Scholar] [CrossRef]
- Strayer, D.L.; Malcom, H.M. Effects of zebra mussels (Dreissena polymorpha) on native bivalves: The beginning of the end or the end of the beginning? J. N. Am. Benthol. Soc. 2007, 26, 111–122. [Google Scholar] [CrossRef]
- Sousa, R.; Antunes, C.; Guilhermino, L. Ecology of the invasive Asian clam Corbicula fluminea (Müller, 1774) in aquatic ecosystems: An overview. Int. J. Lim. 2008, 44, 85–94. [Google Scholar] [CrossRef]
- Sousa, R.; Nogueira, A.J.A.; Gaspar, M.B.; Antunes, C.; Guilhermino, L. Growth and extremely high production of the nonindigenous invasive species Corbicula fluminea (Müller, 1774): Possible implications for ecosystem functioning. Estuar. Coast. Shelf Sci. 2008, 80, 289–295. [Google Scholar] [CrossRef]
- Sousa, R.; Gutiérrez, J.L.; Aldridge, D.C. Non-indigenous invasive bivalves as ecosystem engineers. Biol. Invasions 2009, 11, 2367–2385. [Google Scholar] [CrossRef]
- Sousa, R.; Varandas, S.; Cortes, R.; Teixeira, A.; Lopes-Lima, M.; Machado, J.; Guilhermino, L. Massive die-offs of freshwater bivalves as resource pulses. Ann. Limnol.-Int. J. Lim. 2012, 48, 105–112. [Google Scholar] [CrossRef]
- Ilarri, M.; Sousa, R. Corbicula fluminea Müller (Asian clam). In A Handbook of Global Freshwater Invasive Species; Francis, R.A., Ed.; Routledge: London, UK, 2012; Chapter 15; p. 484. [Google Scholar]
- Anastácio, P.M.; Ribeiro, F.; Capinha, C.; Banha, F.; Gama, M.; Filipe, A.F.; Rebelo, R.; Sousa, R. Non-native freshwater fauna in Portugal: A review. Sci. Total Environ. 2019, 650, 1923–1934. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Summary for policymakers. In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 1–32. [Google Scholar]
- Araujo, R.; Toledo, C.; Machordom, A. Redescription of Unio gibbus Spengler, 1793, a west palaearctic freshwater mussel with hookless glochidia. Malacologia 2009, 51, 131–141. [Google Scholar] [CrossRef]
- Reis, J.; Machordom, A.; Araujo, R. Morphological and molecular diversity of Unionidae (Mollusca, Bivalvia) from Portugal. Graellsia 2013, 69, 17–36. [Google Scholar]
- Lopes-Lima, M.; Sousa, R.; Geist, J.; Aldridge, D.C.; Araujo, R.; Bergengren, J.; Bespalaya, Y.; Bódis, E.; Burlakova, L.; Van Damme, D.; et al. Conservation status of freshwater mussels in Europe: State of the art and future challenges. Biol. Rev. 2017, 92, 572–607. [Google Scholar] [CrossRef] [PubMed]
- Novais, A.; Dias, E.; Sousa, R. Inter- and intraspecific variation of carbon and nitrogen stable isotope ratios in freshwater bivalves. Hydrobiologia 2016, 765, 149–158. [Google Scholar] [CrossRef]
- Modesto, V.; Castro, P.; Lopes-Lima, M.; Antunes, C.; Ilarri, M.; Sousa, R. Potential impacts of the invasive species Corbicula fluminea on the survival of glochidia. Sci. Total Environ. 2019, 673, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, C.C.; Spooner, D.E. Scale-dependent associations between native freshwater mussels and invasive Corbicula. Hydrobiologia 2006, 568, 331–339. [Google Scholar] [CrossRef]
- Rajagopal, S.; Pollux, B.J.A.; Peters, J.L.; Cremers, G.; Moon-van der Staay, S.Y.; van Alen, T.; Eygensteyn, J.; van Hoek, A.; Palau, A.; bij de Vaate, A.; et al. Origin of Spanish invasion by the zebra mussel, Dreissena polymorpha (Pallas, 1771) revealed by amplified fragment length polymorphism (AFLP) fingerprinting. Biol. Invasions 2009, 11, 2147–2159. [Google Scholar] [CrossRef]
- McRae, S.E.; Allan, J.D.; Burch, J.B. Reach and catchment scale determinants of the distribution of freshwater mussels (Bivalvia: Unionidae) in south-eastern Michigan, USA. Freshwat. Biol. 2004, 49, 127–142. [Google Scholar] [CrossRef]
- Obermeyer, B.K. Comparison of Quadrats Versus Timed Snorkel Searches for Assessing Freshwater Mussels. Am. Midl. Nat. 1998, 155, 307–320. [Google Scholar] [CrossRef]
- Schloesser, D.W.; Metcalfe-Smith, J.L.; Kovalak, W.P.; Longton, G.D.; Smithee, R.D. Extirpation of Freshwater Mussels (Bivalvia: Unionidae) Following the Invasion of Dreissenid Mussels in an Interconnecting River of the Laurentian Great Lakes. Am. Midl. Nat. 2006, 155, 307–320.42. [Google Scholar] [CrossRef]
- ASTM D2487—1966 Standard Practice for Classification of Soils for Engineering Purposes (Unified Soil Classification System); Reapproved; ASTM International: West Conshohocken, PA, USA, 2006.
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice-Hall, Inc.: Upper Saddle River, NJ, USA, 1999; p. 663. [Google Scholar]
- Anastácio, P.M.; Banha, F.; Capinha, C.; Bernardo, J.M.; Costa, A.M.; Teixeira, A.; Bruxelas, S. Indicators of movement and space use for two co-occurring invasive crayfish species. Ecol. Indic 2015, 53, 171–181. [Google Scholar] [CrossRef]
- Loughman, Z.J.; Skalican, K.T.; Taylor, N.D. Habitat selection and movement of Cambarus chasmodactylus (Decapoda: Cambaridae) assessed via radio telemetry. Freshwat. Sci. 2013, 32, 1288–1297. [Google Scholar] [CrossRef]
- Smith, D.G. Sistematics and distribution of the recent Margaritiferidae. In Ecology and Evolution of the Freshwater Mussels Unionoida; Springer: Berlin/Heidelberg, Germany, 2001; pp. 33–49. [Google Scholar]
- Araujo, R.; Unio tumidiformis. The IUCN Red List of Threatened Species. 2011. Available online: http://dx.doi.org/10.2305/IUCN.UK.20112.RLTS.T171935A6810869.en (accessed on 10 November 2011).
- Rovira, Q.; Campos, M.; Feo-Quer, C.; Camós, I.; Martí, J.; Angelats, I.; Cros, C.; Casadevall, R.; Dalmau, G.; Cruset, E.; et al. Habitat preferences of Unio mancus and U. ravoisieri in northeast of Catalonia. In Proceedings of the 2nd International Seminar on the Rearing of Unionoid Mussels, Clervaux, Luxembourg, 24 November–27 November 2015. [Google Scholar]
- Patrício, C.I.M. Contribuição para o estudo da bioecologia dos mexilhões de água doce (Unionoida) do Nordeste de Portugal. Master Thesis, University of Oporto, Oporto, Portugal, 2013. [Google Scholar]
- Beran, L. Spreading expansion of Corbicula fluminea (Mollusca: Bivalvia) in the Czech Republic. Heldia 2006, 6, 187–192. [Google Scholar]
- Vohmann, A.; Borcherding, J.; Kureck, A.; bij de Vaate, A.; Arndt, A.; Weitere, M. Strong body mass decrease of the invasive clam Corbicula fluminea during summer. Biol. Invasions 2010, 12, 53–64. [Google Scholar] [CrossRef]
- Way, C.; Hornbach, D.; Miller-Way, C.; Payne, B.; Miller, A.C. Dynamics of filter feeding in Corbicula fluminea (Bivalvia: Corbiculidae). Can. J. Zool. 2011, 68, 115–120. [Google Scholar] [CrossRef]
- Karatayev, A.Y.; Burlakova, L.E.; Kesterson, T.; Padilla, D.K. Dominance of the Asiatic clam, Corbicula fluminea (Müller) in the benthic community of a reservoir. J. Shellfish Res. 2003, 22, 487–493. [Google Scholar]
- Ferreira-Rodriguez, N.; Pardo, I.; Sousa, R. Negative effects of Corbicula fluminea over native freshwater mussels. Hydrobiology 2018, 810, 85–95. [Google Scholar] [CrossRef]
- Graczyk, T.K.; Conn, D.B.; Marcogliese, D.J.; Graczyk, H.; de Lafontaine, Y. Accumulation of human waterborne parasites by zebra mussels (Dreissena polymorpha) and Asian freshwater clams (Corbicula fluminea). Parasitol. Res. 2003, 89, 107–112. [Google Scholar] [PubMed]
- Cherry, D.S.; Scheller, J.L.; Cooper, N.L.; Bidwell, J.R. Potential effects of Asian clam (Corbicula fluminea) die-offs on native freshwater mussels (Unionidae) I: Water-column ammonia levels and ammonia toxicity. J. N. Am. Benthol. Soc. 2005, 24, 369–380. [Google Scholar] [CrossRef]
- Taeubert, A.; Martinez, J.E.; Gum, M.P.B.; Geist, J. The relationship between endangered thick-shelled river mussel (Unio crassus) and its host fishes. Biol. Conserv. 2012, 155, 94–103. [Google Scholar] [CrossRef]
- Correia, A.M.; Bandeira, N.; Anastácio, P.M. Predator-prey interactions of Procambarus clarkii with aquatic macroinvertebrates in single and multiple prey systems. Acta Oecologica 2005, 28, 337–343. [Google Scholar] [CrossRef]
- Reis, J.; Araújo, R. Life history of the freshwater mussel Unio tumidiformis (Bivalvia: Unionidae) in a temporary Mediterranean-type stream. Invert. Biol. 2016, 135, 31–45. [Google Scholar] [CrossRef]
- Weiss, S.; Marić, S.; Snoj, A. Regional structure despite limited mtDNA sequence diversity found in the endangered Huchen, Hucho hucho (Linneaus, 1758). Hydrobiologia 2010, 658, 103–110. [Google Scholar] [CrossRef]
- Meira, A.; Lopes-Lima, M.; Varandas, S.; Teixeira, A.; Arenas, F.; Sousa, R. Invasive crayfishes as a threat to freshwater bivalves: Interspecific differences and conservation implications. Sci. Total Environ. 2018, 649, 938–948. [Google Scholar] [CrossRef]





| Environmental Parameters- | Pools | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | C | D | E | F | |||||||||||||
| Maximum depth range (cm) | 72–80 | 75–90 | 90–160 | 200 | 130–160 | 20–20 | ||||||||||||
| Maximum width range (m) | 17–30 | 7–10 | 21–26 | 23–24 | 16–23 | 12–16 | ||||||||||||
| Maximum length range (m) | 84–102 | 28–32 | 226–268 | 100–113 | 159–236 | 37–88 | ||||||||||||
| Total area (m2) | 3015 | 2411 | 16910 | 4065 | 489393 | 6400 | ||||||||||||
| Exotic vegetation | No | No | Yes | Yes | Yes | Yes | ||||||||||||
| Riparian gallery vegetation cover | Continuous gallery composed of initial succession vegetation, reeds, presence of few trees—Fraxinus angustifolia or shrubs—Nerium oleander. | Gallery almost continuous composed mainly of shrubs—Flueggea tinctoria and N. oleander. | Gallery almost continuous in the left bank with F. angustifolia and Salix spp. and the exotic (Giant cane, Arundo donax). Discontinuous gallery of F. angustifolia and N. oleander in the right bank. | Almost continuous gallery of F. angustifolia and N. oleander in right bank, but very thin transversely. Left bank with sparse vegetation with few individuals of F. angustifolia and N. oleander. Presence of Eucalyptus globulus. | Sparse gallery in both banks, N. oleander in the right rocky bank, F. angustifolia Salix spp. and E. globulus in the left bank. | Continuous gallery composed by initial succession vegetation, reeds, presence of few trees of F. angustifolia Salix spp. and E. globulus. | ||||||||||||
| Cattle presence | Yes | Yes | Yes | Yes | Yes | Yes | ||||||||||||
| Sampling year | 2015 | 2016 | 2017 | 2015 | 2016 | 2017 | 2015 | 2016 | 2017 | 2015 | 2016 | 2017 | 2015 | 2016 | 2017 | 2015 | 2016 | 2017 |
| Submerged area (m2) | 991 | 1172 | 1232 | 136 | 240 | 177 | 3903 | 5188 | 3659 | 1593 | 2158 | 1882 | 2859 | 3641 | 2135 | 488 | 1115 | 368 |
| Site | Species | 2015 | 2016 | 2017 | |||
|---|---|---|---|---|---|---|---|
| Individuals per m2 | C.P.U.E. | Individuals per m2 | C.P.U.E. | Individuals per m2 | C.P.U.E. | ||
| A | U. delphinus | 1.67 | 3.0 | 2.17 | 5.2 | 4.17 | 50.5 |
| C. fluminea | 0 | 0 | 0 | 0 | 0 | 0 | |
| B | U. delphinus | 3.20 | 2.5 | 0.80 | 7.5 | 0.73 | 0 |
| C. fluminea | 0 | 0 | 0 | 0 | 0 | 0 | |
| C | U. delphinus | 0.86 | 18.7 | 1.94 | 4.0 | 1.38 | 5.7 |
| C. fluminea | 0 | 0 | 0 | 0 | 0 | 0 | |
| D | U. delphinus | 2.67 | 6.3 | 2.32 | 15.0 | 6.00 | 0.9 |
| C. fluminea | 0 | 0 | 0 | 0 | 0 | 0 | |
| E | U. delphinus | 2.27 | 42.3 | 2.40 | 36.0 | 1.57 | 0 |
| C. fluminea | 9.93 | 51.3 | 9.73 | 11.0 | 12.71 | 0 | |
| F | U. delphinus | 0 | 20.0 | 0 | 0 | 0 | 4.3 |
| C. fluminea | 12.00 | 2.0 | 6.29 | 20.0 | 11.38 | 10.7 | |
| Statistical Parameters | Dimension 1 | Dimension 2 | ||
|---|---|---|---|---|
| Eigenvalues | 0.2 | 0.21 | ||
| % of variance | 2.01 | 1.90 | ||
| Cumulative % of variance | 2.01 | 3.91 | ||
| Variable | r2 | p value | r2 | p value |
| Distance to near margin (m) | 0.54 | 3.13 × 10−47 | 0.54 | 1.83 × 10−46 |
| C. fluminea | 0.17 | 3.18 × 10−22 | 0.09 | 2.31 × 10−12 |
| Silt/Clay | - | - | 0.42 | 7.94 × 10−63 |
| Riparian gallery cover | - | - | 0.16 | 5.86 × 10−22 |
| Water column depth (m) | 0.65 | 7.87 × 10−79 | 0.56 | 3.32 × 10−56 |
| Coarse Gravel | 0.09 | 2.94 × 10−12 | 0.17 | 2.12 × 10−23 |
| Fine Gravel | 0.18 | 1.37 × 10−24 | 0.05 | 1.00 × 10−06 |
| Boulders | 0.55 | 8.17 × 10−93 | 0.02 | 6.65 × 10−04 |
| Sand | - | - | 0.08 | 3.73 × 10−11 |
| Cobblers | 0.15 | 3.92 × 10−20 | 0.15 | 1.76 × 10−20 |
| U. delphinus | 0.07 | 1.29 × 10−09 | 0.05 | 1.00 × 10−06 |
| Species | Size Class | Year | ||
|---|---|---|---|---|
| 2015 | 2016 | 2017 | ||
| U. delphinus | >30 mm | a | b | b |
| 30–40 mm | a | a | b | |
| 40–50 mm | a | b | c | |
| 50–60 mm | a | a | b | |
| 60–70 mm | a | a | b | |
| <70 mm | a | b | b | |
| C. fluminea | >10 mm | a | b | b |
| 10–20 mm | a | a | b | |
| 20–30 mm | a | b | a | |
| 30–40 mm | a | a | b | |
| 40–50 mm | a | a | b | |
| <50 mm | a | a | b | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gama, M.; Banha, F.; Moreira, C.; Gama, H.; Graça, M.; Anastácio, P. Patterns of Distribution of Bivalve Populations in a Mediterranean Temporary River. Diversity 2020, 12, 158. https://doi.org/10.3390/d12040158
Gama M, Banha F, Moreira C, Gama H, Graça M, Anastácio P. Patterns of Distribution of Bivalve Populations in a Mediterranean Temporary River. Diversity. 2020; 12(4):158. https://doi.org/10.3390/d12040158
Chicago/Turabian StyleGama, Mafalda, Filipe Banha, Cristina Moreira, Henrique Gama, Manuel Graça, and Pedro Anastácio. 2020. "Patterns of Distribution of Bivalve Populations in a Mediterranean Temporary River" Diversity 12, no. 4: 158. https://doi.org/10.3390/d12040158
APA StyleGama, M., Banha, F., Moreira, C., Gama, H., Graça, M., & Anastácio, P. (2020). Patterns of Distribution of Bivalve Populations in a Mediterranean Temporary River. Diversity, 12(4), 158. https://doi.org/10.3390/d12040158