
A Comprehensive Overview of Stress, Resilience, and Neuroplasticity Mechanisms

 , , , and
, , , and
Abstract
1. Introduction
- External stimuli out of the subject’s control that impose demands on the organism;
- Neural processes that evaluate these demands and available resources;
- Physiological, behavioral, and subjective activations indicative of stress;
- Neuroadaptations in brain systems involved in emotion and motivation under chronic stress;
- Cognitive, physiological, and behavioral adaptations in response to stressors.

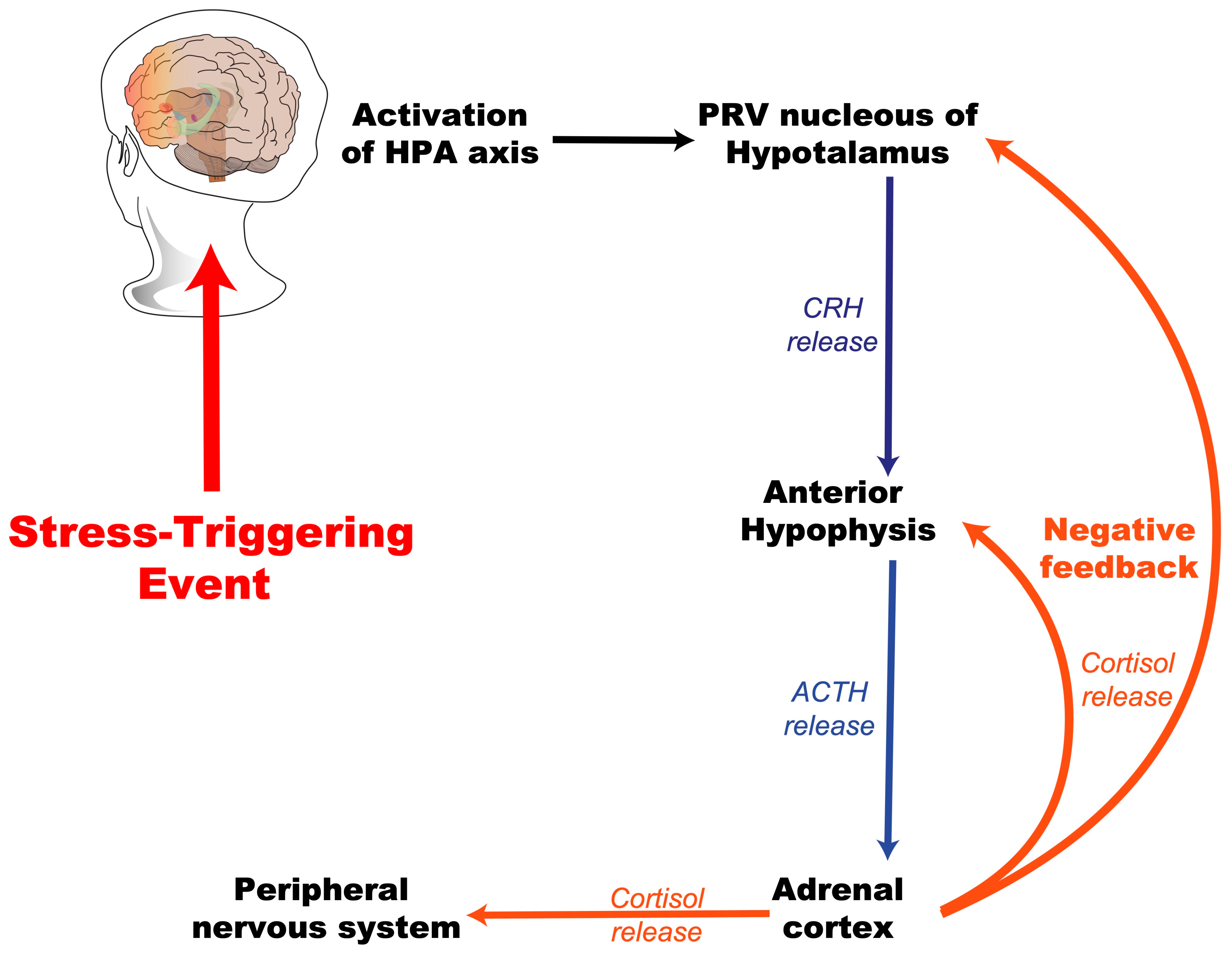
2. Stress
2.1. Early-Life Stress
2.1.1. Match/Mismatch Hypothesis
2.1.2. Cumulative Stress Hypothesis
2.1.3. Three-Hit Concept
2.2. Early Stress and Critical Developmental Periods
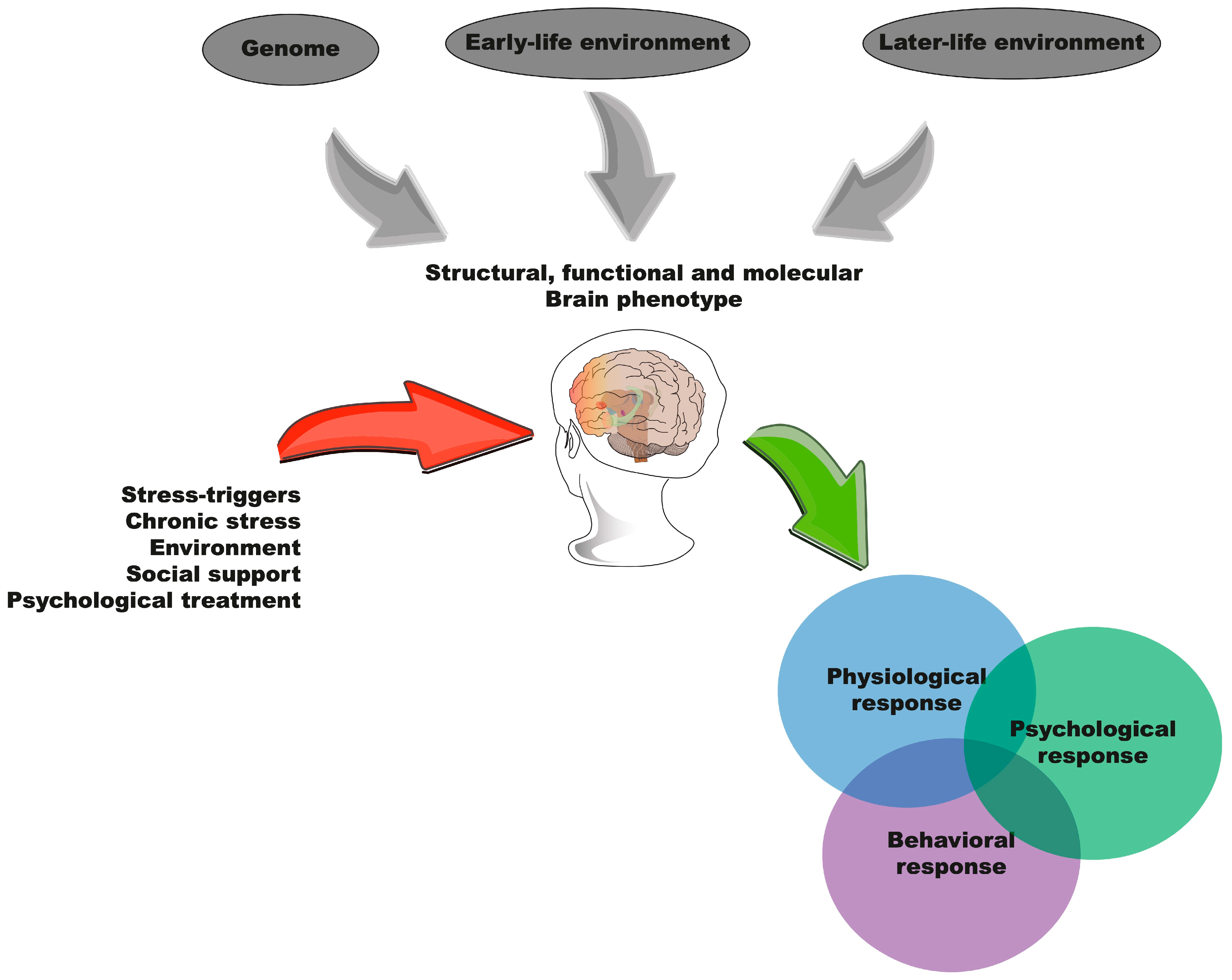
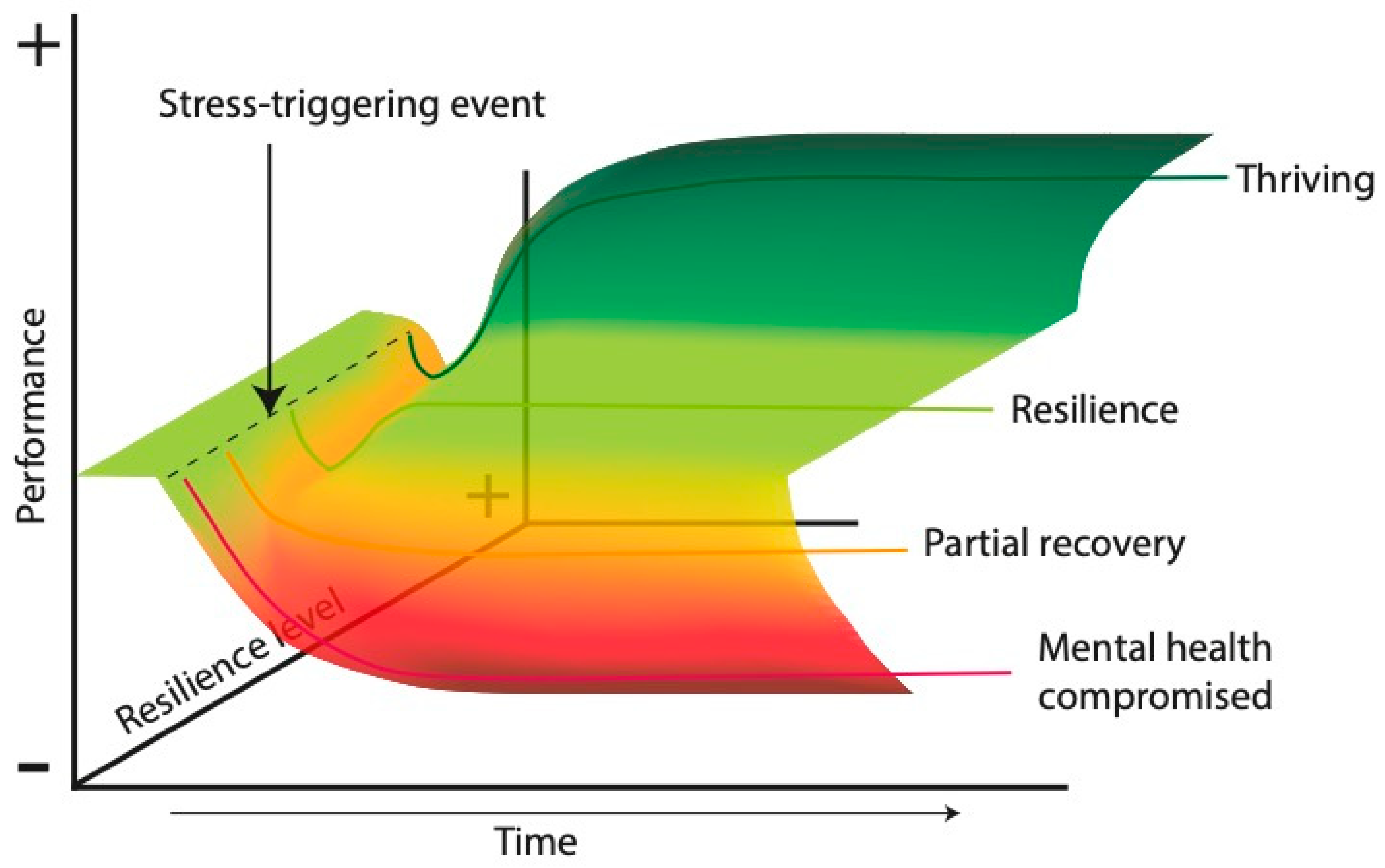
3. Resilience
3.1. Delimitation of the Concept of Stress Resilience
3.2. Models of Stress Resilience
3.2.1. Selective Breeding
3.2.2. Selection of Subpopulations
3.2.3. Transgenic Models
4. Neuroanatomical Structures and Plasticity Mechanisms Involved in Stress and Resilience Responses
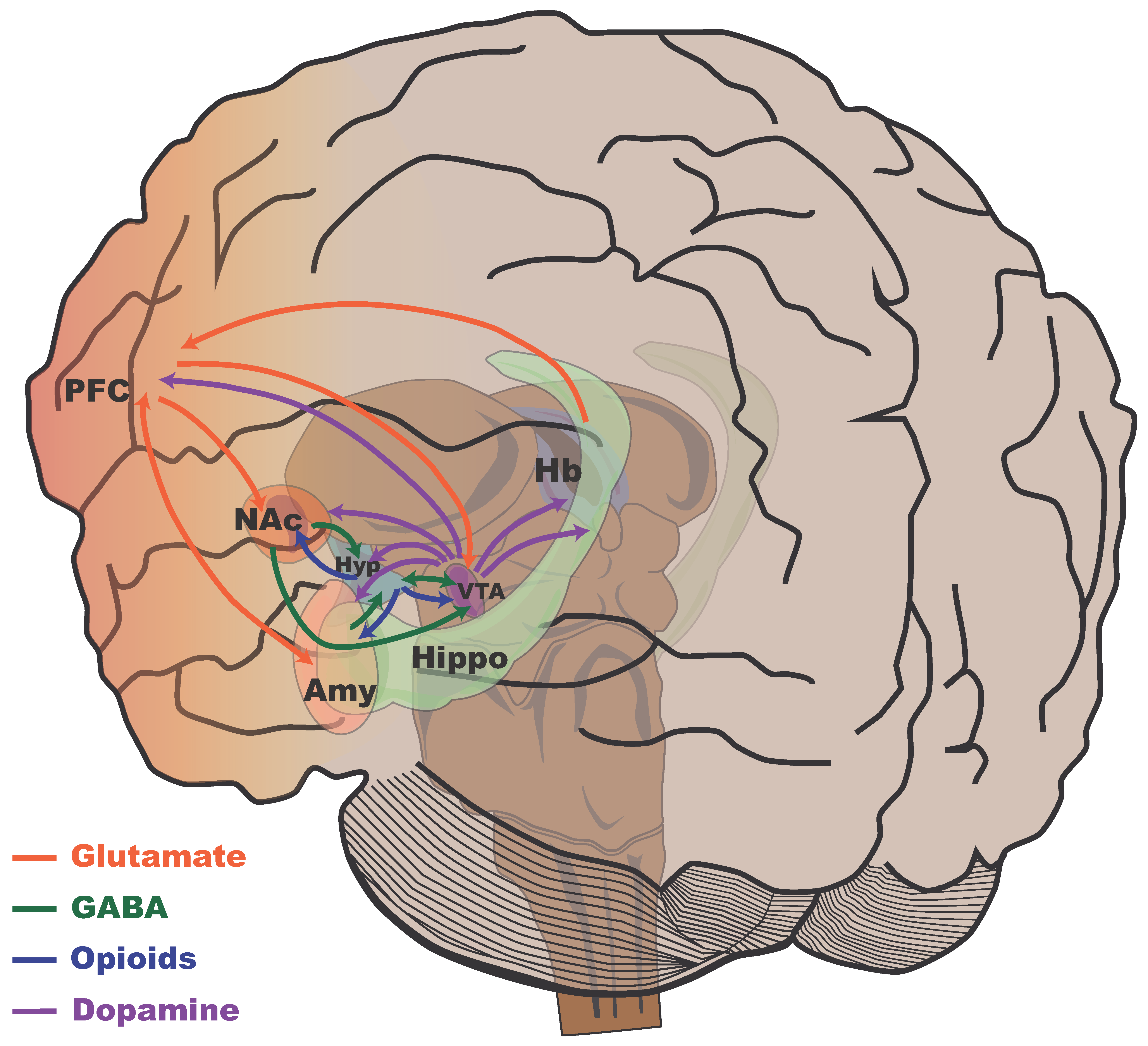
4.1. Reward Circuit
4.2. Reward Circuit, Stress, and Resilience
4.3. Plasticity Mechanisms Associated with Stress and Resilience
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ELS | Early-life stress |
| HPA | Hypothalamic–pituitary–adrenal axis |
| PFC | Prefrontal cortex |
| CORT | Corticosterone |
| EEG | Electroencephalography |
| DNA | Deoxyribonucleic acid |
| FST | Forced swimming test |
| BDNF | Brain-derived neurotrophic factor |
| 5-HT | Serotonin |
| CMS | Chronic mild stress |
| BBB | Blood–brain barrier |
| KO | Knockout |
| GLU | Modulated glutamate |
| VGLUT1 | Vesicular glutamate transporter 1 |
| DA | Dopamine |
| OPIs | Opioid peptides |
| GABA | Gamma-aminobutyric acid |
| VTA | Ventral tegmental area |
| NA | Norepinephrine |
| NAc | Nucleus Accumbens |
| Hyp | Hypothalamus |
| Hb | Habenula |
| LHb | Lateral habenula |
| Amy | Amygdala |
| Hippo | Hippocampus |
| iTRAQ | Isobaric tags for relative and absolute quantitation |
| AHN | Adult hippocampal neurogenesis |
| NPCs | Neural progenitor cells |
| FGF-2 | Fibroblast growth factor 2 |
| VEGF | Vascular endothelial growth factor |
| IGF-1 | Insulin-like growth factor 1 |
References
- Rutter, M. Resilience, Competence, and Coping. Child Abus. Negl. 2007, 31, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Uriarte Arciniega, J.D.D. Resilience. A New Perspective into Developmental Psychopathology. Rev. Psicodidáct. 2005, 10, 61–80. [Google Scholar]
- Sinha, R. Chronic Stress, Drug Use, and Vulnerability to Addiction. Ann. N. Y. Acad. Sci. 2008, 1141, 105–130. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S.; Gianaros, P.J. Stress- and Allostasis-Induced Brain Plasticity. Annu. Rev. Med. 2011, 62, 431–445. [Google Scholar] [CrossRef]
- Osório, C.; Probert, T.; Jones, E.; Young, A.H.; Robbins, I. Adapting to Stress: Understanding the Neurobiology of Resilience. Behav. Med. 2017, 43, 307–322. [Google Scholar] [CrossRef]
- McEwen, B.S.; Nasca, C.; Gray, J.D. Stress Effects on Neuronal Structure: Hippocampus, Amygdala, and Prefrontal Cortex. Neuropsychopharmacology 2016, 41, 3–23. [Google Scholar] [CrossRef]
- Hanchate, N.K. Single-cell Genomics Meets Systems Neuroscience: Insights from Mapping the Brain Circuitry of Stress. J. Neuroendocrinol. 2025, e70005. [Google Scholar] [CrossRef]
- Aldwin, C.M.S. Coping, and Development: An Integrative Perspective, 3rd ed.; Guilford Press: New York, NY, USA, 2009. [Google Scholar]
- Karatsoreos, I.N.; McEwen, B.S. Psychobiological Allostasis: Resistance, Resilience and Vulnerability. Trends Cogn. Sci. 2011, 15, 576–584. [Google Scholar] [CrossRef]
- Torres-Berrio, A.; Cuesta, S.; Lopez-Guzman, S.; Nava-Mesa, M.O. Interaction between Stress and Addiction: Contributions from Latin-American Neuroscience. Front. Psychol. 2018, 9, 2639. [Google Scholar] [CrossRef]
- Cathomas, F.; Murrough, J.W.; Nestler, E.J.; Han, M.H.; Russo, S.J. Neurobiology of Resilience: Interface Between Mind and Body. Biol. Psychiatry 2019, 86, 410–420. [Google Scholar] [CrossRef]
- McEwen, B.S.; Mirsky, A.E.; Hatch, M.M. Physiology and Neurobiology of Stress and Adaptation: Central Role of the Brain. Physiol. Rev. 2007, 87, 873–904. [Google Scholar] [CrossRef] [PubMed]
- Lucassen, P.J.; Oomen, C.A.; Naninck, E.F.G.; Fitzsimons, C.P.; Van Dam, A.M.; Czeh, B.; Korosi, A. Regulation of Adult Neurogenesis and Plasticity by (Early) Stress, Glucocorticoids, and Inflammation. Cold Spring Harb. Perspect. Biol. 2015, 7, a021303. [Google Scholar] [CrossRef] [PubMed]
- Lupien, S.J.; Maheu, F.; Tu, M.; Fiocco, A.; Schramek, T.E. The Effects of Stress and Stress Hormones on Human Cognition: Implications for the Field of Brain and Cognition. Brain Cogn. 2007, 65, 209–237. [Google Scholar] [CrossRef]
- McEwen, B.S. Structural Plasticity of the Adult Brain: How Animal Models Help Us Understand Brain Changes in Depression and Systemic Disorders Related to Depression. Dialogues Clin. Neurosci. 2004, 6, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Selye, H. A Syndrome Produced by Diverse Nocuous Agents. Nature 1936, 138, 32. [Google Scholar] [CrossRef]
- Lu, S.; Wei, F.; Li, G. The Evolution of the Concept of Stress and the Framework of the Stress System. Cell Stress 2021, 5, 76–85. [Google Scholar] [CrossRef]
- Gulyaeva, N.V. Functional Neurochemistry of the Ventral and Dorsal Hippocampus: Stress, Depression, Dementia and Remote Hippocampal Damage. Neurochem. Res. 2019, 44, 1306–1322. [Google Scholar] [CrossRef]
- Horii-Hayashi, N.; Sasagawa, T.; Matsunaga, W.; Matsusue, Y.; Azuma, C.; Nishi, M. Developmental Changes in Desensitisation of C-Fos Expression Induced by Repeated Maternal Separation in Pre-Weaned Mice. J. Neuroendocrinol. 2013, 25, 158–167. [Google Scholar] [CrossRef]
- Jahng, J.W.; Ryu, V.; Yoo, S.B.; Noh, S.J.; Kim, J.Y.; Lee, J.H. Mesolimbic Dopaminergic Activity Responding to Acute Stress Is Blunted in Adolescent Rats That Experienced Neonatal Maternal Separation. Neuroscience 2010, 171, 144–152. [Google Scholar] [CrossRef]
- Ryu, V.; Lee, J.H.; Yoo, S.B.; Gu, X.F.; Moon, Y.W.; Jahng, J.W. Sustained Hyperphagia in Adolescent Rats That Experienced Neonatal Maternal Separation. Int. J. Obes. 2008, 32, 1355–1362. [Google Scholar] [CrossRef]
- Feng, X.; Wang, L.; Yang, S.; Qin, D.; Wang, J.; Li, C.; Lv, L.; Ma, Y.; Hu, X. Maternal Separation Produces Lasting Changes in Cortisol and Behavior in Rhesus Monkeys. Proc. Natl. Acad. Sci. USA 2011, 108, 14312–14317. [Google Scholar] [CrossRef] [PubMed]
- Bevan, K.; Kumari, M. Maternal Separation in Childhood and Hair Cortisol Concentrations in Late Adulthood. Psychoneuroendocrinology 2021, 130, 105253. [Google Scholar] [CrossRef] [PubMed]
- Mackes, N.K.; Golm, D.; Sarkar, S.; Kumsta, R.; Rutter, M.; Fairchild, G.; Mehta, M.A.; Sonuga-Barke, E.J.S. Early Childhood Deprivation Is Associated with Alterations in Adult Brain Structure despite Subsequent Environmental Enrichment. Proc. Natl. Acad. Sci. USA 2020, 117, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Mehta, M.A.; Golembo, N.I.; Nosarti, C.; Colvert, E.; Mota, A.; Williams, S.C.R.; Rutter, M.; Sonuga-Barke, E.J.S. Amygdala, Hippocampal and Corpus Callosum Size Following Severe Early Institutional Deprivation: The English and Romanian Adoptees Study Pilot. J. Child. Psychol. Psychiatry 2009, 50, 943–951. [Google Scholar] [CrossRef]
- Francisco Juruena, M.; Von Werne Baes, C.; Castro Menezes, I.; Guilherme Graeff, F. Early Life Stress in Depressive Patients: Role of Glucocorticoid and Mineralocorti-Coid Receptors and of Hypothalamic-Pituitary-Adrenal Axis Activity. Curr. Pharm. Des. 2015, 21, 1369. [Google Scholar]
- Wesarg, C.; Van Den Akker, A.L.; Oei, N.Y.L.; Hoeve, M.; Wiers, R.W. Identifying Pathways from Early Adversity to Psychopathology: A Review on Dysregulated HPA Axis Functioning and Impaired Self-Regulation in Early Childhood. Eur. J. Dev. Psychol. 2020, 17, 808–827. [Google Scholar] [CrossRef]
- Sapolsky, R.M.; Meaney, M.J. Maturation of the Adrenocortical Stress Response: Neuroendocrine Control Mechanisms and the Stress Hyporesponsive Period. Brain Res. Rev. 1986, 11, 65–76. [Google Scholar]
- Tarry-Adkins, J.L.; Ozanne, S.E. Mechanisms of Early Life Programming: Current Knowledge and Future Directions. Am. J. Clin. Nutr. 2011, 94, S1765–S1771. [Google Scholar] [CrossRef]
- Bale, T.L.; Baram, T.Z.; Brown, A.S.; Goldstein, J.M.; Insel, T.R.; McCarthy, M.M.; Nemeroff, C.B.; Reyes, T.M.; Simerly, R.B.; Susser, E.S.; et al. Early Life Programming and Neurodevelopmental Disorders. Biol. Psychiatry 2010, 68, 314–319. [Google Scholar] [CrossRef]
- Daskalakis, N.P.; Bagot, R.C.; Parker, K.J.; Vinkers, C.H.; de Kloet, E.R. The Three-Hit Concept of Vulnerability and Resilience: Toward Understanding Adaptation to Early-Life Adversity Outcome. Psychoneuroendocrinology 2013, 38, 1858–1873. [Google Scholar] [CrossRef]
- Nesse, R.M. On the Difficulty of Defining Disease: A Darwinian Perspective. Med. Health Care Philos. 2001, 4, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Nederhof, E.; Schmidt, M.V. Mismatch or Cumulative Stress: Toward an Integrated Hypothesis of Programming Effects. Physiol. Behav. 2012, 106, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.V. Animal Models for Depression and the Mismatch Hypothesis of Disease. Psychoneuroendocrinology 2011, 36, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.E. Mechanisms Linking Early Life Stress to Adult Health Outcomes. Proc. Natl. Acad. Sci. USA 2010, 107, 8507–8512. [Google Scholar]
- Champagne, D.L.; Bagot, R.C.; Van Hasselt, F.; Ramakers, G.; Meaney, M.J.; De Kloet, E.R.; Joëls, M.; Krugers, H. Maternal Care and Hippocampal Plasticity: Evidence for Experience-Dependent Structural Plasticity, Altered Synaptic Functioning, and Differential Responsiveness to Glucocorticoids and Stress. J. Neurosci. 2008, 28, 6037–6045. [Google Scholar] [CrossRef]
- Bagot, R.C.; van Hasselt, F.N.; Champagne, D.L.; Meaney, M.J.; Krugers, H.J.; Joëls, M. Maternal Care Determines Rapid Effects of Stress Mediators on Synaptic Plasticity in Adult Rat Hippocampal Dentate Gyrus. Neurobiol. Learn. Mem. 2009, 92, 292–300. [Google Scholar] [CrossRef]
- Lloyd, D.A.; Turner, R.J. Cumulative Lifetime Adversities and Alcohol Dependence in Adolescence and Young Adulthood. Drug Alcohol. Depend. 2008, 93, 217–226. [Google Scholar]
- Bergström, A.; Jayatissa, M.N.; Mørk, A.; Wiborg, O. Stress Sensitivity and Resilience in the Chronic Mild Stress Rat Model of Depression; An in Situ Hybridization Study. Brain Res. 2008, 1196, 41–52. [Google Scholar] [CrossRef]
- Ho, T.C.; King, L.S. Mechanisms of Neuroplasticity Linking Early Adversity to Depression: Developmental Considerations. Transl. Psychiatry 2021, 11, 517. [Google Scholar] [CrossRef]
- Murthy, S.; Kane, G.A.; Katchur, N.J.; Lara Mejia, P.S.; Obiofuma, G.; Buschman, T.J.; McEwen, B.S.; Gould, E. Perineuronal Nets, Inhibitory Interneurons, and Anxiety-Related Ventral Hippocampal Neuronal Oscillations Are Altered by Early Life Adversity. Biol. Psychiatry 2019, 85, 1011–1020. [Google Scholar] [CrossRef]
- Abbink, M.R.; Naninck, E.F.G.; Lucassen, P.J.; Korosi, A. Early-Life Stress Diminishes the Increase in Neurogenesis after Exercise in Adult Female Mice. Hippocampus 2017, 27, 839–844. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, C.F.; Phifer, J.; Bradley, B.; Ressler, K.J. Risk and Resilience: Genetic and Environmental Influences on Development of the Stress Response. Depress. Anxiety 2009, 26, 984–992. [Google Scholar] [CrossRef] [PubMed]
- MacKinnon, N.; Kingsbury, M.; Mahedy, L.; Evans, J.; Colman, I. The Association Between Prenatal Stress and Externalizing Symptoms in Childhood: Evidence From the Avon Longitudinal Study of Parents and Children. Biol. Psychiatry 2018, 83, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Ramsteijn, A.S.; Verkaik-Schakel, R.N.; Houwing, D.J.; Plösch, T.; Olivier, J.D.A. Perinatal Exposure to Fluoxetine and Maternal Adversity Affect Myelin-Related Gene Expression and Epigenetic Regulation in the Corticolimbic Circuit of Juvenile Rats. Neuropsychopharmacology 2022, 47, 1620–1632. [Google Scholar] [CrossRef]
- Triplett, R.L.; Lean, R.E.; Parikh, A.; Miller, J.P.; Alexopoulos, D.; Kaplan, S.; Meyer, D.; Adamson, C.; Smyser, T.A.; Rogers, C.E.; et al. Association of Prenatal Exposure to Early-Life Adversity with Neonatal Brain Volumes at Birth. JAMA Netw. Open 2022, 5, E227045. [Google Scholar] [CrossRef]
- Gómez-González, B.; Escobar, A. Altered Functional Development of the Blood-Brain Barrier after Early Life Stress in the Rat. Brain Res. Bull. 2009, 79, 376–387. [Google Scholar] [CrossRef]
- Gómez-González, B.; Escobar, A. Prenatal Stress Alters Microglial Development and Distribution in Postnatal Rat Brain. Acta Neuropathol. 2010, 119, 303–315. [Google Scholar] [CrossRef]
- Weinstock, M. Prenatal Stressors in Rodents: Effects on Behavior. Neurobiol. Stress 2017, 6, 3–13. [Google Scholar] [CrossRef]
- Class, Q.A.; Abel, K.M.; Khashan, A.S.; Rickert, M.E.; Dalman, C.; Larsson, H.; Hultman, C.M.; Långström, N.; Lichtenstein, P.; D’Onofrio, B.M. Offspring Psychopathology Following Preconception, Prenatal and Postnatal Maternal Bereavement Stress. Psychol. Med. 2014, 44, 71–84. [Google Scholar] [CrossRef]
- Humphreys, K.L.; Camacho, M.C.; Roth, M.C.; Estes, E.C. Prenatal Stress Exposure and Multimodal Assessment of Amygdala–Medial Prefrontal Cortex Connectivity in Infants. Dev. Cogn. Neurosci. 2020, 46, 100877. [Google Scholar] [CrossRef]
- Graham, A.M.; Rasmussen, J.M.; Entringer, S.; Ben Ward, E.; Rudolph, M.D.; Gilmore, J.H.; Styner, M.; Wadhwa, P.D.; Fair, D.A.; Buss, C. Maternal Cortisol Concentrations During Pregnancy and Sex-Specific Associations with Neonatal Amygdala Connectivity and Emerging Internalizing Behaviors. Biol. Psychiatry 2019, 85, 172–181. [Google Scholar] [CrossRef]
- Benoit, J.D.; Rakic, P.; Frick, K.M. Prenatal Stress Induces Spatial Memory Deficits and Epigenetic Changes in the Hippocampus Indicative of Heterochromatin Formation and Reduced Gene Expression. Behav. Brain Res. 2015, 281, 1–8. [Google Scholar] [CrossRef]
- Monk, C.; Feng, T.; Lee, S.; Krupska, I.; Champagne, F.A.; Tycko, B. Distress during Pregnancy: Epigenetic Regulation of Placenta Glucocorticoid-Related Genes and Fetal Neurobehavior. Am. J. Psychiatry 2016, 173, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Moog, N.K.; Entringer, S.; Rasmussen, J.M.; Styner, M.; Gilmore, J.H.; Kathmann, N.; Heim, C.M.; Wadhwa, P.D.; Buss, C. Intergenerational Effect of Maternal Exposure to Childhood Maltreatment on Newborn Brain Anatomy. Biol. Psychiatry 2018, 83, 120–127. [Google Scholar] [CrossRef] [PubMed]
- U.S. Department of Health & Human Services; Administration for Children and Families; Administration on Children, Youth and Families. Children’s Bureau Child Maltreatment 2022. Available online: https://www.acf.hhs.gov/cb/data-research/child-maltreatment (accessed on 9 March 2025).
- McLaughlin, K.A.; Sheridan, M.A.; Nelson, C.A. Neglect as a Violation of Species-Expectant Experience: Neurodevelopmental Consequences. Biol. Psychiatry 2017, 82, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Vanderwert, R.E.; Zeanah, C.H.; Fox, N.A.; Nelson, C.A. Normalization of EEG Activity among Previously Institutionalized Children Placed into Foster Care: A 12-Year Follow-up of the Bucharest Early Intervention Project. Dev. Cogn. Neurosci. 2016, 17, 68–75. [Google Scholar] [CrossRef]
- Alberini, C.M.; Travaglia, A. Infantile Amnesia: A Critical Period of Learning to Learn and Remember. J. Neurosci. 2017, 37, 5783–5795. [Google Scholar] [CrossRef]
- Sheridan, M.A.; Shi, F.; Miller, A.B.; Salhi, C.; McLaughlin, K.A. Network Structure Reveals Clusters of Associations between Childhood Adversities and Development Outcomes. Dev. Sci. 2020, 23, e12934. [Google Scholar] [CrossRef]
- Gunnar, M.R.; Hostinar, C.E.; Sanchez, M.M.; Tottenham, N.; Sullivan, R.M. Parental Buffering of Fear and Stress Neurobiology: Reviewing Parallels across Rodent, Monkey, and Human Models. Soc. Neurosci. 2015, 10, 474–478. [Google Scholar] [CrossRef]
- Pfeiffer, E.; Sukale, T.; Müller, L.R.F.; Plener, P.L.; Rosner, R.; Fegert, J.M.; Sachser, C.; Unterhitzenberger, J. The Symptom Representation of Posttraumatic Stress Disorder in a Sample of Unaccompanied and Accompanied Refugee Minors in Germany: A Network Analysis. Eur. J. Psychotraumatol. 2019, 10, 1675990. [Google Scholar] [CrossRef]
- Scharpf, F.; Saupe, L.; Crombach, A.; Haer, R.; Ibrahim, H.; Neuner, F.; Peltonen, K.; Qouta, S.; Saile, R.; Hecker, T. The Network Structure of Posttraumatic Stress Symptoms in War-Affected Children and Adolescents. JCPP Adv. 2023, 3, e12124. [Google Scholar] [CrossRef] [PubMed]
- Dickson, M.J.; Vargo, K.K. Training Kindergarten Students Lockdown Drill Procedures Using Behavioral Skills Training. J. Appl. Behav. Anal. 2017, 50, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Davis, E.P.; Stout, S.A.; Molet, J.; Vegetabile, B.; Glynn, L.M.; Sandman, C.A.; Heins, K.; Stern, H.; Baram, T.Z. Exposure to Unpredictable Maternal Sensory Signals Influences Cognitive Development across Species. Proc. Natl. Acad. Sci. USA 2017, 114, 10390–10395. [Google Scholar] [CrossRef] [PubMed]
- Eiland, L.; Ramroop, J.; Hill, M.N.; Manley, J.; McEwen, B.S. Chronic Juvenile Stress Produces Corticolimbic Dendritic Architectural Remodeling and Modulates Emotional Behavior in Male and Female Rats. Psychoneuroendocrinology 2012, 37, 39–47. [Google Scholar] [CrossRef]
- Breiner, K.; Li, A.; Cohen, A.O.; Steinberg, L.; Bonnie, R.J.; Scott, E.S.; Taylor-Thompson, K.; Rudolph, M.D.; Chein, J.; Richeson, J.A.; et al. Combined Effects of Peer Presence, Social Cues, and Rewards on Cognitive Control in Adolescents. Dev. Psychobiol. 2018, 60, 292–302. [Google Scholar] [CrossRef]
- Gunnar, M.R.; Depasquale, C.E.; Reid, B.M.; Donzella, B.; Miller, B.S. Pubertal Stress Recalibration Reverses the Effects of Early Life Stress in Postinstitutionalized Children. Proc. Natl. Acad. Sci. USA 2019, 116, 23984–23988. [Google Scholar] [CrossRef]
- King, L.S.; Graber, M.G.; Colich, N.L.; Gotlib, I.H. Associations of Waking Cortisol with DHEA and Testosterone across the Pubertal Transition: Effects of Threat-Related Early Life Stress. Psychoneuroendocrinology 2020, 115, 104651. [Google Scholar] [CrossRef]
- Weaver, I.C.G.; Diorio, J.; Seckl, J.R.; Szyf, M.; Meaney, M.J. Early Environmental Regulation of Hippocampal Glucocorticoid Receptor Gene Expression: Characterization of Intracellular Mediators and Potential Genomic Target Sites. Ann. N. Y. Acad. Sci. 2004, 1024, 182–212. [Google Scholar] [CrossRef]
- Murgatroyd, C.; Patchev, A.V.; Wu, Y.; Micale, V.; Bockmühl, Y.; Fischer, D.; Holsboer, F.; Wotjak, C.T.; Almeida, O.F.X.; Spengler, D. Dynamic DNA Methylation Programs Persistent Adverse Effects of Early-Life Stress. Nat. Neurosci. 2009, 12, 1559–1566. [Google Scholar]
- Rutter, M. Social Context: Meanings, Measures and Mechanisms. Eur. Rev. 1999, 7, 139–149. [Google Scholar]
- Masten, A.S.; Cicchetti, D. Resilience in Development: Progress and Transformation. Dev. Psychopathol. 2016, 4, 271–333. [Google Scholar]
- Southwick, S.M.; Bonanno, G.A.; Masten, A.S.; Panter-Brick, C.; Yehuda, R. Resilience Definitions, Theory, and Challenges: Interdisciplinary Perspectives. Eur. J. Psychotraumatol. 2014, 5, 25338. [Google Scholar] [CrossRef] [PubMed]
- Russo-Neustadt, A.A.; Chen, M.J. Brain-Derived Neurotrophic Factor and Antidepressant Activity. Curr. Pharm. Des. 2005, 11, 1495–1510. [Google Scholar] [CrossRef] [PubMed]
- Spivey, J.; Barrett, D.; Padilla, E.; Gonzalez-Lima, F. Mother-Infant Separation Leads to Hypoactive Behavior in Adolescent Holtzman Rats. Behav. Process. 2008, 79, 59–65. [Google Scholar] [CrossRef]
- Lyons, D.M.; Parker, K.J.; Schatzberg, A.F. Animal Models of Early Life Stress: Implications for Understanding Resilience. Dev. Psychobiol. 2010, 52, 402–410. [Google Scholar] [CrossRef]
- Wu, G.; Feder, A.; Cohen, H.; Kim, J.J.; Calderon, S.; Charney, D.S.; Mathé, A.A. Understanding Resilience. Front. Behav. Neurosci. 2013, 7, 10. [Google Scholar] [CrossRef]
- Cabanyes Truffino, J. Resilience: An Approach to the Concept. Rev. Psiquiatr. Salud Ment. 2010, 3, 145–151. [Google Scholar] [CrossRef]
- Carver, C.S. Resilience and Thriving: Issues, Models, and Linkages. J. Social Issues 1998, 54, 245–266. [Google Scholar] [CrossRef]
- Russo, S.J.; Murrough, J.W.; Han, M.H.; Charney, D.S.; Nestler, E.J. Neurobiology of Resilience. Nat. Neurosci. 2012, 15, 1475–1484. [Google Scholar] [CrossRef]
- Jaimes-Hoy, L.; Pérez-Maldonado, A.; Bahena, E.N.; de la Cruz Guarneros, N.; Rodríguez-Rodríguez, A.; Charli, J.L.; Soberón, X.; Joseph-Bravo, P. Sex Dimorphic Changes in Trh Gene Methylation and Thyroid-Axis Response to Energy Demands in Maternally Separated Rats. Endocrinology 2021, 162, bqab110. [Google Scholar] [CrossRef]
- Carnevali, L.; Koenig, J.; Sgoifo, A.; Ottaviani, C. Autonomic and Brain Morphological Predictors of Stress Resilience. Front. Neurosci. 2018, 12, 228. [Google Scholar] [CrossRef]
- Feder, A.; Nestler, E.J.; Charney, D.S. Psychobiology and Molecular Genetics of Resilience. Nat. Rev. Neurosci. 2009, 10, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Kim-Cohen, J.; Gold, A.L. Measured Gene-Environment Interactions and Mechanisms Promoting Resilient. Psychol. Sci. 2009, 18, 138–142. [Google Scholar]
- Maras, P.M.; Hebda-Bauer, E.K.; Hagenauer, M.H.; Hilde, K.L.; Blandino, P.; Watson, S.J.; Akil, H. Differences in Microglia Morphological Profiles Reflect Divergent Emotional Temperaments: Insights from a Selective Breeding Model. Transl. Psychiatry 2022, 12, 105. [Google Scholar] [CrossRef]
- Stedenfeld, K.A.; Clinton, S.M.; Kerman, I.A.; Akil, H.; Watson, S.J.; Sved, A.F. Novelty-Seeking Behavior Predicts Vulnerability in a Rodent Model of Depression. Physiol. Behav. 2011, 103, 210–216. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, C.; Ji, Y.; Yang, L. Biological and Psychological Perspectives of Resilience: Is It Possible to Improve Stress Resistance? Front. Hum. Neurosci. 2018, 12, 326. [Google Scholar] [CrossRef]
- Touma, C.; Bunck, M.; Glasl, L.; Nussbaumer, M.; Palme, R.; Stein, H.; Wolferstätter, M.; Zeh, R.; Zimbelmann, M.; Holsboer, F.; et al. Mice Selected for High versus Low Stress Reactivity: A New Animal Model for Affective Disorders. Psychoneuroendocrinology 2008, 33, 839–862. [Google Scholar] [CrossRef]
- Scott, P.A.; Cierpial, M.A.; Kilts, C.D.; Weiss, J.M. Susceptibility and Resistance of Rats to Stress-Induced Decreases in Swim-Test Activity: A Selective Breeding Study. Brain Res. 1996, 725, 217–230. [Google Scholar]
- Panocka, I.; Marek, P.; Sadowski, B. Differentiation of Neurochemical Basis of Stress-Induced Analgesia in Mice by Selective Breeding. Brain Res. 1986, 397, 156–160. [Google Scholar]
- Lankford, S.E.; Weber, G.M. Associations between Plasma Growth Hormone, Insulin-Like Growth Factor-I, and Cortisol with Stress Responsiveness and Growth Performance in a Selective Breeding Program for Rainbow Trout. N. Am. J. Aquac. 2006, 68, 151–159. [Google Scholar] [CrossRef]
- Weber, G.M.; Silverstein, J.T. Evaluation of a Stress Response for Use in a Selective Breeding Program for Improved Growth and Disease Resistance in Rainbow Trout. N. Am. J. Aquac. 2007, 69, 69–79. [Google Scholar] [CrossRef]
- Bunck, M.; Czibere, L.; Horvath, C.; Graf, C.; Frank, E.; Keßler, M.S.; Murgatroyd, C.; Müller-Myhsok, B.; Gonik, M.; Weber, P.; et al. A Hypomorphic Vasopressin Allele Prevents Anxiety-Related Behavior. PLoS ONE 2009, 4, e5129. [Google Scholar] [CrossRef]
- Hodes, G.E.; Kana, V.; Menard, C.; Merad, M.; Russo, S.J. Neuroimmune Mechanisms of Depression. Nat. Neurosci. 2015, 18, 1386–1393. [Google Scholar] [CrossRef] [PubMed]
- Felger, J.C.; Li, Z.; Haroon, E.; Woolwine, B.J.; Jung, M.Y.; Hu, X.; Miller, A.H. Inflammation Is Associated with Decreased Functional Connectivity within Corticostriatal Reward Circuitry in Depression. Mol. Psychiatry 2016, 21, 1358–1365. [Google Scholar] [CrossRef]
- Ménard, C.; Pfau, M.L.; Hodes, G.E.; Russo, S.J. Immune and Neuroendocrine Mechanisms of Stress Vulnerability and Resilience. Neuropsychopharmacology 2017, 42, 62–80. [Google Scholar] [CrossRef]
- Christensen, T.; Bisgaard, C.F.; Wiborg, O. Biomarkers of Anhedonic-like Behavior, Antidepressant Drug Refraction, and Stress Resilience in a Rat Model of Depression. Neuroscience 2011, 196, 66–79. [Google Scholar] [CrossRef]
- Delgado Y Palacios, R.; Campo, A.; Henningsen, K.; Verhoye, M.; Poot, D.; Dijkstra, J.; Van Audekerke, J.; Benveniste, H.; Sijbers, J.; Wiborg, O.; et al. Magnetic Resonance Imaging and Spectroscopy Reveal Differential Hippocampal Changes in Anhedonic and Resilient Subtypes of the Chronic Mild Stress Rat Model. Biol. Psychiatry 2011, 70, 449–457. [Google Scholar] [CrossRef]
- Scharf, S.H.; Schmidt, M.V. Animal Models of Stress Vulnerability and Resilience in Translational Research. Curr. Psychiatry Rep. 2012, 14, 159–165. [Google Scholar] [CrossRef]
- Wagner, K.V.; Wang, X.-D.; Liebl, C.; Scharf, S.H.; Müller, M.B.; Schmidt, M.V. Pituitary glucocorticoid receptor deletion reduces vulnerability to chronic stress. Psychoneuroendocrinology 2011, 36, 579–587. [Google Scholar] [CrossRef]
- Garcia-Garcia, A.L.; Elizalde, N.; Matrov, D.; Harro, J.; Wojcik, S.M.; Venzala, E.; Ramírez, M.J.; Del Rio, J.; Tordera, R.M. Increased Vulnerability to Depressive-Like Behavior of Mice with Decreased Expression of VGLUT1. Biol. Psychiatry 2009, 66, 275–282. [Google Scholar] [CrossRef]
- Willner, P. Dopamine and Depression: A Review of Recent Evidente. II. Theoretical Approaches. Brain Res. Rev. 1983, 6, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Juarez, J.; Barrios De Tomasi, E.; Muñoz-Villegas, P.; Buenrostro, M. Adicción Farmacológica y Conductual. In Cerebro y Conducta; Manual Moderno: Guadalajara, Mexico, 2015. [Google Scholar]
- Salmanzadeh, H.; Ahmadi-Soleimani, S.M.; Pachenari, N.; Azadi, M.; Halliwell, R.F.; Rubino, T.; Azizi, H. Adolescent Drug Exposure: A Review of Evidence for the Development of Persistent Changes in Brain Function. Brain Res. Bull. 2020, 156, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Yehuda, S.; Wurtman, R.J.; Yehuda, S.; Wurtman, R.J. Dopaminergic Neurons in the Nigro-Striatal and Mesolimbic Pathways: Mediation of Specific Effects of D-Amphetamine. Eur. J. Pharmacol. 1975, 30, 154–158. [Google Scholar] [CrossRef]
- Feltenstein, M.W.; See, R.E. Systems Level Neuroplasticity in Drug Addiction. Cold Spring Harb. Perspect. Med. 2013, 3, a011916. [Google Scholar] [CrossRef] [PubMed]
- Juárez, J.; Buenrostro-Jáuregui, M.H. Circuitos Cerebrales Involucrados En El Consumo de Drogas de Abuso. In Circuitos Cerebrales Implicados en la Cognición y Conducta; Hernández, M., Sanz, A., Guevara, M., Eds.; Manual Moderno: Guadalajara, Mexico, 2015; pp. 103–146. [Google Scholar]
- Iversen, S.; Kupfermann, I.; Kandel, E.R. Emotional States and Feelings. In Principles of Neural Science; Kandel, E.R., Schwartz, J.H., Jessell, T.M., Siegelbaum, S., Hudspeth, A.J., Mack, S., Eds.; McGraw-Hill: New York, NY, USA, 2000; pp. 982–997. [Google Scholar]
- Brodie, M.S.; Pesold, C.; Appel, S.B. Ethanol Directly Excites Dopaminergic Ventral Tegmental Area Reward Neurons. Alcohol. Clin. Exp. Res. 1999, 23, 1848–1852. [Google Scholar] [CrossRef]
- Corominas, M.; Roncero, C.; Ribases, M.; Castells, X.; Casas, M. Brain Derived Neurotrophic Factor (BDNF) and Its Intracellular Signalling Pathways in Cocaine Addiction. Neuropsychobiology 2007, 55, 2–13. [Google Scholar]
- Ahmad, T.; Sun, N.; Lyons, D.; Laviolette, S.R. Bi-Directional Cannabinoid Signalling in the Basolateral Amygdala Controls Rewarding and Aversive Emotional Processing via Functional Regulation of the Nucleus Accumbens. Addict. Biol. 2017, 22, 1218–1231. [Google Scholar] [CrossRef]
- Chau, B.K.H.; Jarvis, H.; Law, C.-K.; Chong, T.T.-J. Dopamine and Reward: A View from the Prefrontal Cortex. Behav. Pharmacol. 2018, 29, 569–583. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Liu, T.Y.; Qu, W.M.; Hong, Z.Y.; Urade, Y.; Huang, Z.L. Dopamine Is Involved in Food-Anticipatory Activity in Mice. J. Biol. Rhythm. 2012, 27, 398–409. [Google Scholar] [CrossRef]
- Pfaus, J.G.; Phillips, A.G. Role of Dopamine in Anticipatory and Consummatory Aspects of Sexual Behavior in the Male Rat. Behav. Neurosci. 1991, 105, 727–743. [Google Scholar] [CrossRef]
- Berridge, K.C. Food Reward: Brain Substrates of Wanting and Liking. Neurosci. Biobehav. Rev. 1996, 20, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Berridge, K.C. “Liking” and “Wanting” Food Rewards: Brain Substrates and Roles in Eating Disorders. Physiol. Behav. 2009, 97, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Barbano, M.F.; Cador, M. Opioids for Hedonic Experience and Dopamine to Get Ready for It. Psychopharmacology 2007, 191, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Margolis, E.B.; Hjelmstad, G.O.; Fujita, W.; Fields, H.L. Direct Bidirectional μ-Opioid Control of Midbrain Dopamine Neurons. J. Neurosci. 2014, 34, 14707–14716. [Google Scholar] [CrossRef]
- Febbraro, F.; Svenningsen, K.; Tran, T.P.; Wiborg, O. Neuronal Substrates Underlying Stress Resilience and Susceptibility in Rats. PLoS ONE 2017, 12, e0179434. [Google Scholar] [CrossRef]
- Anacker, C.; Scholz, J.; O’Donnell, K.J.; Allemang-Grand, R.; Diorio, J.; Bagot, R.C.; Nestler, E.J.; Hen, R.; Lerch, J.P.; Meaney, M.J. Neuroanatomic Differences Associated with Stress Susceptibility and Resilience. Biol. Psychiatry 2016, 79, 840–849. [Google Scholar] [CrossRef]
- Dias, C.; Feng, J.; Sun, H.; Shao, N.Y.; Mazei-Robison, M.S.; Damez-Werno, D.; Scobie, K.; Bagot, R.; Labonté, B.; Ribeiro, E.; et al. β-Catenin Mediates Stress Resilience through Dicer1/MicroRNA Regulation. Nature 2014, 516, S1–S5. [Google Scholar] [CrossRef]
- Tang, M.; Huang, H.; Li, S.; Zhou, M.; Liu, Z.; Huang, R.; Liao, W.; Xie, P.; Zhou, J. Hippocampal Proteomic Changes of Susceptibility and Resilience to Depression or Anxiety in a Rat Model of Chronic Mild Stress. Transl. Psychiatry 2019, 9, 260. [Google Scholar] [CrossRef]
- Sousa, V.C.; Vital, J.; Costenla, A.R.; Batalha, V.L.; Sebastião, A.M.; Ribeiro, J.A.; Lopes, L.V. Maternal Separation Impairs Long Term-Potentiation in CA1-CA3 Synapses and Hippocampal-Dependent Memory in Old Rats. Neurobiol. Aging 2014, 35, 1680–1685. [Google Scholar] [CrossRef]
- Reshetnikov, V.V.; Kovner, A.V.; Lepeshko, A.A.; Pavlov, K.S.; Grinkevich, L.N.; Bondar, N.P. Stress Early in Life Leads to Cognitive Impairments, Reduced Numbers of CA3 Neurons and Altered Maternal Behavior in Adult Female Mice. Genes Brain Behav. 2020, 19, e12541. [Google Scholar] [CrossRef]
- Leal-Galicia, P.; Romo-Parra, H.; Rodríguez-Serrano, L.M.; Buenrostro-Jáuregui, M. Regulation of Adult Hippocampal Neurogenesis Exerted by Sexual, Cognitive and Physical Activity: An Update. J. Chem. Neuroanat. 2019, 101, 101667. [Google Scholar] [CrossRef] [PubMed]
- Mirescu, C.; Peters, J.D.; Gould, E. Early Life Experience Alters Response of Adult Neurogenesis to Stress. Nat. Neurosci. 2004, 7, 841–846. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.X.; Su, Y.A.; Wang, Q.; Zheng, J.Y.; Zhang, C.C.; Wang, T.; Liu, X.; Ma, Y.N.; Li, X.X.; Zhang, X.Q.; et al. The Causal Involvement of the BDNF-TrkB Pathway in Dentate Gyrus in Early-Life Stress-Induced Cognitive Deficits in Male Mice. Transl. Psychiatry 2023, 13, 173. [Google Scholar] [CrossRef] [PubMed]
- Veena, J.; Rao, B.S.S.; Srikumar, B.N. Regulation of Adult Neurogenesis in the Hippocampus by Stress, Acetylcholine and Dopamine. J. Nat. Sci. Biol. Med. 2011, 2, 26–37. [Google Scholar] [CrossRef]
- Leuner, B.; Caponiti, J.M.; Gould, E. Oxytocin Stimulates Adult Neurogenesis Even under Conditions of Stress and Elevated Glucocorticoids. Hippocampus 2012, 22, 861–868. [Google Scholar] [CrossRef]
- Naninck, E.F.G.; Hoeijmakers, L.; Kakava-Georgiadou, N.; Meesters, A.; Lazic, S.E.; Lucassen, P.J.; Korosi, A. Chronic Early Life Stress Alters Developmental and Adult Neurogenesis and Impairs Cognitive Function in Mice. Hippocampus 2015, 25, 309–328. [Google Scholar] [CrossRef]
- Pencea, V.; Bingaman, K.D.; Wiegand, S.J.; Luskin, M.B. Infusion of Brain-Derived Neurotrophic Factor into the Lateral Ventricle of the Adult Rat Leads to New Neurons in the Parenchyma of the Striatum, Septum, Thalamus, and Hypothalamus. J. Neurosci. 2001, 21, 6706–6717. [Google Scholar] [CrossRef]
- Sachs, B.D.; Caron, M.G. Chronic Fluoxetine Increases Extra-Hippocampal Neurogenesis in Adult Mice. Int. J. Neuropsychopharmacol. 2015, 18, pyu029. [Google Scholar] [CrossRef]
- Fakhoury, M. The Dorsal Diencephalic Conduction System in Reward Processing: Spotlight on the Anatomy and Functions of the Habenular Complex. Behav. Brain Res. 2018, 348, 115–126. [Google Scholar] [CrossRef]
- Hikosaka, O. The Habenula: From Stress Evasion to Value-Based Decision-Making. Nat. Rev. Neurosci. 2010, 11, 503–513. [Google Scholar] [CrossRef]
- Sosa, R.; Mata-Luévanos, J.; Buenrostro-Jáuregui, M. The Role of the Lateral Habenula in Inhibitory Learning from Reward Omission. eNeuro 2021, 8, 0016-21. [Google Scholar] [CrossRef]
- Marzola, P.; Melzer, T.; Pavesi, E.; Gil-Mohapel, J.; Brocardo, P.S. Exploring the Role of Neuroplasticity in Development, Aging, and Neurodegeneration. Brain Sci. 2023, 13, 1610. [Google Scholar] [CrossRef] [PubMed]
- Puderbaugh, M.; Emmady, P.D. Neuroplasticity. Available online: https://www.ncbi.nlm.nih.gov/books/NBK557811/ (accessed on 9 March 2025).
- Seager, M.A.; Johnson, L.D.; Chabot, E.S.; Asaka, Y.; Berry, S.D.; Thompson, R.F. Oscillatory Brain States and Learning: Impact of Hippocampal Theta-Contingent Training. Proc. Natl. Acad. Sci. USA 2002, 99, 1616. [Google Scholar] [PubMed]
- Kempermann, G.; Jessberger, S.; Steiner, B.; Kronenberg, G. Milestones of Neuronal Development in the Adult Hippocampus. Trends Neurosci. 2004, 27, 447–452. [Google Scholar] [CrossRef]
- Leal-Galicia, P.; Saldívar-González, A.; Arias, C.; Morimoto, S. Exposure to Environmental Enrichment Elicits Differential Hippocampal Cell Proliferation: Role of Individual Responsiveness to Anxiety. Dev. Neurobiol. 2007, 67, 395–405. [Google Scholar] [CrossRef]
- Franklin, T.B.; Saab, B.J.; Mansuy, I.M. Neural Mechanisms of Stress Resilience and Vulnerability. Neuron 2012, 75, 747–761. [Google Scholar] [CrossRef]
- Cameron, H.A.; Mcewen, B.S.; Gould, E. Regulation of Adult Neurogenesis by Excitatory Input and NMDA Receptor Activation in the Dentate Gyrus. J. Neurosci. 1995, 75, 46874692. [Google Scholar]
- Yu, S.; Patchev, A.V.; Wu, Y.; Lu, J.; Holsboer, F.; Zhang, J.Z.; Sousa, N.; Almeida, O.F.X. Depletion of the Neural Precursor Cell Pool by Glucocorticoids. Ann. Neurol. 2010, 67, 21–30. [Google Scholar] [CrossRef]
- Leal-galicia, P.; Chávez-hernández, M.E.; Mata, F.; Mata-luévanos, J.; Rodríguez-serrano, L.M.; Tapia-de-jesús, A.; Buenrostro-jáuregui, M.H. Adult Neurogenesis: A Story Ranging from Controversial New Neurogenic Areas and Human Adult Neurogenesis to Molecular Regulation. Int. J. Mol. Sci. 2021, 22, 11489. [Google Scholar] [CrossRef]
- Fournier, N.M.; Duman, R.S. Role of Vascular Endothelial Growth Factor in Adult Hippocampal Neurogenesis: Implications for the Pathophysiology and Treatment of Depression. Behav. Brain Res. 2012, 227, 440–449. [Google Scholar] [CrossRef]
- Lee, E.; Son, H. Adult Hippocampal Neurogenesis and Related Neurotrophic Factors. BMB Rep. 2009, 42, 239–244. [Google Scholar] [PubMed]
- Castrén, E.; Rantamäki, T. The Role of BDNF and Its Receptors in Depression and Antidepressant Drug Action: Reactivation of Developmental Plasticity. Dev. Neurobiol. 2010, 70, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, F.F.; Ribeiro, F.F.; Rodrigues, R.S.; Sebastião, A.M.; Xapelli, S. Brain-Derived Neurotrophic Factor (BDNF) Role in Cannabinoid-Mediated Neurogenesis. Front. Cell. Neurosci. 2018, 12, 441. [Google Scholar] [CrossRef]
- Jin, K.; Zhu, Y.; Sun, Y.; Mao, X.O.; Xie, L.; Greenberg, D.A. Vascular Endothelial Growth Factor (VEGF) Stimulates Neurogenesis in Vitro and in Vivo. Proc. Natl. Acad. Sci. USA 2002, 99, 11946–11950. [Google Scholar]
- O’kusky, J.R.; Ye, P.; Joseph D’ercole, A. Insulin-Like Growth Factor-I Promotes Neurogenesis and Synaptogenesis in the Hippocampal Dentate Gyrus during Postnatal Development. J. Neurosci. 2000, 20, 8435–8442. [Google Scholar]
- Drake, C.T.; Chavkin, C.; Milner, T.A. Opioid Systems in the Dentate Gyrus. Prog. Brain Res. 2007, 163, 245–814. [Google Scholar] [CrossRef]
- Lutz, P.E.; Kieffer, B.L. Opioid Receptors: Distinct Roles in Mood Disorders. Trends Neurosci. 2013, 36, 195–206. [Google Scholar] [CrossRef]
- Sierra, A.; Encinas, J.M.; Maletic-Savatic, M. Adult Human Neurogenesis: From Microscopy to Magnetic Resonance Imaging. Front. Neurosci. 2011, 5, 9725. [Google Scholar] [CrossRef]
- Sutherland, R.J. The Dorsal Diencephalic Conduction System: A Review of the Anatomy and Functions of the Habenular Complex. Neurosci. Biobehav. Rev. 1982, 6, 1–13. [Google Scholar] [CrossRef]
- Rodríguez-Barrera, R.; Ibarra, A.; Rivas-González, M.; García-Sánchez, J.; Mojica-Torres, D. Neurogenesis after Spinal Cord Injury: State of the Art. Cells 2021, 10, 1499. [Google Scholar] [CrossRef]
- Martins-Macedo, J.; Salgado, A.J.; Gomes, E.D.; Pinto, L. Adult Brain Cytogenesis in the Context of Mood Disorders: From Neurogenesis to the Emergent Role of Gliogenesis. Neurosci. Biobehav. Rev. 2021, 131, 411–428. [Google Scholar] [CrossRef] [PubMed]
- Qi, C.; Luo, L.D.; Feng, I.; Ma, S. Molecular Mechanisms of Synaptogenesis. Front. Synaptic Neurosci. 2022, 14, 939793. [Google Scholar] [CrossRef] [PubMed]
- Faust, T.E.; Gunner, G.; Schafer, D.P. Mechanisms Governing Activity-Dependent Synaptic Pruning in the Developing Mammalian CNS. Nat. Rev. Neurosci. 2021, 22, 657–673. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dimension | Description | Refs. |
|---|---|---|
| General Definition | Multifaceted capacity of an organism or individual to adapt, recover, and/or grow after adversity. | [8,72,80] |
| Essential Components | Significant risk: exposure to major stressors threatening well-being. Positive adjustment: adaptive response and/or growth following adversity. | [5,72] |
| Perspectives of the Concept | Resilience as recovery: return to the preadversity state. Resilience as growth: surpassing the previous level of functioning. Resilience as an adaptive process: dynamic interactions between internal and external factors. | [8,79,80] |
| Modulating Factors | Neurobiological factors: efficient regulation of the HPA axis, synaptic plasticity, and allostatic responses. Genetic and epigenetic factors: gene expression is influenced by stress experiences. Environmental and social factors: social support, relationship quality, and resource availability. | [9,74,81] |
| Conceptual Differentiation | Invulnerability: absolute resistance (not synonymous with resilience). Stress resistance: coping without necessarily improving. Mental toughness: psychological competency in facing challenges. | [79,80] |
| Bidimensional Aspect | Significant risk: magnitude of the adverse event. Positive adjustment: functional positive outcome in social or behavioral contexts. | [72,78] |
| Behavioral Indicators | Social avoidance, anhedonia, academic or social success, controlled emotional responses. | [78,81] |
| Measurement and Evaluation | Currently based on psychometric scales and subjective reports, with a need for objective methods (e.g., biomarkers). | [5,77] |
| Animal Models | Maternal separation Chronic social defeat stress Forced swimming tests | [76,81,82,83] |
| Challenges and Limitations | Lack of a unified definition. Limited standardization of evaluation methods. Inadequate use of the term in studies. | [5,74] |
| Gene–Environment Interaction | The social environment and genetic predisposition work together to shape resilience responses. | [84,85] |
| Future Applications | Development of personalized interventions to foster resilience. Creation of biomarkers for an objective evaluation. Inclusion of interdisciplinary approaches in its study. | [74,78] |
| Brain Structure | Synthesized Functions | Refs. |
|---|---|---|
| Reward System | Controls motivation and pleasure, activated by natural rewards and drugs of abuse. Plasticity in the VTA and NAc modulates reward and counteracts anhedonia from stress. | [103,104,105] |
| Ventral Tegmental Area (VTA) | Origin of dopamine for reward and motivation. Projects to limbic and cortical regions. Its volume correlates with post-stress social avoidance. | [103,104,105,112] |
| Nucleus Accumbens (NAc) | Key for reward and motivation, receives dopamine signals from the VTA. Its plasticity counteracts anhedonia. Its volume is inversely related to social avoidance. β-Catenin promotes stress resilience. | [121,122] |
| Amygdala | Processes emotional responses and stimulus valence. Greater connectivity with the PFC in resilient individuals (better emotional regulation). Its activity increases in chronic stress susceptibility. Dendritic changes due to chronic stress affect fear learning. | [6,40,120,121] |
| Hippocampus | Involved in memory and navigation. Its plasticity is related to reward. Its activity is suppressed in chronic stress susceptibility. Its volume correlates with social avoidance. Early stress alters its structure and function (memory, maternal behavior, reduced neurogenesis). Adult neurogenesis influences anxiety and resilience, which is regulated by neurotrophic factors and social experiences. Chronic juvenile stress causes atrophy. | [6,42,120,121,124,125,126,127,128,129,130,131] |
| Prefrontal Cortex (PFC) | Involved in executive functions and planning. Greater connectivity with the amygdala is observed in resilient individuals (better emotional regulation). It receives dopamine signals. Its activity is suppressed in individuals with chronic stress resilience and increased in susceptible individuals in specific areas. Its volume is related to social avoidance. Chronic juvenile stress causes atrophy. | [6,40,113,120,121] |
| Habenula | Processes negative feedback and aversion. Its volume correlates with social avoidance. BDNF influences cell proliferation in this region. Neurogenesis in this area might buffer stress responses. | [121,132,133,134,135,136] |
| Hypothalamus | Regulates the stress response and homeostasis. Its volume correlates with social avoidance. The hippocampus relates to it differently in stress resilience and susceptibility. BDNF may promote neurogenesis in this area. | [121,132] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buenrostro-Jáuregui, M.H.; Muñóz-Sánchez, S.; Rojas-Hernández, J.; Alonso-Orozco, A.I.; Vega-Flores, G.; Tapia-de-Jesús, A.; Leal-Galicia, P. A Comprehensive Overview of Stress, Resilience, and Neuroplasticity Mechanisms. Int. J. Mol. Sci. 2025, 26, 3028. https://doi.org/10.3390/ijms26073028
Buenrostro-Jáuregui MH, Muñóz-Sánchez S, Rojas-Hernández J, Alonso-Orozco AI, Vega-Flores G, Tapia-de-Jesús A, Leal-Galicia P. A Comprehensive Overview of Stress, Resilience, and Neuroplasticity Mechanisms. International Journal of Molecular Sciences. 2025; 26(7):3028. https://doi.org/10.3390/ijms26073028
Chicago/Turabian StyleBuenrostro-Jáuregui, Mario Humberto, Sinuhé Muñóz-Sánchez, Jorge Rojas-Hernández, Adriana Ixel Alonso-Orozco, German Vega-Flores, Alejandro Tapia-de-Jesús, and Perla Leal-Galicia. 2025. "A Comprehensive Overview of Stress, Resilience, and Neuroplasticity Mechanisms" International Journal of Molecular Sciences 26, no. 7: 3028. https://doi.org/10.3390/ijms26073028
APA StyleBuenrostro-Jáuregui, M. H., Muñóz-Sánchez, S., Rojas-Hernández, J., Alonso-Orozco, A. I., Vega-Flores, G., Tapia-de-Jesús, A., & Leal-Galicia, P. (2025). A Comprehensive Overview of Stress, Resilience, and Neuroplasticity Mechanisms. International Journal of Molecular Sciences, 26(7), 3028. https://doi.org/10.3390/ijms26073028