

Protective Effect of Melatonin Against Bisphenol A Toxicity


Abstract
1. Introduction
1.1. Overview of BPA Toxicity
1.2. Overview of Melatonin and Its Potential Protective Effects
1.3. Purpose
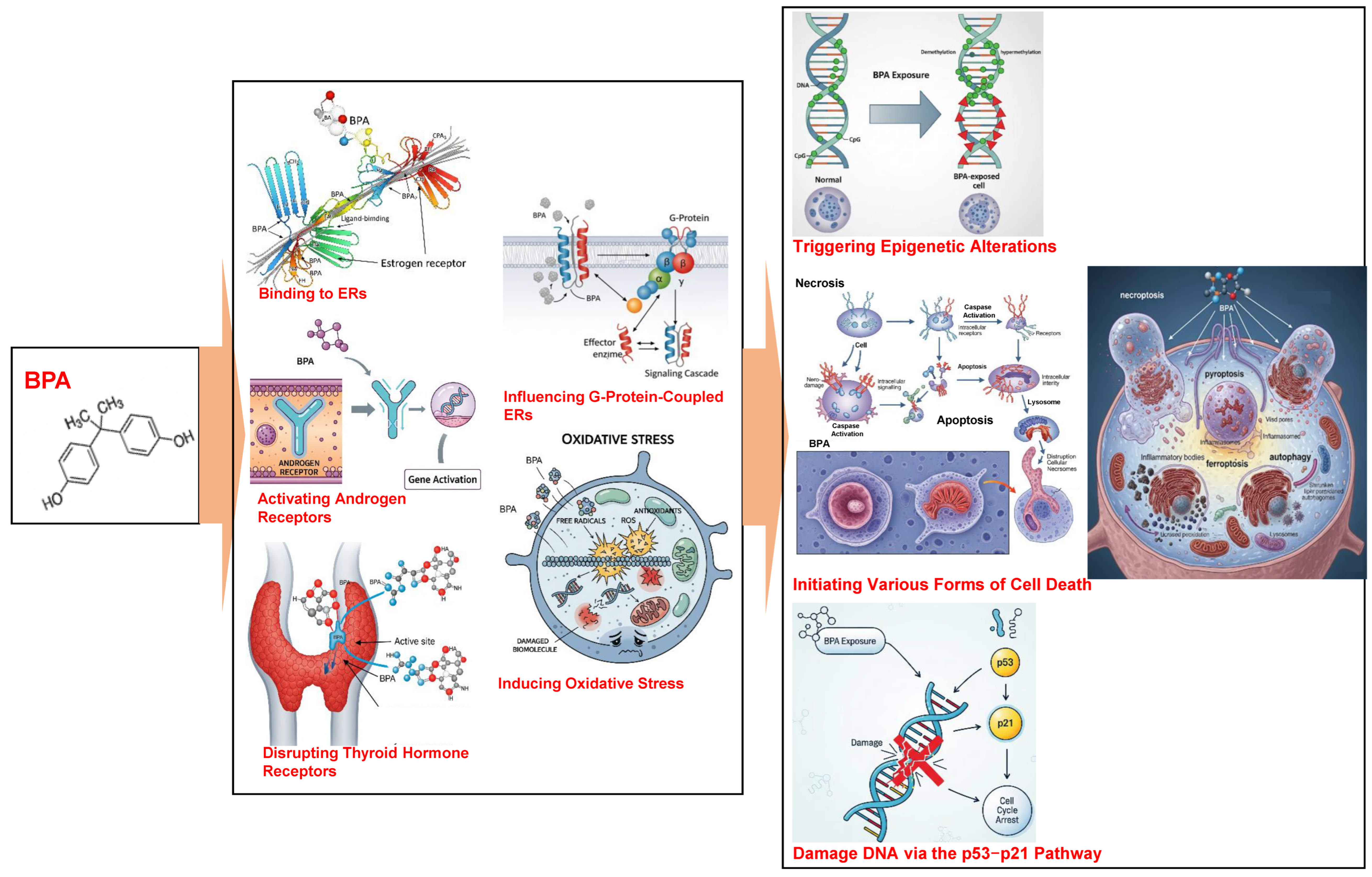
2. BPA Toxicity
2.1. Sources of BPA Exposure
2.2. Mechanisms of BPA Toxicity
2.3. Effects of BPA Exposure on Health
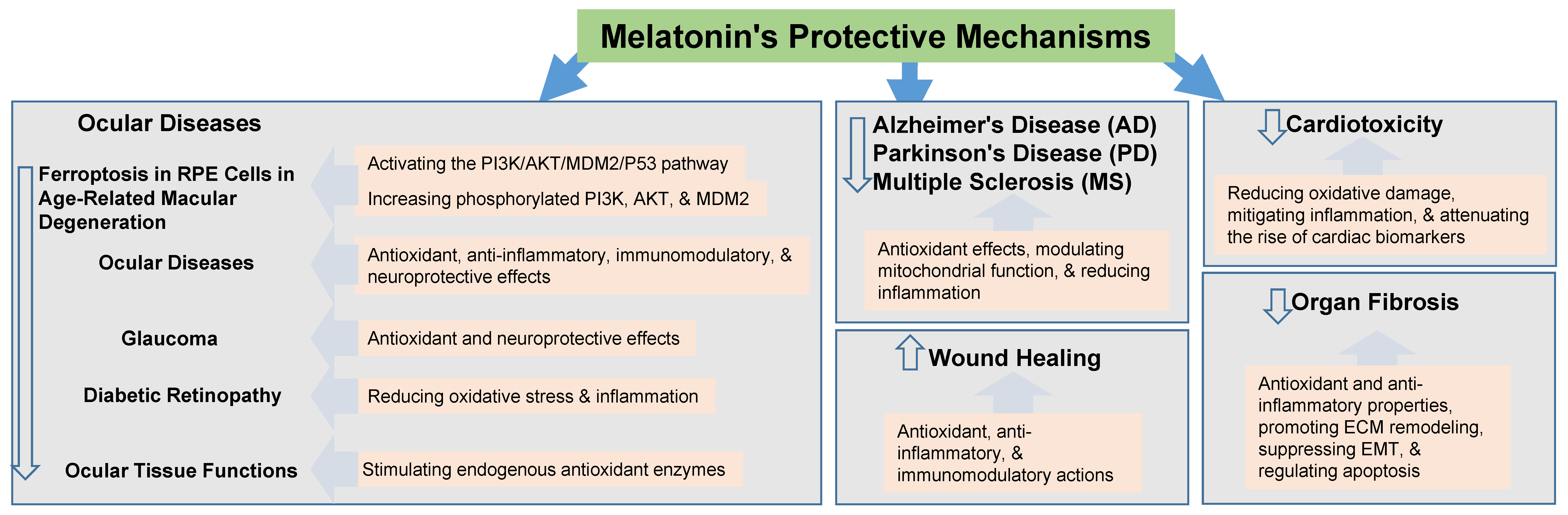
3. Melatonin and Its Protective Effects
3.1. What Is Melatonin?
3.2. Antioxidant Properties of Melatonin
3.3. Anti-Inflammatory Properties of Melatonin
3.4. Other Mechanisms Underlying Protective Effects of Melatonin
4. Evidence of Protective Effects of Melatonin Against BPA Toxicity
4.1. In Vitro Studies
4.2. Animal and Preclinical Studies
5. Potential Applications of Melatonin for BPA-Induced Toxicity
5.1. Melatonin Dosage and Timing in Animal Models
5.2. Safety Considerations in Human
6. Comprehensive Research Framework
6.1. Summary of Findings
6.2. Implications for Public Health and Policy
6.3. Future Research Directions
6.3.1. Modulation of Specific Reproductive Signaling Pathways
6.3.2. Inhibition of ER Stress Pathways
6.3.3. Regulation of Autophagy via Transcription Factor EB (TFEB) and p38 MAPK Pathway
6.3.4. Inhibition of Ferroptosis Through KEAP1/NRF2/PTGS2 Axis and Ferritinophagy
6.3.5. Regulation of Mitochondrial Dynamics and Quality Control via SIRT1/PGC-1α Pathway
6.3.6. New Methods in Future Research
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tsai, W.T. Human health risk on environmental exposure to Bisphenol-A: A review. J. Environ. Sci. Health Part C Environ. Carcinog. Ecotoxicol. Rev. 2006, 24, 225–255. [Google Scholar] [CrossRef] [PubMed]
- Rubin, B.S. Bisphenol A: An endocrine disruptor with widespread exposure and multiple effects. J. Steroid Biochem. Mol. Biol. 2011, 127, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Coppola, L.; La Rocca, C. Molecular Mechanisms of Bisphenol A Toxicity and Effects of Environmental Levels on Health. Int. J. Mol. Sci. 2023, 24, 8028. [Google Scholar] [CrossRef] [PubMed]
- Yukta, S.; Milan, K.L.; Harithpriya, K.; Zong, C.; Sahabudeen, S.; Ichihara, G.; Ramkumar, K.M. Disruptive multiple cell death pathways of bisphenol-A. Toxicol. Mech. Methods 2025, 35, 430–443. [Google Scholar] [CrossRef]
- EFSA Panel on Food Contact Materials, Enzymes and Processing Aids (CEP); Lambré, C.; Barat Baviera, J.M.; Bolognesi, C.; Chesson, A.; Cocconcelli, P.S.; Crebelli, R.; Gott, D.M.; Grob, K.; Lampi, E.; et al. Re-evaluation of the risks to public health related to the presence of bisphenol A (BPA) in foodstuffs. EFSA J. 2023, 21, e06857. [Google Scholar] [CrossRef]
- Mhaouty-Kodja, S.; Zalko, D.; Tait, S.; Testai, E.; Viguié, C.; Corsini, E.; Grova, N.; Buratti, F.M.; Cabaton, N.J.; Coppola, L.; et al. A critical review to identify data gaps and improve risk assessment of bisphenol A alternatives for human health. Crit. Rev. Toxicol. 2024, 54, 696–753. [Google Scholar] [CrossRef]
- Huang, W.; Zheng, J.; Wang, M.; Du, L.Y.; Bai, L.; Tang, H. The potential therapeutic role of melatonin in organ fibrosis: A comprehensive review. Front. Med. 2024, 11, 1502368. [Google Scholar] [CrossRef]
- Sieminski, M.; Reimus, M.; Kałas, M.; Stępniewska, E. Antioxidant and Anti-Inflammatory Properties of Melatonin in Secondary Traumatic Brain Injury. Antioxidants 2024, 14, 25. [Google Scholar] [CrossRef]
- Ebrahimi, R.; Shokrzadeh, M.; Ghassemi Barghi, N. Effects of melatonin on the Bisphenol-A- induced cytotoxicity and genetic toxicity in colon cancer cell lines, normal gingival cell lines, and bone marrow stem cell lines. Cancer Inform. 2021, 20, 11769351211056295. [Google Scholar] [CrossRef]
- Slominski, A.T.; Kim, T.K.; Janjetovic, Z.; Slominski, R.M.; Ganguli-Indra, G.; Athar, M.; Indra, A.K.; Reiter, R.J.; Kleszczyński, K. Melatonin and the Skin: Current Progress and Perspectives for Human Health. J. Invest. Dermatol. 2025, 145, 1345–1360.e2. [Google Scholar] [CrossRef]
- Kim, T.K.; Slominski, R.M.; Pyza, E.; Kleszczynski, K.; Tuckey, R.C.; Reiter, R.J.; Holick, M.F.; Slominski, A.T. Evolutionary formation of melatonin and vitamin D in early life forms: Insects take centre stage. Biol. Rev. Camb. Philos. Soc. 2024, 99, 1772–1790. [Google Scholar] [CrossRef]
- Slominski, A.T.; Semak, I.; Fischer, T.W.; Kim, T.K.; Kleszczyński, K.; Hardeland, R.; Reiter, R.J. Metabolism of melatonin in the skin: Why is it important? Exp. Dermatol. 2017, 26, 563–568. [Google Scholar] [CrossRef]
- Kim, T.K.; Kleszczynski, K.; Janjetovic, Z.; Sweatman, T.; Lin, Z.; Li, W.; Reiter, R.J.; Fischer, T.W.; Slominski, A.T. Metabolism of melatonin and biological activity of intermediates of melatoninergic pathway in human skin cells. FASEB J. 2013, 27, 2742–2755. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Chen, D.; He, Y.; Zhu, W.; Zhou, W.; Zhang, J. Bisphenol-A and Female Infertility: A Possible Role of Gene-Environment Interactions. Int. J. Environ. Res. Public Health 2015, 12, 11101–11116. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhou, Z.; Qing, D.; He, Y.; Wu, T.; Miao, M.; Wang, J.; Weng, X.; Ferber, J.R.; Herrinton, L.J.; et al. Occupational exposure to bisphenol-A (BPA) and the risk of self-reported male sexual dysfunction. Hum. Reprod. 2010, 25, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Machtinger, R.; Combelles, C.M.; Missmer, S.A.; Correia, K.F.; Williams, P.; Hauser, R.; Racowsky, C. Bisphenol-A and human oocyte maturation in vitro. Hum. Reprod. 2013, 28, 2735–2745. [Google Scholar] [CrossRef]
- Acconcia, F.; Pallottini, V.; Marino, M. Molecular Mechanisms of Action of BPA. Dose Response 2015, 13, 1559325815610582. [Google Scholar] [CrossRef]
- La Merrill, M.A.; Vandenberg, L.N.; Smith, M.T.; Goodson, W.; Browne, P.; Patisaul, H.B.; Guyton, K.Z.; Kortenkamp, A.; Cogliano, V.J.; Woodruff, T.J.; et al. Consensus on the key characteristics of endocrine-disrupting chemicals as a basis for hazard identification. Nat. Rev. Endocrinol. 2020, 16, 45–57. [Google Scholar] [CrossRef]
- Pathak, R.K.; Jung, D.W.; Shin, S.H.; Ryu, B.Y.; Lee, H.S.; Kim, J.M. Deciphering the mechanisms and interactions of the endocrine disruptor bisphenol A and its analogs with the androgen receptor. J. Hazard. Mater. 2024, 469, 133935. [Google Scholar] [CrossRef]
- Guo, J.; Zhao, M.H.; Shin, K.T.; Niu, Y.J.; Ahn, Y.D.; Kim, N.H.; Cui, X.S. The possible molecular mechanisms of bisphenol A action on porcine early embryonic development. Sci. Rep. 2017, 7, 8632. [Google Scholar] [CrossRef]
- Rifa, R.A.; Rojo, M.G.; Lavado, R. Mechanisms of toxicity caused by bisphenol analogs in human in vitro cell models. Chem. Biol. Interact. 2025, 412, 111475. [Google Scholar] [CrossRef]
- Manzoor, M.F.; Tariq, T.; Fatima, B.; Sahar, A.; Tariq, F.; Munir, S.; Khan, S.; Nawaz Ranjha, M.M.A.; Sameen, A.; Zeng, X.A.; et al. An insight into bisphenol A, food exposure and its adverse effects on health: A review. Front. Nutr. 2022, 9, 1047827. [Google Scholar] [CrossRef]
- Fonseca, M.I.; Lorigo, M.; Cairrao, E. Endocrine-Disrupting Effects of Bisphenol A on the Cardiovascular System: A Review. J. Xenobiot. 2022, 12, 181–213. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Kim, J.H.; Lim, Y.H.; Park, H.Y.; Hong, Y.C. Associations of bisphenol A exposure with heart rate variability and blood pressure. Hypertension 2012, 60, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Cho, Y.H.; Hong, Y.C. MicroRNA expression in response to bisphenol A is associated with high blood pressure. Environ. Int. 2020, 141, 105791. [Google Scholar] [CrossRef] [PubMed]
- Lakind, J.S.; Goodman, M.; Mattison, D.R. Bisphenol A and indicators of obesity, glucose metabolism/type 2 diabetes and cardiovascular disease: A systematic review of epidemiologic research. Crit. Rev. Toxicol. 2014, 44, 121–150. [Google Scholar] [CrossRef]
- Legeay, S.; Faure, S. Is bisphenol A an environmental obesogen? Fundam. Clin. Pharmacol. 2017, 31, 594–609. [Google Scholar] [CrossRef]
- Liu, B.; Lehmler, H.J.; Sun, Y.; Xu, G.; Liu, Y.; Zong, G.; Sun, Q.; Hu, F.B.; Wallace, R.B.; Bao, W. Bisphenol A substitutes and obesity in US adults: Analysis of a population-based, cross-sectional study. Lancet Planet. Health 2017, 1, e114–e122. [Google Scholar] [CrossRef]
- Minatoya, M.; Sasaki, S.; Araki, A.; Miyashita, C.; Itoh, S.; Yamamoto, J.; Matsumura, T.; Mitsui, T.; Moriya, K.; Cho, K.; et al. Cord Blood Bisphenol A Levels and Reproductive and Thyroid Hormone Levels of Neonates: The Hokkaido Study on Environment and Children’s Health. Epidemiology 2017, 28 (Suppl. S1), S3–S9. [Google Scholar] [CrossRef]
- Cantonwine, D.E.; Hauser, R.; Meeker, J.D. Bisphenol A and Human Reproductive Health. Expert. Rev. Obstet. Gynecol. 2013, 8, 329–335. [Google Scholar] [CrossRef]
- Jašarević, E.; Sieli, P.T.; Twellman, E.E.; Welsh, T.H., Jr.; Schachtman, T.R.; Roberts, R.M.; Geary, D.C.; Rosenfeld, C.S. Disruption of adult expression of sexually selected traits by developmental exposure to bisphenol A. Proc. Natl. Acad. Sci. USA 2011, 108, 11715–11720. [Google Scholar] [CrossRef]
- Covaci, A.; Den Hond, E.; Geens, T.; Govarts, E.; Koppen, G.; Frederiksen, H.; Knudsen, L.E.; Mørck, T.A.; Gutleb, A.C.; Guignard, C.; et al. Urinary BPA measurements in children and mothers from six European member states: Overall results and determinants of exposure. Environ. Res. 2015, 141, 77–85. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Maffini, M.V.; Sonnenschein, C.; Rubin, B.S.; Soto, A.M. Bisphenol-A and the great divide: A review of controversies in the field of endocrine disruption. Endocr. Rev. 2009, 30, 75–95. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Chahoud, I.; Heindel, J.J.; Padmanabhan, V.; Paumgartten, F.J.; Schoenfelder, G. Urinary, circulating, and tissue biomonitoring studies indicate widespread exposure to bisphenol A. Environ. Health Perspect. 2010, 118, 1055–1070. [Google Scholar] [CrossRef]
- Wang, X.; Nag, R.; Brunton, N.P.; Siddique, M.A.B.; Harrison, S.M.; Monahan, F.J.; Cummins, E. Human health risk assessment of bisphenol A (BPA) through meat products. Environ. Res. 2022, 213, 113734. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, S.M.; House, C.D. Bisphenol A and DDT disrupt adipocyte function in the mammary gland: Implications for breast cancer risk and progression. Front. Oncol. 2025, 15, 1490898. [Google Scholar] [CrossRef] [PubMed]
- Stillwater, B.J.; Bull, A.C.; Romagnolo, D.F.; Neumayer, L.A.; Donovan, M.G.; Selmin, O.I. Bisphenols and Risk of Breast Cancer: A Narrative Review of the Impact of Diet and Bioactive Food Components. Front. Nutr. 2020, 7, 581388. [Google Scholar] [CrossRef]
- Dong, Z.; He, L.; Wu, J.; Xie, C.; Geng, S.; Wu, J.; Zhong, C.; Li, X. Bisphenol A-induced cancer-associated adipocytes promotes breast carcinogenesis via CXCL12/AKT signaling. Mol. Cell. Endocrinol. 2025, 599, 112473. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wei, C.; Liu, X.; Zhang, Z.; Wu, Y.; Zeng, B.; Jin, Y.; Shi, Y.; Mo, Z.; Cheng, J.; et al. Revealing the role of bisphenol A on prostate cancer progression and identifying potential targets: A comprehensive analysis from population cohort to molecular mechanism. Ecotoxicol. Environ. Saf. 2025, 296, 118209. [Google Scholar] [CrossRef]
- Xu, S.; Yuan, X.; Wang, Y.; Fu, Z.; Chen, K.; Cui, Z.; Xu, L.; Zhang, H.; Xia, D.; Wu, Y. Bisphenols exposure at environmentally relevant dose promoted ovarian cancer progression and modulated tumor microenvironment through β-catenin/SPP1 axis. J. Hazard. Mater. 2025, 490, 137824. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Rajaura, S.; Singh, A.; Rambabu; Nivedita; Ahmed, M.Z. Bisphenol-A-induced ovarian cancer: Changes in epithelial diversity, apoptosis, antioxidant and anti-inflammatory mechanisms. Reprod. Toxicol. 2025, 135, 108909. [Google Scholar] [CrossRef]
- Rajaura, S.; Bhardwaj, N.; Singh, A.; Babu, R.; Gupta, N.; Ahmed, M.Z. Bisphenol A-induced oxidative stress increases the production of ovarian cancer stem cells in mice. Reprod. Toxicol. 2024, 130, 108724. [Google Scholar] [CrossRef]
- Leung, Y.K.; Biesiada, J.; Govindarajah, V.; Ying, J.; Kendler, A.; Medvedovic, M.; Ho, S.M. Low-Dose Bisphenol A in a Rat Model of Endometrial Cancer: A CLARITY-BPA Study. Environ. Health Perspect. 2020, 128, 127005. [Google Scholar] [CrossRef]
- Hong, X.; Wang, S.; Zhang, Q.; Li, L.; Liu, H.; Yang, H.; Wu, D.; Liu, X.; Shen, T. Bisphenol A exacerbates colorectal cancer progression through enhancing ceramide synthesis. Toxicology 2025, 511, 154054. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.F.S.; Abdullah, M.N.H.; Lim, V.; Bahari, H.; Khoo, B.Y.; Tan, J.J.; Yong, Y.K. Exploring the role of Bisphenol A in obesity-driven colorectal cancer progression: Network toxicology and multi-organ pathology in animal models. Toxicol. Appl. Pharmacol. 2025, 495, 117227. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chen, S.; Li, Y.; Qiu, Y.; Chen, X.; Wu, Y.; Deng, X.; Chen, M.; Wang, C.; Hong, Z.; et al. Upregulation of GOLPH3 mediated by Bisphenol a promotes colorectal cancer proliferation and migration: Evidence based on integrated analysis. Front. Pharmacol. 2024, 15, 1337883. [Google Scholar] [CrossRef] [PubMed]
- Nair, V.A.; Malhab, L.J.B.; Abdel-Rahman, W.M. Characterization of the Molecular Alterations Induced by the Prolonged Exposure of Normal Colon Mucosa and Colon Cancer Cells to Low-Dose Bisphenol A. Int. J. Mol. Sci. 2022, 23, 11620. [Google Scholar] [CrossRef]
- Wang, Y.; Su, X.; Wang, Q.; Zhang, L.; Yu, Y.; Zhao, Y.; Liu, Z. Bisphenol A exposure enhances proliferation and tumorigenesis of papillary thyroid carcinoma through ROS generation and activation of NOX4 signaling pathways. Ecotoxicol. Environ. Saf. 2025, 292, 117946. [Google Scholar] [CrossRef]
- Wiszpolska, M.; Lepiarczyk, E.; Maździarz, M.A.; Paukszto, Ł.; Makowczenko, K.G.; Lipka, A.; Łopieńska-Biernat, E.; Makowska, K.; Gonkowski, S.; Correia-de-Sá, P.; et al. The Carcinogenic Potential of Bisphenol A in the Liver Based on Transcriptomic Studies. Cancers 2023, 15, 5014. [Google Scholar] [CrossRef]
- Ismael, L.Q.; Abdulhameed, A.R.; Keong, Y.Y.; Abdullah, M.N.H.; Bahari, H.; Jie, T.J.; Yin, K.B. Bisphenol A is a carcinogen that induces lipid accumulation, peroxisome proliferator-activated receptor-γ expression and liver disease. Exp. Ther. Med. 2022, 24, 735. [Google Scholar] [CrossRef]
- Ebihara, S.; Marks, T.; Hudson, D.J.; Menaker, M. Genetic control of melatonin synthesis in the pineal gland of the mouse. Science 1986, 231, 491–493. [Google Scholar] [CrossRef]
- Klein, D.C.; Coon, S.L.; Roseboom, P.H.; Weller, J.L.; Bernard, M.; Gastel, J.A.; Zatz, M.; Iuvone, P.M.; Rodriguez, I.R.; Bégay, V.; et al. The melatonin rhythm-generating enzyme: Molecular regulation of serotonin N-acetyltransferase in the pineal gland. Recent. Prog. Horm. Res. 1997, 52, 307–357. [Google Scholar]
- Reiter, R.J.; Tan, D.X.; Fuentes-Broto, L. Melatonin: A multitasking molecule. Prog Brain Res. 2010, 181, 127–151. [Google Scholar] [CrossRef]
- Comai, S.; Gobbi, G. Melatonin, Melatonin Receptors and Sleep: Moving Beyond Traditional Views. J. Pineal Res. 2024, 76, e13011. [Google Scholar] [CrossRef]
- Vanecek, J. Cellular mechanisms of melatonin action. Physiol. Rev. 1998, 78, 687–721. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.B.; Ali, A.; Bilal, M.; Rashid, S.M.; Wani, A.B.; Bhat, R.R.; Rehman, M.U. Melatonin and Health: Insights of Melatonin Action, Biological Functions, and Associated Disorders. Cell. Mol. Neurobiol. 2023, 43, 2437–2458. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; Srinivasan, V.; Maestroni, G.J.; Cardinali, D.P.; Poeggeler, B.; Hardeland, R. Melatonin: Nature’s most versatile biological signal? FEBS J. 2006, 273, 2813–2838. [Google Scholar] [CrossRef] [PubMed]
- Chitimus, D.M.; Popescu, M.R.; Voiculescu, S.E.; Panaitescu, A.M.; Pavel, B.; Zagrean, L.; Zagrean, A.M. Melatonin’s Impact on Antioxidative and Anti-Inflammatory Reprogramming in Homeostasis and Disease. Biomolecules 2020, 10, 1211. [Google Scholar] [CrossRef]
- Cho, J.H.; Bhutani, S.; Kim, C.H.; Irwin, M.R. Anti-inflammatory effects of melatonin: A systematic review and meta-analysis of clinical trials. Brain Behav. Immun. 2021, 93, 245–253. [Google Scholar] [CrossRef]
- Mauriz, J.L.; Collado, P.S.; Veneroso, C.; Reiter, R.J.; González-Gallego, J. A review of the molecular aspects of melatonin’s anti-inflammatory actions: Recent insights and new perspectives. J. Pineal Res. 2013, 54, 1–14. [Google Scholar] [CrossRef]
- Xia, M.Z.; Liang, Y.L.; Wang, H.; Chen, X.; Huang, Y.Y.; Zhang, Z.H.; Chen, Y.H.; Zhang, C.; Zhao, M.; Xu, D.X.; et al. Melatonin modulates TLR4-mediated inflammatory genes through MyD88- and TRIF-dependent signaling pathways in lipopolysaccharide-stimulated RAW264.7 cells. J. Pineal Res. 2012, 53, 325–334. [Google Scholar] [CrossRef]
- Gitto, E.; Reiter, R.J.; Amodio, A.; Romeo, C.; Cuzzocrea, E.; Sabatino, G.; Buonocore, G.; Cordaro, V.; Trimarchi, G.; Barberi, I. Early indicators of chronic lung disease in preterm infants with respiratory distress syndrome and their inhibition by melatonin. J. Pineal Res. 2004, 36, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Frungieri, M.B.; Calandra, R.S.; Matzkin, M.E.; Rossi, S.P. Melatonin as a natural anti-inflammatory and anti-oxidant therapy in the testis: A focus on infertility and aging. Biol. Reprod. 2024, 111, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.S.; Chung, S.H.; Lee, S.K.; Kim, J.H.; Kim, J.B.; Kim, T.K.; Kim, D.S.; Baik, H.W. Melatonin improves experimental colitis with sleep deprivation. Int. J. Mol. Med. 2015, 35, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Vaghari-Tabari, M.; Moein, S.; Alipourian, A.; Qujeq, D.; Malakoti, F.; Alemi, F.; Yousefi, B.; Khazaie, S. Melatonin and inflammatory bowel disease: From basic mechanisms to clinical application. Biochimie 2023, 209, 20–36. [Google Scholar] [CrossRef]
- MacDonald, I.J.; Huang, C.C.; Liu, S.C.; Tang, C.H. Reconsidering the Role of Melatonin in Rheumatoid Arthritis. Int. J. Mol. Sci. 2020, 21, 2877. [Google Scholar] [CrossRef]
- Konečná, B.; Chobodová, P.; Janko, J.; Baňasová, L.; Bábíčková, J.; Celec, P.; Tóthová, Ľ. The Effect of Melatonin on Periodontitis. Int. J. Mol. Sci. 2021, 22, 2390. [Google Scholar] [CrossRef]
- Tobeiha, M.; Jafari, A.; Fadaei, S.; Mirazimi, S.M.A.; Dashti, F.; Amiri, A.; Khan, H.; Asemi, Z.; Reiter, R.J.; Hamblin, M.R.; et al. Evidence for the Benefits of Melatonin in Cardiovascular Disease. Front. Cardiovasc. Med. 2022, 9, 888319. [Google Scholar] [CrossRef]
- Qi, Q.; Feng, L.; Liu, J.; Xu, D.; Wang, G.; Pan, X. Melatonin Alleviates BPA-Induced Testicular Apoptosis and Endoplasmic Reticulum Stress. Front. Biosci. 2024, 29, 95. [Google Scholar] [CrossRef]
- Sevastre-Berghian, A.C.; Casandra, C.; Gheban, D.; Olteanu, D.; Olanescu Vaida Voevod, M.C.; Rogojan, L.; Filip, G.A.; Bâldea, I. Neurotoxicity of Bisphenol A and the Impact of Melatonin Administration on Oxidative Stress, ERK/NF-kB Signaling Pathway, and Behavior in Rats. Neurotox. Res. 2022, 40, 1882–1894. [Google Scholar] [CrossRef]
- Ishtiaq, A.; Ali, T.; Bakhtiar, A.; Bibi, R.; Bibi, K.; Mushtaq, I.; Li, S.; Khan, W.; Khan, U.; Anis, R.A.; et al. Melatonin abated Bisphenol A-induced neurotoxicity via p53/PUMA/Drp-1 signaling. Environ. Sci. Pollut. Res. 2021, 28, 17789–17801. [Google Scholar] [CrossRef]
- Slominski, A.T.; Kim, T.K.; Slominski, R.M.; Song, Y.; Qayyum, S.; Placha, W.; Janjetovic, Z.; Kleszczyński, K.; Atigadda, V.; Song, Y.; et al. Melatonin and Its Metabolites Can Serve as Agonists on the Aryl Hydrocarbon Receptor and Peroxisome Proliferator-Activated Receptor Gamma. Int. J. Mol. Sci. 2023, 24, 15496. [Google Scholar] [CrossRef]
- Wu, P.; Zhao, L.; Du, Y.; Lu, J.; He, Y.; Shu, Q.; Peng, H.; Wang, X. Melatonin protects retinal pigment epithelium cells against ferroptosis in AMD via the PI3K/AKT/MDM2/P53 pathway. Front. Pharmacol. 2025, 16, 1543575. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Wang, Q.; Wu, W.; Zeng, W.; Feng, Y. Therapeutic Effects of Melatonin on Ocular Diseases: Knowledge Map and Perspective. Front. Pharmacol. 2021, 12, 721869. [Google Scholar] [CrossRef] [PubMed]
- Rusciano, D.; Russo, C. The Therapeutic Trip of Melatonin Eye Drops: From the Ocular Surface to the Retina. Pharmaceuticals 2024, 17, 441. [Google Scholar] [CrossRef] [PubMed]
- Galvani, F.; Cammarota, M.; Vacondio, F.; Rivara, S.; Boscia, F. Protective Activity of Melatonin Combinations and Melatonin-Based Hybrid Molecules in Neurodegenerative Diseases. J. Pineal Res. 2024, 76, e70008. [Google Scholar] [CrossRef]
- Amiri, M.; Khazaie, H.; Mohammadi, M. The protective effects of melatonin against electromagnetic waves of cell phones in animal models: A systematic review. Anim. Model. Exp. Med. 2025, 8, 629–637. [Google Scholar] [CrossRef]
- Sohn, E.H.; Kim, S.N.; Lee, S.R. Melatonin’s Impact on Wound Healing. Antioxidants 2024, 13, 1197. [Google Scholar] [CrossRef]
- Bocheva, G.; Slominski, R.M.; Janjetovic, Z.; Kim, T.K.; Böhm, M.; Steinbrink, K.; Reiter, R.J.; Kleszczyński, K.; Slominski, A.T. Protective Role of Melatonin and Its Metabolites in Skin Aging. Int. J. Mol. Sci. 2022, 23, 1238. [Google Scholar] [CrossRef]
- Samra, T.; Gomez-Gomez, T.; Linowiecka, K.; Akhundlu, A.; Lopez de Mendoza, G.; Gompels, M.; Lee, W.W.; Gherardini, J.; Chéret, J.; Paus, R. Melatonin Exerts Prominent, Differential Epidermal and Dermal Anti-Aging Properties in Aged Human Eyelid Skin Ex Vivo. Int. J. Mol. Sci. 2023, 24, 15963. [Google Scholar] [CrossRef]
- Mansoori, R.; Ashrafpour, M.; Asghari, M.H.; Golchoobian, R.; Hosseini, S.M.; Reiter, R.J.; Karim, B.; Moghadamnia, A.A.; Kazemi, S. Protective effects of melatonin against 5-fluorouracil-induced cardiotoxicity in rats: A comprehensive evaluation of oxidative, inflammatory, and apoptotic pathways. Toxicol. Appl. Pharmacol. 2025, 499, 117343. [Google Scholar] [CrossRef]
- Kobylińska, A.; Bernat, P.; Posmyk, M.M. Melatonin Mitigates Lead-Induced Oxidative Stress and Modifies Phospholipid Profile in Tobacco BY-2 Suspension Cells. Int. J. Mol. Sci. 2024, 25, 5064. [Google Scholar] [CrossRef]
- Fatemeh, G.; Sajjad, M.; Niloufar, R.; Neda, S.; Leila, S.; Khadijeh, M. Effect of melatonin supplementation on sleep quality: A systematic review and meta-analysis of randomized controlled trials. J. Neurol. 2022, 269, 205–216. [Google Scholar] [CrossRef]
- Peña-Corona, S.I.; Chávez-Corona, J.I.; Pérez-Caltzontzin, L.E.; Vargas-Estrada, D.; Mendoza-Rodríguez, C.A.; Ramos-Martínez, E.; Cerbón-Gutiérrez, J.L.; Herrera-Barragán, J.A.; Quintanar-Guerrero, D.; Leyva-Gómez, G. Melatonin and Vitamins as Protectors against the Reproductive Toxicity of Bisphenols: Which Is the Most Effective? A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2023, 24, 14930. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, N.D.; Parsa, S.; Nozari, F.; Shahlaee, M.A.; Maktabi, A.; Sayadi, M.; Sadeghi, A.; Azarpira, N. Protective effects of melatonin against the toxic effects of environmental pollutants and heavy metals on testicular tissue: A systematic review and meta-analysis of animal studies. Front. Endocrinol. 2023, 14, 1119553. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, K.K.A.C.; Shiroma, M.E.; Damous, L.L.; Simões, M.J.; Simões, R.D.S.; Cipolla-Neto, J.; Baracat, E.C.; Soares, J.M., Jr. Antioxidant Actions of Melatonin: A Systematic Review of Animal Studies. Antioxidants 2024, 13, 439. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, S.M.; Rashid, H. Melatonin Ameliorates BPA Induced Oxidative Stress in Human Red Blood Cells: An In vitro Study. Endocr. Metab. Immune Disord. Drug Targets 2020, 20, 1321–1327. [Google Scholar] [CrossRef]
- Song, D.; Liu, Y.; Yao, Y.; Liu, F.; Tao, W.; Zhou, X.; Li, R.; Zhang, X.; Li, X. Melatonin improves bisphenol A-induced cell apoptosis, oxidative stress and autophagy impairment via inhibition of the p38 MAPK signaling pathway in FLK-BLV cells. Environ. Toxicol. 2022, 37, 1551–1562. [Google Scholar] [CrossRef]
- Park, H.J.; Park, S.Y.; Kim, J.W.; Yang, S.G.; Kim, M.J.; Jegal, H.G.; Kim, I.S.; Choo, Y.K.; Koo, D.B. Melatonin Improves Oocyte Maturation and Mitochondrial Functions by Reducing Bisphenol A-Derived Superoxide in Porcine Oocytes In Vitro. Int. J. Mol. Sci. 2018, 19, 3422. [Google Scholar] [CrossRef]
- Akarca-Dizakar, S.Ö.; Erdoğan, D.; Peker, T.; Coşkun Akçay, N.; Türkoğlu, I.; Eşmekaya, M.A.; Ömeroğlu, S. Effects of co-administered melatonin, fructose and bisphenol A (BPA) on rat epididymis and sperm characteristics. Biotech. Histochem. 2020, 95, 18–26. [Google Scholar] [CrossRef]
- Mi, P.; Zhang, Q.P.; Li, S.B.; Liu, X.Y.; Zhang, S.H.; Li, M.; Chen, D.Y.; Zhao, X.; Feng, D.F.; Feng, X.Z. Melatonin protects embryonic development and maintains sleep/wake behaviors from the deleterious effects of fluorene-9-bisphenol in zebrafish (Danio rerio). J. Pineal Res. 2019, 66, e12530. [Google Scholar] [CrossRef]
- Amjad, S.; Rahman, M.S.; Pang, M.G. Role of Antioxidants in Alleviating Bisphenol A Toxicity. Biomolecules 2020, 10, 1105. [Google Scholar] [CrossRef]
- Qi, Q.; Yang, J.; Li, S.; Liu, J.; Xu, D.; Wang, G.; Feng, L.; Pan, X. Melatonin alleviates oxidative stress damage in mouse testes induced by bisphenol A. Front. Cell Dev. Biol. 2024, 12, 1338828. [Google Scholar] [CrossRef] [PubMed]
- Kobroob, A.; Peerapanyasut, W.; Chattipakorn, N.; Wongmekiat, O. Damaging Effects of Bisphenol A on the Kidney and the Protection by Melatonin: Emerging Evidences from In Vivo and In Vitro Studies. Oxidative Med. Cell. Longev. 2018, 2018, 3082438. [Google Scholar] [CrossRef] [PubMed]
- Bameri, B.; Armandeh, M.; Haghi-Aminjan, H.; Rahimifard, M.; Hassani, S.; Gholami, M.; Ataei, M.; Samadi, M.; Daniali, M.; Hooshangi Shayesteh, M.R.; et al. Insights into the role of melatonin on the reproductive system toxicity induced by bisphenol A: A systematic review of preclinical studies. Toxin Rev. 2024, 43, 137–156. [Google Scholar] [CrossRef]
- Kadir, E.R.; Yakub, A.D.; Imam, A.; Olajide, O.J.; Lawal, I.A.; Oyewole, A.L.; Hussein, M.O.; Ojulari, L.S.; Ajao, M.S. Restorative Effects of Melatonin on Bisphenol A-induced Interference of Gene Expression in Hypothalamic Pituitary Axis Following Early Exposure. Int. J. Health Sci. 2024, 8, 1293–1315. [Google Scholar] [CrossRef]
- El-Missiry, M.A.; Othman, A.I.; Al-Abdan, M.A.; El-Sayed, A.A. Melatonin ameliorates oxidative stress, modulates death receptor pathway proteins, and protects the rat cerebrum against bisphenol-A-induced apoptosis. J. Neurol. Sci. 2014, 347, 251–256. [Google Scholar] [CrossRef]
- Olukole, S.G.; Ajani, S.O.; Ola-Davies, E.O.; Lanipekun, D.O.; Aina, O.O.; Oyeyemi, M.O.; Oke, B.O. Melatonin ameliorates bisphenol A-induced perturbations of the prostate gland of adult Wistar rats. Biomed. Pharmacother. 2018, 105, 73–82. [Google Scholar] [CrossRef]
- Kanwar, M.K.; Xie, D.; Yang, C.; Ahammed, G.J.; Qi, Z.; Hasan, M.K.; Reiter, R.J.; Yu, J.Q.; Zhou, J. Melatonin promotes metabolism of bisphenol A by enhancing glutathione-dependent detoxification in Solanum lycopersicum L. J. Hazard. Mater. 2020, 388, 121727. [Google Scholar] [CrossRef]
- Meli, R.; Monnolo, A.; Annunziata, C.; Pirozzi, C.; Ferrante, M.C. Oxidative Stress and BPA Toxicity: An Antioxidant Approach for Male and Female Reproductive Dysfunction. Antioxidants 2020, 9, 405. [Google Scholar] [CrossRef]
- Tain, Y.L.; Hsu, C.N. Melatonin Use during Pregnancy and Lactation Complicated by Oxidative Stress: Focus on Offspring’s Cardiovascular-Kidney-Metabolic Health in Animal Models. Antioxidants 2024, 13, 226. [Google Scholar] [CrossRef]
- Cohen, P.A.; Avula, B.; Wang, Y.H.; Katragunta, K.; Khan, I. Quantity of Melatonin and CBD in Melatonin Gummies Sold in the US. JAMA 2023, 329, 1401–1402. [Google Scholar] [CrossRef]
- Lelak, K.; Vohra, V.; Neuman, M.I.; Toce, M.S.; Sethuraman, U. Pediatric Melatonin Ingestions–United States, 2012–2021. MMWR Morb. Mortal. Wkly. Rep. 2022, 71, 725–729. [Google Scholar] [CrossRef]
- Verteramo, R.; Pierdomenico, M.; Greco, P.; Milano, C. The Role of Melatonin in Pregnancy and the Health Benefits for the Newborn. Biomedicines 2022, 10, 3252. [Google Scholar] [CrossRef] [PubMed]
- Händel, M.N.; Andersen, H.K.; Ussing, A.; Virring, A.; Jennum, P.; Debes, N.M.; Laursen, T.; Baandrup, L.; Gade, C.; Dettmann, J.; et al. The short-term and long-term adverse effects of melatonin treatment in children and adolescents: A systematic review and GRADE assessment. eClinicalMedicine 2023, 61, 102083. [Google Scholar] [CrossRef] [PubMed]
- Zwart, T.C.; Smits, M.G.; Egberts, T.C.G.; Rademaker, C.M.A.; van Geijlswijk, I.M. Long-Term Melatonin Therapy for Adolescents and Young Adults with Chronic Sleep Onset Insomnia and Late Melatonin Onset: Evaluation of Sleep Quality, Chronotype, and Lifestyle Factors Compared to Age-Related Randomly Selected Population Cohorts. Healthcare 2018, 6, 23. [Google Scholar] [CrossRef] [PubMed]
- Alers, N.O.; Jenkin, G.; Miller, S.L.; Wallace, E.M. Antenatal melatonin as an antioxidant in human pregnancies complicated by fetal growth restriction–a phase I pilot clinical trial: Study protocol. BMJ Open 2013, 3, e004141. [Google Scholar] [CrossRef]
- Hobson, S.R.; Lim, R.; Gardiner, E.E.; Alers, N.O.; Wallace, E.M. Phase I pilot clinical trial of antenatal maternally administered melatonin to decrease the level of oxidative stress in human pregnancies affected by pre-eclampsia (PAMPR): Study protocol. BMJ Open 2013, 3, e003788. [Google Scholar] [CrossRef]
- Boutin, J.A.; Audinot, V.; Ferry, G.; Delagrange, P. Molecular tools to study melatonin pathways and actions. Trends Pharmacol. Sci. 2005, 26, 412–419. [Google Scholar] [CrossRef]
- Givler, D.; Givler, A.; Luther, P.M.; Wenger, D.M.; Ahmadzadeh, S.; Shekoohi, S.; Edinoff, A.N.; Dorius, B.K.; Baptiste, C.J.; Cornett, E.M.; et al. Chronic Administration of Melatonin: Physiological and Clinical Considerations. Neurol. Int. 2023, 15, 518–533. [Google Scholar] [CrossRef]
- Lui, M.F.G.; Chow, H.K.D.; Wong, W.M.K.; Tsang, W.N.W. Melatonin Affects Postural Control in Community-Dwelling Older Adults While Dual Tasking: A Randomized Observation Study. J. Aging Phys. Act. 2018, 27, 102–107. [Google Scholar] [CrossRef]
- Gooneratne, N.S.; Edwards, A.Y.; Zhou, C.; Cuellar, N.; Grandner, M.A.; Barrett, J.S. Melatonin pharmacokinetics following two different oral surge-sustained release doses in older adults. J. Pineal Res. 2012, 52, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Papagiannidou, E.; Skene, D.J.; Ioannides, C. Potential drug interactions with melatonin. Physiol. Behav. 2014, 131, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Minich, D.M.; Henning, M.; Darley, C.; Fahoum, M.; Schuler, C.B.; Frame, J. Is Melatonin the “Next Vitamin D”?: A Review of Emerging Science, Clinical Uses, Safety, and Dietary Supplements. Nutrients 2022, 14, 3934. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Vico, A.; Lardone, P.J.; Alvarez-Sánchez, N.; Rodríguez-Rodríguez, A.; Guerrero, J.M. Melatonin: Buffering the immune system. Int. J. Mol. Sci. 2013, 14, 8638–8683. [Google Scholar] [CrossRef]
- Sheldon, S.H. Pro-convulsant effects of oral melatonin in neurologically disabled children. Lancet 1998, 351, 1254. [Google Scholar] [CrossRef]
- Maghbooli, M.; NajafAbadi, S.A.; MalekMahmoudi, G.; Molseghi, M.H. Effect of add-on melatonin on seizure outcomes and quality of sleep in epilepsy with idiopathic generalized tonic-clonic seizures alone in adult patients: Cross-sectional, randomized, double-blind, placebo-controlled clinical trial. Brain Behav. 2023, 13, e2860. [Google Scholar] [CrossRef]
- Wirtz, P.H.; Spillmann, M.; Bärtschi, C.; Ehlert, U.; von Känel, R. Oral melatonin reduces blood coagulation activity: A placebo-controlled study in healthy young men. J. Pineal Res. 2008, 44, 127–133. [Google Scholar] [CrossRef]
- Carman, J.S.; Post, R.M.; Buswell, R.; Goodwin, F.K. Negative effects of melatonin on depression. Am. J. Psychiatry 1976, 133, 1181–1186. [Google Scholar] [CrossRef]
- Boutin, J.A.; Kennaway, D.J.; Jockers, R. Melatonin: Facts, Extrapolations and Clinical Trials. Biomolecules 2023, 13, 943. [Google Scholar] [CrossRef]
- Anjum, S.; Rahman, S.; Kaur, M.; Ahmad, F.; Rashid, H.; Ansari, R.A.; Raisuddin, S. Melatonin ameliorates bisphenol A-induced biochemical toxicity in testicular mitochondria of mouse. Food Chem. Toxicol. 2011, 49, 2849–2854. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Specificity | Proposed Mechanism | BPA Concentration | Reference |
|---|---|---|---|---|
| Breast Cancer | General * | Endocrine disruption; promoting adipogenesis, lipogenesis, and adipokine secretion; creating a pro-inflammatory, nutrient-rich environment | Not specified | [36] |
| ERα-positive cells | Synergizes with genistein to induce estrogenic responses, potentially epigenetically reprogramming breast cells | 50 nM | [37] | |
| General | Induces differentiation of adipose cells into cancer-associated adipocyte-like cells, promoting epithelial–mesenchymal transition (EMT) via the CXCL12/AKT pathway | 10 nM | [38] | |
| Prostate Cancer | General | Promotes biochemical recurrence and death by disrupting mitochondrial energy homeostasis, potentially via ESR1-PFKFB4 axis | In vitro and in vivo: 10 nM | [39] |
| Ovarian Cancer | General | Promotes migration and invasion by activating the Wnt/β-catenin/SPP1 axis, leading to osteopontin secretion and transformation of fibroblasts into cancer-associated fibroblasts | In vitro: 10, 100 nM | [40] |
| General | Alters epithelial diversity; induces apoptosis and necrosis; disrupts antioxidant, apoptotic, and inflammatory gene expression | Low dose 1 mg/kg body weight (BW), high dose 5 mg/kg BW | [41] | |
| General | Induces oxidative stress, increasing production of ovarian cancer stem cells | Low dose 1 mg/kg BW, high dose 5 mg/kg BW | [42] | |
| Endometrial Cancer | General | Low-dose BPA alters estrous cycle and uterine pathology in rats, with a gene signature predictive of survival in human patients with endometrial cancer | 25 and 250 μg/kg BW/day | [43] |
| Colorectal Cancer | General | Enhances de novo ceramide synthesis, exacerbating tumor progression and EMT | In vitro: 0.01, 0.1, 1 µM; in vivo: low dose 0.62–1.14 μg/g, high dose 6.65–20.56 μg/g | [44] |
| Obese rats | May worsen progression through the PI3K–AKT pathway and increase IL-1β levels | 25 mg/kg | [45] | |
| General | Upregulates GOLPH3, promoting proliferation and migration | 1 µM | [46] | |
| Colon epithelial cells | Increases cellular invasiveness and anchorage-independent cell growth, potentially through phosphorylation of various protein kinases | Low dose 0.0043 nM | [47] | |
| Thyroid Cancer | Papillary thyroid carcinoma | Enhances proliferation and tumorigenesis through ROS generation and activation of NOX4 signaling pathways | 0.1 and 0.5 µM | [48] |
| Liver Cancer | Hepatocellular carcinoma (in mice) | Alters gene expression in the liver, predicting hepatocellular carcinoma | 50 mg/kg | [49] |
| Liver cells | Induces chemosensitivity and is associated with increased PPARγ expression in digestive system cancers | ~33.70 µg/mL | [50] |
| Specificity of Protective Effect | Proposed Mechanism | Melatonin Concentration | Reference | |
|---|---|---|---|---|
| Ocular Diseases | Protects against ferroptosis in RPE cells in age-related macular degeneration | PI3K/AKT/MDM2/p53 pathway: increases phosphorylated PI3K, AKT, and MDM2, inhibiting p53 and restoring SLC7A11 expression | In vitro: 10–200 μM; in vivo: 10–40 mg/kg | [73] |
| General protective and therapeutic potential for ocular diseases | Antioxidant, anti-inflammatory, immunomodulatory, neuroprotective, regulation of intraocular pressure, and VEGF secretion | Varies depending on disease and study | [74] | |
| IOP-lowering and neuroprotection in glaucoma | Stimulation of melatonin receptors in the ciliary body; antioxidant and neuroprotective effects | Topical formulations: 0.1–17.2 mM | [75] | |
| Antioxidant and neuroprotective effects in diabetic retinopathy | Reduction in oxidative stress and inflammation | Varies depending on study | [74] | |
| Antioxidant protection and regulates ocular tissue functions | Direct scavenging of ROS, stimulation of antioxidant enzymes, interaction with melatonin receptors | 0.07–86 mM | [75] | |
| Neurodegenerative Diseases | Protective activity in Alzheimer’s disease, Parkinson’s disease, and multiple sclerosis | Antioxidant, modulates mitochondrial function and inflammation, synergistic effects with other neuroprotective agents | Optimal dosage has not yet been established in clinical settings | [76] |
| Protection Against Electromagnetic Waves | Protects various organs (brain, skin, eyes, testis, kidney) against cell phone-induced electromagnetic waves | Strengthens cellular antioxidant system, mitigates oxidative stress and cell death | Animal studies: 2–100 mg/kg | [77] |
| Wound Healing | Increases general wound healing (cuts, burns, ulcers) | Antioxidant, anti-inflammatory, and immunomodulatory actions, regulates vascular reactivity and angiogenesis | Physiological (pM) to pharmacological (μM) levels | [78] |
| Skin Aging | Protection against age-related skin deterioration | Combats oxidative stress, shields from UV damage, curbs melanin production, and influences collagen synthesis and mitochondrial activity | Ex vivo: 100–200 µM | [79] |
| Differential antiaging effects on epidermis and dermis | Downregulates mTORC1 activity and MMP-1, increases VEGF-A and fibrillin-1 expressions | Ex vivo: 100–200 µM | [80] | |
| Cardiotoxicity | Protects against 5-fluorouracil-induced cardiotoxicity | Reduces oxidative damage, mitigates inflammation, attenuates the increase in cardiac biomarker levels | Rats: 2.5, 5, and 10 mg/kg/day | [81] |
| Organ Fibrosis | Inhibition of fibrosis in liver, lung, heart, kidney, and skin | Antioxidant, anti-inflammatory effects, remodels extracellular matrix, suppresses EMT, regulates apoptosis | Oral administration: up to 500 mg/day | [7] |
| Other Protective Effects | Mitigation of lead-induced oxidative stress in tobacco BY-2 suspension cells | Protects against lipid profile modification and membrane integrity | Not specified | [82] |
| Increase in sleep quality in adults with various diseases | Not specified | Varies depending on study | [83] |
| Specificity | Proposed Mechanism | Melatonin Concentration | BPA Concentration | Functions | References |
|---|---|---|---|---|---|
| Renal protection | Diminishing oxidative stress, maintaining redox equilibrium within mitochondria, sustaining mitochondrial function and architecture | In vivo: 10 mg/kg; in vitro preincubation: 0.1, 0.5, 1, 10, 100 µM; in vitro after BPA exposure: 0.5 µM | In vivo: 50, 100, 150 mg/kg; in vitro: 1–1000 µM; in vitro with melatonin: 125 µM | Damaging effects of BPA on the kidney and the protection by melatonin: emerging evidence from in vivo and in vitro studies | [94] |
| Protection of testicular mitochondria | Antioxidant properties, direct free radical scavenging activity, lowering mitochondrial lipid peroxidation, restoring mitochondrial enzyme activity, and ameliorating decreased enzymatic and nonenzymatic antioxidants | In vivo: 1–10 mg/kg BW | In vivo: 10 mg/kg BW; environmentally relevant BPA doses: 25 μg/kg | Melatonin ameliorates BPA-induced biochemical toxicity in testicular mitochondria of mice | [95,121] |
| Protection against cytotoxicity and genotoxicity in HGF, MKN45, and MSC cell lines | Antioxidant and free radical scavenging properties, reducing ROS and MDA levels, increasing GSH content, and preventing DNA damage | In vitro: 12, 23, 46, and 93 µg/mL | In vitro: 0.5, 5, 50, and 100 µg/mL; control for melatonin protection: 50 µg/mL | The primary route of human exposure to BPA is oral intake, which is associated with genotoxicity, oxidative stress, endocrine disruption, mutagenicity, and carcinogenicity in in vitro and in vivo models | [9] |
| Protection against reproductive toxicity | Increases in sperm concentration, sperm viability, and testosterone serum levels, potentially through antioxidant properties and interaction with hormone receptors, antiapoptotic effects, and hormonal modulation | Varies across studies: 10–20 mg/kg BW | Varies across included studies: 25–250 μg/kg BW | Melatonin and vitamins as protectors against the reproductive toxicity of bisphenols | [84,93] |
| Neuro-protection in rats | Reduction in oxidative stress (diminished MDA levels), modulation of ERK/NF-kB signaling pathway, attenuation of histopathological alterations in the hippocampus, improvement in behavioral changes | In vivo, oral administration: 20 mg/kg BW (MEL I), 40 mg/kg BW (MEL II) | In vivo, intraperitoneal administration: 1 mg/kg BW | Neurotoxicity of BPA and impact of melatonin administration on oxidative stress, ERK/NF-kB signaling pathway, and behavior in rats | [70,71] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joo, S.S.; Yoo, Y.-M. Protective Effect of Melatonin Against Bisphenol A Toxicity. Int. J. Mol. Sci. 2025, 26, 7526. https://doi.org/10.3390/ijms26157526
Joo SS, Yoo Y-M. Protective Effect of Melatonin Against Bisphenol A Toxicity. International Journal of Molecular Sciences. 2025; 26(15):7526. https://doi.org/10.3390/ijms26157526
Chicago/Turabian StyleJoo, Seong Soo, and Yeong-Min Yoo. 2025. "Protective Effect of Melatonin Against Bisphenol A Toxicity" International Journal of Molecular Sciences 26, no. 15: 7526. https://doi.org/10.3390/ijms26157526
APA StyleJoo, S. S., & Yoo, Y.-M. (2025). Protective Effect of Melatonin Against Bisphenol A Toxicity. International Journal of Molecular Sciences, 26(15), 7526. https://doi.org/10.3390/ijms26157526