
Antioxidants and Reactive Oxygen Species: Shaping Human Health and Disease Outcomes

 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Bioactive Reactive Species
2.1. Production of Reactive Oxygen and Nitrogen Species
2.1.1. Reactive Oxygen Species (ROS)
Endogenous Sources
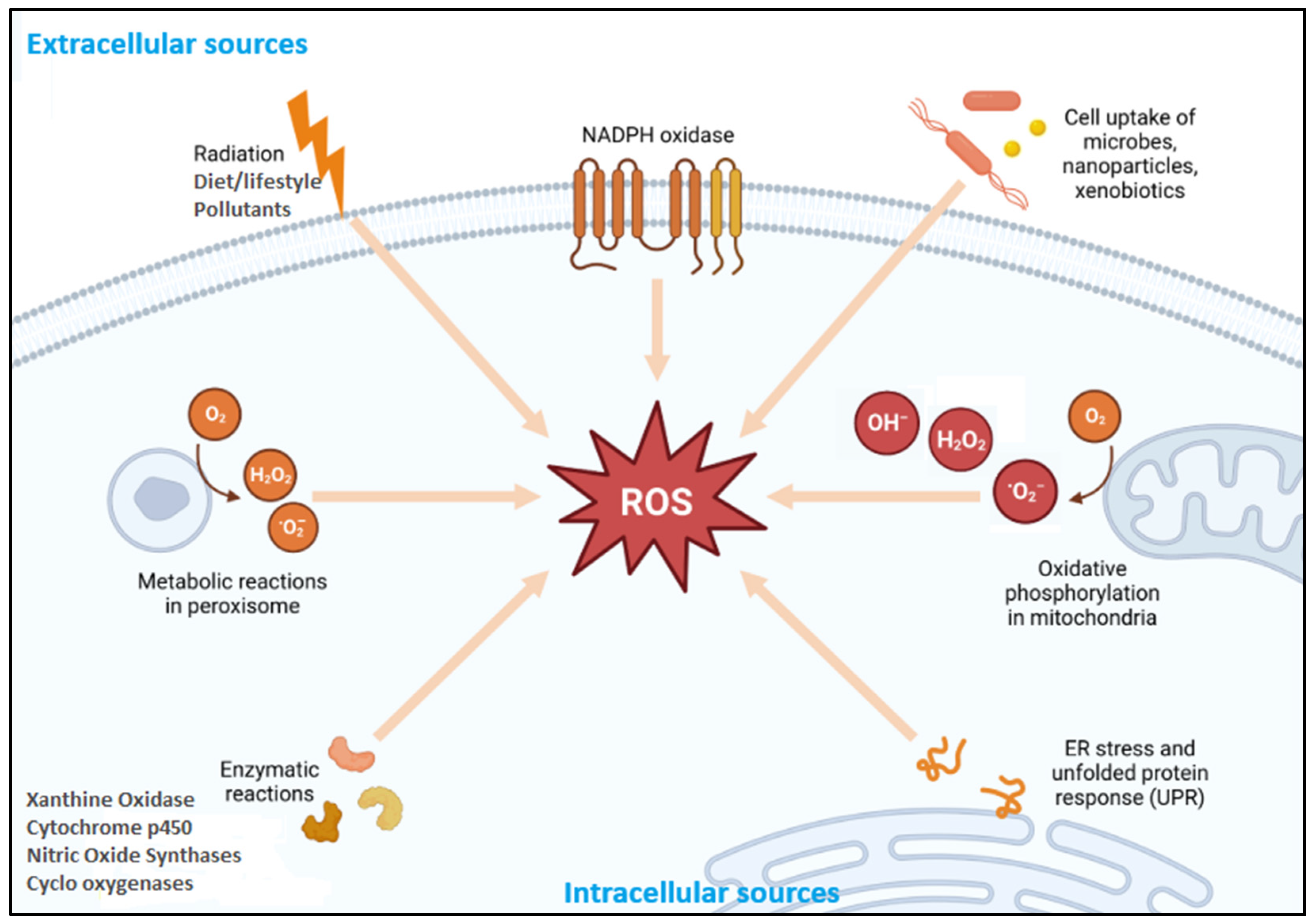
Exogenous Sources
2.1.2. Reactive Nitrogen Species (RNS)
Endogenous Sources
Exogenous Sources
3. Reactive Species in the Pathophysiology of Human Health
3.1. ROS in Cellular Signaling and Oxidative Stress
3.1.1. Physiological Roles of ROS
3.1.2. Pathological Roles of ROS
3.2. RNS in Cellular Signaling and Nitrosative Stress
3.2.1. Physiological Roles of RNS
3.2.2. Pathological Roles of RNS
3.3. Synergistic Effects of Reactive Oxygen and Nitrogen Species
4. Antioxidant Modulation of Oxidative and Nitrosative Stress
4.1. Types of Antioxidant Systems
4.1.1. Endogenous Antioxidants
Enzymatic Antioxidants
Non-Enzymatic Antioxidants
4.1.2. Exogenous Antioxidants
Vitamins as Antioxidants
Polyphenols as Antioxidants
Essential Trace Minerals as Antioxidant Cofactors
Dietary Patterns and Antioxidant Synergy
4.2. Mechanisms of Action of Antioxidants
5. Application of Antioxidant-Based Therapies
5.1. Efficacy and Safety Concerns
5.2. Inconsistent Clinical Trial Outcomes
5.3. Controversies in Cancer Therapy
5.4. Pro-Oxidant Effects and Redox Imbalance
5.5. Biological Complexity and Limitations in Trial Design
6. Emerging Advances in Antioxidant Research: Innovative Solutions to Clinical Limitations
6.1. Nanotechnology-Driven Antioxidant Delivery
6.2. Synergistic and Combination Therapies
6.3. Personalized and Precision Antioxidant Medicine
6.4. Enzyme-Mimetic Antioxidants
6.5. Redox Biomarkers and Real-Time Monitoring Tools
6.6. Future Outlook
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| GPx | Glutathione peroxidase; |
| SOD | Superoxide dismutase; |
| COX-2 | Cyclooxygenase-2; |
| CAT | Catalase; |
| EGCG | Epigallocatechin-3-gallate; |
| DNA | Deoxyribonucleic acid; |
| Nrf2 | Nuclear factor erythroid 2-related factor 2; |
| TNF-α | Tumor necrosis factor-alpha; |
| IL-6 | Interleukin-6; |
| NF-κB | Nuclear factor kappa B; |
| GSTs | Glutathione S-transferases; |
| HO-1 | Heme oxygenase-1; |
| NQO1 | NAD(P)H:quinone oxidoreductase 1; |
| TrxR | Thioredoxin reductase; |
| SIRT1 | Sirtuin 1; |
| CoQ10 | Coenzyme Q10; |
| GSH | Glutathione; |
| PON2 | Paraoxonase-2; |
| HIF-1α | Hypoxia-inducible factor 1-alpha; |
| STAT3 | Signal transducer and activator of transcription 3; |
| FOXO | Forkhead box O; |
| p53 | Tumor protein 53; |
| IKK | IκB kinase; |
| AP-1 | Activator protein 1; |
| Keap1 | Kelch-like ECH-associated protein 1. |
References
- Thomas, C.; Wurzer, L.; Malle, E.; Ristow, M.; Madreiter-Sokolowski, C.T. Modulation of reactive oxygen species homeostasis as a pleiotropic effect of commonly used drugs. Front. Aging 2022, 3, 905261. [Google Scholar] [CrossRef]
- Jakubek, P.; Parchem, K.; Wieckowski, M.R.; Bartoszek, A. The interplay between endogenous and foodborne pro-oxidants and antioxidants in shaping redox homeostasis. Int. J. Mol. Sci. 2024, 25, 7827. [Google Scholar] [CrossRef]
- Pham, T.-P.-T.; Le, T.-H.-T.; Pham, H.-T.-X.; Tran, T.-T.; Pham, V.-T.; Mafruhah, O.R.; Ha, H.-A. Comparative efficacy of antioxidant therapies for sepsis and septic shock in the intensive care unit: A frequentist network meta-analysis. Heliyon 2024, 10, e31447. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.J.; Perfetti, T.A.; King, J.A. Indirect oxidative stress from pulmonary inflammation exceeds direct oxidative stress from chemical damage to mitochondria. Toxicol. Res. Appl. 2019, 3, 2397847319842845. [Google Scholar] [CrossRef]
- Zhao, F.; Li, B.; Yang, W.; Ge, T.; Cui, R. Brain–immune interaction mechanisms: Implications for cognitive dysfunction in psychiatric disorders. Cell Prolif. 2022, 55, e13295. [Google Scholar] [CrossRef] [PubMed]
- Hernansanz-Agustín, P.; Enríquez, J.A. Generation of reactive oxygen species by mitochondria. Antioxidants 2021, 10, 415. [Google Scholar] [CrossRef]
- Morris, G.; Gevezova, M.; Sarafian, V.; Maes, M. Redox regulation of the immune response. Cell. Mol. Immunol. 2022, 19, 1079–1101. [Google Scholar] [CrossRef] [PubMed]
- Andrés, C.M.C.; Pérez de la Lastra, J.M.; Andrés Juan, C.; Plou, F.J.; Pérez-Lebeña, E. Superoxide anion chemistry—Its role at the core of the innate immunity. Int. J. Mol. Sci. 2023, 24, 1841. [Google Scholar] [CrossRef]
- Mukherjee, A.; Ghosh, K.K.; Chakrabortty, S.; Gulyás, B.; Padmanabhan, P.; Ball, W.B. Mitochondrial reactive oxygen species in infection and immunity. Biomolecules 2024, 14, 670. [Google Scholar] [CrossRef]
- Han, C.; Wu, Y.; Rong, J.; Xia, Q.; Du, D. Unveiling the Emerging Role of Xanthine Oxidase in Acute Pancreatitis: Beyond Reactive Oxygen Species. Antioxidants 2025, 14, 95. [Google Scholar] [CrossRef]
- Tang, S.-P.; Mao, X.-L.; Chen, Y.-H.; Yan, L.-L.; Ye, L.-P.; Li, S.-W. Reactive oxygen species induce fatty liver and ischemia-reperfusion injury by promoting inflammation and cell death. Front. Immunol. 2022, 13, 870239. [Google Scholar] [CrossRef]
- Wang, C.-M.; Chen, J.; Zhao, J.; Hu, S.-S.; Zhang, S.-Q.; Mi, X.-Q.; Shi, X.; Cao, X.-H.; Li, Z. Xanthohumol induces ROS through NADPH oxidase, causes cell cycle arrest and apoptosis. Oxidative Med. Cell. Longev. 2021, 2021, 9877170. [Google Scholar] [CrossRef]
- Fukai, T.; Ushio-Fukai, M. Cross-talk between NADPH oxidase and mitochondria: Role in ROS signaling and angiogenesis. Cells 2020, 9, 1849. [Google Scholar] [CrossRef]
- Massart, J.; Begriche, K.; Hartman, J.H.; Fromenty, B. Role of mitochondrial cytochrome P450 2E1 in healthy and diseased liver. Cells 2022, 11, 288. [Google Scholar] [CrossRef]
- Gonzalez, M.; Clayton, S.; Wauson, E.; Christian, D.; Tran, Q.-K. Promotion of nitric oxide production: Mechanisms, strategies, and possibilities. Front. Physiol. 2025, 16, 1545044. [Google Scholar] [CrossRef] [PubMed]
- Abdal Dayem, A.; Hossain, M.K.; Lee, S.B.; Kim, K.; Saha, S.K.; Yang, G.-M.; Choi, H.Y.; Cho, S.-G. The role of reactive oxygen species (ROS) in the biological activities of metallic nanoparticles. Int. J. Mol. Sci. 2017, 18, 120. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Hwang, J.; Park, C.; Park, R. Redox system and ROS-related disorders in peroxisomes. Free Radic. Res. 2024, 58, 662–675. [Google Scholar] [CrossRef]
- Brahma, M.K.; Gilglioni, E.H.; Zhou, L.; Trépo, E.; Chen, P.; Gurzov, E.N. Oxidative stress in obesity-associated hepatocellular carcinoma: Sources, signaling and therapeutic challenges. Oncogene 2021, 40, 5155–5167. [Google Scholar] [CrossRef] [PubMed]
- Rey, F.; Berardo, C.; Maghraby, E.; Mauri, A.; Messa, L.; Esposito, L.; Casili, G.; Ottolenghi, S.; Bonaventura, E.; Cuzzocrea, S. Redox imbalance in neurological disorders in adults and children. Antioxidants 2023, 12, 965. [Google Scholar] [CrossRef]
- Shoham, S.; Pintel, N.; Avni, D. Oxidative Stress, Gut Bacteria, and Microalgae: A Holistic Approach to Manage Inflammatory Bowel Diseases. Antioxidants 2025, 14, 697. [Google Scholar] [CrossRef]
- Wei, M.; He, X.; Liu, N.; Deng, H. Role of reactive oxygen species in ultraviolet-induced photodamage of the skin. Cell Div. 2024, 19, 1. [Google Scholar] [CrossRef]
- Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Arancibia-Hernández, Y.L.; Hernández-Cruz, E.Y.; Pedraza-Chaverri, J. RONS and oxidative stress: An overview of basic concepts. Oxygen 2022, 2, 437–478. [Google Scholar] [CrossRef]
- Talapko, J.; Talapko, D.; Katalinić, D.; Kotris, I.; Erić, I.; Belić, D.; Vasilj Mihaljević, M.; Vasilj, A.; Erić, S.; Flam, J. Health Effects of Ionizing Radiation on the Human Body. Medicina 2024, 60, 653. [Google Scholar] [CrossRef]
- Daiber, A.; Kuntic, M.; Hahad, O.; Delogu, L.G.; Rohrbach, S.; Di Lisa, F.; Schulz, R.; Münzel, T. Effects of air pollution particles (ultrafine and fine particulate matter) on mitochondrial function and oxidative stress–Implications for cardiovascular and neurodegenerative diseases. Arch. Biochem. Biophys. 2020, 696, 108662. [Google Scholar] [CrossRef]
- Seo, Y.-S.; Park, J.-M.; Kim, J.-H.; Lee, M.-Y. Cigarette smoke-induced reactive oxygen species formation: A concise review. Antioxidants 2023, 12, 1732. [Google Scholar] [CrossRef]
- Teschke, R. Aluminum, arsenic, beryllium, cadmium, chromium, cobalt, copper, iron, lead, mercury, molybdenum, nickel, platinum, thallium, titanium, vanadium, and zinc: Molecular aspects in experimental liver injury. Int. J. Mol. Sci. 2022, 23, 12213. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, K.; Song, X.; Huang, Q.; Lin, S.; Deng, S.; Qi, M.; Yang, Y.; Lu, Q.; Zhao, D. TLR4 overexpression aggravates bacterial lipopolysaccharide-induced apoptosis via excessive autophagy and NF-κB/MAPK signaling in transgenic mammal models. Cells 2023, 12, 1769. [Google Scholar] [CrossRef] [PubMed]
- Youn, D.Y.; Xiaoli, A.M.; Kwon, H.; Yang, F.; Pessin, J.E. The subunit assembly state of the Mediator complex is nutrient-regulated and is dysregulated in a genetic model of insulin resistance and obesity. J. Biol. Chem. 2019, 294, 9076–9083. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.K.; Yates, E.; Lilly, K.; Dhanda, A.D. Oxidative stress in alcohol-related liver disease. World J. Hepatol. 2020, 12, 332. [Google Scholar] [CrossRef]
- Nakai, K.; Tsuruta, D. What are reactive oxygen species, free radicals, and oxidative stress in skin diseases? Int. J. Mol. Sci. 2021, 22, 10799. [Google Scholar] [CrossRef]
- Dash, U.C.; Nayak, V.; Navani, H.S.; Samal, R.R.; Agrawal, P.; Singh, A.K.; Majhi, S.; Mogare, D.G.; Duttaroy, A.K.; Jena, A.B. Understanding the molecular bridges between the drugs and immune cell. Pharmacol. Ther. 2025, 267, 108805. [Google Scholar] [CrossRef] [PubMed]
- Lushchak, V.I.; Lushchak, O. Interplay between reactive oxygen and nitrogen species in living organisms. Chem.-Biol. Interact. 2021, 349, 109680. [Google Scholar] [CrossRef] [PubMed]
- Iova, O.-M.; Marin, G.-E.; Lazar, I.; Stanescu, I.; Dogaru, G.; Nicula, C.A.; Bulboacă, A.E. Nitric oxide/nitric oxide synthase system in the pathogenesis of neurodegenerative disorders—An overview. Antioxidants 2023, 12, 753. [Google Scholar] [CrossRef]
- Casella, A.C.; Prolo, C.; Pereyra, J.; Ríos, N.; Piacenza, L.; Radi, R.; Álvarez, M.N. Superoxide, nitric oxide and peroxynitrite production by macrophages under different physiological oxygen tensions. Free Radic. Biol. Med. 2024, 212, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.E.; Lee, J.S. Advances in the Regulation of Inflammatory Mediators in Nitric Oxide Synthase: Implications for Disease Modulation and Therapeutic Approaches. Int. J. Mol. Sci. 2025, 26, 1204. [Google Scholar] [CrossRef]
- Beigloo, F.; Khoshkar-Vandi, S.A.; Pourmand, E.; Heydari, M.; Molaabasi, F.; Gharib, N.; Zare, Y.; Rhee, K.Y.; Park, S.-J. Electrochemical micro-and nanobiosensors for in vivo reactive oxygen/nitrogen species measurement in the brain. Nanotechnol. Rev. 2023, 12, 20230124. [Google Scholar] [CrossRef]
- Crochemore, C.; Cimmaruta, C.; Fernández-Molina, C.; Ricchetti, M. Reactive species in progeroid syndromes and aging-related processes. Antioxid. Redox Signal. 2022, 37, 208–228. [Google Scholar] [CrossRef]
- Khelfi, A. Reactive Species. In Biomarkers of Oxidative Stress: Basics and Measurement of Oxidative Stress; Springer: Berlin/Heidelberg, Germany, 2024; pp. 25–68. [Google Scholar]
- Hawkins, C.L.; Davies, M.J. Role of myeloperoxidase and oxidant formation in the extracellular environment in inflammation-induced tissue damage. Free Radic. Biol. Med. 2021, 172, 633–651. [Google Scholar] [CrossRef]
- da Fonseca, C.A.R.; Paltian, J.J.; da Motta, K.P.; Martins, C.C.v.o.; Kazmierczak, J.C.; Schumacher, R.F.; Soares, M.P.; Luchese, C.; Wilhelm, E.A. Aging is a Risk Factor for Rheumatoid Arthritis in Rats: Therapeutic Potential of 4-(Phenylselanyl)-2H-chromen-2-one. ACS Omega 2025, 10, 25990–26005. [Google Scholar] [CrossRef]
- Bortolotti, M.; Polito, L.; Battelli, M.G.; Bolognesi, A. Xanthine oxidoreductase: One enzyme for multiple physiological tasks. Redox Biol. 2021, 41, 101882. [Google Scholar] [CrossRef]
- Joshi, B.; Singh, S.; Sharma, P.; Mohapatra, T.; Kumar, P. Effect of Cigarette Smoking on Selected Antioxidant Enzymes and Oxidative Stress Biomarkers. J. Clin. Diagn. Res. 2020, 14, 19. [Google Scholar] [CrossRef]
- Biedrzycki, G.; Wolszczak-Biedrzycka, B.; Dorf, J.; Maciejczyk, M. The antioxidant barrier, oxidative/nitrosative stress, and protein glycation in allergy: From basic research to clinical practice. Front. Immunol. 2024, 15, 1440313. [Google Scholar] [CrossRef] [PubMed]
- Karwowska, M.; Kononiuk, A. Nitrates/nitrites in food—Risk for nitrosative stress and benefits. Antioxidants 2020, 9, 241. [Google Scholar] [CrossRef]
- de Andrade Júnior, F.P.; Sobreira de Cabral, A.L.; Dantas de Araújo, J.M.; Cordeiro, L.V.; de Barros Cândido, M.; Pontes da Silva, A.; de Medeiros Lima, B.T.; Braga Dantas, B. Food nitrates and nitrites as possible causes of cancer: A review. Rev. Colomb. De Cienc. Químico-Farm. 2021, 50, 269–291. [Google Scholar] [CrossRef]
- Olas, B. The cardioprotective role of nitrate-rich vegetables. Foods 2024, 13, 691. [Google Scholar] [CrossRef] [PubMed]
- Iliadis, S.; Papanikolaou, N.A. Reactive Oxygen Species Mechanisms that Regulate Protein–Protein Interactions in Cancer. Int. J. Mol. Sci. 2024, 25, 9255. [Google Scholar] [CrossRef]
- Santos, A.L.; Sinha, S.; Lindner, A.B. The good, the bad, and the ugly of ROS: New insights on aging and aging-related diseases from eukaryotic and prokaryotic model organisms. Oxidative Med. Cell. Longev. 2018, 2018, 1941285. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The chemistry of reactive oxygen species (ROS) revisited: Outlining their role in biological macromolecules (DNA, lipids and proteins) and induced pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]
- Chandimali, N.; Bak, S.G.; Park, E.H.; Lim, H.-J.; Won, Y.-S.; Kim, E.-K.; Park, S.-I.; Lee, S.J. Free radicals and their impact on health and antioxidant defenses: A review. Cell Death Discov. 2025, 11, 19. [Google Scholar] [CrossRef]
- Liu, H.-M.; Cheng, M.-Y.; Xun, M.-H.; Zhao, Z.-W.; Zhang, Y.; Tang, W.; Cheng, J.; Ni, J.; Wang, W. Possible mechanisms of oxidative stress-induced skin cellular senescence, inflammation, and cancer and the therapeutic potential of plant polyphenols. Int. J. Mol. Sci. 2023, 24, 3755. [Google Scholar] [CrossRef]
- Selvaraj, N.R.; Nandan, D.; Nair, B.G.; Nair, V.A.; Venugopal, P.; Aradhya, R. Oxidative Stress and Redox Imbalance: Common Mechanisms in Cancer Stem Cells and Neurodegenerative Diseases. Cells 2025, 14, 511. [Google Scholar] [CrossRef]
- Ibrahim, N.I.; Naina Mohamed, I. Interdependence of anti-inflammatory and antioxidant properties of squalene–implication for cardiovascular health. Life 2021, 11, 103. [Google Scholar] [CrossRef]
- Liu, Z.-Y.; Song, K.; Tu, B.; Lin, L.-C.; Sun, H.; Zhou, Y.; Li, R.; Shi, Y.; Yang, J.-J.; Zhang, Y. Crosstalk between oxidative stress and epigenetic marks: New roles and therapeutic implications in cardiac fibrosis. Redox Biol. 2023, 65, 102820. [Google Scholar] [CrossRef] [PubMed]
- Di Carlo, E.; Sorrentino, C. Oxidative stress and age-related tumors. Antioxidants 2024, 13, 1109. [Google Scholar] [CrossRef]
- Baird, L.; Yamamoto, M. The Keap1-Nrf2 pathway: From mechanism to medical applications. In Oxidative Stress; Elsevier: Amsterdam, The Netherlands, 2020; pp. 125–147. [Google Scholar]
- Zhang, D.D. Thirty years of NRF2: Advances and therapeutic challenges. Nat. Rev. Drug Discov. 2025, 24, 421–444. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Xiao, Y.; Miao, J.; Zhang, X.; Liu, M.; Zhu, L.; Liu, H.; Shen, X.; Wang, J.; Xie, B. Oxidative Stress and Inflammation: Drivers of Tumorigenesis and Therapeutic Opportunities. Antioxidants 2025, 14, 735. [Google Scholar] [CrossRef] [PubMed]
- Atsaves, V.; Leventaki, V.; Rassidakis, G.Z.; Claret, F.X. AP-1 transcription factors as regulators of immune responses in cancer. Cancers 2019, 11, 1037. [Google Scholar] [CrossRef]
- Lai, J.-Q.; Zhao, L.-L.; Hong, C.; Zou, Q.-M.; Su, J.-X.; Li, S.-J.; Zhou, X.-F.; Li, Z.-S.; Deng, B.; Cao, J. Baicalein triggers ferroptosis in colorectal cancer cells via blocking the JAK2/STAT3/GPX4 axis. Acta Pharmacol. Sin. 2024, 45, 1715–1726. [Google Scholar] [CrossRef]
- Bae, T.; Hallis, S.P.; Kwak, M.-K. Hypoxia, oxidative stress, and the interplay of HIFs and NRF2 signaling in cancer. Exp. Mol. Med. 2024, 56, 501–514. [Google Scholar] [CrossRef]
- Kim, M.E.; Kim, D.H.; Lee, J.S. FoxO transcription factors: Applicability as a novel immune cell regulators and therapeutic targets in oxidative stress-related diseases. Int. J. Mol. Sci. 2022, 23, 11877. [Google Scholar] [CrossRef]
- Recknagel, R.O.; Glende, E.A.; Britton, R.S. Free radical damage and lipid peroxidation. In Hepatotoxicology; CRC Press: Boca Raton, FL, USA, 2020; pp. 401–436. [Google Scholar]
- Wang, B.; Wang, Y.; Zhang, J.; Hu, C.; Jiang, J.; Li, Y.; Peng, Z. ROS-induced lipid peroxidation modulates cell death outcome: Mechanisms behind apoptosis, autophagy, and ferroptosis. Arch. Toxicol. 2023, 97, 1439–1451. [Google Scholar] [CrossRef] [PubMed]
- Bellanti, F.; Coda, A.R.D.; Trecca, M.I.; Lo Buglio, A.; Serviddio, G.; Vendemiale, G. Redox Imbalance in Inflammation: The Interplay of Oxidative and Reductive Stress. Antioxidants 2025, 14, 656. [Google Scholar] [CrossRef]
- Al-Taie, A.; Sancar, M.; Izzettin, F.V. 8-Hydroxydeoxyguanosine: A valuable predictor of oxidative DNA damage in cancer and diabetes mellitus. In Cancer; Elsevier: Amsterdam, The Netherlands, 2021; pp. 179–187. [Google Scholar]
- Andrabi, S.M.; Sharma, N.S.; Karan, A.; Shahriar, S.S.; Cordon, B.; Ma, B.; Xie, J. Nitric oxide: Physiological functions, delivery, and biomedical applications. Adv. Sci. 2023, 10, 2303259. [Google Scholar] [CrossRef] [PubMed]
- Lourenço, C.F.; Laranjinha, J. Nitric oxide pathways in neurovascular coupling under normal and stress conditions in the brain: Strategies to rescue aberrant coupling and improve cerebral blood flow. Front. Physiol. 2021, 12, 729201. [Google Scholar] [CrossRef]
- Andrés, C.M.C.; Pérez de la Lastra, J.M.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. The role of reactive species on innate immunity. Vaccines 2022, 10, 1735. [Google Scholar] [CrossRef]
- Halabitska, I.; Oksenych, V.; Kamyshnyi, O. Exploring the efficacy of alpha-lipoic acid in Comorbid Osteoarthritis and Type 2 diabetes Mellitus. Nutrients 2024, 16, 3349. [Google Scholar] [CrossRef]
- Kopacz, A.; Kloska, D.; Forman, H.J.; Jozkowicz, A.; Grochot-Przeczek, A. Beyond repression of Nrf2: An update on Keap1. Free Radic. Biol. Med. 2020, 157, 63–74. [Google Scholar] [CrossRef]
- Wang, F.; Yuan, Q.; Chen, F.; Pang, J.; Pan, C.; Xu, F.; Chen, Y. Fundamental mechanisms of the cell death caused by nitrosative stress. Front. Cell Dev. Biol. 2021, 9, 742483. [Google Scholar] [CrossRef]
- Prolo, C.; Piacenza, L.; Radi, R. Peroxynitrite: A multifaceted oxidizing and nitrating metabolite. Curr. Opin. Chem. Biol. 2024, 80, 102459. [Google Scholar] [CrossRef]
- Griswold-Prenner, I.; Kashyap, A.K.; Mazhar, S.; Hall, Z.W.; Fazelinia, H.; Ischiropoulos, H. Unveiling the human nitroproteome: Protein tyrosine nitration in cell signaling and cancer. J. Biol. Chem. 2023, 299, 105038. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Oh, C.-k.; Zhang, X.; Lipton, S.A. Protein S-nitrosylation and oxidation contribute to protein misfolding in neurodegeneration. Free Radic. Biol. Med. 2021, 172, 562–577. [Google Scholar] [CrossRef]
- Guil-Luna, S.; Sanchez-Montero, M.T.; Rodríguez-Ariza, A. S-Nitrosylation at the intersection of metabolism and autophagy: Implications for cancer. Biochim. Et Biophys. Acta (BBA)-Rev. Cancer 2023, 1878, 189012. [Google Scholar] [CrossRef]
- Sharma, V.; Fernando, V.; Letson, J.; Walia, Y.; Zheng, X.; Fackelman, D.; Furuta, S. S-Nitrosylation in tumor microenvironment. Int. J. Mol. Sci. 2021, 22, 4600. [Google Scholar] [CrossRef]
- Andrés, C.M.C.; Lastra, J.M.P.d.l.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Chemical insights into oxidative and nitrative modifications of DNA. Int. J. Mol. Sci. 2023, 24, 15240. [Google Scholar] [CrossRef]
- Ushio-Fukai, M.; Ash, D.; Nagarkoti, S.; Belin de Chantemèle, E.J.; Fulton, D.J.; Fukai, T. Interplay between reactive oxygen/reactive nitrogen species and metabolism in vascular biology and disease. Antioxid. Redox Signal. 2021, 34, 1319–1354. [Google Scholar] [CrossRef]
- Jurkovicova, D.; Neophytou, C.M.; Gašparović, A.Č.; Gonçalves, A.C. DNA damage response in cancer therapy and resistance: Challenges and opportunities. Int. J. Mol. Sci. 2022, 23, 14672. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, P.; Janmeda, P.; Docea, A.O.; Yeskaliyeva, B.; Abdull Razis, A.F.; Modu, B.; Calina, D.; Sharifi-Rad, J. Oxidative stress, free radicals and antioxidants: Potential crosstalk in the pathophysiology of human diseases. Front. Chem. 2023, 11, 1158198. [Google Scholar] [CrossRef] [PubMed]
- Batty, M.; Bennett, M.R.; Yu, E. The role of oxidative stress in atherosclerosis. Cells 2022, 11, 3843. [Google Scholar] [CrossRef] [PubMed]
- Griendling, K.K.; Camargo, L.L.; Rios, F.J.; Alves-Lopes, R.; Montezano, A.C.; Touyz, R.M. Oxidative stress and hypertension. Circ. Res. 2021, 128, 993–1020. [Google Scholar] [CrossRef]
- Shah, A.K.; Bhullar, S.K.; Elimban, V.; Dhalla, N.S. Oxidative Stress as A Mechanism for Functional Alterations in Cardiac Hypertrophy and Heart Failure. Antioxidants 2021, 10, 931. [Google Scholar] [CrossRef]
- Zheng, J.; Winderickx, J.; Franssens, V.; Liu, B. A mitochondria-associated oxidative stress perspective on Huntington’s disease. Front. Mol. Neurosci. 2018, 11, 329. [Google Scholar] [CrossRef]
- Gil-Mohapel, J.; S. Brocardo, P.; R. Christie, B. The role of oxidative stress in Huntington’s disease: Are antioxidants good therapeutic candidates? Curr. Drug Targets 2014, 15, 454–468. [Google Scholar] [CrossRef]
- Bai, R.; Guo, J.; Ye, X.-Y.; Xie, Y.; Xie, T. Oxidative stress: The core pathogenesis and mechanism of Alzheimer’s disease. Ageing Res. Rev. 2022, 77, 101619. [Google Scholar] [CrossRef]
- Guo, J.D.; Zhao, X.; Li, Y.; Li, G.R.; Liu, X.L. Damage to dopaminergic neurons by oxidative stress in Parkinson’s disease. Int. J. Mol. Med. 2018, 41, 1817–1825. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative stress in cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Sathyapalan, T.; Atkin, S.L.; Sahebkar, A. Molecular mechanisms linking oxidative stress and diabetes mellitus. Oxidative Med. Cell. Longev. 2020, 2020, 8609213. [Google Scholar] [CrossRef] [PubMed]
- Darenskaya, M.; Kolesnikova, L.a.; Kolesnikov, S. Oxidative stress: Pathogenetic role in diabetes mellitus and its complications and therapeutic approaches to correction. Bull. Exp. Biol. Med. 2021, 171, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Zamudio-Cuevas, Y.; Martínez-Flores, K.; Martínez-Nava, G.A.; Clavijo-Cornejo, D.; Fernández-Torres, J.; Sánchez-Sánchez, R. Rheumatoid arthritis and oxidative stress. Cell. Mol. Biol. 2022, 68, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Muro, P.; Zhang, L.; Li, S.; Zhao, Z.; Jin, T.; Mao, F.; Mao, Z. The emerging role of oxidative stress in inflammatory bowel disease. Front. Endocrinol. 2024, 15, 1390351. [Google Scholar] [CrossRef]
- Sahoo, D.K.; Heilmann, R.M.; Paital, B.; Patel, A.; Yadav, V.K.; Wong, D.; Jergens, A.E. Oxidative stress, hormones, and effects of natural antioxidants on intestinal inflammation in inflammatory bowel disease. Front. Endocrinol. 2023, 14, 1217165. [Google Scholar] [CrossRef]
- Saunders, R.M.; Biddle, M.; Amrani, Y.; Brightling, C.E. Stressed out-The role of oxidative stress in airway smooth muscle dysfunction in asthma and COPD. Free Radic. Biol. Med. 2022, 185, 97–119. [Google Scholar] [CrossRef]
- Lewis, B.W.; Ford, M.L.; Rogers, L.K.; Britt, R.D., Jr. Oxidative stress promotes corticosteroid insensitivity in asthma and COPD. Antioxidants 2021, 10, 1335. [Google Scholar] [CrossRef]
- Nucera, F.; Mumby, S.; Paudel, K.R.; Dharwal, V.; Stefano, A.D.; Casolaro, V.; Hansbro, P.M.; Adcock, I.M.; Caramori, G. Role of oxidative stress in the pathogenesis of COPD. Minerva Medica 2022, 113, 370–404. [Google Scholar] [CrossRef] [PubMed]
- Piko, N.; Bevc, S.; Hojs, R.; Ekart, R. The role of oxidative stress in kidney injury. Antioxidants 2023, 12, 1772. [Google Scholar] [CrossRef]
- Jabeen, S.; Noor, S. Oxidative stress and chronic kidney disease. Fundam. Princ. Oxidative Stress Metab. Reprod. 2024, 151–165. [Google Scholar] [CrossRef]
- Ruan, Y.; Jiang, S.; Gericke, A. Age-related macular degeneration: Role of oxidative stress and blood vessels. Int. J. Mol. Sci. 2021, 22, 1296. [Google Scholar] [CrossRef] [PubMed]
- Terao, R.; Ahmed, T.; Suzumura, A.; Terasaki, H. Oxidative stress-induced cellular senescence in aging retina and age-related macular degeneration. Antioxidants 2022, 11, 2189. [Google Scholar] [CrossRef]
- Delli Bovi, A.P.; Marciano, F.; Mandato, C.; Siano, M.A.; Savoia, M.; Vajro, P. Oxidative stress in non-alcoholic fatty liver disease. An updated mini review. Front. Med. 2021, 8, 595371. [Google Scholar] [CrossRef]
- Lian, D.; Chen, M.-M.; Wu, H.; Deng, S.; Hu, X. The role of oxidative stress in skeletal muscle myogenesis and muscle disease. Antioxidants 2022, 11, 755. [Google Scholar] [CrossRef]
- Zhang, H.; Qi, G.; Wang, K.; Yang, J.; Shen, Y.; Yang, X.; Chen, X.; Yao, X.; Gu, X.; Qi, L. Oxidative stress: Roles in skeletal muscle atrophy. Biochem. Pharmacol. 2023, 214, 115664. [Google Scholar] [CrossRef]
- Joffre, J.; Hellman, J. Oxidative stress and endothelial dysfunction in sepsis and acute inflammation. Antioxid. Redox Signal. 2021, 35, 1291–1307. [Google Scholar] [CrossRef] [PubMed]
- Harshithkumar, R.; Shah, P.; Jadaun, P.; Mukherjee, A. ROS chronicles in HIV infection: Genesis of oxidative stress, associated pathologies, and therapeutic strategies. Curr. Issues Mol. Biol. 2024, 46, 8852–8873. [Google Scholar] [CrossRef]
- Perl, A. Oxidative stress in the pathology and treatment of systemic lupus erythematosus. Nat. Rev. Rheumatol. 2013, 9, 674–686. [Google Scholar] [CrossRef]
- Jomova, K.; Raptova, R.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: Chronic diseases and aging. Arch. Toxicol. 2023, 97, 2499–2574. [Google Scholar] [CrossRef]
- Kvedariene, V.; Vaskovic, M.; Semyte, J.B. Role of Oxidative Stress and Antioxidants in the Course of Atopic Dermatitis. Int. J. Mol. Sci. 2025, 26, 4210. [Google Scholar] [CrossRef]
- Gulcin, İ. Antioxidants: A comprehensive review. Arch. Toxicol. 2025, 99, 1893–1997. [Google Scholar] [CrossRef] [PubMed]
- Tumilaar, S.G.; Hardianto, A.; Dohi, H.; Kurnia, D. A comprehensive review of free radicals, oxidative stress, and antioxidants: Overview, clinical applications, global perspectives, future directions, and mechanisms of antioxidant activity of flavonoid compounds. J. Chem. 2024, 2024, 5594386. [Google Scholar] [CrossRef]
- Khan, A.; Gothalwal, R.; Kotnis, A. Reactive Oxygen Species: Generation and Signaling Pathways. In The Role of Reactive Oxygen Species in Human Health and Disease; IGI Global Scientific Publishing: Hershey, PA, USA, 2025; pp. 1–42. [Google Scholar]
- Janssen, W.J.; Nozik-Grayck, E. Power of place: Intravascular superoxide dismutase for prevention of acute respiratory distress syndrome. Am. J. Respir. Cell Mol. Biol. 2017, 56, 147–149. [Google Scholar] [CrossRef]
- Palma, F.R.; He, C.; Danes, J.M.; Paviani, V.; Coelho, D.R.; Gantner, B.N.; Bonini, M.G. Mitochondrial superoxide dismutase: What the established, the intriguing, and the novel reveal about a key cellular redox switch. Antioxid. Redox Signal. 2020, 32, 701–714. [Google Scholar] [CrossRef]
- Kowalczyk, P.; Sulejczak, D.; Kleczkowska, P.; Bukowska-Ośko, I.; Kucia, M.; Popiel, M.; Wietrak, E.; Kramkowski, K.; Wrzosek, K.; Kaczyńska, K. Mitochondrial oxidative stress—A causative factor and therapeutic target in many diseases. Int. J. Mol. Sci. 2021, 22, 13384. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Kelley, J.; Ye, Y.; Ye, Z.-W.; Townsend, D.M.; Zhang, J.; Wu, Y. REDOX Imbalance and Oxidative Stress in the Intervertebral Disc: The Effect of Mechanical Stress and Cigarette Smoking on ER Stress and Mitochondrial Dysfunction. Cells 2025, 14, 613. [Google Scholar] [CrossRef]
- Pei, J.; Pan, X.; Wei, G.; Hua, Y. Research progress of glutathione peroxidase family (GPX) in redoxidation. Front. Pharmacol. 2023, 14, 1147414. [Google Scholar] [CrossRef] [PubMed]
- Stolwijk, J.M.; Falls-Hubert, K.C.; Searby, C.C.; Wagner, B.A.; Buettner, G.R. Simultaneous detection of the enzyme activities of GPx1 and GPx4 guide optimization of selenium in cell biological experiments. Redox Biol. 2020, 32, 101518. [Google Scholar] [CrossRef]
- Parween, F.; Gupta, R.D. Insights into the role of paraoxonase 2 in human pathophysiology. J. Biosci. 2022, 47, 4. [Google Scholar] [CrossRef]
- Campagna, R.; Serritelli, E.N.; Salvolini, E.; Schiavoni, V.; Cecati, M.; Sartini, D.; Pozzi, V.; Emanuelli, M. Contribution of the Paraoxonase-2 Enzyme to Cancer Cell Metabolism and Phenotypes. Biomolecules 2024, 14, 208. [Google Scholar] [CrossRef] [PubMed]
- Fumarola, S.; Cecati, M.; Sartini, D.; Ferretti, G.; Milanese, G.; Galosi, A.B.; Pozzi, V.; Campagna, R.; Morresi, C.; Emanuelli, M. Bladder cancer chemosensitivity is affected by paraoxonase-2 expression. Antioxidants 2020, 9, 175. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Meza, H.; Vilchis-Landeros, M.M.; Vázquez-Carrada, M.; Uribe-Ramírez, D.; Matuz-Mares, D. Cellular compartmentalization, glutathione transport and its relevance in some pathologies. Antioxidants 2023, 12, 834. [Google Scholar] [CrossRef]
- Townsend, D.M.; Tew, K.D.; Tapiero, H. The importance of glutathione in human disease. Biomed. Pharmacother. 2003, 57, 145–155. [Google Scholar] [CrossRef]
- Tram, N.K.; McLean, R.M.; Swindle-Reilly, K.E. Glutathione improves the antioxidant activity of vitamin C in human lens and retinal epithelial cells: Implications for vitreous substitutes. Curr. Eye Res. 2021, 46, 470–481. [Google Scholar] [CrossRef]
- Biswas, P.; Dellanoce, C.; Vezzoli, A.; Mrakic-Sposta, S.; Malnati, M.; Beretta, A.; Accinni, R. Antioxidant activity with increased endogenous levels of vitamin C, E and A following dietary supplementation with a combination of glutathione and resveratrol precursors. Nutrients 2020, 12, 3224. [Google Scholar] [CrossRef]
- Pinz, M.P.; Medeiros, I.; Carvalho, L.A.d.C.; Meotti, F.C. Is uric acid a true antioxidant? Identification of uric acid oxidation products and their biological effects. Redox Rep. 2025, 30, 2498105. [Google Scholar] [CrossRef]
- Du, L.; Zong, Y.; Li, H.; Wang, Q.; Xie, L.; Yang, B.; Pang, Y.; Zhang, C.; Zhong, Z.; Gao, J. Hyperuricemia and its related diseases: Mechanisms and advances in therapy. Signal Transduct. Target. Ther. 2024, 9, 212. [Google Scholar] [CrossRef] [PubMed]
- Nitti, M.; Furfaro, A.L.; Mann, G.E. Heme oxygenase dependent bilirubin generation in vascular cells: A role in preventing endothelial dysfunction in local tissue microenvironment? Front. Physiol. 2020, 11, 23. [Google Scholar] [CrossRef] [PubMed]
- Vitek, L.; Hinds, T.D.; Stec, D.E.; Tiribelli, C. The physiology of bilirubin: Health and disease equilibrium. Trends Mol. Med. 2023, 29, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Dini, I. The potential of Dietary antioxidants. Antioxidants 2021, 10, 1752. [Google Scholar] [CrossRef]
- Dini, I. Contribution of nanoscience research in antioxidants delivery used in nutricosmetic sector. Antioxidants 2022, 11, 563. [Google Scholar] [CrossRef]
- Martelli, A.; Testai, L.; Colletti, A.; Cicero, A.F. Coenzyme Q10: Clinical applications in cardiovascular diseases. Antioxidants 2020, 9, 341. [Google Scholar] [CrossRef]
- Wang, X.; Xiong, X. Mitochondrial Reactive Oxygen Species (mROS) Generation and Cancer: Emerging Nanoparticle Therapeutic Approaches. Int. J. Nanomed. 2025, 6085–6119. [Google Scholar] [CrossRef]
- Fu, M.; Yoon, K.-S.; Ha, J.; Kang, I.; Choe, W. Crosstalk between antioxidants and adipogenesis: Mechanistic pathways and their roles in metabolic health. Antioxidants 2025, 14, 203. [Google Scholar] [CrossRef]
- Zhitkovich, A. Nuclear and cytoplasmic functions of vitamin C. Chem. Res. Toxicol. 2020, 33, 2515–2526. [Google Scholar] [CrossRef]
- Briggs, M.A. From foods to chemotherapeutics: The antioxidant potential of dietary phytochemicals. Processes 2022, 10, 1222. [Google Scholar] [CrossRef]
- Niki, E. Lipid oxidation that is, and is not, inhibited by vitamin E: Consideration about physiological functions of vitamin E. Free Radic. Biol. Med. 2021, 176, 1–15. [Google Scholar] [CrossRef]
- Yang, C.; Jiang, Q. Vitamin E δ-tocotrienol inhibits TNF-α-stimulated NF-κB activation by up-regulation of anti-inflammatory A20 via modulation of sphingolipid including elevation of intracellular dihydroceramides. J. Nutr. Biochem. 2019, 64, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Maboud, M.; Menshawy, A.; Menshawy, E.; Emara, A.; Alshandidy, M.; Eid, M. The efficacy of vitamin E in reducing non-alcoholic fatty liver disease: A systematic review, meta-analysis, and meta-regression. Ther. Adv. Gastroenterol. 2020, 13, 1756284820974917. [Google Scholar] [CrossRef]
- Pérez-Gálvez, A.; Viera, I.; Roca, M. Carotenoids and chlorophylls as antioxidants. Antioxidants 2020, 9, 505. [Google Scholar] [CrossRef]
- Blanco, A.; Amengual, J. Immunometabolic effects of β-carotene and vitamin A in atherogenesis. Immunometabolism 2024, 6, e00051. [Google Scholar] [CrossRef]
- Farhana, A.; Khan, Y.S.; Alsrhani, A.; Manni, E.; Alameen, A.A.; Derafa, W.; Alhathlaul, N.; Atif, M.; Eltayeb, L.B. Antioxidant and Prooxidant Functions of Carotenoids in Human Health: Trigger Factors, Mechanism and Application. In Recent Advances in Phytochemical Research; IntechOpen: London, UK, 2025. [Google Scholar]
- Khan, U.M.; Sevindik, M.; Zarrabi, A.; Nami, M.; Ozdemir, B.; Kaplan, D.N.; Selamoglu, Z.; Hasan, M.; Kumar, M.; Alshehri, M.M. Lycopene: Food sources, biological activities, and human health benefits. Oxidative Med. Cell. Longev. 2021, 2021, 2713511. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Redondo-Flórez, L.; Martín-Rodríguez, A.; Curiel-Regueros, A.; Rubio-Zarapuz, A.; Tornero-Aguilera, J.F. Impact of vegan and vegetarian diets on neurological health: A critical review. Nutrients 2025, 17, 884. [Google Scholar] [CrossRef]
- Nguyen, D.; Thrimawithana, T.; Piva, T.J.; Grando, D.; Huynh, T. Benefits of plant carotenoids against age-related macular degeneration. J. Funct. Foods 2023, 106, 105597. [Google Scholar] [CrossRef]
- Bekavac, N.; Krog, K.; Stanić, A.; Šamec, D.; Šalić, A.; Benković, M.; Jurina, T.; Gajdoš Kljusurić, J.; Valinger, D.; Jurinjak Tušek, A. Valorization of Food Waste: Extracting Bioactive Compounds for Sustainable Health and Environmental Solutions. Antioxidants 2025, 14, 714. [Google Scholar] [CrossRef] [PubMed]
- Jenča, A.; Mills, D.K.; Ghasemi, H.; Saberian, E.; Karimi Forood, A.M.; Petrášová, A.; Jenčová, J.; Jabbari Velisdeh, Z.; Zare-Zardini, H. Herbal therapies for cancer treatment: A review of phytotherapeutic efficacy. Biol. Targets Ther. 2024, 18, 229–255. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, A.; Kupczyński, R.; Gałęska, E.; Araujo, J.P.; Czerniawska-Piątkowska, E. Clinical application of bioextracts in supporting the reproductive system of animals and humans: Potential and Limitations. Evid.-Based Complement. Altern. Med. 2022, 2022, 4766409. [Google Scholar] [CrossRef]
- Yeung, A.W.K.; Solka, M.; Jóźwik, A.; Ksepka, N.; Matin, M.; Wang, D.; Zielińska, A.; Mohanasundaram, A.; Vejux, A.; Zarrouk, A. Anthocyanins-dietary natural products with a variety of bioactivities for the promotion of human and animal health. Anim. Sci. Pap. Rep. 2024, 42, 5–33. [Google Scholar] [CrossRef]
- Zhang, Y.-M.; Zhang, Z.-Y.; Wang, R.-X. Protective mechanisms of quercetin against myocardial ischemia reperfusion injury. Front. Physiol. 2020, 11, 956. [Google Scholar] [CrossRef]
- Aghababaei, F.; Hadidi, M. Recent advances in potential health benefits of quercetin. Pharmaceuticals 2023, 16, 1020. [Google Scholar] [CrossRef]
- Grinán-Ferré, C.; Bellver-Sanchis, A.; Izquierdo, V.; Corpas, R.; Roig-Soriano, J.; Chillón, M.; Andres-Lacueva, C.; Somogyvári, M.; Sőti, C.; Sanfeliu, C. The pleiotropic neuroprotective effects of resveratrol in cognitive decline and Alzheimer’s disease pathology: From antioxidant to epigenetic therapy. Ageing Res. Rev. 2021, 67, 101271. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Tang, P.; Lv, H. Targeting Oxidative Stress in Diabetic Retinopathy: Mechanisms, Pathology, and Novel Treatment Approaches. Front. Immunol. 2025, 16, 1571576. [Google Scholar] [CrossRef]
- Murakami, A. Impact of hormesis to deepen our understanding of the mechanisms underlying the bioactivities of polyphenols. Curr. Opin. Biotechnol. 2024, 86, 103074. [Google Scholar] [CrossRef]
- Simon, P.; Török, É.; Szalontai, K.; Kari, B.; Neuperger, P.; Zavala, N.; Kanizsai, I.; Puskás, L.G.; Török, S.; Szebeni, G.J. Nutritional Support of Chronic Obstructive Pulmonary Disease. Nutrients 2025, 17, 1149. [Google Scholar] [CrossRef]
- Barchielli, G.; Capperucci, A.; Tanini, D. The role of selenium in pathologies: An updated review. Antioxidants 2022, 11, 251. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Yu, P.; Chan, W.N.; Xie, F.; Zhang, Y.; Liang, L.; Leung, K.T.; Lo, K.W.; Yu, J.; Tse, G.M. Cellular zinc metabolism and zinc signaling: From biological functions to diseases and therapeutic targets. Signal Transduct. Target. Ther. 2024, 9, 6. [Google Scholar] [CrossRef]
- Costa, M.I.; Sarmento-Ribeiro, A.B.; Gonçalves, A.C. Zinc: From biological functions to therapeutic potential. Int. J. Mol. Sci. 2023, 24, 4822. [Google Scholar] [CrossRef]
- Liu, M.; Sun, X.; Chen, B.; Dai, R.; Xi, Z.; Xu, H. Insights into manganese superoxide dismutase and human diseases. Int. J. Mol. Sci. 2022, 23, 15893. [Google Scholar] [CrossRef]
- Munteanu, C.; Schwartz, B. Interactions between dietary antioxidants, dietary Fiber and the gut microbiome: Their putative role in inflammation and Cancer. Int. J. Mol. Sci. 2024, 25, 8250. [Google Scholar] [CrossRef] [PubMed]
- Salem, A.; Ammar, A.; Kerkeni, M.; Boujelbane, M.A.; Uyar, A.M.; Köbel, L.M.; Selvaraj, S.; Zare, R.; Heinrich, K.M.; Jahrami, H. Short-Term Beetroot Juice Supplementation Enhances Strength, Reduces Fatigue, and Promotes Recovery in Physically Active Individuals: A Randomized, Double-Blind, Crossover Trial. Nutrients 2025, 17, 1720. [Google Scholar] [CrossRef]
- Dominguez, L.J.; Di Bella, G.; Veronese, N.; Barbagallo, M. Impact of Mediterranean diet on chronic non-communicable diseases and longevity. Nutrients 2021, 13, 2028. [Google Scholar] [CrossRef]
- Helm, M.M.; Alaba, T.; Klimis-Zacas, D.; Izuora, K.; Basu, A. Effect of Dietary Berry Supplementation on Antioxidant Biomarkers in Adults with Cardiometabolic Risks: A Systematic Review of Clinical Trials. Antioxidants 2023, 12, 1182. [Google Scholar] [CrossRef]
- García-Cordero, J.; Martinez, A.; Blanco-Valverde, C.; Pino, A.; Puertas-Martín, V.; San Román, R.; de Pascual-Teresa, S. Regular consumption of cocoa and red berries as a strategy to improve cardiovascular biomarkers via modulation of microbiota metabolism in healthy aging adults. Nutrients 2023, 15, 2299. [Google Scholar] [CrossRef]
- O’Riordan, K.J.; Schneider, E.; Clarke, G.; Cryan, J.F. Diet and the Microbiome–Gut–Brain Axis: Feeding Your Microbes for Mental Health Benefit. In Nutritional Psychiatry: A Primer for Clinicians; Dinan, T., Ed.; Cambridge University Press: New York, NY, USA, 2023; pp. 15–38. [Google Scholar]
- Seminotti, B.; Grings, M.; Tucci, P.; Leipnitz, G.; Saso, L. Nuclear factor erythroid-2-related factor 2 signaling in the neuropathophysiology of inherited metabolic disorders. Front. Cell. Neurosci. 2021, 15, 785057. [Google Scholar] [CrossRef] [PubMed]
- Munteanu, C.; Galaction, A.I.; Onose, G.; Turnea, M.; Rotariu, M. The Janus Face of Oxidative Stress and Hydrogen Sulfide: Insights into Neurodegenerative Disease Pathogenesis. Antioxidants 2025, 14, 360. [Google Scholar] [CrossRef] [PubMed]
- Dera, A.A.; Al Fayi, M. CB5712809, a novel Keap1 inhibitor upregulates SQSTM1/p62 mediated Nrf2 activation to induce cell death in colon cancer cells. Discov. Oncol. 2025, 16, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.-T.; Wan, C.; Lin, J.-H.; Hammes, H.-P.; Zhang, C. Mitochondrial oxidative stress and cell death in podocytopathies. Biomolecules 2022, 12, 403. [Google Scholar] [CrossRef] [PubMed]
- Fantone, S.; Marzioni, D.; Tossetta, G. NRF2/KEAP1 signaling inhibitors in gynecologic cancers. Expert Rev. Anticancer. Ther. 2024, 24, 1191–1194. [Google Scholar] [CrossRef] [PubMed]
- Tossetta, G.; Fantone, S.; Marzioni, D.; Mazzucchelli, R. Cellular modulators of the NRF2/KEAP1 signaling pathway in prostate cancer. Front. Biosci. 2023, 28, 143. [Google Scholar] [CrossRef]
- Andrés, C.M.C.; Pérez de la Lastra, J.M.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Antioxidant metabolism pathways in vitamins, polyphenols, and selenium: Parallels and divergences. Int. J. Mol. Sci. 2024, 25, 2600. [Google Scholar] [CrossRef]
- Hashim, N.T.; Babiker, R.; Chaitanya, N.C.; Mohammed, R.; Priya, S.P.; Padmanabhan, V.; Ahmed, A.; Dasnadi, S.P.; Islam, M.S.; Gismalla, B.G. New Insights in Natural Bioactive Compounds for Periodontal Disease: Advanced Molecular Mechanisms and Therapeutic Potential. Molecules 2025, 30, 807. [Google Scholar] [CrossRef]
- Divyajanani, S.; Harithpriya, K.; Ganesan, K.; Ramkumar, K.M. Dietary polyphenols remodel DNA methylation patterns of NRF2 in chronic disease. Nutrients 2023, 15, 3347. [Google Scholar] [CrossRef]
- Požgajová, M.; Klongová, L.; Kovár, M.; Navrátilová, A. Cell Protection by Oxidative Stress Mitigation Using Substances with Bioactive Properties. In The Power of Antioxidants-Unleashing Nature’s Defense Against Oxidative Stress; IntechOpen: London, UK, 2024. [Google Scholar]
- Kilicarslan You, D.; Fuwad, A.; Lee, K.H.; Kim, H.K.; Kang, L.; Kim, S.M.; Jeon, T.-J. Evaluation of the Protective Role of Vitamin E against ROS-Driven Lipid Oxidation in Model Cell Membranes. Antioxidants 2024, 13, 1135. [Google Scholar] [CrossRef]
- Miazek, K.; Beton, K.; Śliwińska, A.; Brożek-Płuska, B. The effect of β-carotene, tocopherols and ascorbic acid as anti-oxidant molecules on human and animal in vitro/in vivo studies: A review of research design and analytical techniques used. Biomolecules 2022, 12, 1087. [Google Scholar] [CrossRef]
- Reed, D.J. Interaction of vitamin E, ascorbic acid, and glutathione in protection against oxidative damage. In Vitamin E in Health and Disease; CRC Press: Boca Raton, FL, USA, 2023; pp. 269–282. [Google Scholar]
- Kükürt, A.; Gelen, V. Understanding vitamin C: Comprehensive examination of its biological significance and antioxidant properties. In Ascorbic Acid-Biochemistry and Functions; IntechOpen: London, UK, 2024. [Google Scholar]
- Shen, Y.; Huang, H.; Wang, Y.; Yang, R.; Ke, X. Antioxidant effects of Se-glutathione peroxidase in alcoholic liver disease. J. Trace Elem. Med. Biol. 2022, 74, 127048. [Google Scholar] [CrossRef]
- Wang, X.; Qi, Y.; Zheng, H. Dietary polyphenol, gut microbiota, and health benefits. Antioxidants 2022, 11, 1212. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Si, H.; Jia, Z.; Liu, D. Dietary anti-aging polyphenols and potential mechanisms. Antioxidants 2021, 10, 283. [Google Scholar] [CrossRef]
- Kıran, T.R.; Otlu, O.; Karabulut, A.B. Oxidative stress and antioxidants in health and disease. J. Lab. Med. 2023, 47, 1–11. [Google Scholar] [CrossRef]
- Hofer, S.J.; Daskalaki, I.; Bergmann, M.; Friščić, J.; Zimmermann, A.; Mueller, M.I.; Abdellatif, M.; Nicastro, R.; Masser, S.; Durand, S. Spermidine is essential for fasting-mediated autophagy and longevity. Nat. Cell Biol. 2024, 26, 1571–1584. [Google Scholar] [CrossRef] [PubMed]
- Sartini, D.; Campagna, R.; Lucarini, G.; Pompei, V.; Salvolini, E.; Mattioli-Belmonte, M.; Molinelli, E.; Brisigotti, V.; Campanati, A.; Bacchetti, T. Differential immunohistochemical expression of paraoxonase-2 in actinic keratosis and squamous cell carcinoma. Hum. Cell 2021, 34, 1929–1931. [Google Scholar] [CrossRef]
- Cui, B.; Wang, Y.; Jin, J.; Yang, Z.; Guo, R.; Li, X.; Yang, L.; Li, Z. Resveratrol treats UVB-induced photoaging by anti-MMP expression, through anti-inflammatory, antioxidant, and antiapoptotic properties, and treats photoaging by upregulating VEGF-B expression. Oxidative Med. Cell. Longev. 2022, 2022, 6037303. [Google Scholar] [CrossRef] [PubMed]
- Torres-Contreras, A.M.; Garcia-Baeza, A.; Vidal-Limon, H.R.; Balderas-Renteria, I.; Ramírez-Cabrera, M.A.; Ramirez-Estrada, K. Plant secondary metabolites against skin photodamage: Mexican plants, a potential source of uv-radiation protectant molecules. Plants 2022, 11, 220. [Google Scholar] [CrossRef]
- Zhou, L.; Long, J.; Sun, Y.; Chen, W.; Qiu, R.; Yuan, D. Resveratrol ameliorates atherosclerosis induced by high-fat diet and LPS in ApoE−/− mice and inhibits the activation of CD4+ T cells. Nutr. Metab. 2020, 17, 41. [Google Scholar] [CrossRef]
- Chassot, L.N.; Scolaro, B.; Roschel, G.G.; Cogliati, B.; Cavalcanti, M.F.; Abdalla, D.S.; Castro, I.A. Comparison between red wine and isolated trans-resveratrol on the prevention and regression of atherosclerosis in LDLr(−/−) mice. J. Nutr. Biochem. 2018, 61, 48–55. [Google Scholar] [CrossRef]
- Phie, J.; Krishna, S.M.; Moxon, J.V.; Omer, S.M.; Kinobe, R.; Golledge, J. Flavonols reduce aortic atherosclerosis lesion area in apolipoprotein E deficient mice: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0181832. [Google Scholar] [CrossRef]
- Madokoro, Y.; Kamikokuryo, C.; Niiyama, S.; Ito, T.; Hara, S.; Ichinose, H.; Kakihana, Y. Early ascorbic acid administration prevents vascular endothelial cell damage in septic mice. Front. Pharmacol. 2022, 13, 929448. [Google Scholar] [CrossRef]
- Juraschek, S.P.; Guallar, E.; Appel, L.J.; Miller III, E.R. Effects of vitamin C supplementation on blood pressure: A meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2012, 95, 1079–1088. [Google Scholar] [CrossRef]
- Mohammadipoor, N.; Shafiee, F.; Rostami, A.; Kahrizi, M.S.; Soleimanpour, H.; Ghodsi, M.; Ansari, M.J.; Bokov, D.O.; Jannat, B.; Mosharkesh, E. Resveratrol supplementation efficiently improves endothelial health: A systematic review and meta-analysis of randomized controlled trials. Phytother. Res. 2022, 36, 3529–3539. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.; Luo, J.; Zhu, Y.; An, P.; Luo, Y.; Xing, Q. The effect of antioxidant polyphenol supplementation on cardiometabolic risk factors: A systematic review and meta-analysis. Nutrients 2024, 16, 4206. [Google Scholar] [CrossRef] [PubMed]
- Lagou, V.; Greyling, A.; Ferruzzi, M.G.; Skene, S.S.; Dubost, J.; Demirkan, A.; Prokopenko, I.; Shlisky, J.; Rodriguez-Mateos, A.; Heiss, C. Impact of flavan-3-ols on blood pressure and endothelial function in diverse populations: A systematic review and meta-analysis of randomized controlled trials. Eur. J. Prev. Cardiol. 2025, zwaf173. [Google Scholar] [CrossRef] [PubMed]
- Sesso, H.D.; Buring, J.E.; Christen, W.G.; Kurth, T.; Belanger, C.; MacFadyen, J.; Bubes, V.; Manson, J.E.; Glynn, R.J.; Gaziano, J.M. Vitamins E and C in the prevention of cardiovascular disease in men: The Physicians’ Health Study II randomized controlled trial. JAMA 2008, 300, 2123–2133. [Google Scholar] [CrossRef]
- Gaziano, J.M.; Glynn, R.J.; Christen, W.G.; Kurth, T.; Belanger, C.; MacFadyen, J.; Bubes, V.; Manson, J.E.; Sesso, H.D.; Buring, J.E. Vitamins E and C in the prevention of prostate and total cancer in men: The Physicians’ Health Study II randomized controlled trial. JAMA 2009, 301, 52–62. [Google Scholar] [CrossRef]
- Myung, S. Efficacy of vitamin and antioxidant supplements in prevention. Evid. Val. Health 2016, 2, 144–151. [Google Scholar]
- Schürks, M.; Glynn, R.J.; Rist, P.M.; Tzourio, C.; Kurth, T. Effects of vitamin E on stroke subtypes: Meta-analysis of randomised controlled trials. BMJ 2010, 341, c5702. [Google Scholar] [CrossRef]
- Garcia-Alloza, M.; Borrelli, L.; Rozkalne, A.; Hyman, B.; Bacskai, B. Curcumin labels amyloid pathology in vivo, disrupts existing plaques, and partially restores distorted neurites in an Alzheimer mouse model. J. Neurochem. 2007, 102, 1095–1104. [Google Scholar] [CrossRef]
- Asadbegi, M.; Komaki, H.; Faraji, N.; Taheri, M.; Safari, S.; Raoufi, S.; Kourosh-Arami, M.; Golipoor, Z.; Komaki, A. Effectiveness of coenzyme Q10 on learning and memory and synaptic plasticity impairment in an aged Aβ-induced rat model of Alzheimer’s disease: A behavioral, biochemical, and electrophysiological study. Psychopharmacology 2023, 240, 951–967. [Google Scholar] [CrossRef]
- Hosseini, L.; Majdi, A.; Sadigh-Eteghad, S.; Farajdokht, F.; Ziaee, M.; Aghsan, S.R.; Farzipour, M.; Mahmoudi, J. Coenzyme Q10 ameliorates aging-induced memory deficits via modulation of apoptosis, oxidative stress, and mitophagy in aged rats. Exp. Gerontol. 2022, 168, 111950. [Google Scholar] [CrossRef]
- Komaki, H.; Faraji, N.; Komaki, A.; Shahidi, S.; Etaee, F.; Raoufi, S.; Mirzaei, F. Investigation of protective effects of coenzyme Q10 on impaired synaptic plasticity in a male rat model of Alzheimer’s disease. Brain Res. Bull. 2019, 147, 14–21. [Google Scholar] [CrossRef]
- Shults, C.W.; Oakes, D.; Kieburtz, K.; Beal, M.F.; Haas, R.; Plumb, S.; Juncos, J.L.; Nutt, J.; Shoulson, I.; Carter, J. Effects of coenzyme Q10 in early Parkinson disease: Evidence of slowing of the functional decline. Arch. Neurol. 2002, 59, 1541–1550. [Google Scholar] [CrossRef]
- Müller, T.; Büttner, T.; Gholipour, A.-F.; Kuhn, W. Coenzyme Q10 supplementation provides mild symptomatic benefit in patients with Parkinson’s disease. Neurosci. Lett. 2003, 341, 201–204. [Google Scholar] [CrossRef]
- Yoritaka, A.; Kawajiri, S.; Yamamoto, Y.; Nakahara, T.; Ando, M.; Hashimoto, K.; Nagase, M.; Saito, Y.; Hattori, N. Randomized, double-blind, placebo-controlled pilot trial of reduced coenzyme Q10 for Parkinson’s disease. Park. Relat. Disord. 2015, 21, 911–916. [Google Scholar] [CrossRef]
- Beal, M.F.; Oakes, D.; Shoulson, I.; Henchcliffe, C.; Galpern, W.R.; Haas, R.; Juncos, J.L.; Nutt, J.G.; Voss, T.S.; Ravina, B. A randomized clinical trial of high-dosage coenzyme Q10 in early Parkinson disease: No evidence of benefit. JAMA Neurol. 2014, 71, 543–552. [Google Scholar] [PubMed]
- Farina, N.; Llewellyn, D.; Isaac, M.; Tabet, N. Vitamin E for Alzheimer’s dementia and mild cognitive impairment. Cochrane Database Syst. Rev. 2017, 4, CD002854. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Yu, S.; Yu, Y.; Xu, G. (−)-Epigallocatechin-3-gallate suppresses prostate cancer cell growth via activating miR-520a-3p. Rev. Bras. De Farmacogn. 2020, 30, 528–536. [Google Scholar] [CrossRef]
- Mokhtari, H.; Yaghmaei, B.; Sirati-Sabet, M.; Jafari, N.; Mardomi, A.; Abediankenari, S.; Mahrooz, A. Epigallocatechin-3-gallate Enhances the Efficacy of MicroRNA-34a Mimic and MicroRNA-93 Inhibitor Co-transfection in Prostate Cancer Cell Line. Iran. J. Allergy Asthma Immunol. 2020, 19, 612–623. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, F.; Chen, J. Application and potential value of curcumin in prostate cancer: A meta-analysis based on animal models. Front. Pharmacol. 2024, 15, 1379389. [Google Scholar] [CrossRef] [PubMed]
- Howells, L.; Malhotra Mukhtyar, R.; Theofanous, D.; Pepper, C.; Thomas, A.; Brown, K.; Khan, S. A systematic review assessing clinical utility of curcumin with a focus on cancer prevention. Mol. Nutr. Food Res. 2021, 65, 2000977. [Google Scholar] [CrossRef]
- Ahmad, T.; Khan, I.; Sultana, A.; Rizvi, M.M.A. A comparative study to evaluate the therapeutic effects of nutraceuticals in oral leukoplakia:-A randomized clinical trail. Natl. J. Maxillofac. Surg. 2023, 14, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Kuriakose, M.A.; Ramdas, K.; Dey, B.; Iyer, S.; Rajan, G.; Elango, K.K.; Suresh, A.; Ravindran, D.; Kumar, R.R.; Ramachandran, S. A randomized double-blind placebo-controlled phase IIB trial of curcumin in oral leukoplakia. Cancer Prev. Res. 2016, 9, 683–691. [Google Scholar] [CrossRef]
- Klein, E.A.; Thompson, I.M.; Tangen, C.M.; Crowley, J.J.; Lucia, M.S.; Goodman, P.J.; Minasian, L.M.; Ford, L.G.; Parnes, H.L.; Gaziano, J.M. Vitamin E and the risk of prostate cancer: The Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA 2011, 306, 1549–1556. [Google Scholar] [CrossRef]
- Khosravi, M.A.; Seifert, R. Clinical trials on curcumin in relation to its bioavailability and effect on malignant diseases: Critical analysis. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2024, 397, 3477–3491. [Google Scholar] [CrossRef]
- Gutsche, L.C.; Dörfler, J.; Hübner, J. Curcumin as a complementary treatment in oncological therapy: A systematic review. Eur. J. Clin. Pharmacol. 2025, 81, 1–33. [Google Scholar] [CrossRef]
- Li, X.; Matsumoto, T.; Takuwa, M.; Saeed Ebrahim Shaiku Ali, M.; Hirabashi, T.; Kondo, H.; Fujino, H. Protective effects of astaxanthin supplementation against ultraviolet-induced photoaging in hairless mice. Biomedicines 2020, 8, 18. [Google Scholar] [CrossRef]
- Komatsu, T.; Sasaki, S.; Manabe, Y.; Hirata, T.; Sugawara, T. Preventive effect of dietary astaxanthin on UVA-induced skin photoaging in hairless mice. PLoS ONE 2017, 12, e0171178. [Google Scholar] [CrossRef] [PubMed]
- Boyera, N.; Galey, I.; Bernard, B. Effect of vitamin C and its derivatives on collagen synthesis and cross-linking by normal human fibroblasts. Int. J. Cosmet. Sci. 1998, 20, 151–158. [Google Scholar] [CrossRef]
- Boo, Y.C. Ascorbic acid (vitamin C) as a cosmeceutical to increase dermal collagen for skin antiaging purposes: Emerging combination therapies. Antioxidants 2022, 11, 1663. [Google Scholar] [CrossRef]
- Humbert, P.G.; Haftek, M.; Creidi, P.; Lapière, C.; Nusgens, B.; Richard, A.; Schmitt, D.; Rougier, A.; Zahouani, H. Topical ascorbic acid on photoaged skin. Clinical, topographical and ultrastructural evaluation: Double-blind study vs. placebo. Exp. Dermatol. 2003, 12, 237–244. [Google Scholar] [CrossRef]
- Sauermann, K.; Jaspers, S.; Koop, U.; Wenck, H. Topically applied vitamin C increases the density of dermal papillae in aged human skin. BMC Dermatol. 2004, 4, 13. [Google Scholar] [CrossRef]
- Pullar, J.M.; Carr, A.C.; Vissers, M. The roles of vitamin C in skin health. Nutrients 2017, 9, 866. [Google Scholar] [CrossRef]
- Rouse, M.; Younès, A.; Egan, J.M. Resveratrol and curcumin enhance pancreatic β-cell function by inhibiting phosphodiesterase activity. J. Endocrinol. 2014, 223, 107. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.-J.; Zhang, H.; Song, D.-Q.; Xue, R.; Zhao, W.; Wei, J.; Wang, Y.-M.; Shan, N.; Zhou, Z.-X.; Yang, P. Berberine reduces insulin resistance through protein kinase C–dependent up-regulation of insulin receptor expression. Metabolism 2009, 58, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Asghari, K.M.; Saleh, P.; Salekzamani, Y.; Dolatkhah, N.; Aghamohammadzadeh, N.; Hashemian, M. The effect of curcumin and high-content eicosapentaenoic acid supplementations in type 2 diabetes mellitus patients: A double-blinded randomized clinical trial. Nutr. Diabetes 2024, 14, 14. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Feng, B.; Tian, Z. The effect of curcumin on lipid profile and glycemic status of patients with type 2 diabetes mellitus: A systematic review and meta-analysis. Evid. -Based Complement. Altern. Med. 2022, 2022, 8278744. [Google Scholar] [CrossRef]
- Khachik, F.; Bernstein, P.S.; Garland, D.L. Identification of lutein and zeaxanthin oxidation products in human and monkey retinas. Investig. Ophthalmol. Vis. Sci. 1997, 38, 1802–1811. [Google Scholar]
- Bian, Q.; Gao, S.; Zhou, J.; Qin, J.; Taylor, A.; Johnson, E.J.; Tang, G.; Sparrow, J.R.; Gierhart, D.; Shang, F. Lutein and zeaxanthin supplementation reduces photooxidative damage and modulates the expression of inflammation-related genes in retinal pigment epithelial cells. Free Radic. Biol. Med. 2012, 53, 1298–1307. [Google Scholar] [CrossRef]
- Chew, E.Y.; Clemons, T.E.; Agrón, E.; Domalpally, A.; Keenan, T.D.; Vitale, S.; Weber, C.; Smith, D.C.; Christen, W.; SanGiovanni, J.P. Long-term outcomes of adding lutein/zeaxanthin and ω-3 fatty acids to the AREDS supplements on age-related macular degeneration progression: AREDS2 report 28. JAMA Ophthalmol. 2022, 140, 692–698. [Google Scholar] [CrossRef]
- Li, S.-S.; Wang, H.-H.; Zhang, D. Efficacy of different nutrients in age-related macular degeneration: A systematic review and network meta-analysis. In Seminars in Ophthalmology; Taylor & Francis: Abingdon, UK, 2022; pp. 515–523. [Google Scholar]
- Group, A.-R.E.D.S.R. Lutein+ zeaxanthin and omega-3 fatty acids for age-related macular degeneration: The Age-Related Eye Disease Study 2 (AREDS2) randomized clinical trial. JAMA 2013, 309, 2005–2015. [Google Scholar]
- Ma, L.; Dou, H.-L.; Wu, Y.-Q.; Huang, Y.-M.; Huang, Y.-B.; Xu, X.-R.; Zou, Z.-Y.; Lin, X.-M. Lutein and zeaxanthin intake and the risk of age-related macular degeneration: A systematic review and meta-analysis. Br. J. Nutr. 2012, 107, 350–359. [Google Scholar] [CrossRef]
- Evans, J.R.; Lawrenson, J.G. Antioxidant vitamin and mineral supplements for slowing the progression of age-related macular degeneration. Cochrane Database Syst. Rev. 2023, 11, CD000254. [Google Scholar] [CrossRef]
- Group, A.-R.E.D.S.R.; Chew, E.Y.; SanGiovanni, J.P.; Ferris, F.L., III; Wong, W.T.; Agron, E.; Clemons, T.E.; Sperduto, R.; Danis, R.; Chandra, S.R. Lutein/zeaxanthin for the treatment of age-related cataract: AREDS2 randomized trial report no. 4. JAMA Ophthalmol. 2013, 131, 843–850. [Google Scholar]
- Clichici, S.; Olteanu, D.; Nagy, A.-L.; Oros, A.; Filip, A.; Mircea, P.A. Silymarin inhibits the progression of fibrosis in the early stages of liver injury in CCl4-treated rats. J. Med. Food 2015, 18, 290–298. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.-A.; Chen, G.-M.; Liu, Y.; Chen, Y.-X.; Wu, H.-Y.; Chen, J.; Xiong, Y.-L.; Tian, C.; Wang, G.-Y.; Jia, B. Inhibitory effect of silymarin on CCl4-induced liver fibrosis by reducing Ly6Chi monocytes infiltration. Int. J. Clin. Exp. Pathol. 2017, 10, 11941. [Google Scholar]
- Gharbia, S.; Balta, C.; Herman, H.; Rosu, M.; Váradi, J.; Bácskay, I.; Vecsernyés, M.; Gyöngyösi, S.; Fenyvesi, F.; Voicu, S.N. Enhancement of silymarin anti-fibrotic effects by complexation with hydroxypropyl (HPBCD) and randomly methylated (RAMEB) β-cyclodextrins in a mouse model of liver fibrosis. Front. Pharmacol. 2018, 9, 883. [Google Scholar] [CrossRef]
- Chalasani, N.; Sanyal, A.; Kowdley, K.; Robuck, P.; Hoofnagle, J.; Kleiner, D. Pioglitazone vs. vitamin E vs. placebo for treatment of non-diabetic patients with nonalcoholic steatohepatitis (PIVENS). N. Engl. J. Med. 2010, 362, 1675–1685. [Google Scholar]
- Sanyal, A.J.; Chalasani, N.; Kowdley, K.V.; McCullough, A.; Diehl, A.M.; Bass, N.M.; Neuschwander-Tetri, B.A.; Lavine, J.E.; Tonascia, J.; Unalp, A. Pioglitazone, vitamin E, or placebo for nonalcoholic steatohepatitis. N. Engl. J. Med. 2010, 362, 1675–1685. [Google Scholar] [CrossRef] [PubMed]
- Vadarlis, A.; Antza, C.; Bakaloudi, D.R.; Doundoulakis, I.; Kalopitas, G.; Samara, M.; Dardavessis, T.; Maris, T.; Chourdakis, M. Systematic review with meta-analysis: The effect of vitamin E supplementation in adult patients with non-alcoholic fatty liver disease. J. Gastroenterol. Hepatol. 2021, 36, 311–319. [Google Scholar] [CrossRef]
- Chee, N.M.Z.; Sinnanaidu, R.P.; Chan, W.K. Vitamin E improves serum markers and histology in adults with metabolic dysfunction-associated steatotic liver disease: Systematic review and meta-analysis. J. Gastroenterol. Hepatol. 2024, 39, 2545–2554. [Google Scholar] [CrossRef] [PubMed]
- Masoumi, M.; Salehi, M.; Angaji, S.A.; Hashemi, M. Effects of coenzyme Q10 and diamond nanoparticles on ischemia-reperfusion-induced testicular damages in rats. Galen Med. J. 2022, 10, e2029. [Google Scholar] [CrossRef] [PubMed]
- Said, R.S.; Mohamed, H.A.; Kamal, M.M. Coenzyme Q10 mitigates ionizing radiation-induced testicular damage in rats through inhibition of oxidative stress and mitochondria-mediated apoptotic cell death. Toxicol. Appl. Pharmacol. 2019, 383, 114780. [Google Scholar] [CrossRef] [PubMed]
- Li, K.-p.; Yang, X.-s.; Wu, T. The effect of antioxidants on sperm quality parameters and pregnancy rates for idiopathic male infertility: A network meta-analysis of randomized controlled trials. Front. Endocrinol. 2022, 13, 810242. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Qu, H.; Cao, Y.; Zhu, J.; Zhang, S.-z.; Wu, J.; Jiao, Y.-z. Effect of antioxidants on sperm quality parameters in subfertile men: A systematic review and network meta-analysis of randomized controlled trials. Adv. Nutr. 2022, 13, 586–594. [Google Scholar] [CrossRef]
- Showell, M.G.; Mackenzie-Proctor, R.; Jordan, V.; Hart, R.J. Antioxidants for female subfertility. Cochrane Database Syst. Rev. 2020, 8, CD007807. [Google Scholar] [CrossRef]
- Ntumy, M.; Maya, E.; Lizneva, D.; Adanu, R.; Azziz, R. The pressing need for standardization in epidemiologic studies of PCOS across the globe. Gynecol. Endocrinol. 2019, 35, 1–3. [Google Scholar] [CrossRef]
- Caro, A.A.; Bell, M.; Ejiofor, S.; Zurcher, G.; Petersen, D.R.; Ronis, M.J. N-acetylcysteine inhibits the up-regulation of mitochondrial biogenesis genes in livers from rats fed ethanol chronically. Alcohol. Clin. Exp. Res. 2014, 38, 2896–2906. [Google Scholar] [CrossRef]
- Kim, K.; Cort, T.A.; Kunz, E.M.; Moerschel, J.; Palzkill, V.R.; Dong, G.; Moparthy, C.N.; Anderson, E.M.; Fazzone, B.; O’Malley, K.A. N-acetylcysteine treatment attenuates hemodialysis access-related limb pathophysiology in mice with chronic kidney disease. Am. J. Physiol.-Ren. Physiol. 2023, 325, F271–F282. [Google Scholar] [CrossRef]
- Bolt, J.; Sandhu, S.; Mohammadi, A. Effect of coenzyme Q10 supplementation on sarcopenia, frailty, and falls: A scoping review. J. Nutr. Health Aging 2023, 27, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Petrofsky, J.; Laymon, M.; Khowailed, I.; Lee, H. Synergistic effects of continuous low level heat wraps and vitamins in improving balance and gait in adults. Int. J. Vitam. Nutr. Res. 2016, 86, 152–160. [Google Scholar] [CrossRef]
- Ganguly, A. Role of Jumpstart Nutrition®, a dietary supplement, to ameliorate calcium-to-Phosphorus ratio and parathyroid hormone of patients with osteoarthritis. Med. Sci. 2019, 7, 105. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Cabrera, M.-C.; Domenech, E.; Romagnoli, M.; Arduini, A.; Borras, C.; Pallardo, F.V.; Sastre, J.; Vina, J. Oral administration of vitamin C decreases muscle mitochondrial biogenesis and hampers training-induced adaptations in endurance performance. Am. J. Clin. Nutr. 2008, 87, 142–149. [Google Scholar] [CrossRef]
- Close, G.L.; Ashton, T.; Cable, T.; Doran, D.; Holloway, C.; McArdle, F.; MacLaren, D.P. Ascorbic acid supplementation does not attenuate post-exercise muscle soreness following muscle-damaging exercise but may delay the recovery process. Br. J. Nutr. 2006, 95, 976–981. [Google Scholar] [CrossRef]
- Braakhuis, A.J.; Hopkins, W.G.; Lowe, T.E. Effects of dietary antioxidants on training and performance in female runners. Eur. J. Sport Sci. 2014, 14, 160–168. [Google Scholar] [CrossRef]
- Bryant, R.J.; Ryder, J.; Martino, P.; Kim, J.; Craig, B.W. Effects of vitamin E and C supplementation either alone or in combination on exercise-induced lipid peroxidation in trained cyclists. J. Strength Cond. Res. 2003, 17, 792–800. [Google Scholar]
- Paulsen, G.; Cumming, K.T.; Hamarsland, H.; Børsheim, E.; Berntsen, S.; Raastad, T. Can supplementation with vitamin C and E alter physiological adaptations to strength training? BMC Sports Sci. Med. Rehabil. 2014, 6, 28. [Google Scholar] [CrossRef]
- Gliemann, L.; Schmidt, J.F.; Olesen, J.; Biensø, R.S.; Peronard, S.L.; Grandjean, S.U.; Mortensen, S.P.; Nyberg, M.; Bangsbo, J.; Pilegaard, H. Resveratrol blunts the positive effects of exercise training on cardiovascular health in aged men. J. Physiol. 2013, 591, 5047–5059. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C.; Henson, D.A.; McAnulty, S.R.; McAnulty, L.S.; Morrow, J.D.; Ahmed, A.; Heward, C.B. Vitamin E and immunity after the Kona Triathlon World Championship. Med. Sci. Sports Exerc. 2004, 36, 1328–1335. [Google Scholar] [CrossRef]
- Laaksonen, R.; Fogelholm, M.; Himberg, J.-J.; Laakso, J.; Salorinne, Y. Ubiquinone supplementation and exercise capacity in trained young and older men. Eur. J. Appl. Physiol. Occup. Physiol. 1995, 72, 95–100. [Google Scholar] [CrossRef]
- Parker, A.; Cuddihy, S.L.; Son, T.G.; Vissers, M.C.; Winterbourn, C.C. Roles of superoxide and myeloperoxidase in ascorbate oxidation in stimulated neutrophils and H2O2-treated HL60 cells. Free Radic. Biol. Med. 2011, 51, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Xu, X.; Zhang, K.; Ye, Q. The effect of intravenous vitamin C on clinical outcomes in patients with sepsis or septic shock: A meta-analysis of randomized controlled trials. Front. Nutr. 2022, 9, 964484. [Google Scholar] [CrossRef]
- Alissa, A.; Alrashed, M.A.; Alshaya, A.I.; Al Sulaiman, K.; Alharbi, S. Reevaluating vitamin C in sepsis and septic shock: A potential benefit in severe cases? Front. Med. 2024, 11, 1476242. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Chiu, C.-T.; Lee, H.-S.; Lai, C.-C. The impact of vitamin C-containing treatment on the mortality of patients with sepsis: A systematic review and meta-analysis of randomized controlled trials. J. Infect. Public Health 2022, 15, 1514–1520. [Google Scholar] [CrossRef] [PubMed]
- Inoue, R.; Miura, M.; Yanai, S.; Nishimune, H. Coenzyme Q10 supplementation improves the motor function of middle-aged mice by restoring the neuronal activity of the motor cortex. Sci. Rep. 2023, 13, 4323. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, N.; Hussain, H.K.; Rehman, Z.; Sabir, A.; Ashraf, W.; Ahmad, T.; Alqahtani, F.; Imran, I. Co-administration of coenzyme Q10 and curcumin mitigates cognitive deficits and exerts neuroprotective effects in aluminum chloride-induced Alzheimer’s disease in aged mice. Exp. Gerontol. 2025, 199, 112659. [Google Scholar] [CrossRef]
- Ahmadi-Soleimani, S.M.; Ghasemi, S.; Rahmani, M.A.; Gharaei, M.; Mohammadi Bezanaj, M.; Beheshti, F. Oral administration of coenzyme Q10 ameliorates memory impairment induced by nicotine-ethanol abstinence through restoration of biochemical changes in male rat hippocampal tissues. Sci. Rep. 2024, 14, 11413. [Google Scholar] [CrossRef]
- Rodríguez-Bies, E.; Tung, B.T.; Navas, P.; López-Lluch, G. Resveratrol primes the effects of physical activity in old mice. Br. J. Nutr. 2016, 116, 979–988. [Google Scholar] [CrossRef]
- Alehagen, U.; Alexander, J.; Aaseth, J. Supplementation with selenium and coenzyme Q10 reduces cardiovascular mortality in elderly with low selenium status. A secondary analysis of a randomised clinical trial. PLoS ONE 2016, 11, e0157541. [Google Scholar] [CrossRef]
- Dunning, B.J.; Bourgonje, A.R.; Bulthuis, M.L.; Alexander, J.; Aaseth, J.O.; Larsson, A.; van Goor, H.; Alehagen, U. Selenium and coenzyme Q10 improve the systemic redox status while reducing cardiovascular mortality in elderly population-based individuals. Free Radic. Biol. Med. 2023, 204, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Alehagen, U.; Aaseth, J.; Alexander, J.; Johansson, P.; Larsson, A. Supplemental selenium and coenzyme Q10 reduce glycation along with cardiovascular mortality in an elderly population with low selenium status–A four-year, prospective, randomised, double-blind placebo-controlled trial. J. Trace Elem. Med. Biol. 2020, 61, 126541. [Google Scholar] [CrossRef] [PubMed]
- Alehagen, U.; Aaseth, J.O.; Schomburg, L.; Opstad, T.B.; Larsson, A.; Alexander, J. Selenium and Coenzyme Q10 Supplementation and Sex Differences in Cardiovascular Mortality Results from a Prospective Randomized Double-Blind Placebo-Controlled Trial in Elderly People Low in Selenium. Antioxidants 2025, 14, 685. [Google Scholar] [CrossRef] [PubMed]
- Funk, J.L.; Frye, J.B.; Oyarzo, J.N.; Kuscuoglu, N.; Wilson, J.; McCaffrey, G.; Stafford, G.; Chen, G.; Lantz, R.C.; Jolad, S.D. Efficacy and mechanism of action of turmeric supplements in the treatment of experimental arthritis. Arthritis Rheum. Off. J. Am. Coll. Rheumatol. 2006, 54, 3452–3464. [Google Scholar] [CrossRef]
- Zhong, Y.; Tu, Y.; Ma, Q.; Chen, L.; Zhang, W.; Lu, X.; Yang, S.; Wang, Z.; Zhang, L. Corrigendum: Curcumin alleviates experimental colitis in mice by suppressing necroptosis of intestinal epithelial cells. Front. Pharmacol. 2023, 14, 1240661. [Google Scholar] [CrossRef]
- Yue, W.; Liu, Y.; Li, X.; Lv, L.; Huang, J.; Liu, J. Curcumin ameliorates dextran sulfate sodium-induced colitis in mice via regulation of autophagy and intestinal immunity. Turk. J. Gastroenterol. 2019, 30, 290. [Google Scholar] [CrossRef]
- Panahi, Y.; Hosseini, M.S.; Khalili, N.; Naimi, E.; Simental-Mendía, L.E.; Majeed, M.; Sahebkar, A. Effects of curcumin on serum cytokine concentrations in subjects with metabolic syndrome: A post-hoc analysis of a randomized controlled trial. Biomed. Pharmacother. 2016, 82, 578–582. [Google Scholar] [CrossRef]
- Belcaro, G.; Cesarone, M.R.; Dugall, M.; Pellegrini, L.; Ledda, A.; Grossi, M.G.; Togni, S.; Appendino, G. Efficacy and safety of Meriva®, a curcumin-phosphatidylcholine complex, during extended administration in osteoarthritis patients. Altern. Med. Rev. 2010, 15, 337–344. [Google Scholar]
- Zeng, L.; Yang, T.; Yang, K.; Yu, G.; Li, J.; Xiang, W.; Chen, H. Efficacy and safety of curcumin and curcuma longa extract in the treatment of arthritis: A systematic review and meta-analysis of randomized controlled trial. Front. Immunol. 2022, 13, 891822. [Google Scholar] [CrossRef]
- Li, C.; Chen, X.; Yao, J.; Zha, W.; Li, M.; Shen, J.; Jiang, H.; Tian, P. Curcumin modulated gut microbiota and alleviated renal fibrosis in 5/6 nephrectomy-induced chronic kidney disease rats. PLoS One 2025, 20, e0314029. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Lang, X.; Cheng, D.; Yang, Z. Curcumin ameliorates chronic renal failure in 5/6 nephrectomized rats by regulation of the mTOR/HIF-1α/VEGF signaling pathway. Biol. Pharm. Bull. 2019, 42, 886–891. [Google Scholar] [CrossRef]
- Jun, M.; Venkataraman, V.; Razavian, M.; Cooper, B.; Zoungas, S.; Ninomiya, T.; Webster, A.C.; Perkovic, V. Antioxidants for chronic kidney disease. Cochrane Database Syst. Rev. 2012, 10, CD008176. [Google Scholar] [CrossRef]
- Chiu, A.-H.; Wang, C.-J.; Lin, Y.-L.; Wang, C.-L.; Chiang, T.-I. N-Acetylcysteine Alleviates the Progression of Chronic Kidney Disease: A Three-Year Cohort Study. Medicina 2023, 59, 1983. [Google Scholar] [CrossRef]
- Ye, M.; Lin, W.; Zheng, J.; Lin, S. N-acetylcysteine for chronic kidney disease: A systematic review and meta-analysis. Am. J. Transl. Res. 2021, 13, 2472. [Google Scholar] [PubMed]
- Ahmadi, F.; Abbaszadeh, M.; Razeghi, E.; Maziar, S.; Khoidaki, S.D.; Najafi, M.T.; Lessan-Pezeshki, M. Effectiveness of N-acetylcysteine for preserving residual renal function in patients undergoing maintenance hemodialysis: Multicenter randomized clinical trial. Clin. Exp. Nephrol. 2017, 21, 342–349. [Google Scholar] [CrossRef] [PubMed]
- Colombijn, J.M.; Hooft, L.; Jun, M.; Webster, A.C.; Bots, M.L.; Verhaar, M.C.; Vernooij, R.W. Antioxidants for adults with chronic kidney disease. Cochrane Database Syst. Rev. 2023, 11, CD008176. [Google Scholar] [CrossRef]
- Xu, L.; Cai, B.-q.; Zhu, Y.-J. Pathogenesis of cigarette smoke-induced chronic obstructive pulmonary disease and therapeutic effects of glucocorticoids and N-acetylcysteine in rats. Chin. Med. J. 2004, 117, 1611–1619. [Google Scholar]
- Cai, S.; Chen, P.; Zhang, C.; CHEN, J.B.; Wu, J. Oral N-acetylcysteine attenuates pulmonary emphysema and alveolar septal cell apoptosis in smoking-induced COPD in rats. Respirology 2009, 14, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wiegman, C.; Seiffert, J.M.; Zhu, J.; Clarke, C.; Chang, Y.; Bhavsar, P.; Adcock, I.; Zhang, J.; Zhou, X. Effects of N-acetylcysteine in ozone-induced chronic obstructive pulmonary disease model. PLoS ONE 2013, 8, e80782. [Google Scholar] [CrossRef]
- Zhu, L.; Xu, F.; Kang, X.; Zhou, J.; Yao, Q.; Lin, Y.; Zhang, W. The antioxidant N-acetylcysteine promotes immune response and inhibits epithelial-mesenchymal transition to alleviate pulmonary fibrosis in chronic obstructive pulmonary disease by suppressing the VWF/p38 MAPK axis. Mol. Med. 2021, 27, 97. [Google Scholar] [CrossRef]
- Jiang, C.; Zou, J.; Lv, Q.; Yang, Y. Systematic review and meta-analysis of the efficacy of N-acetylcysteine in the treatment of acute exacerbation of chronic obstructive pulmonary disease. Ann. Palliat. Med. 2021, 10, 6564576–6566576. [Google Scholar] [CrossRef]
- Huang, C.; Kuo, S.; Lin, L.; Yang, Y. The efficacy of N-acetylcysteine in chronic obstructive pulmonary disease patients: A meta-analysis. Ther. Adv. Respir. Dis. 2023, 17, 17534666231158563. [Google Scholar] [CrossRef]
- De Backer, J.; Vos, W.; Van Holsbeke, C.; Vinchurkar, S.; Claes, R.; Parizel, P.M.; De Backer, W. Effect of high-dose N-acetylcysteine on airway geometry, inflammation, and oxidative stress in COPD patients. Int. J. Chronic Obstr. Pulm. Dis. 2013, 8, 569–579. [Google Scholar] [CrossRef]
- Guo, X.; Xu, Y.; Geng, R.; Qiu, J.; He, X. Curcumin alleviates dextran sulfate sodium-induced colitis in mice through regulating gut microbiota. Mol. Nutr. Food Res. 2022, 66, 2100943. [Google Scholar] [CrossRef]
- Lang, A.; Salomon, N.; Wu, J.C.; Kopylov, U.; Lahat, A.; Har-Noy, O.; Ching, J.Y.; Cheong, P.K.; Avidan, B.; Gamus, D. Curcumin in combination with mesalamine induces remission in patients with mild-to-moderate ulcerative colitis in a randomized controlled trial. Clin. Gastroenterol. Hepatol. 2015, 13, 1444–1449.e1441. [Google Scholar] [CrossRef] [PubMed]
- Holt, P.R.; Katz, S.; Kirshoff, R. Curcumin therapy in inflammatory bowel disease: A pilot study. Dig. Dis. Sci. 2005, 50, 2191–2193. [Google Scholar] [CrossRef]
- Shen, C.-L.; Yeh, J.K.; Cao, J.J.; Tatum, O.L.; Dagda, R.Y.; Wang, J.-S. Green tea polyphenols mitigate bone loss of female rats in a chronic inflammation-induced bone loss model. J. Nutr. Biochem. 2010, 21, 968–974. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.-L.; Smith, B.J.; Li, J.; Cao, J.J.; Song, X.; Newhardt, M.F.; Corry, K.A.; Tomison, M.D.; Tang, L.; Wang, J.-S. Effect of long-term green tea polyphenol supplementation on bone architecture, turnover, and mechanical properties in middle-aged ovariectomized rats. Calcif. Tissue Int. 2019, 104, 285–300. [Google Scholar] [CrossRef]
- Besora-Moreno, M.; Llauradó, E.; Valls, R.M.; Tarro, L.; Pedret, A.; Solà, R. Antioxidant-rich foods, antioxidant supplements, and sarcopenia in old-young adults ≥ 55 years old: A systematic review and meta-analysis of observational studies and randomized controlled trials. Clin. Nutr. 2022, 41, 2308–2324. [Google Scholar] [CrossRef] [PubMed]
- Nejadhosseinian, M.; Djalalinia, S.; Haerian, H.; Alikhani, M.; Mansour, A.; Mousavian, A.-H.; Mardani-Fard, H.A.; Kasaeian, A.; Faezi, S.T. The effects of antioxidants on knee osteoarthritis: A systematic review and meta-analysis. Front. Nutr. 2022, 9, 1026450. [Google Scholar] [CrossRef]
- Karimi, S.M.; Abbaspour, F.; Tabatabaei-Malazy, O.; Qolami, H.; Fayyaz, F.; Fana, S.E.; Rahimi, R.; Salari, P.; Larijani, B. Impact of Medicinal plants on Bone Health; a systematic review and meta-analysis of clinical studies. J. Agric. Food Res. 2024, 101545. [Google Scholar] [CrossRef]
- Austermann, K.; Baecker, N.; Zwart, S.R.; Fimmers, R.; Frippiat, J.-P.; Stehle, P.; Smith, S.M.; Heer, M. Antioxidant supplementation does not affect bone turnover markers during 60 days of 6 head-down tilt bed rest: Results from an exploratory randomized controlled trial. J. Nutr. 2021, 151, 1527–1538. [Google Scholar] [CrossRef]
- Li, Y.; Qi, H.; Huang, X.; Lu, G.; Pan, H. Exogenous and endogenous antioxidants in osteoporosis risk: Causal associations unveiled by Mendelian Randomization analysis. Front. Physiol. 2024, 15, 1411148. [Google Scholar] [CrossRef] [PubMed]
- Ashrafpour, S.; Ashrafpour, M. The double-edged sword of nutraceuticals: Comprehensive review of protective agents and their hidden risks. Front. Nutr. 2025, 12, 1524627. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Lykkesfeldt, J. On the effect of vitamin C intake on human health: How to (mis) interprete the clinical evidence. Redox Biol. 2020, 34, 101532. [Google Scholar] [CrossRef]
- Doseděl, M.; Jirkovský, E.; Macáková, K.; Krčmová, L.K.; Javorská, L.; Pourová, J.; Mercolini, L.; Remião, F.; Nováková, L.; Mladěnka, P. Vitamin C—sources, physiological role, kinetics, deficiency, use, toxicity, and determination. Nutrients 2021, 13, 615. [Google Scholar] [CrossRef]
- Guan, R.; Van Le, Q.; Yang, H.; Zhang, D.; Gu, H.; Yang, Y.; Sonne, C.; Lam, S.S.; Zhong, J.; Jianguang, Z. A review of dietary phytochemicals and their relation to oxidative stress and human diseases. Chemosphere 2021, 271, 129499. [Google Scholar] [CrossRef]
- Sahraeian, S.; Rashidinejad, A.; Golmakani, M.-T. Recent advances in the conjugation approaches for enhancing the bioavailability of polyphenols. Food Hydrocoll. 2024, 146, 109221. [Google Scholar] [CrossRef]
- Owen, K.N.; Dewald, O. Vitamin E Toxicity. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Bano, I.; Hassan, M.F.; Kieliszek, M. A Comprehensive Review of Selenium as a Key Regulator in Thyroid Health. Biol. Trace Elem. Res. 2025, 1–15. [Google Scholar] [CrossRef]
- Lian, W.; Yang, X.; Duan, Q.; Li, J.; Zhao, Y.; Yu, C.; He, T.; Sun, T.; Zhao, Y.; Wang, W. The Biological Activity of Ganoderma lucidum on Neurodegenerative Diseases: The Interplay between Different Active Compounds and the Pathological Hallmarks. Molecules 2024, 29, 2516. [Google Scholar] [CrossRef]
- Hasnain, B.I.; Mooradian, A.D. Recent trials of antioxidant therapy: What should we be telling our patients? Clevel. Clin. J. Med. 2004, 71, 327–334. [Google Scholar] [CrossRef]
- Steinhubl, S.R. Why have antioxidants failed in clinical trials? Am. J. Cardiol. 2008, 101, S14–S19. [Google Scholar] [CrossRef]
- Cook, N.R.; Albert, C.M.; Gaziano, J.M.; Zaharris, E.; MacFadyen, J.; Danielson, E.; Buring, J.E.; Manson, J.E. A randomized factorial trial of vitamins C and E and beta carotene in the secondary prevention of cardiovascular events in women: Results from the Women’s Antioxidant Cardiovascular Study. Arch. Intern. Med. 2007, 167, 1610–1618. [Google Scholar] [CrossRef]
- Shahidi, F.; Pan, Y. Influence of food matrix and food processing on the chemical interaction and bioaccessibility of dietary phytochemicals: A review. Crit. Rev. Food Sci. Nutr. 2022, 62, 6421–6445. [Google Scholar] [CrossRef] [PubMed]
- Middha, P.; Weinstein, S.J.; Männistö, S.; Albanes, D.; Mondul, A.M. β-carotene supplementation and lung cancer incidence in the alpha-tocopherol, beta-carotene cancer prevention study: The role of tar and nicotine. Nicotine Tob. Res. 2019, 21, 1045–1050. [Google Scholar] [CrossRef] [PubMed]
- Woldeselassie, M.; Tamene, A. Therapeutic controversies over use of antioxidant supplements during cancer treatment: A scoping review. Front. Nutr. 2024, 11, 1480780. [Google Scholar] [CrossRef] [PubMed]
- George, S.; Abrahamse, H. Redox potential of antioxidants in cancer progression and prevention. Antioxidants 2020, 9, 1156. [Google Scholar] [CrossRef]
- Calvani, M.; Pasha, A.; Favre, C. Nutraceutical boom in cancer: Inside the labyrinth of reactive oxygen species. Int. J. Mol. Sci. 2020, 21, 1936. [Google Scholar] [CrossRef]
- Talib, W.H.; Ahmed Jum’AH, D.A.; Attallah, Z.S.; Jallad, M.S.; Al Kury, L.T.; Hadi, R.W.; Mahmod, A.I. Role of vitamins A, C, D, E in cancer prevention and therapy: Therapeutic potentials and mechanisms of action. Front. Nutr. 2024, 10, 1281879. [Google Scholar] [CrossRef]
- Li, Y.; Lin, Q.; Lu, X.; Li, W. Post-diagnosis use of antioxidant vitamin supplements and breast cancer prognosis: A systematic review and meta-analysis. Clin. Breast Cancer 2021, 21, 477–485. [Google Scholar] [CrossRef]
- Ambrosone, C.B.; Zirpoli, G.R.; Hutson, A.D.; McCann, W.E.; McCann, S.E.; Barlow, W.E.; Kelly, K.M.; Cannioto, R.; Sucheston-Campbell, L.E.; Hershman, D.L. Dietary supplement use during chemotherapy and survival outcomes of patients with breast cancer enrolled in a cooperative group clinical trial (SWOG S0221). J. Clin. Oncol. 2020, 38, 804–814. [Google Scholar] [CrossRef]
- Kordiak, J.; Bielec, F.; Jabłoński, S.; Pastuszak-Lewandoska, D. Role of beta-carotene in lung cancer primary chemoprevention: A systematic review with meta-analysis and meta-regression. Nutrients 2022, 14, 1361. [Google Scholar] [CrossRef] [PubMed]
- Kaźmierczak-Barańska, J.; Boguszewska, K.; Adamus-Grabicka, A.; Karwowski, B.T. Two faces of vitamin C—Antioxidative and pro-oxidative agent. Nutrients 2020, 12, 1501. [Google Scholar] [CrossRef]
- Cobley, J.N.; McHardy, H.; Morton, J.P.; Nikolaidis, M.G.; Close, G.L. Influence of vitamin C and vitamin E on redox signaling: Implications for exercise adaptations. Free Radic. Biol. Med. 2015, 84, 65–76. [Google Scholar] [CrossRef]
- Li, S.; Fasipe, B.; Laher, I. Potential harms of supplementation with high doses of antioxidants in athletes. J. Exerc. Sci. Fit. 2022, 20, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Several lines of antioxidant defense against oxidative stress: Antioxidant enzymes, nanomaterials with multiple enzyme-mimicking activities, and low-molecular-weight antioxidants. Arch. Toxicol. 2024, 98, 1323–1367. [Google Scholar] [CrossRef] [PubMed]
- García-Sánchez, A.; Miranda-Díaz, A.G.; Cardona-Muñoz, E.G. The role of oxidative stress in physiopathology and pharmacological treatment with pro-and antioxidant properties in chronic diseases. Oxidative Med. Cell. Longev. 2020, 2020, 2082145. [Google Scholar] [CrossRef]
- Xu, Y.; Zheng, H.; Slabu, I.; Liehn, E.A.; Rusu, M. Vitamin C in cardiovascular disease: From molecular mechanisms to clinical evidence and therapeutic applications. Antioxidants 2025, 14, 506. [Google Scholar] [CrossRef]
- Yao, J.; Dai, X.; Yv, X.; Zheng, L.; Zheng, J.; Kuang, B.; Teng, W.; Yu, W.; Li, M.; Cao, H. The role of potential oxidative biomarkers in the Prognosis of Intracerebral Hemorrhage and the exploration antioxidants as possible Preventive and Treatment options. Front. Mol. Biosci. 2025, 12, 1541230. [Google Scholar] [CrossRef]
- Shen, Y.-J.; Huang, Y.-C.; Cheng, Y.-C. Advancements in Antioxidant-Based Therapeutics for Spinal Cord Injury: A Critical Review of Strategies and Combination Approaches. Antioxidants 2024, 14, 17. [Google Scholar] [CrossRef]
- Yusuf, A.; Almotairy, A.R.Z.; Henidi, H.; Alshehri, O.Y.; Aldughaim, M.S. Nanoparticles as drug delivery systems: A review of the implication of nanoparticles’ physicochemical properties on responses in biological systems. Polymers 2023, 15, 1596. [Google Scholar] [CrossRef]
- Martinelli, C.; Pucci, C.; Battaglini, M.; Marino, A.; Ciofani, G. Antioxidants and nanotechnology: Promises and limits of potentially disruptive approaches in the treatment of central nervous system diseases. Adv. Healthc. Mater. 2020, 9, 1901589. [Google Scholar] [CrossRef]
- Ammar, H.O.; Shamma, R.N.; Elbatanony, R.S.; Khater, B. Antioxidants in cancer therapy: Recent trends in application of nanotechnology for enhanced delivery. Sci. Pharm. 2020, 88, 5. [Google Scholar] [CrossRef]
- Sangiovanni, E.; Dell’Agli, M. Anti-Inflammatory Activity of Plant Polyphenols 2.0. Biomedicines 2021, 10, 37. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Palanisamy, C.P.; Natarajan, P.M.; Umapathy, V.R.; Roy, J.R.; Srinivasan, G.P.; Panagal, M.; Jayaraman, S. Curcumin-loaded polymeric nanomaterials as a novel therapeutic strategy for Alzheimer’s disease: A comprehensive review. Ageing Res. Rev. 2024, 99, 102393. [Google Scholar] [CrossRef]
- Bateni, Z.; Rahimi, H.R.; Hedayati, M.; Afsharian, S.; Goudarzi, R.; Sohrab, G. The effects of nano-curcumin supplementation on glycemic control, blood pressure, lipid profile, and insulin resistance in patients with the metabolic syndrome: A randomized, double-blind clinical trial. Phytother. Res. 2021, 35, 3945–3953. [Google Scholar] [CrossRef]
- Maniam, G.; Mai, C.-W.; Zulkefeli, M.; Dufès, C.; Tan, D.M.-Y.; Fu, J.-Y. Challenges and opportunities of nanotechnology as delivery platform for tocotrienols in cancer therapy. Front. Pharmacol. 2018, 9, 1358. [Google Scholar] [CrossRef]
- Chavda, V.P.; Nalla, L.V.; Balar, P.; Bezbaruah, R.; Apostolopoulos, V.; Singla, R.K.; Khadela, A.; Vora, L.; Uversky, V.N. Advanced phytochemical-based nanocarrier systems for the treatment of breast cancer. Cancers 2023, 15, 1023. [Google Scholar] [CrossRef] [PubMed]
- Sultana, A.; Zare, M.; Thomas, V.; Kumar, T.S.; Ramakrishna, S. Nano-based drug delivery systems: Conventional drug delivery routes, recent developments and future prospects. Med. Drug Discov. 2022, 15, 100134. [Google Scholar] [CrossRef]
- Fleming, E.; Luo, Y. Co-delivery of synergistic antioxidants from food sources for the prevention of oxidative stress. J. Agric. Food Res. 2021, 3, 100107. [Google Scholar] [CrossRef]
- Block, K.I.; Koch, A.C.; Mead, M.N.; Tothy, P.K.; Newman, R.A.; Gyllenhaal, C. Impact of antioxidant supplementation on chemotherapeutic efficacy: A systematic review of the evidence from randomized controlled trials. Cancer Treat. Rev. 2007, 33, 407–418. [Google Scholar] [CrossRef] [PubMed]
- Sepidarkish, M.; Akbari-Fakhrabadi, M.; Daneshzad, E.; Yavari, M.; Rezaeinejad, M.; Morvaridzadeh, M.; Heshmati, J. Effect of omega-3 fatty acid plus vitamin E Co-Supplementation on oxidative stress parameters: A systematic review and meta-analysis. Clin. Nutr. 2020, 39, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Boaru, D.L.; Fraile-Martinez, O.; De Leon-Oliva, D.; Garcia-Montero, C.; De Castro-Martinez, P.; Miranda-Gonzalez, A.; Saez, M.A.; Muñon-Zamarron, L.; Castillo-Ruiz, E.; Barrena-Blázquez, S. Harnessing the Anti-Inflammatory Properties of Polyphenols in the Treatment of Inflammatory Bowel Disease. Int. J. Biol. Sci. 2024, 20, 5608. [Google Scholar] [CrossRef]
- Gao, X.; Feng, X.; Hou, T.; Huang, W.; Ma, Z.; Zhang, D. The roles of flavonoids in the treatment of inflammatory bowel disease and extraintestinal manifestations: A review. Food Biosci. 2024, 62, 105431. [Google Scholar] [CrossRef]
- Nikic, P.; Dragicevic, D.; Savic-Radojevic, A.; Pljesa-Ercegovac, M.; Coric, V.; Jovanovic, D.; Bumbasirevic, U.; Pekmezovic, T.; Simic, T.; Dzamic, Z. Association between GPX1 and SOD2 genetic polymorphisms and overall survival in patients with metastatic urothelial bladder cancer: A single-center study in Serbia. J. BUON 2018, 23, 1130–1135. [Google Scholar]
- Nikic, P.; Dragicevic, D.; Jerotic, D.; Savic, S.; Djukic, T.; Stankovic, B.; Kovacevic, L.; Simic, T.; Matic, M. Polymorphisms of antioxidant enzymes SOD2 (rs4880) and GPX1 (rs1050450) are associated with bladder cancer risk or its aggressiveness. Medicina 2023, 59, 131. [Google Scholar] [CrossRef]
- Rossman, M.J.; Santos-Parker, J.R.; Steward, C.A.; Bispham, N.Z.; Cuevas, L.M.; Rosenberg, H.L.; Woodward, K.A.; Chonchol, M.; Gioscia-Ryan, R.A.; Murphy, M.P. Chronic supplementation with a mitochondrial antioxidant (MitoQ) improves vascular function in healthy older adults. Hypertension 2018, 71, 1056–1063. [Google Scholar] [CrossRef] [PubMed]
- Looi, D.; Moorthy, M.; Chaiyakunapruk, N.; Palanisamy, U.D. Impact of ellagitannin-rich fruit consumption on blood pressure: A systematic review and meta-analysis of randomized controlled trials. J. Funct. Foods 2022, 99, 105320. [Google Scholar] [CrossRef]
- Banc, R.; Rusu, M.E.; Filip, L.; Popa, D.-S. The impact of ellagitannins and their metabolites through gut microbiome on the gut health and brain wellness within the gut–brain axis. Foods 2023, 12, 270. [Google Scholar] [CrossRef]
- Tragni, V.; Primiano, G.; Tummolo, A.; Cafferati Beltrame, L.; La Piana, G.; Sgobba, M.N.; Cavalluzzi, M.M.; Paterno, G.; Gorgoglione, R.; Volpicella, M. Personalized medicine in mitochondrial health and disease: Molecular basis of therapeutic approaches based on nutritional supplements and their analogs. Molecules 2022, 27, 3494. [Google Scholar] [CrossRef] [PubMed]
- Tretter, V.; Hochreiter, B.; Zach, M.L.; Krenn, K.; Klein, K.U. Understanding Cellular Redox Homeostasis: A Challenge for Precision Medicine. Int. J. Mol. Sci. 2022, 23, 106. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.M.; Lee, C.M.; Saunders, D.; Curtis, A.E.; Dunlap, N.E.; Nangia, C.; Lee, A.S.; Kovoor, P.; Bar-Ad, V.; Pedadda, A.V., Jr. Two-year tumor outcomes of a phase 2B, randomized, double-blind trial of avasopasem manganese (GC4419) versus placebo to reduce severe oral mucositis owing to concurrent radiation therapy and cisplatin for head and neck cancer. Int. J. Radiat. Oncol. Biol. Phys. 2022, 114, 416–421. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Hegde, M.; Parama, D.; Girisa, S.; Kumar, A.; Daimary, U.D.; Garodia, P.; Yenisetti, S.C.; Oommen, O.V.; Aggarwal, B.B. Role of turmeric and curcumin in prevention and treatment of chronic diseases: Lessons learned from clinical trials. ACS Pharmacol. Transl. Sci. 2023, 6, 447–518. [Google Scholar] [CrossRef]
- Kil, J.; Lobarinas, E.; Spankovich, C.; Griffiths, S.K.; Antonelli, P.J.; Lynch, E.D.; Le Prell, C.G. Safety and efficacy of ebselen for the prevention of noise-induced hearing loss: A randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2017, 390, 969–979. [Google Scholar] [CrossRef]
- Sharpley, A.L.; Williams, C.; Holder, A.A.; Godlewska, B.R.; Singh, N.; Shanyinde, M.; MacDonald, O.; Cowen, P.J. A phase 2a randomised, double-blind, placebo-controlled, parallel-group, add-on clinical trial of ebselen (SPI-1005) as a novel treatment for mania or hypomania. Psychopharmacology 2020, 237, 3773–3782. [Google Scholar] [CrossRef]
- Wang, J.; Wang, P.; Dong, C.; Zhao, Y.; Zhou, J.; Yuan, C.; Zou, L. Mechanisms of ebselen as a therapeutic and its pharmacology applications. Future Med. Chem. 2020, 12, 2141–2160. [Google Scholar] [CrossRef]
- Zhou, S.; Cai, H.; He, X.; Tang, Z.; Lu, S. Enzyme-mimetic antioxidant nanomaterials for ROS scavenging: Design, classification, and biological applications. Coord. Chem. Rev. 2024, 500, 215536. [Google Scholar] [CrossRef]
- Zhang, S. Current development on wearable biosensors towards biomedical applications. J. Agric. Food Res. 2023, 11, 1264337. [Google Scholar] [CrossRef]
- Kim, J.; Campbell, A.S.; de Ávila, B.E.-F.; Wang, J. Wearable biosensors for healthcare monitoring. Nat. Biotechnol. 2019, 37, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Fan, R.; Zhang, Y.; Jia, Z.; Zhang, J.; Pan, H.; Wang, Q. Oxidative stress in the brain–lung crosstalk: Cellular and molecular perspectives. Front. Aging Neurosci. 2024, 16, 1389454. [Google Scholar] [CrossRef] [PubMed]
- Nuñez-Selles, A.J.; Nuñez-Musa, R.A.; Guillen-Marmolejos, R.A. Linking oxidative stress biomarkers to disease progression and antioxidant therapy in hypertension and diabetes mellitus. Front. Mol. Biosci. 2025, 12, 1611842. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yang, Y.; Min, J.; Song, Y.; Tu, J.; Mukasa, D.; Ye, C.; Xu, C.; Heflin, N.; McCune, J.S. A wearable electrochemical biosensor for the monitoring of metabolites and nutrients. Nat. Biomed. Eng. 2022, 6, 1225–1235. [Google Scholar] [CrossRef] [PubMed]
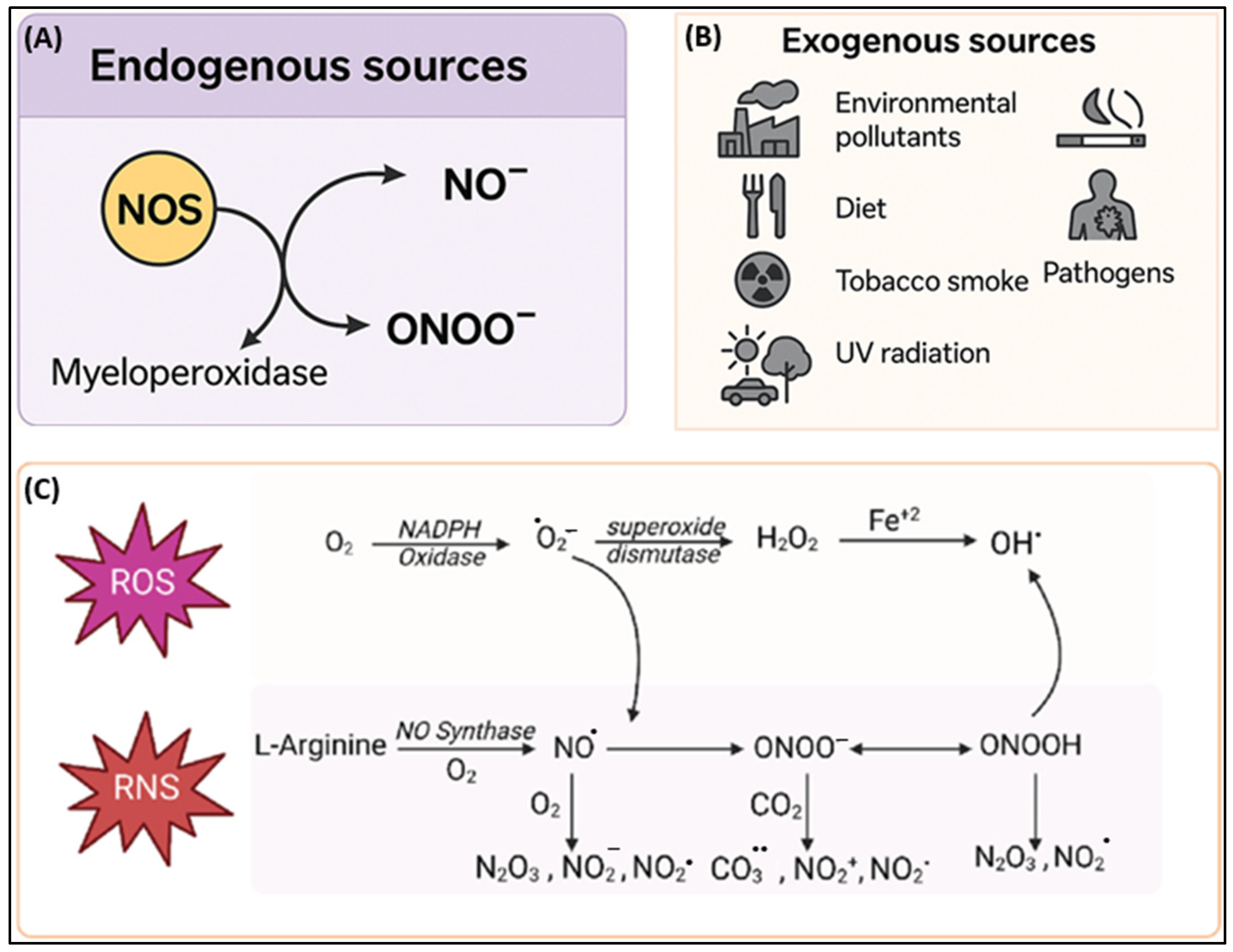
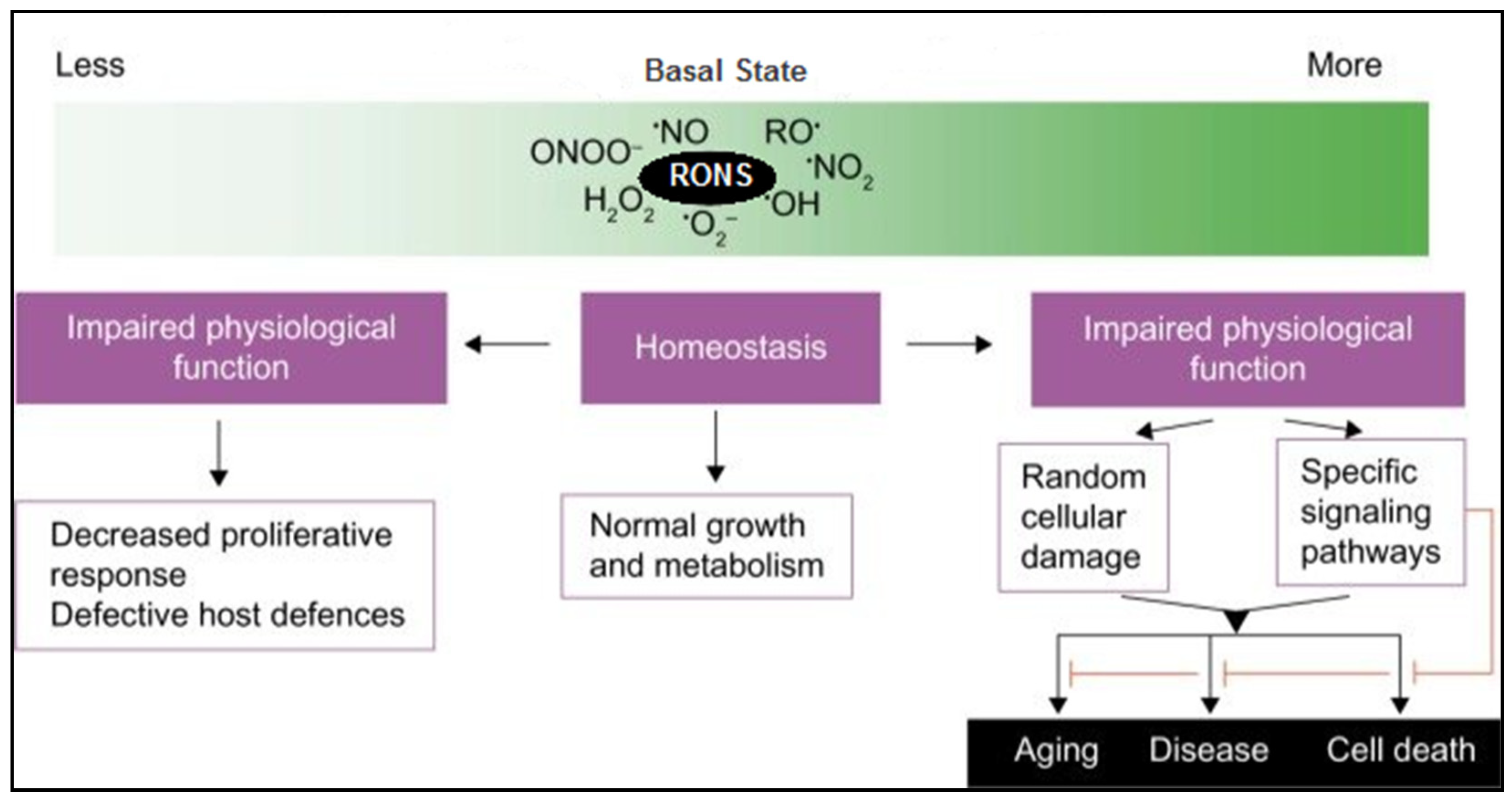

{kind=link}
{kind=link}
{kind=link}
| Radicals | Formula | Non-Radicals | Formula |
|---|---|---|---|
| Superoxide anion * | O2•− | Hydrogen peroxide * | H2O2 |
| Hydroxyl radical * | •OH | Singlet oxygen * | 1O2 |
| Nitric oxide † | •NO | Ozone/trioxygen * | O3 |
| Nitrogen dioxide † | •NO2 | Organic hydroperoxides * | ROOH |
| Organic radicals | R• | Hypochlorite * | ClO− |
| Peroxyl radicals * | ROO• | Hypochlorous acid * | HOCl |
| Alkoxyl radicals * | RO• | Peroxynitrite † | ONOO− |
| Thiyl radicals | RS• | Akyl peroxynitrite † | ROONO |
| Sulfonyl radicals | ROS• | Nitrosoperoxycarbonate anion † | O=NOOCO2− |
| Thiyl peroxyl radicals * | RSOO• | Nitrocarbonate anion † | O2NOCO2− |
| Hydroperoxyl * | HOO• | Dinitrogen dioxide † | N2O2 |
| Nitrogen dioxide † | •NOO | Nitronium † | NO2+ |
| Dinitrogen trioxide † | N2O3 | ||
| Nitrous acid † | HNO2 | ||
| Nitroxyl anion † | NO− | ||
| Nitrosyl cation † | NO+ | ||
| Nitryl chloride † | NOCl | ||
| Dinitrogen tetraoxide † | N2O4 | ||
| Disulfides | RSSR |
| Disease Category | Specific Diseases | Role of Oxidative/Nitrosative Stress | Study Limitations | References |
|---|---|---|---|---|
| Cardiovascular Diseases | Atherosclerosis, Hypertension, Heart Failure | Oxidative stress induces LDL oxidation, leading to foam cell formation and plaque buildup. ROS/RNS impair nitric oxide (NO) signaling, causing endothelial dysfunction and vascular tone dysregulation. In heart failure, oxidative/nitrosative stress promotes myocardial remodeling and apoptosis. | Predominantly based on animal or in vitro models; causal pathways in humans remain underexplored. | [82,83,84] |
| Neurodegenerative Diseases | Alzheimer’s Disease, Parkinson’s Disease, Huntington’s Disease | ROS and RNS contribute to neurotoxicity by inducing lipid peroxidation, protein aggregation (e.g., amyloid-beta, alpha-synuclein), and mitochondrial dysfunction. These species activate microglia, leading to chronic neuroinflammation and synaptic loss. | Translational challenges from animal models to humans; complexity of disease progression limits mechanistic clarity. | [85,86,87,88] |
| Cancers | Lung, Breast, Colon, Prostate, etc. | Oxidative and nitrosative stress contribute to tumor initiation by inducing DNA mutations and genomic instability. They also activate pro-oncogenic transcription factors (e.g., NF-κB, AP-1) and promote angiogenesis and tumor progression. | Multifactorial nature of cancer makes it difficult to attribute causality to oxidative stress alone. | [89] |
| Diabetes | Type 1 and Type 2 Diabetes | In type 1 diabetes, ROS mediate autoimmune destruction of pancreatic beta cells. In type 2 diabetes, oxidative stress impairs insulin signaling, enhances adipose tissue inflammation, and contributes to endothelial dysfunction. | Limited longitudinal human studies; antioxidant therapy trials yield inconsistent results. | [90,91] |
| Chronic Inflammatory Diseases | Rheumatoid Arthritis, Inflammatory Bowel Disease (IBD) | ROS and RNS sustain chronic inflammation by activating NF-κB and MAPK pathways, leading to cytokine overproduction and tissue damage in joints or the gastrointestinal tract. They also perpetuate immune dysregulation. | Confounding from co-existing inflammatory processes; lack of robust antioxidant-targeted clinical trials. | [92,93,94] |
| Respiratory Diseases | COPD, Asthma | Inhaled pollutants and allergens enhance ROS/RNS generation, which triggers epithelial injury, mucus hypersecretion, airway remodeling, and inflammation. NO-derived species exacerbate bronchoconstriction and hyperreactivity. | Evidence largely from animal models and ex vivo studies; need for large-scale human trials. | [95,96,97] |
| Renal Diseases | Chronic Kidney Disease (CKD), Acute Kidney Injury (AKI) | ROS and RNS mediate tubular cell apoptosis, glomerular injury, and interstitial fibrosis. They impair mitochondrial function and activate profibrotic signaling pathways (e.g., TGF-β). | Lack of standardization in oxidative stress biomarkers; heterogeneous patient populations in studies. | [98,99] |
| Age-Related Diseases | Macular Degeneration, Sarcopenia | Accumulated oxidative damage to retinal pigment epithelium (in AMD) and skeletal muscle proteins (in sarcopenia) disrupts tissue integrity and accelerates functional decline with age. | Multifactorial influences of aging (e.g., metabolic, genetic) complicate interpretation of oxidative stress roles. | [100,101] |
| Liver Diseases | Non-alcoholic liver disease (NAFLD), Cirrhosis, Hepatitis | Hepatic steatosis and inflammation are driven by lipid peroxidation and mitochondrial ROS/RNS production. Oxidative stress triggers stellate cell activation, leading to fibrosis and cirrhosis. | Few well-powered clinical trials evaluating antioxidant interventions; heterogeneity in liver disease subtypes. | [102] |
| Muscle Diseases | Muscular Dystrophy, Sarcopenia | ROS and RNS disrupt calcium homeostasis, impair mitochondrial respiration, and induce muscle fiber necrosis. Chronic oxidative stress leads to muscle wasting and fatigue. | Studies limited by small sample sizes and short durations; genetic variability complicates interpretation. | [103,104] |
| Infectious Diseases | HIV, Sepsis | Overproduction of ROS/RNS during infection leads to oxidative damage of immune and endothelial cells, contributing to immunosuppression and multiorgan failure. Nitrosative stress disrupts mitochondrial and barrier function. | Difficult to isolate oxidative stress from pathogen-induced effects; clinical data remain inconclusive. | [105,106] |
| Autoimmune Diseases | Systemic Lupus Erythematosus (SLE), Multiple Sclerosis | ROS and RNS act as secondary messengers that enhance autoreactive immune responses, promote DNA fragmentation, and damage target organs such as the kidneys (SLE) or CNS (MS). | Mechanistic overlap between immune activation and oxidative stress; lack of validated redox biomarkers. | [104,107,108] |
| Antioxidant Class | Representative Examples | Site of Action | Primary Mechanisms of Action |
|---|---|---|---|
| Endogenous Antioxidants | |||
| Enzymatic | Superoxide Dismutase (SOD1, SOD2, SOD3) | Cytosol (SOD1), mitochondria (SOD2), extracellular space (SOD3) | Converts superoxide (O2•−) to hydrogen peroxide (H2O2) |
| Catalase (CAT) | Peroxisomes | Decomposes H2O2 into water and oxygen | |
| Glutathione Peroxidase (GPx) | Cytosol and mitochondria | Reduces H2O2 and lipid hydroperoxides using glutathione (GSH) | |
| Paraoxonase-2 (PON2) | Mitochondrial and plasma membranes | Inhibits lipid peroxidation and modulates redox signaling | |
| Non-enzymatic | Glutathione (GSH) | Cytosol, nucleus, mitochondria | Reduces peroxides and regenerates oxidized vitamins C and E |
| Uric Acid | Plasma, extracellular fluid | Scavenges singlet oxygen and peroxynitrite; protects against lipid and DNA oxidation | |
| Bilirubin | Plasma, liver, brain | Scavenges peroxyl radicals; anti-inflammatory and neuroprotective functions | |
| Coenzyme Q10 (Ubiquinone) | Mitochondrial membranes | Prevents lipid peroxidation in mitochondrial inner membrane; regenerates vitamin E | |
| Exogenous Antioxidants | |||
| Vitamins | Vit C (Ascorbic acid) | Cytosol, plasma | Neutralizes ROS (•OH, ROO•); regenerates vitamin E and GSH |
| Vit E (α-Tocopherol) | Lipid membranes | Terminates lipid peroxidation chain reactions; stabilizes membrane integrity | |
| Vit A | Eyes, skin, adipose tissue | Quench singlet oxygen; protect against UV- and light-induced oxidative damage | |
| Polyphenols | Quercetin, Resveratrol, EGCG | Cytosol, nucleus | Scavenge ROS/RNS; inhibit pro-inflammatory enzymes; activate Nrf2 signaling |
| Carotenoids | β-Carotene, Lutein, Zeaxanthin, Lycopene | Retina, skin | Absorb blue light; quench singlet oxygen and prevent lipid oxidation |
| Trace Minerals | Selenium | All tissues (especially liver and kidney) | Cofactor for GPx and thioredoxin reductase; supports peroxide detoxification |
| Zinc | Cytosol, membranes | Cofactor for Cu/Zn-SOD; stabilizes proteins and membranes; inhibits NADPH oxidase | |
| Manganese | Mitochondria | Cofactor for Mn-SOD; prevents mitochondrial oxidative damage | |
| Condition | Antioxidant | Mechanism | Disease | Preclinical Outcomes | Clinical Outcomes | Inconsistent/Limited Outcome |
|---|---|---|---|---|---|---|
| Cardiovascular System | Vit C, Vit E, Flavonoids, Resveratrol | Inhibit LDL oxidation, boost NO•, reduce vascular inflammation | Atherosclerosis, hypertension, endothelial dysfunction | Resveratrol reduced atherosclerosis induced by HFD in mice [188] Resveratrol reduced oxidative stress in mice submitted to atherogenic diet leading to CV protection [189]. Meta-analysis: Flavonols reduced atherosclerotic lesion in apolipoprotein E-deficient mice [190]. Vit C suppressed vascular endothelial cell damage; improved endothelial function in adult septic mice [191]. | Vit C reduced blood pressure in short-term RCT [192]. Meta-analysis: Improved endothelial function and cardiovascular health with resveratrol [193]. RCT: Antioxidant polyphenol [194] and flavan-3-ol-rich food [195] supplementation reduced oxidative stress and improved CV health. | Meta-analyses: No evidence in support of vitamin and antioxidant supplements for prevention of CVDs [196,197,198]. Meta-analysis: High-dose Vit E supplementation linked to hemorrhagic stroke [199]. |
| Neurodegeneration | Curcumin, CoQ10, Resveratrol, Melatonin, Vit E | Scavenge ROS, stabilize mitochondria, reduce protein aggregation | AD, PD, HD | Curcumin reduced and reversed existing amyloid pathology and associated neurotoxicity in mouse model of AD [200]. CoQ10 suppressed progression of neuro-degeneration and restored synaptic plasticity and mitochondrial function in aged β-amyloid (Aβ)-induced AD rats [201,202,203]. | CoQ10 slowed PD progression in early-stage RCT [204,205]. RCT: Ubiquinol-10 (reduced form of CoQ10) conferred motor benefit in early PD [206]. | Meta-analysis: No cognitive benefit or prevention of dementia in AD with Vit E supplementation due to low CNS bioavailability [207,208]. |
| Cancer Prevention | EGCG, Curcumin, Sulforaphane, Selenium | Support detox enzymes, induce apoptosis, inhibit angiogenesis | Breast, prostate, colon, lung | EGCG suppressed tumor growth and prostate cancer in xenografts [209,210]. Meta-analyses: Curcumin inhibited prostate cancer growth in animal models [211,212]. | RCT: Curcumin effective for treatment of oral leukoplakia [213]. RCT: Curcumin effective for treating oral leukoplakia [214]. | RCT (SELECT): Increased prostate cancer risk with selenium and vitamin E [215]. Meta-analysis: Curcumin had no significant/positive effect on the therapy of malignant diseases due to limited bioavailability [216,217]. |
| Skin Health | Vit C, Vit E, Astaxanthin (AST), CoQ10, Polyphenols | Scavenge UV-ROS, enhance collagen, reduce matrix metalloproteinases (MMP) activity | Photoaging, wrinkles, hyperpigmentation | AST reduced/ prevented UV-induced photoaging and wrinkles in skin of hairless mice [218,219] Vit C exerted anti-aging properties on human fibroblast cells [220]. Vit C improved dermal collagen in hairless skin mice [221]. | Topical Vit C improved photodamaged skin and ultrastructure in RCT [222]. Topical application of Vit C in RCT significantly increased the density of dermal papillae in aged human skin [223]. | Inconsistent outcomes in meta-analysis due to variable formulations and limited transdermal absorption [224]. |
| Metabolic Disorders | Resveratrol, Curcumin, Berberine, Quercetin | Enhance insulin signaling, reduce β-cell apoptosis, inhibit AGEs | T2DM, diabetic neuropathy | Curcumin and resveratrol enhanced pancreatic β-cell function [225]. Berberine lowered FPG and improved insulin sensitivity and glucose metabolism in HFD rats [226]. | Curcumin adjunct therapy in RCT improved HbA1c by 0.4% over 12 weeks [227]. | Meta-analysis: Mild HbA1c reduction (<0.5%) from curcumin supplementation with significant heterogeneity [228]. |
| Ocular Health | Lutein, Zeaxanthin, Vit C, Vit E, Zinc, Vit A | Filter blue light, protect RPE, reduce lipid peroxidation | AMD, cataracts | Lutein and zeaxanthin delayed AMD progression and protected photoreceptors in light-exposed RPE cell models [229,230]. | RCT: AREDS2 slight reduction in AMD progression from lutein and zeaxanthin [231]. Network meta-analysis supports efficacy in early AMD by improving visual acuity [232]. | RCTs: No significant reduction in early AMD with dietary lutein and zeaxanthin [233,234]. Large long-term RCT (AREDS2): No benefit in advanced cataracts from lutein and zeaxanthin [235,236]. RCT (AREDS2), dietary lutein, and zeaxanthin effective only in early/intermediate AMD [236]. |
| Liver Diseases | Silymarin, NAC, Vit E, Glutathione | Restore mitochondrial redox, inhibit lipid peroxidation | NAFLD, NASH, hepatitis | Silymarin reduced hepatic inflammation and fibrosis in NAFLD in CCl4-induced injury mouse models [237,238,239]. | RCT: Vit E effective treating NASH in adults without diabetes [240]. Meta-analyses: Vit E effective for treatment of NASH and NAFLD in adult and pediatric patients [139,241,242,243]. | Large long-term RCT (SELECT): Increased prostate cancer risk with selenium and vit E with modest drop in hepatic inflammation [215]. |
| Reproductive Health | Vit E, CoQ10, Zinc, L-Carnitine | Reduce sperm DNA damage, supports oocyte quality, enhance mitochondrial energetics | Male/female infertility, PCOS | CoQ10 conserved against oxidative stress injury in germinal cells of rats. CoQ10 improved sperm count and motility in rats [244,245]. | Meta-analysis: CoQ10 improved sperm motility, concentration, and morphology [246,247]. | Meta-analysis: Limited evidence in support of supplemental oral antioxidants for subfertile women; non-significant live birth rate improvements [248]. PCOS studies underpowered due to lack of standardization of methodologies across phenotypes [249] |
| Muscle Physiology | NAC, Vit C, Vit E, Polyphenols, CoQ10 | Scavenge exercise-induced ROS, support mitochondrial biogenesis | Muscle fatigue, f | NAC reduced neuromotor pathology, improved endurance and mitochondrial markers in ethanol and adenine fed mice [250,251]. | RCTs: CoQ10 improved physical robustness, balance, gait speed, and symmetry [252,253,254]. | High-dose antioxidants blunt training adaptation in RCTs [255,256,257,258,259,260,261]. RCT: CoQ10 ineffective as an ergogenic aid in both young and older trained men [262]. |
| 10. Immune Function | Vit C, Vit A, Zinc, Selenium | Regulate redox-sensitive pathways, support leukocyte and NK cell function | Infections, immunosenescence | Vit C enhanced neutrophil activity in stimulated neutrophils and H2O2-treated HL60 cells [263]. | Meta-analysis: Vit C reduced ICU mortality rate in sepsis and ARDS [264,265]. | Meta-analyses: No significant benefit on immune function from Vit C in patients with sepsis, viral infections, and common colds [266]. |
| Aging & Frailty | CoQ10, Vit E, Polyphenols | Reduce mitochondrial ROS, preserve telomeres and aging processes | Cognitive decline, frailty, sarcopenia | CoQ10, curcumin, and revesterol extended lifespan and improved cognitive and motor function in aged rodents [267,268,269,270]. | RCTs: CoQ10 and selenium reduced serum R-SH levels, reduced cardiac mortality, and improved CV health in elderly [271,272,273,274]. | Lifespan extension not seen in human RCTs [207]. Meta-analysis: Minor improvement in physical robustness-related outcomes, but frailty endpoints still exploratory [252]. |
| Inflammatory Diseases | Curcumin, ω-3 FAs, Quercetin, Resveratrol | Inhibit NF-κB, COX-2; ↓ IL-6, TNF-α | RA, IBD, asthma, lupus | Curcumin reduced joint inflammation in mice [275]. Curcumin suppressed TNF-α and IL-6 in DSS-induced colitis and CIA models [276,277]. | RCT shows that curcumin decreased serum pro-inflammatory cytokines in subjects with metabolic syndrome [278]. RCT: Meriva® curcumin reduced CRP and DAS28 in RA over 24 weeks [279]. | Meta-analysis: Benefit in RA from curcumin but limited in IBD due to low absorption, poor bioavailability, and heterogeneity in RCT endpoints [280]. |
| Renal Disorders | Curcumin, NAC, Vit E, Selenium | Inhibit oxidative glomerular injury | CKD, diabetic mephropathy | Curcumin exhibited renoprotective effects by reducing fibrosis in nephrectomized rats [281,282] | Meta-analysis: Curcumin reduced development of ESKD [283]. RCTs: NAC reduced CKD progression and ESRD in CKD patients [284,285,286]. | Meta-analysis: No reduced death or improved kidney transplant outcomes or proteinuria in patients with CKD [283,287]. Curcumin dose adjustment essential in renal impairment. Risk of nephrotoxicity with overdose; hydration status affects outcomes. |
| Respiratory Disorders | NAC, Quercetin, Vit E, Curcumin | Reduce airway ROS, inhibit cytokine-driven mucus production | COPD, asthma, pulmonary fibrosis | NAC attenuated lung damage, pulmonary emphysema, mucus, and inflammation in COPD rats [288,289,290,291]. | Meta-analysis: Inhaled NAC improved FEV1 by 7% and reduced exacerbations in moderate COPD [292]. | Meta-analysis: NAC did not reduce the risk of acute exacerbation or ameliorate the decline in lung volume in COPD patients [293]. NAC effective only in chronic use, inhaled forms superior to oral. Mixed COPD results with oral NAC; limited bioavailability [294]. |
| Gastrointestinal Health | Curcumin, Sulforaphane, Polyphenols | Preserve gut barrier, modulate microbiota, reduce redox-driven inflammation | IBD, CRC, gastritis | Curcumin maintained intestinal barrier function and effectively alleviated colitis injury in DSS colitis mice [276,295]. | RCT: Curcumin + mesalamine induced remission in patients with ulcerative colitis [296]. | RCTs are small and heterogeneous; formulation issues common [297]. |
| Skeletal Health | Vit K, Resveratrol, Vit C | Inhibit osteoclast activity, enhance osteoblast mineral deposition | Osteoporosis, inflammatory bone loss | GTP reduced bone loss and improved mineral density and bone mechanical properties in chronic inflammation-induced bone loss mice [298,299]. | Meta-analyses: Antioxidants effective for sarcopenia, especially improving muscle strength and function [300,301,302]. | RCT: Antioxidant supplementation did not affect bone turnover markers [303,304]; Human data limited; limited FDA-approved antioxidant sfor bone health. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manful, C.F.; Fordjour, E.; Subramaniam, D.; Sey, A.A.; Abbey, L.; Thomas, R. Antioxidants and Reactive Oxygen Species: Shaping Human Health and Disease Outcomes. Int. J. Mol. Sci. 2025, 26, 7520. https://doi.org/10.3390/ijms26157520
Manful CF, Fordjour E, Subramaniam D, Sey AA, Abbey L, Thomas R. Antioxidants and Reactive Oxygen Species: Shaping Human Health and Disease Outcomes. International Journal of Molecular Sciences. 2025; 26(15):7520. https://doi.org/10.3390/ijms26157520
Chicago/Turabian StyleManful, Charles F., Eric Fordjour, Dasinaa Subramaniam, Albert A. Sey, Lord Abbey, and Raymond Thomas. 2025. "Antioxidants and Reactive Oxygen Species: Shaping Human Health and Disease Outcomes" International Journal of Molecular Sciences 26, no. 15: 7520. https://doi.org/10.3390/ijms26157520
APA StyleManful, C. F., Fordjour, E., Subramaniam, D., Sey, A. A., Abbey, L., & Thomas, R. (2025). Antioxidants and Reactive Oxygen Species: Shaping Human Health and Disease Outcomes. International Journal of Molecular Sciences, 26(15), 7520. https://doi.org/10.3390/ijms26157520