

Nanoplastic Disrupts Intestinal Homeostasis in Immature Rats by Altering the Metabolite Profile and Gene Expression

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Accumulation of PS-NPs in the Jejunum of Exposed Rats
2.2. Metabolomics of the PS-NP-Induced Jejunum of Immature Rats
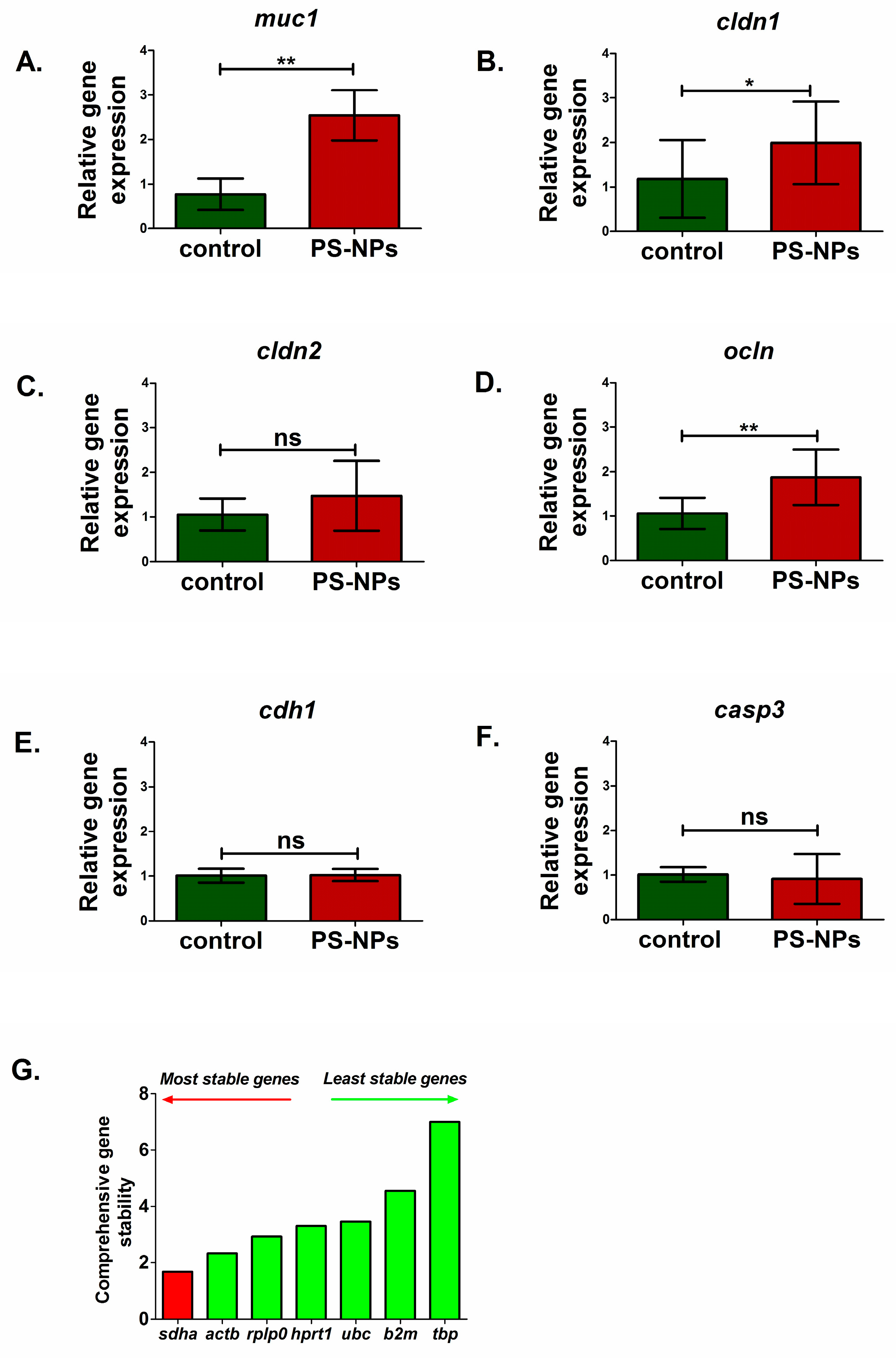
2.3. PS-NPs-Induced Alterations in the Expression of Selected Genes Linked to Jejunum Functions
2.4. Oxidative Stress and Inflammation Are the Main Pathological Processes Induced by PS-NPs in the Jejunum of Immature Rats
3. Discussion
3.1. Accumulation of PS-NPs in the Jejunum of Exposed Immature Rats and Their Effects on Metabolomics and Gene Expression
3.2. PS-NPs Induced Oxidative Stress and Inflammation in the Jejunum of Immature Rats
4. Materials and Methods
4.1. PS-NPs and Experimental Design
4.2. Sample Preparation for NMR Examinations
4.3. NMR Spectroscopy Measurements
4.4. Visualization of PS-NPs in the Jejunum by Confocal Microscopy
4.5. Transmission Electron Microscopy (TEM) Analysis
4.6. Analysis of Relative Gene Expression by qRT–PCR
4.6.1. RNA Isolation
4.6.2. RT
4.6.3. SYBR GREEN Gene Expression Assays

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | GenBank Number | Primer Sequence | Amplicon Length |
|---|---|---|---|---|
| fabp2 | Fatty acid binding protein 2 | NM_013068.1 | ATCATGGCATTTGATGGCACTTG | 150 |
| TCCTGTGTGATCGTCAGTTTCAA | ||||
| chga | Chromogranin A | NM_021655.2 | CGGAAGTATTTGAGAACCAGAGC | 113 |
| ATCCTGTTGCCCCTTGTCAG | ||||
| lgr5 | Leucine-rich repeat-containing G protein-coupled receptor 5 | NM_001106784.1 | ATACTGTCACTGTGAGCTGGATG | 132 |
| TGACTGATGTTGTTCATACTGAGGT | ||||
| lyz1 | Lysozyme | NM_012771.3 | AAGGAATGGGATGTCTGGCT | 127 |
| CCCATAGTCGGTGCTTTGGT | ||||
| muc2 | Mucin 2 | NM_022174.1 | GATGTGTGGGACCGGACAAT | 118 |
| GCACACTTCTTTGGTTGGCA | ||||
| muc1 | Mucin 1 | NM_001398538.1 | CTCGGAAGTCAATGTGAATGAGATG | |
| CAATGAGATAGACGATGACCAAAGC | 131 | |||
| s100β | S100 calcium-binding protein B | NM_013191.2 | TGTCTACCCTCCTAGTCCTCG | 94 |
| CCTTCTCCAGCTCAGACATCC | ||||
| cspg4 | Chondroitin sulfate proteoglycan 4 | NM_031022.2 | CTCCAGTTCTCCACATCGCA | 119 |
| TTTTGTCCCAGGGCAAGTCT | ||||
| map2 | Microtubule-associated protein 2 | NM_013066.1 | GCCGGGGCCATGATCTTTC | 80 |
| GTAATCATCTCCTTCATCCATCGTT | ||||
| Il1b | Interleukin 1 beta | NM_031512.2 | GCTATGGCAACTGTCCCTGA | 134 |
| TCTGGACAGCCCAAGTCAAG | ||||
| tnf | Tumor necrosis factor | NM_012675.3 | ATGGGCTCCCTCTCATCAGT | 106 |
| GCTTGGTGGTTTGCTACGAC | ||||
| Il23a | Interleukin 23 subunit alpha | NM_130410.2 | AAAAGTGACGTGCCCCGTAT | 98 |
| AGACCTTGGCGGATCCTTTG | ||||
| ocln | Occludin | NM_031329.3 | GCAGTGAACAAGCTGTGTCTAAA | 179 |
| CGGCTAAAACAGACCAAACTGG | ||||
| cldn1 | Claudin 1 | NM_031699.3 | GACTGCTCAGGCCATCTACG | 132 |
| ACCATCAAGGCTCTGGTTGC | ||||
| cldn2 | Claudin 2 | NM_001106846.2 | CGAGAAAGAACAGCTCCGTTT | 105 |
| TCACAGTGTCTCTGGCAAGC | ||||
| cdh1 | Cadherin 1 | NM_031334.1 | ATTACAAGTTCCCGCCATCCTT | 148 |
| ATACACATTGTCCCGGGTATCG | ||||
| casp3 | Caspase 3 | NM_012922.2 | GGAGCTTGGAACGCGAAGAAAAG | 135 |
| CTGCTGTCCAGATATATTCCAGAGT | ||||
| sdha | Succinate dehydrogenase complex flavoprotein subunit A | NM_130428.1 | GTATTTCGCACTGGATCTTCTGATG | 97 |
| CTTTGCTCTTATTCGGTGTATGGAC | ||||
| actb | Actin, beta | NM_031144.3 | TACAACCTTCTTGCAGCTCCTC | 200 |
| TGACCCATACCCACCATCACAC | ||||
| rplp0 | Ribosomal protein lateral stalk subunit P0 | NM_022402.2 | CATCAATGGATACAAAAGGGTCCTG | 271 |
| TCTTTCTCAAATTAAGCAGGCTGAC | ||||
| hprt1 | Hypoxanthine phosphoribosyltransferase 1 | NM_012583.2 | CTTCCTCCTCAGACCGCTTTTC | 169 |
| ATCAGTCCATGAGGAATAAACACCT | ||||
| ubc | Ubiquitin C | NM_001399781.1 | AAGATACTCGTACCTTTCTCACCAC | 88 |
| AAAACTAAGACACCTCCCCATCAAA | ||||
| b2m | beta-2 microglobulin | NM_012512.2 | ATGTTAGGATGAAAGCCCAGGTATT | 141 |
| CCAACAGAATTTACCAGGAAACACA | ||||
| tbp | TATA box binding protein | NM_001004198.1 | CACCGTACATCTCAGCTGCTTC | 134 |
| ATCTCCTTAGAAACGTCTTCGACTT | ||||
| TaqMan probes | ||||
| Gene symbol | Gene name | Chromosome Location | Assay ID | Manufacturer |
| sod1 | Superoxide dismutase 1 | Chr.11: 30363282–30368858 on Build Rnor_6.0 | Rn00566938_m1 | Thermo Fisher Scientific (Warsaw, Poland) |
| sod2 | Superoxide dismutase 2 | Chr.1: 47914757–47921587 on Build Rnor_6.0 | Rn00690588_g1 | Thermo Fisher Scientific (Warsaw, Poland) |
| cat | Catalase | Chr.3: 93379872–93412058 on Build Rnor_6.0 | Rn01512560_m1 | Thermo Fisher Scientific (Warsaw, Poland) |
| gpx1 | Glutathione peroxidase 1 | Chr.8: 117117430–117118528 on Build Rnor_6.0 | Rn00577994_g1 | Thermo Fisher Scientific (Warsaw, Poland) |
| gpx4 | Glutathione peroxidase 4 | Chr.7: 12516357–12519154 on Build Rnor_6.0 | Rn00820818_g1 | Thermo Fisher Scientific (Warsaw, Poland) |
4.6.4. TaqMan Gene Expression Assays
4.7. Western Blot Analysis
4.8. Measurements of Oxidative Stress Markers
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, C.; Qiu, M.; Wang, S.; Luo, J.; Huang, L.; Deng, Q.; Fang, Z.; Sun, L.; Gooneratne, R. Gut-Microbiota-Derived Butyric Acid Overload Contributes to Ileal Mucosal Barrier Damage in Late Phase of Chronic Unpredictable Mild Stress Mice. Int. J. Mol. Sci. 2024, 25, 12998. [Google Scholar] [CrossRef]
- Wang, L.; Wu, W.M.; Bolan, N.S.; Tsang, D.C.W.; Li, Y.; Qin, M.; Hou, D. Environmental fate, toxicity and risk management strategies of nanoplastics in the environment: Current status and future perspectives. J. Hazard. Mater. 2021, 401, 123415. [Google Scholar] [CrossRef]
- Yan, Z.; Liu, Y.; Zhang, T.; Zhang, F.; Ren, H.; Zhang, Y. Analysis of Microplastics in Human Feces Reveals a Correlation between Fecal Microplastics and Inflammatory Bowel Disease Status. Environ. Sci. Technol. 2022, 56, 414–421. [Google Scholar] [CrossRef]
- Cetin, M.; Miloglu, F.D.; Baygutalp, N.K.; Ceylan, O.; Yildirim, S.; Eser, G.; Gul, H.I. Higher number of microplastics in tumoral colon tissues from patients with colorectal adenocarcinoma. Environ. Chem. Lett. 2023, 21, 639–646. [Google Scholar] [CrossRef]
- Bruno, A.; Dovizio, M.; Milillo, C.; Aruffo, E.; Pesce, M.; Gatta, M.; Chiacchiaretta, P.; Di Carlo, P.; Ballerini, P. Orally Ingested Micro- and Nano-Plastics: A Hidden Driver of Inflammatory Bowel Disease and Colorectal Cancer. Cancers 2024, 16, 3079. [Google Scholar] [CrossRef] [PubMed]
- Greven, A.C.; Merk, T.; Karagoz, F.; Mohr, K.; Klapper, M.; Jovanovic, B.; Palic, D. Polycarbonate and polystyrene nanoplastic particles act as stressors to the innate immune system of fathead minnow (Pimephales promelas). Environ. Toxicol. Chem. 2016, 35, 3093–3100. [Google Scholar] [CrossRef] [PubMed]
- Covello, C.; Di Vincenzo, F.; Cammarota, G.; Pizzoferrato, M. Micro(nano)plastics and Their Potential Impact on Human Gut Health: A Narrative Review. Curr. Issues Mol. Biol. 2024, 46, 2658–2677. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, M.; Wang, L.; Gu, W.; Li, X.; Han, Z.; Fu, X.; Wang, X.; Li, X.; Su, Z. Continuous oral exposure to micro- and nanoplastics induced gut microbiota dysbiosis, intestinal barrier and immune dysfunction in adult mice. Environ. Int. 2023, 182, 108353. [Google Scholar] [CrossRef]
- Afzaal, M.; Saeed, F.; Shah, Y.A.; Hussain, M.; Rabail, R.; Socol, C.T.; Hassoun, A.; Pateiro, M.; Lorenzo, J.M.; Rusu, A.V.; et al. Human gut microbiota in health and disease: Unveiling the relationship. Front. Microbiol. 2022, 13, 999001. [Google Scholar] [CrossRef]
- Shi, L.; Feng, Y.; Wang, J.; Xiao, R.; Wang, L.; Tian, P.; Jin, X.; Zhao, J.; Wang, G. Innovative mechanisms of micro- and nanoplastic-induced brain injury: Emphasis on the microbiota-gut-brain axis. Life Sci. 2024, 357, 123107. [Google Scholar] [CrossRef]
- Senathirajah, K.; Attwood, S.; Bhagwat, G.; Carbery, M.; Wilson, S.; Palanisami, T. Estimation of the mass of microplastics ingested—A pivotal first step towards human health risk assessment. J. Hazard. Mater. 2021, 404 Pt B, 124004. [Google Scholar] [CrossRef]
- Priya, A.K.; Jalil, A.A.; Dutta, K.; Rajendran, S.; Vasseghian, Y.; Qin, J.; Soto-Moscoso, M. Microplastics in the environment: Recent developments in characteristic, occurrence, identification and ecological risk. Chemosphere 2022, 298, 134161. [Google Scholar] [CrossRef]
- Struzynska, L.; Dabrowska-Bouta, B.; Sulkowski, G. Developmental neurotoxicity of silver nanoparticles: The current state of knowledge and future directions. Nanotoxicology 2022, 16, 500–525. [Google Scholar] [CrossRef] [PubMed]
- Adamiak, K.; Sidoryk-Wegrzynowicz, M.; Dabrowska-Bouta, B.; Sulkowski, G.; Struzynska, L. Plastic nanoparticles interfere with extracellular vesicle pathway in primary astrocytes. Ecotoxicol. Environ. Saf. 2024, 286, 117180. [Google Scholar] [CrossRef] [PubMed]
- Adamiak, K.; Sidoryk-Wegrzynowicz, M.; Dabrowska-Bouta, B.; Sulkowski, G.; Struzynska, L. Primary astrocytes as a cellular depot of polystyrene nanoparticles. Sci. Rep. 2025, 15, 6502. [Google Scholar] [CrossRef] [PubMed]
- Massey, W.J.; Kay, K.E.; Jaramillo, T.C.; Horak, A.J., 3rd; Cao, S.; Osborn, L.J.; Banerjee, R.; Mrdjen, M.; Hamoudi, M.K.; Silver, D.J.; et al. Metaorganismal choline metabolism shapes olfactory perception. J. Biol. Chem. 2023, 299, 105299. [Google Scholar] [CrossRef]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; Ian McKenzie, C.; Hijikata, A.; Wong, C.; et al. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef]
- Sun, P.Q.; Dong, W.M.; Yuan, Y.F.; Cao, Q.; Chen, X.Y.; Guo, L.L.; Jiang, Y.Y. Targeted metabolomics study of fatty-acid metabolism in lean metabolic-associated fatty liver disease patients. World J. Gastroenterol. 2024, 30, 3290–3303. [Google Scholar] [CrossRef]
- Visser, A.; van Zwol, W.; Kloosterhuis, N.; Huijkman, N.; Smit, M.; Koster, M.; Bloks, V.; Hussain, M.M.; van de Sluis, B.; Kuivenhoven, J.A. ERICH4 is not involved in the assembly and secretion of intestinal lipoproteins. Atherosclerosis 2024, 399, 118635. [Google Scholar] [CrossRef]
- Atanga, R.; Singh, V.; In, J.G. Intestinal Enteroendocrine Cells: Present and Future Druggable Targets. Int. J. Mol. Sci. 2023, 24, 8836. [Google Scholar] [CrossRef]
- Syed, Z.A.; Zhang, L.; Ten Hagen, K.G. In vivo models of mucin biosynthesis and function. Adv. Drug Deliv. Rev. 2022, 184, 114182. [Google Scholar] [CrossRef]
- Hansson, G.C. Mucins and the Microbiome. Annu. Rev. Biochem. 2020, 89, 769–793. [Google Scholar] [CrossRef]
- Croze, M.L.; Soulage, C.O. Potential role and therapeutic interests of myo-inositol in metabolic diseases. Biochimie 2013, 95, 1811–1827. [Google Scholar] [CrossRef]
- Lolicato, F.; Nickel, W.; Haucke, V.; Ebner, M. Phosphoinositide switches in cell physiology—From molecular mechanisms to disease. J. Biol. Chem. 2024, 300, 105757. [Google Scholar] [CrossRef]
- Kaiko, G.E.; Ryu, S.H.; Koues, O.I.; Collins, P.L.; Solnica-Krezel, L.; Pearce, E.J.; Pearce, E.L.; Oltz, E.M.; Stappenbeck, T.S. The Colonic Crypt Protects Stem Cells from Microbiota-Derived Metabolites. Cell 2016, 165, 1708–1720. [Google Scholar] [CrossRef]
- Progatzky, F.; Pachnis, V. The role of enteric glia in intestinal immunity. Curr. Opin. Immunol. 2022, 77, 102183. [Google Scholar] [CrossRef] [PubMed]
- Won, S.Y.; Han, G.P.; Kwon, C.H.; Lee, E.C.; Kil, D.Y. Effect of individual or combination of dietary betaine and glycine on productive performance, stress response, liver health, and intestinal barrier function in broiler chickens raised under heat stress conditions. Poult. Sci. 2023, 102, 102771. [Google Scholar] [CrossRef]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Yimamu, M.; Zhang, S.; Shah, A.M.; Yang, H.; Cai, W.; Li, C.; Lu, X.; Li, F.; Yang, K. Effects of guanidino acetic acid and betaine supplementation on growth, dietary nutrient digestion and intestinal creatine metabolism in sheep. Vet. Med. Sci. 2024, 10, e1470. [Google Scholar] [CrossRef] [PubMed]
- Schermuly, I.I.; Romanet, S.; Patra, A.K.; Mastrototaro, L.; Lemme, A.; Pieper, R.; Zentek, J.; Aschenbach, J.R. Transport of Neutral Amino Acids in the Jejunum of Pigs with Special Consideration of L-Methionine. Nutrients 2024, 16, 3418. [Google Scholar] [CrossRef]
- Yi, Z.; Tan, X.; Wang, Q.; Huang, P.; Li, Y.; Ding, X.; Li, J.; Huang, J.; Yang, H.; Yin, Y. Dietary niacin affects intestinal morphology and functions via modulating cell proliferation in weaned piglets. Food Funct. 2021, 12, 7402–7414. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.H.; Hao, Z.; Qiu, M.; Liu, Y.; Chang, M.; Liu, X.; Wang, Y.; Tang, Y.; Sun, W.; Teng, X.; et al. Amino acid metabolism disorder and oxidative stress took part in EGCG alleviating Mn-caused ferroptosis via miR-9–5p/got1 axis. J. Hazard. Mater. 2025, 489, 137656. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, M.; Vianello, A.; Picker, M.; Simon-Sanchez, L.; Chen, R.; Estevinho, M.M.; Weinstein, K.; Lykkemark, J.; Jess, T.; Peter, I.; et al. Micro- and nano-plastics, intestinal inflammation, and inflammatory bowel disease: A review of the literature. Sci. Total Environ. 2024, 953, 176228. [Google Scholar] [CrossRef] [PubMed]
- Lueschow, S.R.; McElroy, S.J. The Paneth Cell: The Curator and Defender of the Immature Small Intestine. Front. Immunol. 2020, 11, 587. [Google Scholar] [CrossRef]
- Yin, L.; Huang, L.; Kufe, D. MUC1 oncoprotein activates the FOXO3a transcription factor in a survival response to oxidative stress. J. Biol. Chem. 2004, 279, 45721–45727. [Google Scholar] [CrossRef]
- Alaa, M.; Abdel Razek, A.H.; Tony, M.A.; Yassin, A.M.; Warda, M.; Awad, M.A.; Bawish, B.M. Guanidinoacetic acid supplementation and stocking density effects on broiler performance: Behavior, biochemistry, immunity, and small intestinal histomorphology. Acta Vet. Scand. 2024, 66, 62. [Google Scholar] [CrossRef]
- Han, M.; Gao, T.; Liu, G.; Zhu, C.; Zhang, T.; Sun, M.; Li, J.; Ji, F.; Si, Q.; Jiang, Q. The effect of a polystyrene nanoplastic on the intestinal microbes and oxidative stress defense of the freshwater crayfish, Procambarus clarkii. Sci. Total Environ. 2022, 833, 155722. [Google Scholar] [CrossRef]
- Qiu, M.H.; Hao, Z.; Liu, Y.; Liu, Y.; Chang, M.; Lin, X.; Liu, X.; Dong, N.; Sun, W.; Teng, X. ROS acted as an initial role in selenium nanoparticles alleviating insecticide chlorpyrifos-induced oxidative stress, pyroptosis, and intestinal barrier dysfunction in porcine intestinal epithelial cells. Pestic. Biochem. Physiol. 2025, 211, 106418. [Google Scholar] [CrossRef]
- Federico, A.; Cardaioli, E.; Da Pozzo, P.; Formichi, P.; Gallus, G.N.; Radi, E. Mitochondria, oxidative stress and neurodegeneration. J. Neurol. Sci. 2012, 322, 254–262. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, L.; Ding, B.; Liu, Y.; Zhu, H.; Liu, J.; Li, Y.; Kang, P.; Yin, Y.; Wu, G. Alpha-Ketoglutarate and intestinal function. Front. Biosci. 2011, 16, 1186–1196. [Google Scholar] [CrossRef]
- Govindaraju, V.; Young, K.; Maudsley, A.A. Proton NMR chemical shifts and coupling constants for brain metabolites. NMR Biomed. 2000, 13, 129–153. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–428. [Google Scholar] [CrossRef]
- Augustyniak, J.; Lenart, J.; Lipka, G.; Stepien, P.P.; Buzanska, L. Reference Gene Validation via RT-qPCR for Human iPSC-Derived Neural Stem Cells and Neural Progenitors. Mol. Neurobiol. 2019, 56, 6820–6832. [Google Scholar] [CrossRef]
- Zieminska, E.; Toczylowska, B.; Diamandakis, D.; Hilgier, W.; Filipkowski, R.K.; Polowy, R.; Orzel, J.; Gorka, M.; Lazarewicz, J.W. Glutamate, Glutamine and GABA Levels in Rat Brain Measured Using MRS, HPLC and NMR Methods in Study of Two Models of Autism. Front. Mol. Neurosci. 2018, 11, 418. [Google Scholar] [CrossRef]
- Weigl, W.; Milej, D.; Gerega, A.; Toczylowska, B.; Sawosz, P.; Kacprzak, M.; Janusek, D.; Wojtkiewicz, S.; Maniewski, R.; Liebert, A. Confirmation of brain death using optical methods based on tracking of an optical contrast agent: Assessment of diagnostic feasibility. Sci. Rep. 2018, 8, 7332. [Google Scholar] [CrossRef] [PubMed]
- Ellis, D.I.; Dunn, W.B.; Griffin, J.L.; Allwood, J.W.; Goodacre, R. Metabolic fingerprinting as a diagnostic tool. Pharmacogenomics 2007, 8, 1243–1266. [Google Scholar] [CrossRef] [PubMed]

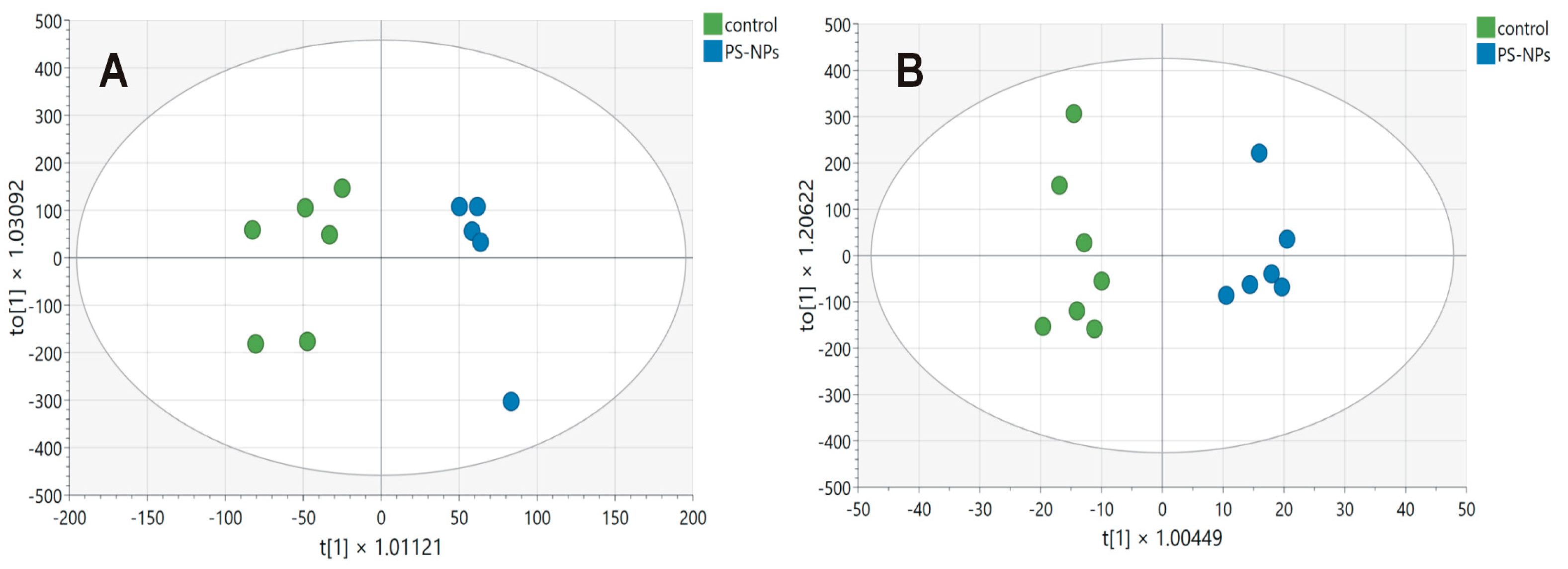




| Hydrophilic Metabolite | PS-NPs vs. Controls [%] | p | VIP |
|---|---|---|---|
| NAD | 61 | 0.248 | |
| Nicotinic acid | 55 | 0.017 | |
| Purine | 75 | 0.303 | |
| IMP/Inosine/Adenosine | 137 | 0.126 | |
| ITP/AMP/IMP/Nicotinic acid | 79 | 0.565 | |
| Hypoxanthine | 100 | 0.662 | |
| Quinolinic acid/Adenine xanthine/Purine | 59 | 0.247 | |
| Uridine | 101 | 0.931 | |
| Guanine | 50 | 0.537 | |
| Phenylalanine | 208 | 0.177 | |
| Cytidine/Cytosine | 182 | 0.126 | |
| Histamine/Serotonin | 123 | 0.387 | |
| Tyrosine | 174 | 0.223 | |
| Fumarate | 66 | 0.322 | |
| AMP/IMP | 119 | 0.547 | |
| Uracil | 91 | 0.429 | |
| α-D glucose | 213 | 0.429 | |
| β-D-glucose | 207 | 0.110 | |
| Guanidinoacetic acid | 183 | 0.229 | 2.02 |
| Myo-inositol | 73 | 0.177 | 1.12 |
| Glycine | 156 | 0.247 | 1.20 |
| Taurine | 80 | 0.380 | |
| Trimethylamine oxide (TMAO)/Betaine | 80 | 0.534 | 2.11 |
| Taurine/Myo-inositol | 95 | 0.829 | |
| Phosphorylcholine | 51 | 0.128 | 2.09 |
| Choline | 89 | 0.329 | 2.52 |
| Ethanolamine/Arginine/Cysteine/Ornithine | 95 | 0.247 | |
| Creatinine | 71 | 0.340 | |
| Creatine | 69 | 0.292 | 1.24 |
| α-ketoisovaleric acid/ α-ketoglutaric acid | 190 | 0.126 | 1.42 |
| Aspartic acid | 49 | 0.013 | |
| Glutamine GLN | 177 | 0.217 | |
| Glutamic acid GLU | 142 | 0.429 | |
| Proline | 159 | 0.082 | |
| α-ketoglutaric acid | 124 | 0.530 | |
| Methionine | 145 | 0.389 | 1.17 |
| Arginine | 164 | 0.265 | |
| Lysine | 175 | 0.126 | 1.20 |
| Alanine | 180 | 0.126 | 2.39 |
| Lactate | 100 | 0.989 | |
| Threonine | 147 | 0.429 | 1.07 |
| 3-hydroxyisovalerate/ Methylmalonate/Lipid | 76 | 0.429 | |
| α-ketoisovaleric aid | 166 | 0.329 | |
| Valine | 157 | 0.302 | 1.35 |
| Isoleucine/Leucine | 192 | 0.247 |
| Hydrophobic Metabolite/Functional Group | PS-NPs vs. Controls [%] | p | VIP |
|---|---|---|---|
| 16-hydroxyestradiol | 145 | 0.324 | |
| Estriol | 107 | 0.945 | |
| Testosterone | 201 | 0.269 | |
| Phosphatidylcholine (PC)/Phosphatidylethanolamine (PE)/Sphingomyelin (SM) | 103 | 0.928 | |
| 1,2-Diacyloglycerol (DAG)/2-Monoacyloglycerol (MAG) | 98 | 0.445 | |
| Phosphatidylcholine/Phosphatidylethanolamine | 81 | 0.445 | |
| Triglycerides (TG) | 45 | 0.132 | 1.23 |
| 1,3-DAG | 120 | 0.534 | |
| SM | 95 | 1.0 | |
| PC | 112 | 0.712 | |
| 1-MAG or 1,2-DAG | 108 | 0.826 | |
| PC/SM | 78 | 0.534 | |
| PUFA (arachidonic, alfa-linolenic, DHA) | 88 | 0.330 | |
| Linoleic acid | 97 | 0.897 | |
| Saturated fatty acid (FA), polyunsaturated FA (PUFA) and monounsaturated FA (MUFA) (lauric, myristic/dodecanoic/palmitic, arachidonic, alfa-linolenic/oleic) | 92 | 0.769 | |
| Hexanoylglycine | 117 | 0.836 | |
| Oleic acid | 76 | 0.394 | 1.65 |
| Vaccenic acid | 102 | 0.836 | |
| Palmitoleic acid | 90 | 0.295 | |
| Free cholesterol and 1,3-DAG, 1-MAG, and FA (dodecanoic acid) | 100 | 0.628 | |
| Saturated FA and PUFA (lauric/palmitic) | 106 | 0.808 | 3.41 |
| Cholestenol | 114 | 0.534 | |
| Saturated FA, PUFA, and MUFA (dodecanonic, palmitic/arachidonic/palmitoleic, oleic, | 107 | 0.445 | 1.72 |
| Pelargonic acid (nonanoic acid) | 99 | 0.628 | |
| 24S- hydroxycholesterol | 129 | 0.366 | |
| Free cholesterol | 120 | 0.376 | 1.40 |
| Compound/ Functional Group | Direction of Changes vs. Control | Metabolic Pathway | Probable Mechanism/Process | Marker Expression * | Direction of Changes vs. Control |
|---|---|---|---|---|---|
| Nicotinic acid | ↓ | Nicotinate and nicotinamide metabolism | -Intestinal inflammation -Barrier function | IL1β/Il1β TNFα/tnfα lgr5 muc1 cldn1 ocln | ↑/↑ ↑/↑ ↑ ↑ ↑ ↑ |
| Guanidinoacetic acid Threonine Glycine Methionine Creatine TMAO/Betaine | ↑ ↑ ↑ ↑ ↓ ↓ | Glycine, serine, threonine and cysteine, methionine metabolisms | -Intestinal barrier damage -Cellular energy disturbance -Oxidative stress -Effect on mucosal cells -Disturbance of the intestinal protein synthesis | ||
| Myo-inositol | ↓ | Inositol phosphate metabolism | -Nerve conduction impairment -Axonal degeneration, demyelination | chga CAT/cat SOD1/sod1 SOD2/sod2 s100b cspg4 fabp2 | ↓ ↓/- ↓/↓ ↑/- ↓ ↓ ↓ |
| Phosphoryl choline choline | ↓ ↓ | Glicero phospholipids metabolism | Oxidative stress | ||
| Ketoisovaleric acid/ ketoglutaric acid | ↑ | TCA cycle | -Cellular energy disturbance -Oxidative stress -Effect on enterocytes -Absorption disorders | ||
| Aspartic acid Alanine | ↓ ↑ | Alanine, aspartate and glutamate metabolism | -Intestinal barrier damage -Absorption disorders -Impact on intestinal microbiota | cldn1 ocln lgr5 muc1 lyz1 fabp2 | ↑ ↑ ↑ ↑ ↑ ↓ |
| Lysine Valine | ↑ ↑ | Valine, leucine, isoleucine biosynthesis/ degradation | -Impact on intestinal microbiota -Intestinal barrier damage -Absorption disorders | ||
| Triglycerides Free cholesterol | ↓ ↑ | Glicerolipids and cholesterol metabolisms | Effect on enterocytes | ||
| Oleic acid Lauric/palmitic acid Dodecanoic/ palmitic/arachidonic/ oleic acid | ↓ ↑ ↑ | Fatty acids biosynthesis/ metabolism | Impact on enterohepatic circulation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Augustyniak, J.; Toczylowska, B.; Dąbrowska-Bouta, B.; Adamiak, K.; Sulkowski, G.; Zieminska, E.; Struzynska, L. Nanoplastic Disrupts Intestinal Homeostasis in Immature Rats by Altering the Metabolite Profile and Gene Expression. Int. J. Mol. Sci. 2025, 26, 7207. https://doi.org/10.3390/ijms26157207
Augustyniak J, Toczylowska B, Dąbrowska-Bouta B, Adamiak K, Sulkowski G, Zieminska E, Struzynska L. Nanoplastic Disrupts Intestinal Homeostasis in Immature Rats by Altering the Metabolite Profile and Gene Expression. International Journal of Molecular Sciences. 2025; 26(15):7207. https://doi.org/10.3390/ijms26157207
Chicago/Turabian StyleAugustyniak, Justyna, Beata Toczylowska, Beata Dąbrowska-Bouta, Kamil Adamiak, Grzegorz Sulkowski, Elzbieta Zieminska, and Lidia Struzynska. 2025. "Nanoplastic Disrupts Intestinal Homeostasis in Immature Rats by Altering the Metabolite Profile and Gene Expression" International Journal of Molecular Sciences 26, no. 15: 7207. https://doi.org/10.3390/ijms26157207
APA StyleAugustyniak, J., Toczylowska, B., Dąbrowska-Bouta, B., Adamiak, K., Sulkowski, G., Zieminska, E., & Struzynska, L. (2025). Nanoplastic Disrupts Intestinal Homeostasis in Immature Rats by Altering the Metabolite Profile and Gene Expression. International Journal of Molecular Sciences, 26(15), 7207. https://doi.org/10.3390/ijms26157207