
Effects of Polypropylene and Polyethylene Terephthalate Microplastics on Trypsin Structure and Function

 ,
,  , , , ,
, , , ,
Abstract
1. Introduction
2. Results
2.1. Binding Analysis of Trypsin to MPs
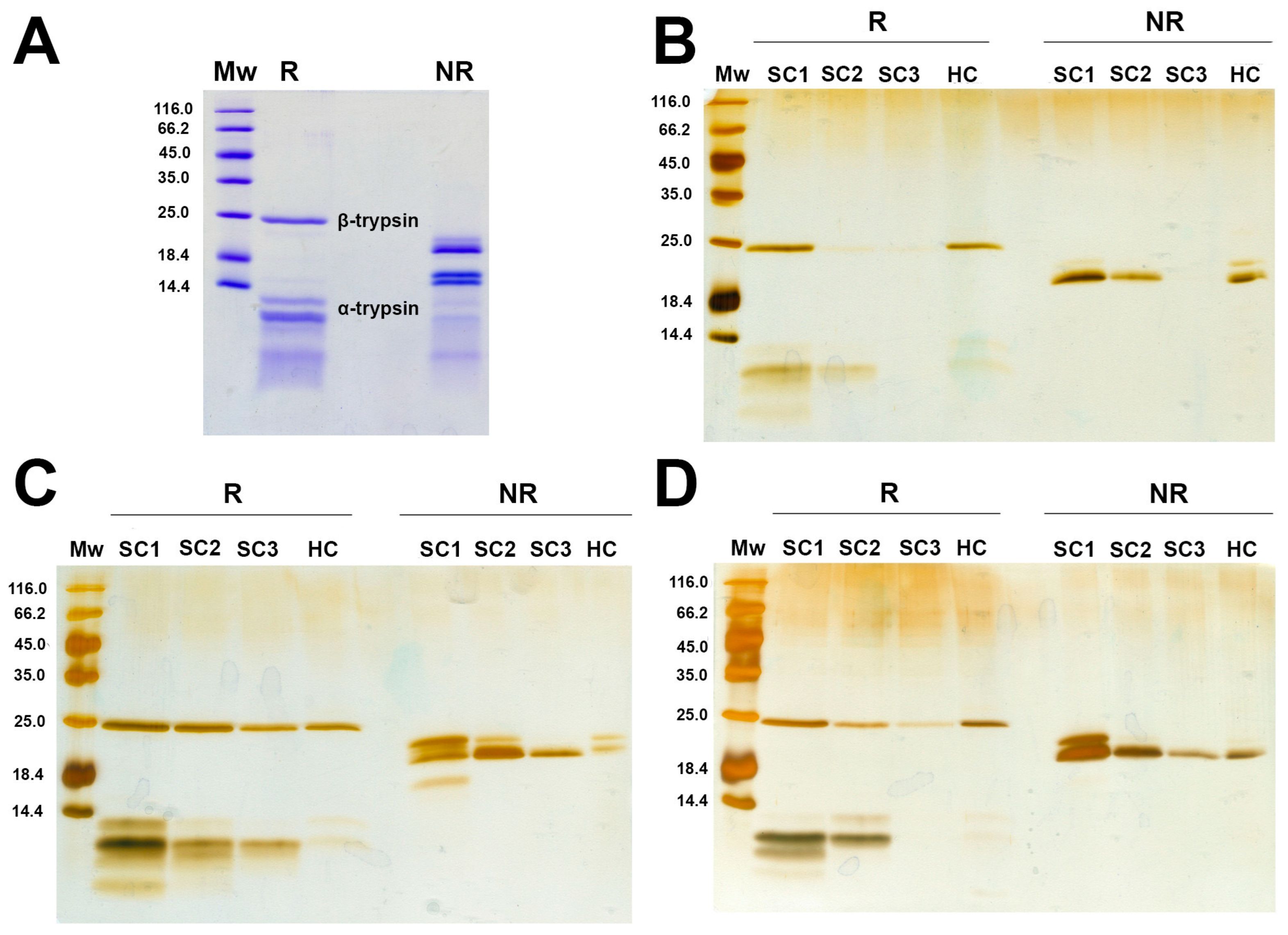
2.2. Soft and Hard Corona Formation by Trypsin onto the Surface of MPs
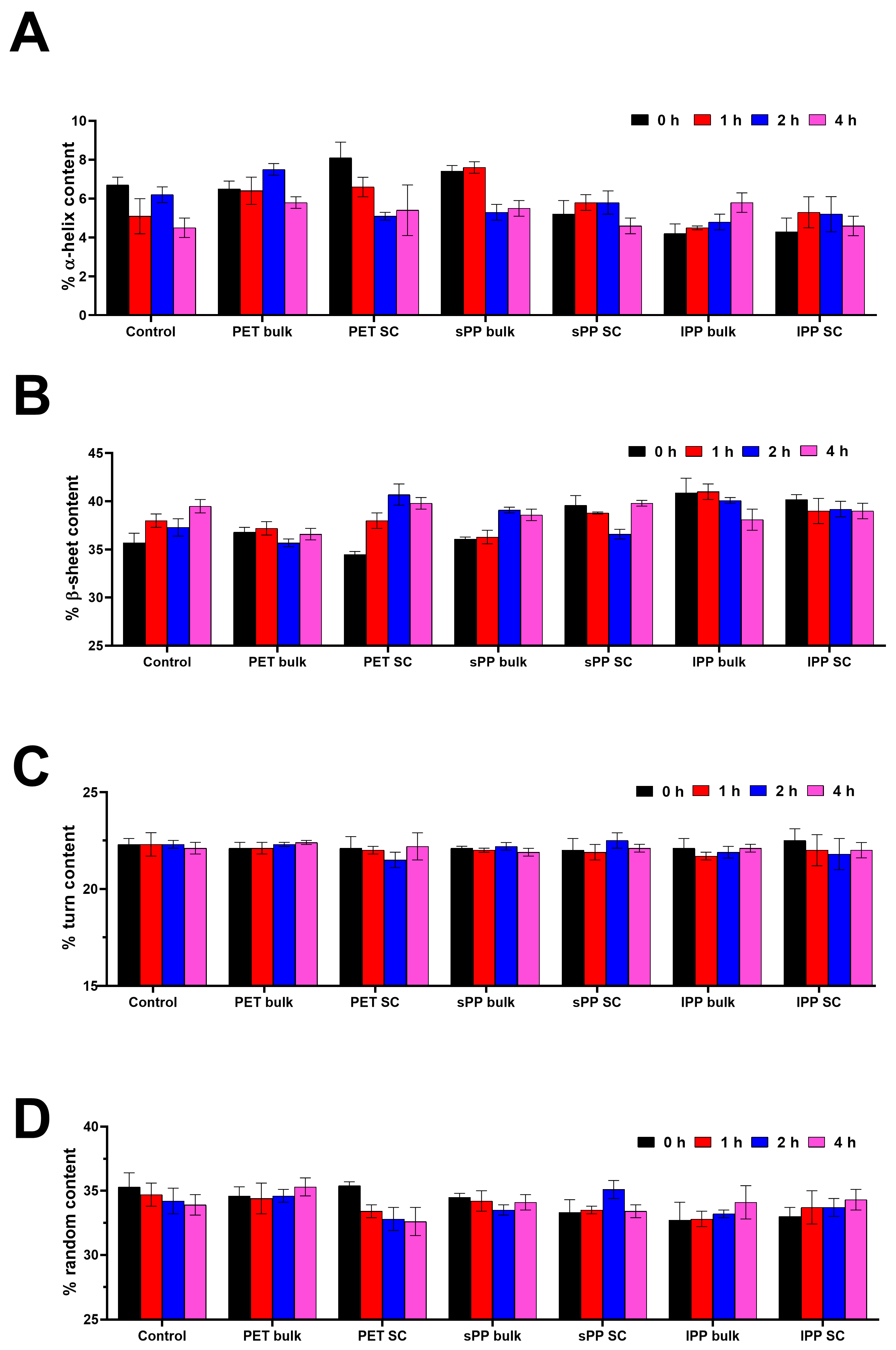
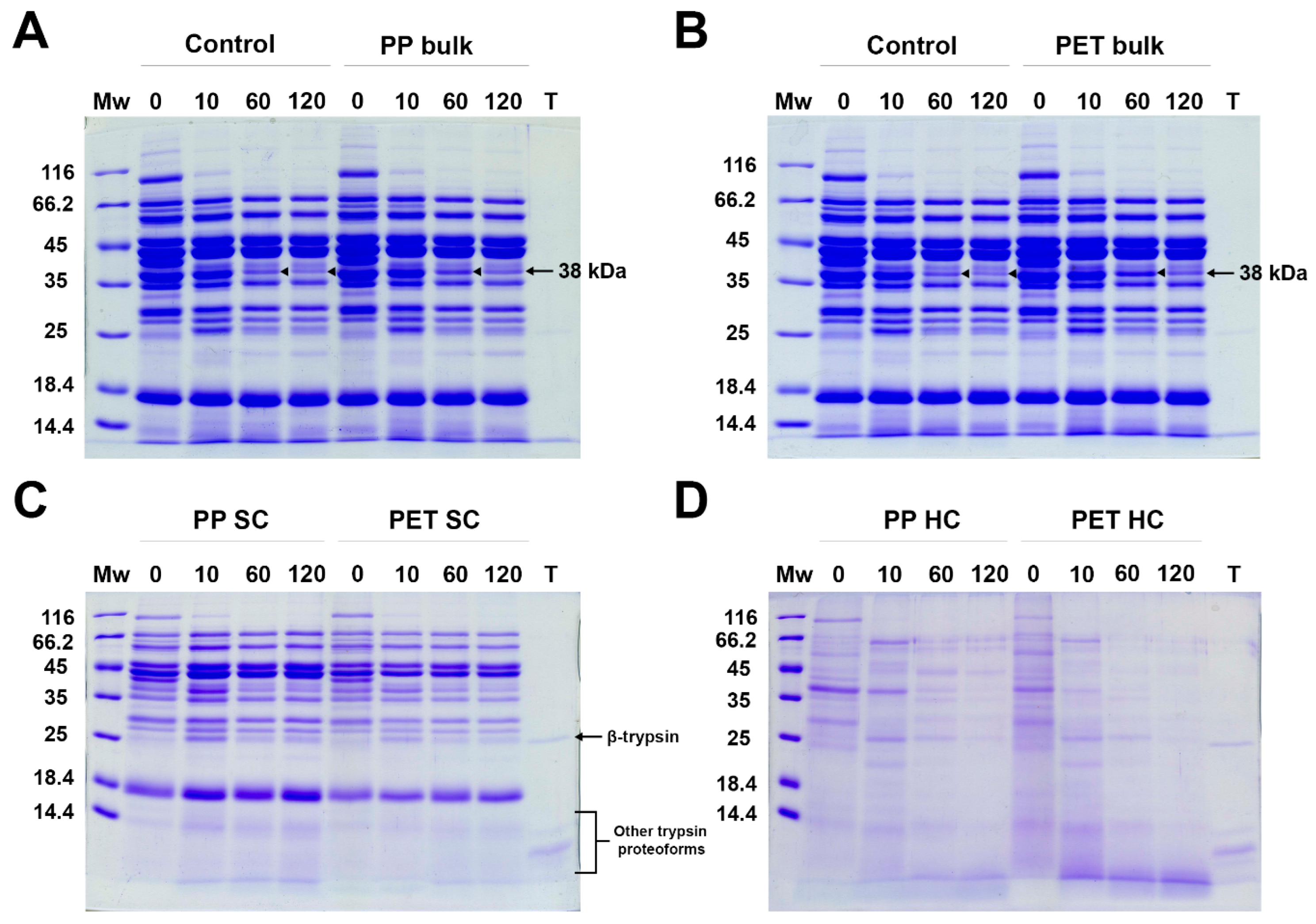
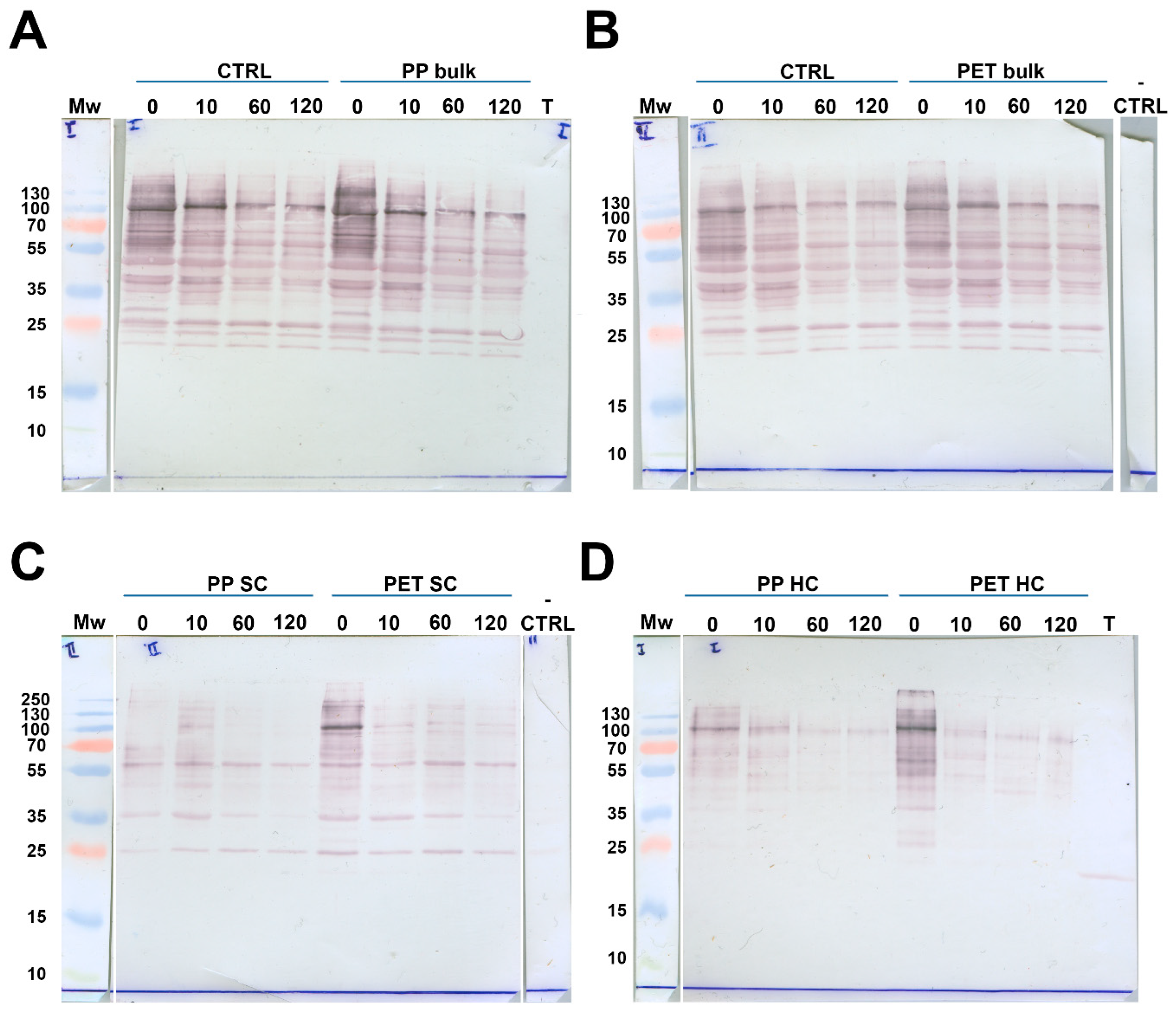
2.3. Structural Analysis of Trypsin After Incubation with MPs
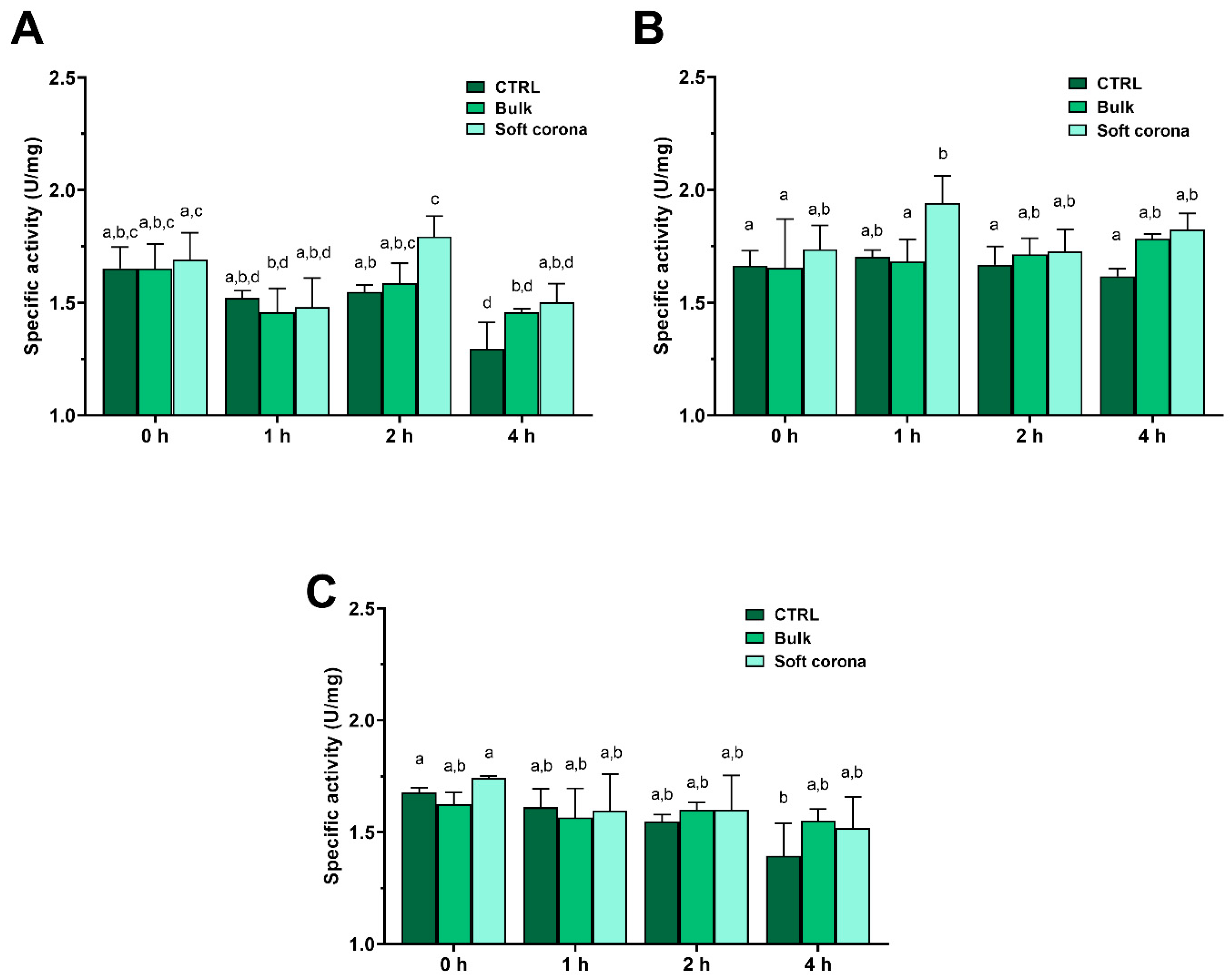
2.4. Influence of MPs on the Activity of Trypsin
2.4.1. Influence of MPs on the Activity of Bulk and Soft Corona Trypsin
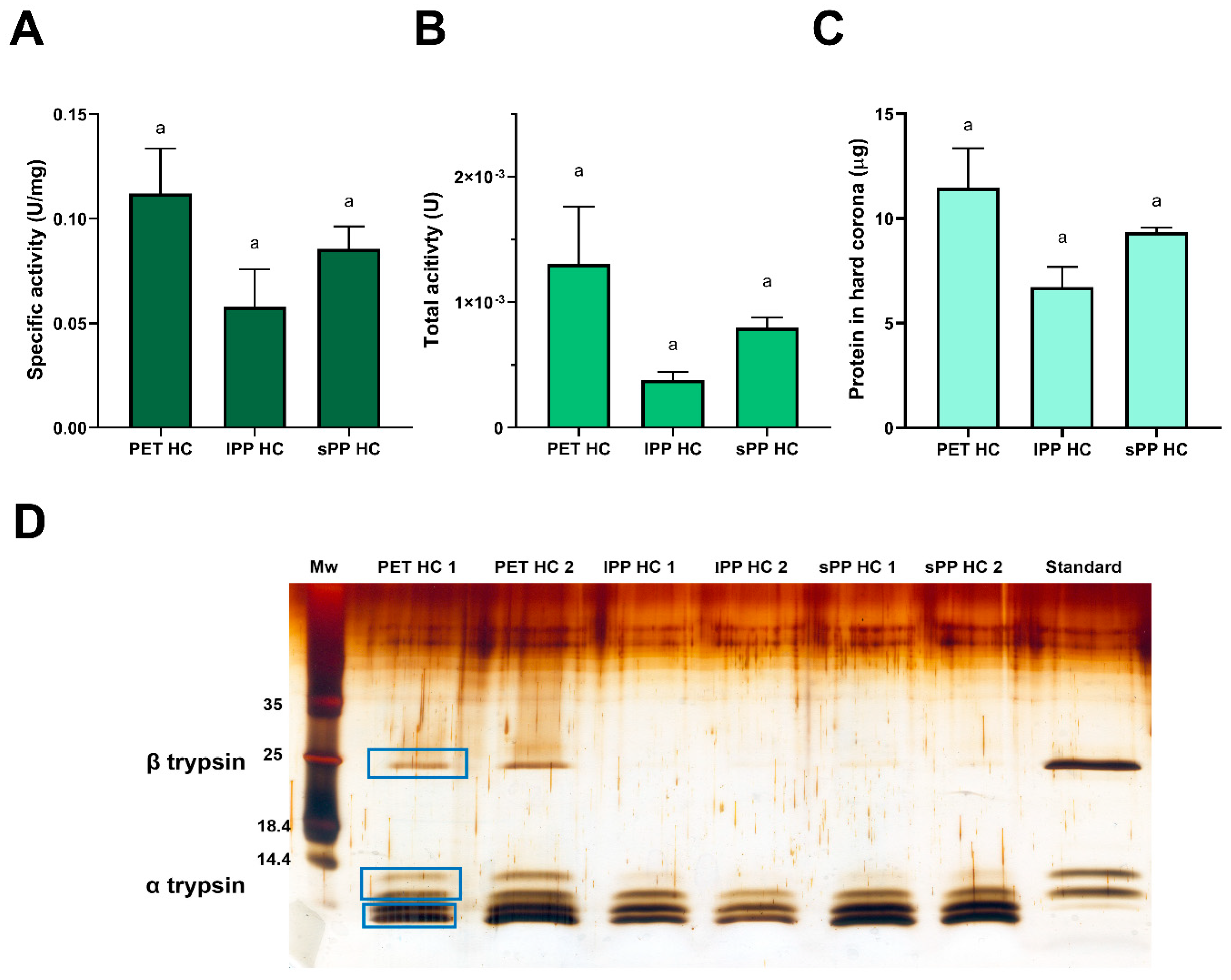
2.4.2. Activity of Trypsin in the Hard Corona
2.5. Influence of MPs on In Vitro Digestion of Meat Extract Proteins by Trypsin
2.6. Influence of MPs on In Vitro Digestion of α-Gal Bearing Meat Allergens from Meat Extract by Trypsin
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Trypsin Binding to MPs
4.3. Formation of Soft and Hard Corona on the Surface of Tested MPs
4.4. Influence of MPs on the Structure of Trypsin Analyzed by Circular Dichroism (CD) Spectrometry
4.5. Influence of MPs on Trypsin Activity
4.5.1. Trypsin Activity in Bulk and MPs Soft Corona
4.5.2. Activity of Trypsin in the Hard Corona
4.6. Influence of MPs on In Vitro Digestion of Meat Extract Proteins with Trypsin
In Vitro Digestion of Meat Extract Proteins with Trypsin
4.7. Statistics and Graph Generation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koelmans, A.A.; Redondo-Hasselerharm, P.E.; Nor, N.H.M.; de Ruijter, V.N.; Mintenig, S.M.; Kooi, M. Risk Assessment of Microplastic Particles. Nat. Rev. Mater. 2022, 7, 138–152. [Google Scholar] [CrossRef]
- Di Fiore, C.; Carriera, F.; Russo, M.V.; Avino, P. Are Microplastics a Macro Issue? A Review on the Sources of Contamination, Analytical Challenges and Impact on Human Health of Microplastics in Food. Foods 2023, 12, 3915. [Google Scholar] [CrossRef] [PubMed]
- Giri, S.; Lamichhane, G.; Khadka, D.; Devkota, H.P. Microplastics Contamination in Food Products: Occurrence, Analytical Techniques and Potential Impacts on Human Health. Curr. Res. Biotechnol. 2024, 7, 100190. [Google Scholar] [CrossRef]
- Kumar, V.; Sharma, N.; Umesh, M.; Gupta, P.; Sharma, P.; Basheer, T.; Huligowda, L.K.D.; Thomas, J.; Bhagat, S.K.; Pasrija, R. Microplastics in Food: Occurrence, Toxicity, Green Analytical Detection Methods and Future Challenges. Green Anal. Chem. 2024, 11, 100152. [Google Scholar] [CrossRef]
- Udovicki, B.; Andjelkovic, M.; Cirkovic-Velickovic, T.; Rajkovic, A. Microplastics in Food: Scoping Review on Health Effects, Occurrence, and Human Exposure. Int. J. Food Contam. 2022, 9, 7. [Google Scholar] [CrossRef]
- Vitali, C.; Peters, R.J.B.; Janssen, H.-G.; Nielen, M.W.F. Microplastics and Nanoplastics in Food, Water, and Beverages; Part I. Occurrence. TrAC Trends Anal. Chem. 2023, 159, 116670. [Google Scholar] [CrossRef]
- Huerta Lwanga, E.; Mendoza Vega, J.; Ku Quej, V.; Chi, J.D.L.A.; Sanchez del Cid, L.; Chi, C.; Escalona Segura, G.; Gertsen, H.; Salánki, T.; van der Ploeg, M.; et al. Field Evidence for Transfer of Plastic Debris along a Terrestrial Food Chain. Sci. Rep. 2017, 7, 14071. [Google Scholar] [CrossRef]
- Habib, R.Z.; Poulose, V.; Alsaidi, R.; al Kendi, R.; Iftikhar, S.H.; Mourad, A.-H.I.; Kittaneh, W.F.; Thiemann, T. Plastic Cutting Boards as a Source of Microplastics in Meat. Food Addit. Contam. 2022, 39, 609–619. [Google Scholar] [CrossRef]
- Kaseke, T.; Lujic, T.; Cirkovic Velickovic, T. Nano- and Microplastics Migration from Plastic Food Packaging into Dairy Products: Impact on Nutrient Digestion, Absorption, and Metabolism. Foods 2023, 12, 3043. [Google Scholar] [CrossRef]
- Kedzierski, M.; Lechat, B.; Sire, O.; Le Maguer, G.; Le Tilly, V.; Bruzaud, S. Microplastic Contamination of Packaged Meat: Occurrence and Associated Risks. Food Packag. Shelf Life 2020, 24, 100489. [Google Scholar] [CrossRef]
- Li, D.; Shi, Y.; Yang, L.; Xiao, L.; Kehoe, D.K.; Gun’ko, Y.K.; Boland, J.J.; Wang, J.J. Microplastic Release from the Degradation of Polypropylene Feeding Bottles during Infant Formula Preparation. Nat. Food 2020, 1, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Mohamed Nor, N.H.; Kooi, M.; Diepens, N.J.; Koelmans, A.A. Lifetime Accumulation of Microplastic in Children and Adults. Environ. Sci. Technol. 2021, 55, 5084–5096. [Google Scholar] [CrossRef]
- Pletz, M. Ingested Microplastics: Do Humans Eat One Credit Card per Week? J. Hazard. Mater. Lett. 2022, 3, 100071. [Google Scholar] [CrossRef]
- Schwabl, P.; Köppel, S.; Königshofer, P.; Bucsics, T.; Trauner, M.; Reiberger, T.; Liebmann, B. Detection of Various Microplastics in Human Stool: A Prospective Case Series. Ann. Intern. Med. 2019, 171, 453–457. [Google Scholar] [CrossRef]
- Yan, Z.; Liu, Y.; Zhang, T.; Zhang, F.; Ren, H.; Zhang, Y. Analysis of Microplastics in Human Feces Reveals a Correlation between Fecal Microplastics and Inflammatory Bowel Disease Status. Environ. Sci. Technol. 2022, 56, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Li, Y.B.; He, H.R.; Zhang, J.F.; Ma, G.S. You Are What You Eat: Microplastics in the Feces of Young Men Living in Beijing. Sci. Total Environ. 2021, 767, 144345. [Google Scholar] [CrossRef]
- Ke, D.; Zheng, J.; Liu, X.; Xu, X.; Zhao, L.; Gu, Y.; Yang, R.; Liu, S.; Yang, S.; Du, J.; et al. Occurrence of Microplastics and Disturbance of Gut Microbiota: A Pilot Study of Preschool Children in Xiamen, China. eBioMedicine 2023, 97, 104828. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, L.; Trasande, L.; Kannan, K. Occurrence of Polyethylene Terephthalate and Polycarbonate Microplastics in Infant and Adult Feces. Environ. Sci. Technol. Lett. 2021, 8, 989–994. [Google Scholar] [CrossRef]
- Özsoy, S.; Gündogdu, S.; Sezigen, S.; Tasalp, E.; Ikiz, D.A.; Kideys, A.E. Presence of Microplastics in Human Stomachs. Forensic Sci. Int. 2024, 364, 112246. [Google Scholar] [CrossRef]
- Cetin, M.; Demirkaya Miloglu, F.; Kilic Baygutalp, N.; Ceylan, O.; Yildirim, S.; Eser, G.; Gul, H.İ. Higher Number of Microplastics in Tumoral Colon Tissues from Patients with Colorectal Adenocarcinoma. Environ. Chem. Lett. 2023, 21, 639–646. [Google Scholar] [CrossRef]
- Ibrahim, Y.S.; Tuan Anuar, S.; Azmi, A.A.; Wan Mohd Khalik, W.M.A.; Lehata, S.; Hamzah, S.R.; Ismail, D.; Ma, Z.F.; Dzulkarnaen, A.; Zakaria, Z.; et al. Detection of Microplastics in Human Colectomy Specimens. JGH Open 2021, 5, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Yin, K.; Wang, Y.; Zhao, H.; Wang, D.; Guo, M.; Mu, M.; Liu, Y.; Nie, X.; Li, B.; Li, J.; et al. A Comparative Review of Microplastics and Nanoplastics: Toxicity Hazards on Digestive, Reproductive and Nervous System. Sci. Total Environ. 2021, 774, 145758. [Google Scholar] [CrossRef]
- Monopoli, M.P.; Åberg, C.; Salvati, A.; Dawson, K.A. Biomolecular Coronas Provide the Biological Identity of Nanosized Materials. Nat. Nanotech. 2012, 7, 779–786. [Google Scholar] [CrossRef]
- Tan, H.; Yue, T.; Xu, Y.; Zhao, J.; Xing, B. Microplastics Reduce Lipid Digestion in Simulated Human Gastrointestinal System. Environ. Sci. Technol. 2020, 54, 12285–12294. [Google Scholar] [CrossRef]
- Zhu, H.; Wu, P.; Hu, Z.; Chen, H.; Wang, N.; Chen, X.D. Unraveling the Impact of Polystyrene Microplastics with Varying Particle Sizes and Concentrations on Lipid In Vitro Digestion and Ex Vivo Absorption. J. Hazard. Mater. 2025, 495, 138821. [Google Scholar] [CrossRef] [PubMed]
- de Guzman, M.K.; Stanic-Vucinic, D.; Gligorijevic, N.; Wimmer, L.; Gasparyan, M.; Lujic, T.; Vasovic, T.; Dailey, L.A.; Van Haute, S.; Cirkovic Velickovic, T. Small Polystyrene Microplastics Interfere with the Breakdown of Milk Proteins during Static In Vitro Simulated Human Gastric Digestion. Environ. Pollut. 2023, 335, 122282. [Google Scholar] [CrossRef]
- Pekar, J.; Ret, D.; Untersmayr, E. Stability of Allergens. Mol. Immunol. 2018, 100, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Wang, Z.; Wu, Y.; Chen, H.; Gao, J. Oral Exposure to Nano- and Microplastics: Potential Effects in Food Allergies? Allergy Med. 2024, 1, 100006. [Google Scholar] [CrossRef]
- Hou, G.; Hu, W.; Zhao, J.; Lu, J.; Zhang, W.; Liu, X.; Lu, S.; Shinichi, Y.; Ebere, E.C.; Wang, Q.; et al. Studies on Adsorption and Synergistic Biological Effects Induced by Microplastic Particles and the Platanus Pollen Allergenic Protein 3 (Pla a3). Environ. Pollut. 2025, 373, 126149. [Google Scholar] [CrossRef]
- DeLoid, G.M.; Cao, X.; Coreas, R.; Bitounis, D.; Singh, D.; Zhong, W.; Demokritou, P. Incineration-Generated Polyethylene Micro-Nanoplastics Increase Triglyceride Lipolysis and Absorption in an In Vitro Small Intestinal Epithelium Model. Environ. Sci. Technol. 2022, 56, 12288–12297. [Google Scholar] [CrossRef]
- Perusko, M.; Grundström, J.; Eldh, M.; Hamsten, C.; Apostolovic, D.; van Hage, M. The α-Gal Epitope—The Cause of a Global Allergic Disease. Front. Immunol. 2024, 15, 1335911. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-N.; Wen, B.; Zhu, J.-G.; Zhang, Y.-S.; Gao, J.-Z.; Chen, Z.-Z. Exposure to Microplastics Impairs Digestive Performance, Stimulates Immune Response and Induces Microbiota Dysbiosis in the Gut of Juvenile Guppy (Poecilia reticulata). Sci. Total. Environ. 2020, 733, 138929. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Huang, W.; Wei, S.; Shang, Y.; Gu, H.; Wu, F.; Lan, Z.; Hu, M.; Shi, H.; Wang, Y. Microplastics Impair Digestive Performance but Show Little Effects on Antioxidant Activity in Mussels under Low pH Conditions. Environ. Pollut. 2020, 258, 113691. [Google Scholar] [CrossRef]
- Xiao, K.; Song, L.; Li, Y.; Li, C.; Zhang, S. Dietary Intake of Microplastics Impairs Digestive Performance, Induces Hepatic Dysfunction, and Shortens Lifespan in the Annual Fish Nothobranchius guentheri. Biogerontology 2023, 24, 207–223. [Google Scholar] [CrossRef]
- Frank, Y.A.; Interesova, E.A.; Solovyev, M.M.; Xu, J.; Vorobiev, D.S. Effect of Microplastics on the Activity of Digestive and Oxidative-Stress-Related Enzymes in Peled Whitefish (Coregonus peled Gmelin) Larvae. Int. J. Mol. Sci. 2023, 24, 10998. [Google Scholar] [CrossRef]
- Romano, N.; Ashikin, M.; Teh, J.C.; Syukri, F.; Karami, A. Effects of Pristine Polyvinyl Chloride Fragments on Whole Body Histology and Protease Activity in Silver Barb Barbodes gonionotus Fry. Environ. Pollut. 2018, 237, 1106–1111. [Google Scholar] [CrossRef]
- Liu, G.; Jiang, Q.; Qin, L.; Zeng, Z.; Zhang, P.; Feng, B.; Liu, X.; Qing, Z.; Qing, T. The Influence of Digestive Tract Protein on Cytotoxicity of Polyvinyl Chloride Microplastics. Sci. Total Environ. 2024, 945, 174023. [Google Scholar] [CrossRef]
- Santos, A.M.C.; de Oliveira, J.S.; Bittar, E.R.; da Silva, A.L.; Guia, M.L.D.M.; Bemquerer, M.P.; Santoro, M.M. Improved Purification Process of β- and α-Trypsin Isoforms by Ion-Exchange Chromatography. Braz. Arch. Biol. Technol. 2008, 51, 711–721. [Google Scholar] [CrossRef]
- Perutka, Z.; Šebela, M. Pseudotrypsin: A Little-Known Trypsin Proteoform. Molecules 2018, 23, 2637. [Google Scholar] [CrossRef]
- Rabe, M.; Verdes, D.; Seeger, S. Understanding Protein Adsorption Phenomena at Solid Surfaces. Adv. Colloid Interface Sci. 2011, 162, 87–106. [Google Scholar] [CrossRef]
- Yue, Y.; Tu, Q.; Guo, Y.; Wang, Y.; Xu, Y.; Zhang, Y.; Liu, J. Comparison of the Interactions of Fanetizole with Pepsin and Trypsin: Spectroscopic and Molecular Docking Approach. J. Mol. Liq. 2022, 365, 120095. [Google Scholar] [CrossRef]
- Gilliland, G.L.; Teplyakov, A. Structural Calcium (Trypsin, Subtilisin). In Encyclopedia of Inorganic and Bioinorganic Chemistry, 2nd ed.; Scott, R.A., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2011; ISBN 978-1-119-95143-8. [Google Scholar]
- Koutsopoulos, S.; Patzsch, K.; Bosker, W.T.E.; Norde, W. Adsorption of Trypsin on Hydrophilic and Hydrophobic Surfaces. Langmuir 2007, 23, 2000–2006. [Google Scholar] [CrossRef]
- Santos, A.M.C.; Santana, M.A.; Gomide, F.T.F.; Miranda, A.A.C.; Oliveira, J.S.; Vilas Boas, F.A.S.; Vasconcelos, A.B.; Bemquerer, M.P.; Santoro, M.M. Physical-Chemical Characterization and Stability Study of Alpha-Trypsin at pH 3.0 by Differential Scanning Calorimetry. Int. J. Biol. Macromol. 2008, 42, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Covello, C.; Di Vincenzo, F.; Cammarota, G.; Pizzoferrato, M. Micro(Nano)Plastics and Their Potential Impact on Human Gut Health: A Narrative Review. Curr. Issues Mol. Biol. 2024, 46, 2658–2677. [Google Scholar] [CrossRef]
- Jones, L.R.; Wright, S.J.; Gant, T.W. A Critical Review of Microplastics Toxicity and Potential Adverse Outcome Pathway in Human Gastrointestinal Tract Following Oral Exposure. Toxicol. Lett. 2023, 385, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Min, K.; Cuiffi, J.D.; Mathers, R.T. Ranking Environmental Degradation Trends of Plastic Marine Debris Based on Physical Properties and Molecular Structure. Nat. Commun. 2020, 11, 727. [Google Scholar] [CrossRef]
- Anand, G.; Sharma, S.; Dutta, A.K.; Kumar, S.K.; Belfort, G. Conformational Transitions of Adsorbed Proteins on Surfaces of Varying Polarity. Langmuir 2010, 26, 10803–10811. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, Y.; Hu, S.; Zong, W.; Liu, R. New Mechanistic Insights of Nanoplastics Synergistic Cadmium Induced Overactivation of Trypsin: Joint Analysis from Protein Multi-Level Conformational Changes and Computational Modeling. J. Hazard. Mater. 2024, 480, 135817. [Google Scholar] [CrossRef]
- Cordeiro, A.L.; Rückel, M.; Bartels, F.; Maitz, M.F.; Renner, L.D.; Werner, C. Protein Adsorption Dynamics to Polymer Surfaces Revisited-A Multisystems Approach. Biointerphases 2019, 14, 051005. [Google Scholar] [CrossRef]
- Arai, T.; Norde, W. The Behavior of Some Model Proteins at Solid-Liquid Interfaces 1. Adsorption from Single Protein Solutions. Colloids Surf. 1990, 51, 1–15. [Google Scholar] [CrossRef]
- Coglitore, D.; Janot, J.-M.; Balme, S. Protein at Liquid Solid Interfaces: Toward a New Paradigm to Change the Approach to Design Hybrid Protein/Solid-State Materials. Adv. Colloid. Interface Sci. 2019, 270, 278–292. [Google Scholar] [CrossRef] [PubMed]
- Baron, M.H.; Revault, M.; Servagent-Noinville, S.; Abadie, J.; Quiquampoix, H. Chymotrypsin Adsorption on Montmorillonite: Enzymatic Activity and Kinetic FTIR Structural Analysis. J. Colloid Interface Sci. 1999, 214, 319–332. [Google Scholar] [CrossRef]
- Winkler, F.K.; D’Arcy, A.; Hunziker, W. Structure of Human Pancreatic Lipase. Nature 1990, 343, 771–774. [Google Scholar] [CrossRef] [PubMed]
- Roussel, A.; Canaan, S.; Egloff, M.-P.; Rivière, M.; Dupuis, L.; Verger, R.; Cambillau, C. Crystal Structure of Human Gastric Lipase and Model of Lysosomal Acid Lipase, Two Lipolytic Enzymes of Medical Interest. J. Biol. Chem. 1999, 274, 16995–17002. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.F.M.; van Mierlo, C.P.M.; Visser, A.J.W.G. Kinetic and Structural Characterization of Adsorption-Induced Unfolding of Bovine α-Lactalbumin. J. Biol. Chem. 2002, 277, 10922–10930. [Google Scholar] [CrossRef]
- Qiu, Z.; Shi, Y.; Zheng, Y.; Shi, W.; Zhang, L.; Yin, M.; Wang, X. Comparison of in Vitro Digestive Characteristics of Proteins from Different Sources in Simulated Elderly Gastrointestinal Conditions. Food Chem. 2025, 463, 141299. [Google Scholar] [CrossRef]
- Benedé, S.; López-Fandiño, R.; Molina, E. Residual α-Gal in Digested Beef, Pork and Lamb Meat Submitted to Different Cooking Methods. LWT 2024, 204, 116441. [Google Scholar] [CrossRef]
- Jin, Y.; Lu, L.; Tu, W.; Luo, T.; Fu, Z. Impacts of Polystyrene Microplastic on the Gut Barrier, Microbiota and Metabolism of Mice. Sci. Total Environ. 2019, 649, 308–317. [Google Scholar] [CrossRef]
- Lu, L.; Wan, Z.; Luo, T.; Fu, Z.; Jin, Y. Polystyrene Microplastics Induce Gut Microbiota Dysbiosis and Hepatic Lipid Metabolism Disorder in Mice. Sci. Total Environ. 2018, 631–632, 449–458. [Google Scholar] [CrossRef]
- Li, B.; Ding, Y.; Cheng, X.; Sheng, D.; Xu, Z.; Rong, Q.; Wu, Y.; Zhao, H.; Ji, X.; Zhang, Y. Polyethylene Microplastics Affect the Distribution of Gut Microbiota and Inflammation Development in Mice. Chemosphere 2020, 244, 125492. [Google Scholar] [CrossRef]
- Schippa, S.; Conte, M.P. Dysbiotic Events in Gut Microbiota: Impact on Human Health. Nutrients 2014, 6, 5786–5805. [Google Scholar] [CrossRef]
- Herath, M.; Hosie, S.; Bornstein, J.C.; Franks, A.E.; Hill-Yardin, E.L. The Role of the Gastrointestinal Mucus System in Intestinal Homeostasis: Implications for Neurological Disorders. Front. Cell Infect. Microbiol. 2020, 10, 248. [Google Scholar] [CrossRef] [PubMed]
- Phue, W.H.; Xu, K.; George, S. Inorganic Food Additive Nanomaterials Alter the Allergenicity of Milk Proteins. Food Chem. Toxicol. 2022, 162, 112874. [Google Scholar] [CrossRef] [PubMed]
- Maddah, H.A. Polypropylene as a Promising Plastic: A Review. Am. J. Polym. Sci. 2016, 6, 1–11. [Google Scholar] [CrossRef]
- Thomsen, T.B.; Hunt, C.J.; Meyer, A.S. Influence of Substrate Crystallinity and Glass Transition Temperature on Enzymatic Degradation of Polyethylene Terephthalate (PET). N. Biotechnol. 2022, 69, 28–35. [Google Scholar] [CrossRef]
- Ducoli, S.; Federici, S.; Nicsanu, R.; Zendrini, A.; Marchesi, C.; Paolini, L.; Radeghieri, A.; Bergese, P.; Depero, L.E. A Different Protein Corona Cloaks “True-to-Life” Nanoplastics with Respect to Synthetic Polystyrene Nanobeads. Environ. Sci. Nano 2022, 9, 1414–1426. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST Static in Vitro Simulation of Gastrointestinal Food Digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Buck, F.F.; Vithayathil, A.J.; Bier, M.; Nord, F.F. On the Mechanism of Enzyme Action. 73. Studies on Trypsins from Beef, Sheep and Pig Pancreas. Arch. Biochem. Biophys. 1962, 97, 417–424. [Google Scholar] [CrossRef]
- Magrì, D.; Sánchez-Moreno, P.; Caputo, G.; Gatto, F.; Veronesi, M.; Bardi, G.; Catelani, T.; Guarnieri, D.; Athanassiou, A.; Pompa, P.P.; et al. Laser Ablation as a Versatile Tool To Mimic Polyethylene Terephthalate Nanoplastic Pollutants: Characterization and Toxicology Assessment. ACS Nano 2018, 12, 7690–7700. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Chevallet, M.; Luche, S.; Rabilloud, T. Silver Staining of Proteins in Polyacrylamide Gels. Nat. Protoc. 2006, 1, 1852–1858. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Camacho, I.; Rosas-Murrieta, N. Leucyl Aminopeptidase yspII (Yeast). In Handbook of Proteolytic Enzymes, 3rd ed.; Rawlings, N.D., Salvesen, G., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 1476–1480. ISBN 978-0-12-382219-2. [Google Scholar]
- Apostolovic, D.; Tran, T.A.T.; Hamsten, C.; Starkhammar, M.; Cirkovic Velickovic, T.; van Hage, M. Immunoproteomics of Processed Beef Proteins Reveal Novel Galactose-α-1,3-Galactose-Containing Allergens. Allergy 2014, 69, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Hummel, B.C. A Modified Spectrophotometric Determination of Chymotrypsin, Trypsin, and Thrombin. Can. J. Biochem. Physiol. 1959, 37, 1393–1399. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isotherm Model | Parameters of Isotherms PET | |
|---|---|---|
| Langmuir | KL (mL/mg) | 10.15 |
| QL (mg/g) | 4.16 | |
| RL | 0.078 | |
| R2 | 0.9623 | |
| Freundlich | KF ((mg/g) * (mL/mg)1/n) | 4.08 |
| n | 3.7 | |
| R2 | 0.8267 | |
| Redlich–eterson | KR (mL/mg) | 28.2 |
| α (mL/mg) | 1.2 | |
| β | 1.26 | |
| R2 | 0.9970 | |
| Guggenheim–Anderson–de Boer | QG (mg/g) | 7.15 |
| Khard G (mL/mg) | 4.5 | |
| Ksoft G (mL/mg) | −0.56 | |
| R2 | 0.9945 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lujic, T.; Gligorijevic, N.; Stanic-Vucinic, D.; Krstic Ristivojevic, M.; Mutic, T.; Wimmer, L.; Dailey, L.A.; Cirkovic Velickovic, T. Effects of Polypropylene and Polyethylene Terephthalate Microplastics on Trypsin Structure and Function. Int. J. Mol. Sci. 2025, 26, 5974. https://doi.org/10.3390/ijms26135974
Lujic T, Gligorijevic N, Stanic-Vucinic D, Krstic Ristivojevic M, Mutic T, Wimmer L, Dailey LA, Cirkovic Velickovic T. Effects of Polypropylene and Polyethylene Terephthalate Microplastics on Trypsin Structure and Function. International Journal of Molecular Sciences. 2025; 26(13):5974. https://doi.org/10.3390/ijms26135974
Chicago/Turabian StyleLujic, Tamara, Nikola Gligorijevic, Dragana Stanic-Vucinic, Maja Krstic Ristivojevic, Tamara Mutic, Lukas Wimmer, Lea Ann Dailey, and Tanja Cirkovic Velickovic. 2025. "Effects of Polypropylene and Polyethylene Terephthalate Microplastics on Trypsin Structure and Function" International Journal of Molecular Sciences 26, no. 13: 5974. https://doi.org/10.3390/ijms26135974
APA StyleLujic, T., Gligorijevic, N., Stanic-Vucinic, D., Krstic Ristivojevic, M., Mutic, T., Wimmer, L., Dailey, L. A., & Cirkovic Velickovic, T. (2025). Effects of Polypropylene and Polyethylene Terephthalate Microplastics on Trypsin Structure and Function. International Journal of Molecular Sciences, 26(13), 5974. https://doi.org/10.3390/ijms26135974