
Regulators and Conductors of Immunity: Natural Immune System in Health and Autoimmunity


Abstract
1. Introduction
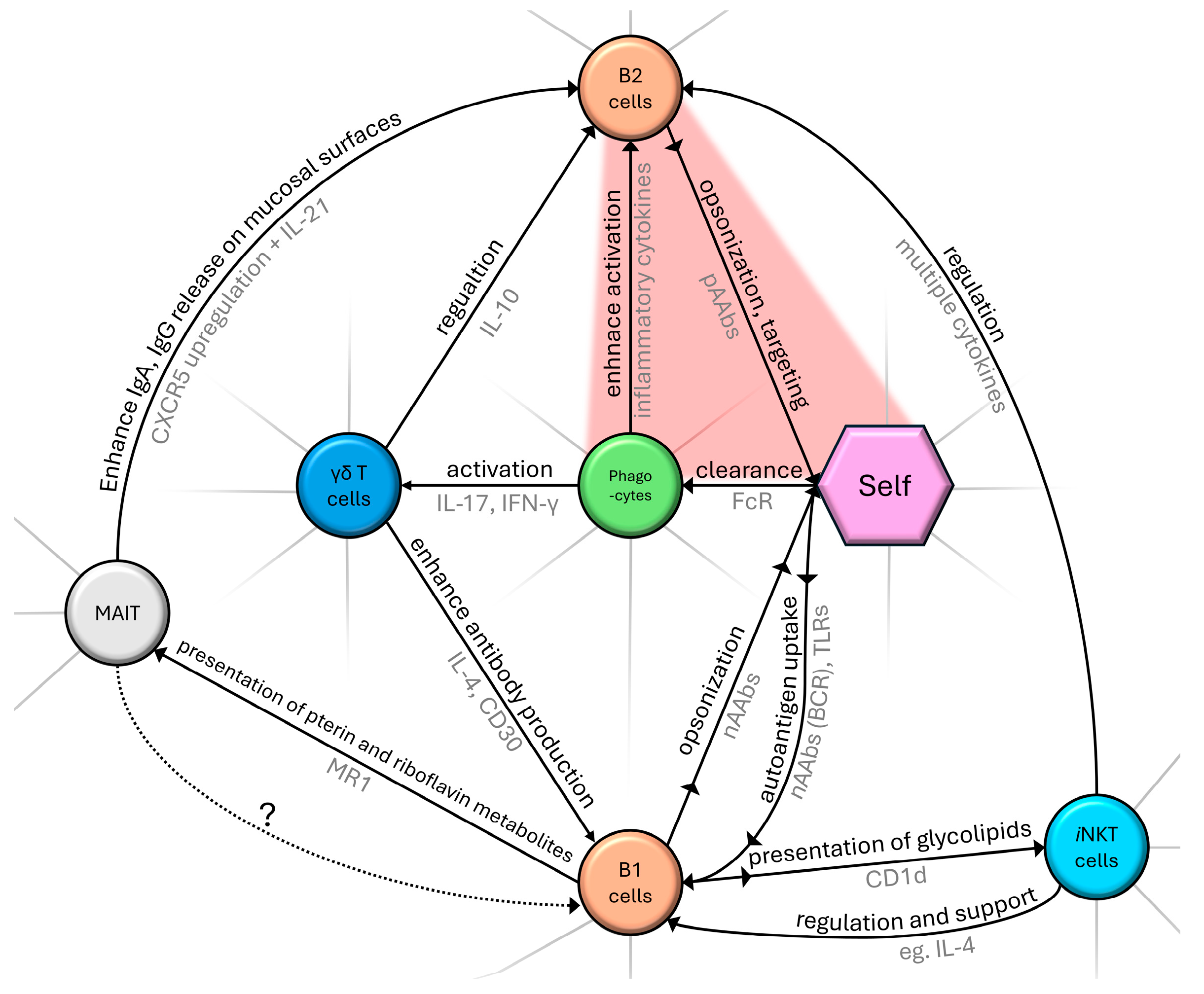
2. Cellular and Humoral Components and Their Role in the Natural Immune System
2.1. Potentially Mutual Influence of γδ T Cells and nAAb Network
2.2. Connections of iNKT and B-1 Cells
2.3. Conjecture of MAIT and B-1 Cell Interaction
2.4. B-1 Cells and Natural Autoantibodies (nAAbs)
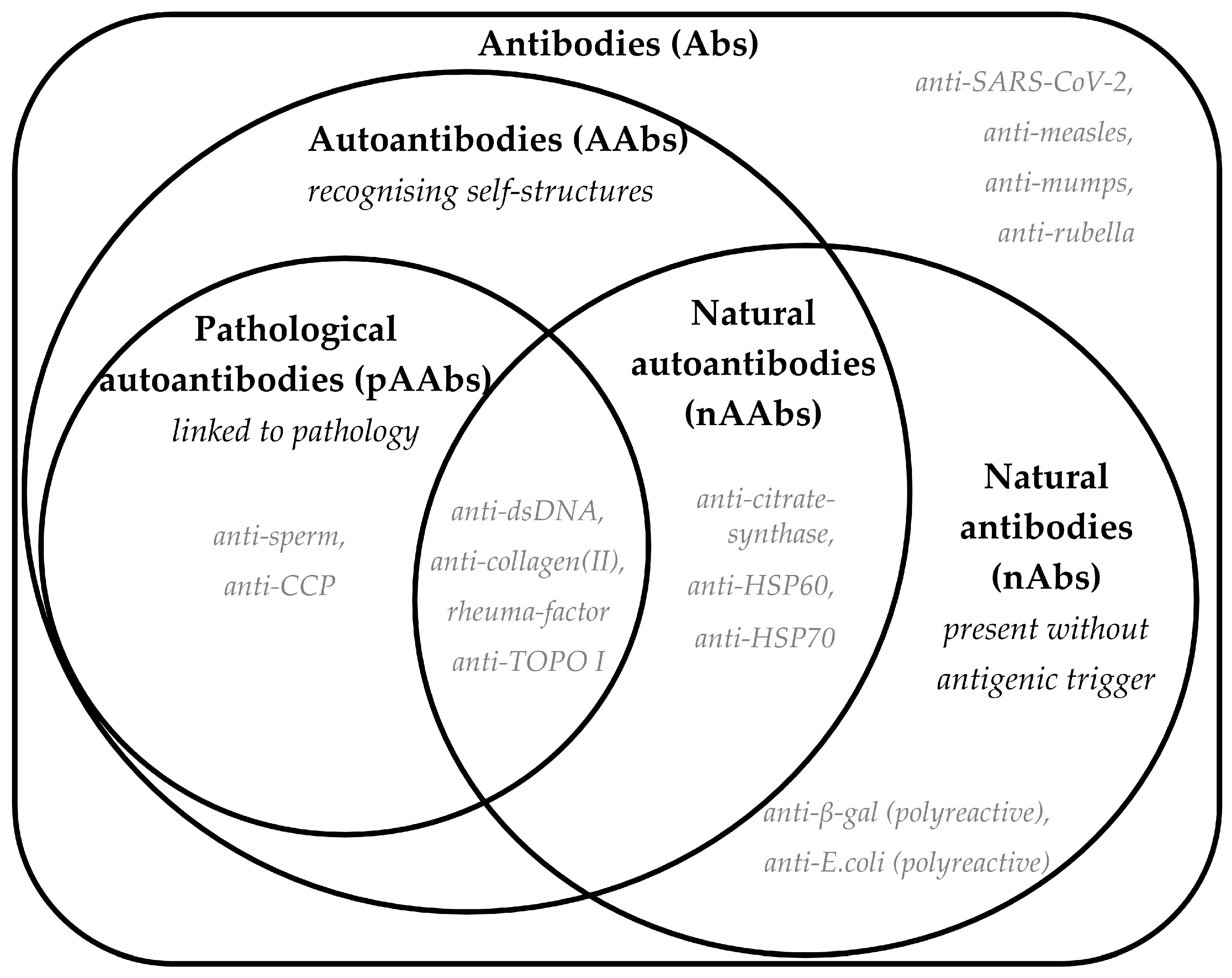
2.5. Role of Natural Autoantibodies (nAAbs)
2.6. Natural IgM Autoantibodies
2.7. The Dual Nature of Natural IgG Autoantibodies: Implications for Immune Tolerance and Autoimmune Disease Development
3. Regulatory Role of the Natural Immune System in Pathological Conditions
3.1. From Clonal Selection to Self-Assessment: The Development of Autoreactivity in Immunology
3.2. Shifting Balance Between Physiological and Pathological Autoimmunity
3.3. Challenging Conventional Views: Natural Autoantibodies and Their Dynamic Responses in Health and Disease
3.4. Natural Autoantibodies in Health and Disease: Interplay Between Immunological Response and Pathogenesis
3.5. From BCG and SARS-CoV-2 to Natural Autoantibodies: Investigating the Non-Specific Immune Enhancements and Their Mechanisms
3.6. Natural Autoantibodies as Biomarkers and Modulators in Autoimmune Disorders: From Systemic Sclerosis Through Type-1 Diabetes to Hashimoto Thyroiditis in Pregnancy
3.7. Antibodies Against Complex Self-Patterns: The Case of AMPAs and aPls
- By either:
- ∘
- ∘
- Both processes are hallmarked by chronic activation [3].
4. Concluding Remarks
5. Implications of the Study
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| aCarb-β2-GPI | anti-carbamylated-β2-glycoprotein I |
| aCL | anti-cardiolipin |
| ACPA | anti-citrullinated protein antibody |
| AMPA | anti-modified protein antibody |
| aPl | anti-phospholipid |
| aPS/PT | anti-phosphatidylserine/prothrombin complex |
| aVim/CL | anti-vimentin/cardiolipin complex |
| aβ2-GPI | anti-β2-GP I |
| BCG vaccine | Bacille Calmette–Guérin vaccine |
| BCR | B cell receptor |
| CCP | cyclic-citrullinated peptide |
| CCR | chemokine receptor |
| CD | cluster of differentiation |
| CS | citrate synthase |
| DAMP | damage-associated molecular pattern |
| dsDNA | double-stranded deoxyribonucleic acid |
| dsDNA | double-stranded DNA |
| F4 | fragment 4 of the human topoisomerase I |
| HCV | hepatitis C virus |
| HSP | heat-shock protein |
| IFN | interferon |
| Ig | immunoglobulin |
| IL | interleukin |
| iNKTs | invariant natural killer T cells |
| IVIG | intravenous immunoglobulin |
| LAC | lupus anticoagulant |
| LPS | lipopolysaccharide |
| MAIT | mucosa-associated invariant T cells |
| MHC | major histocompatibility complex |
| MMR | mumps, measles, rubella |
| MZ | marginal zone |
| nAAb | natural autoantibody |
| nAb | natural antibody |
| NSE | non-specific effects |
| NZB mouse | New Zealand black mouse |
| pAAbs | pathological autoantibody |
| PTM | post-translationally modified |
| RA | rheumatoid arthritis |
| RBC | red blood cell |
| SARS-CoV-2 | severe acute respiratory syndrome coronavirus 2 |
| SLE | systemic lupus erythematosus |
| SSc | systemic sclerosis |
| ssDNA | single-stranded deoxyribonucleic acid |
| STAT-6 | signal transducer and activator of transcription 6 |
| T1D | type 1 diabetes |
| TCR | T cell receptor |
| TLR | Toll-like receptor |
| TOPO | topoisomerase |
| αβ T cell | alpha–beta T cell |
| γδ T cell | gamma–delta T cell |
Appendix A

{kind=link}
{kind=link}
| Year | Scientific Question | Immunoglobulin Isotype-Related Conclusion (If Applicable) | Reference | |
|---|---|---|---|---|
| 1991 | How might the presence of natural autoantibodies affect the overall health and immune response of an unimmunized animal? | The immune system in unimmunized animals produces polyreactive and monoreactive IgM and IgG natural autoantibodies, encoded by unmutated germ-line genes, which interact with self-antigens to form a dynamic network supporting organismal homeostasis. | [71] | |
| 1995 | In what ways do natural autoantibodies contribute to immune regulation and homeostasis? | Natural autoantibodies are conserved throughout life, maintaining stability without affinity maturation. Encoded by germline genes, these multireactive antibodies are found across individuals and species, supporting immune regulation, homeostasis, infection resistance, and modulation of biological molecules. | [117] | |
| 1998 | What roles do natural autoantibodies (nAAbs) play in the immune system? | Natural antibodies, present in healthy individuals without prior immunization, often target self-antigens and are termed natural autoantibodies (nAAbs). Historically overlooked in immune regulation, nAAbs are now recognized as prevalent across vertebrate species, selected early in development, and essential for immune homeostasis. This study reviews the contemporary (and foundational) understanding of nAAb properties and their potential for therapeutic use. | [174] | |
| 1999 | How do natural antibodies influence pathogen spread and immune response in the body? | Natural antibodies, often dismissed as “background”, are essential for immunity against infections. In antibody-free mice, viral and bacterial titers in vital organs were significantly higher, while titers in lymphoid organs were lower, compared to antibody-competent mice. This suggests that natural antibodies help prevent pathogen spread to vital organs and enhance immunogenicity by improving antigen trapping in secondary lymphoid organs. | [118] | |
| 2006 | Are the anti-mitochondrial citrate synthase autoantibodies components of the natural antibody network? | Natural IgM autoantibodies to citrate synthase (CS) are present from infancy, are stable in adults, potentially function as a first line of defense against pathogens and differ in epitope recognition under pathological conditions like SLE, indicating a possible link between innate immune responses and autoimmune disease processes. | [97] | |
| 2006 | Is the natural antibody repertoire linked to the host biome? | Habitat (wild vs. laboratory) had a stronger impact on immunoglobulin levels than age, strain, or gender, with wild rodents showing heightened protective immune responses similar to both autoimmune (Th1-IgG) and allergic (Th2-IgE) reactions. Wild rats have significantly higher levels of autoreactive, polyreactive IgG, but not IgM, compared to laboratory rats, both quantitatively and qualitatively. | [121] | |
| 2008 | Could the presence of naturally occurring anti-hCS antibodies indicate a broader mechanism linking innate immunity to autoimmunity? | Human sera contain naturally occurring IgM antibodies recognizing human citrate synthase (hCS), with distinct epitope patterns in healthy and SLE conditions. Sera affinity-purified on hCS also cross-reacts with bacterial CS and nucleosome antigens, suggesting that these antibodies may both contribute to the innate defense and recognize autoantigens in systemic autoimmune disease. | [175] | |
| 2008 | What constituents can be found in cryoglobulins appearing during hepatitis C virus infection? | IgM nAAbs against anti-HCV IgG1/Κ Fab (VH1-69) expand upon hepatitis C virus infection, leading to mixed cryoglobulinemia. | [122] | |
| 2010 | How do anti-human Hsp60 autoantibodies (known riskfactor of atherosclerosis) change between a mother’s and newborn’s umbilical cord blood, and in adults? Is it a nAAb? | Levels of IgM anti-Hsp60 did not correlate to maternal concentrations. nAAb IgM anti-HSP60 level does not change in a 5-year-long period, supporting the idea that the immune system in a tightly regulated manner selectively favors autoreactive B cells targeting a core set of immunodominant self-antigens, including Hsp60. | [123] | |
| 2013 | Are there potential IgG autoantibody markers with clinical prevalence in non-immunological diseases? | IgG nAAb diversity in serum increases with age and is generally higher in females than in males, while certain diseases like Alzheimer’s, Parkinson’s, and multiple sclerosis are associated with fewer detectable autoantibodies, possibly reflecting disease-related immune modulation. | [90] | |
| 2016 | What role does natural IgM play in preventing autoimmune responses from autoreactive B and T cells that have bypassed tolerance mechanisms? | Polyreactive natural IgM autoantibodies (IgM-NAA) protect against pathogens and neo-antigens while inhibiting autoimmune inflammation through anti-idiotypic mechanisms, enhancing apoptotic cell clearance, masking neo-antigens, and modulating dendritic and effector cell function. Natural IgM also prevents autoimmune disorders driven by pathogenic IgG autoantibodies, autoreactive B and T cells, and genetic factors, such as in SLE. | [27] | |
| 2017 | Can the immunization of rats with specific allergens enhance natural antibody networks? | nAAbs from immunized rat sera recognized more self-antigens across all organ extracts, with differences less pronounced in IgG than in IgM. | Immunization of laboratory rats with different allergens was found to enhance networks of natural antibodies (with a more marked effect on IgM than on IgG). | [91] |
| 2018 | Do vaccines (like BCG) provide broader health benefits beyond specific disease protection by enhancing innate immune memory through trained immunity? | The non-specific effects (NSEs) of the Bacille Calmette–Guérin (BCG) vaccine may involve both adaptive and innate immune mechanisms, with evidence suggesting a key role for trained immunity. This memory-like feature of innate immune cells results from epigenetic reprogramming after exposure to a primary stimulus like BCG, subsequently influencing cytokine production and cell metabolism. | [141] | |
| 2020 | What functions can B-1a cells perform in anti-viral immunology? | B-1a cells are a unique B lymphocyte subpopulation essential for natural antibody production, innate immunity, and immunoregulation. They produce IgM, IL-10, and GM-CSF, which neutralize pathogens, modulate cytokine storms, and enhance IgM production, respectively. B-1a cells have shown protective effects against infections like influenza, sepsis, and pneumonia, highlighting their potential role in immune defense against SARS-CoV-2 and other infections. | [176] | |
| 2021 | Is there an association between autoimmune disease-specific pathological autoantibodies and the natural autoantibody pool in different autoimmune diseases (SSc, SLE, RA)? | The levels of anti-F4 and anti-CS IgG antibodies were significantly increased in anti-dsDNA IgG-positive compared to anti-dsDNA IgG-negative SLE patients. | The levels of anti-F4 and anti-CS IgM natural antibodies were significantly elevated in anti-dsDNA IgM-positive compared to anti-dsDNA IgM-negative SLE patients. | [147] |
| 2021 | Is there an association between an external antigenic trigger (anti-measles vaccination or natural measles virus infection) and the consequently formed and still persisting antibodies and the nAAb pool in different autoimmune diseases (SSc, SLE, RA)? | Significantly higher levels of natural anti-CS IgG were detected in anti-measles IgG-seropositive compared to -seronegative samples in RA, SLE, and SSc. | [147] | |
| 2021 | How do B cell subgroups and anti-citrate synthase nAAbs reflect immunological dysregulation present in systemic sclerosis? | IgG anti-CS nAAbs are associated with active systemic sclerosis (SSc) and may indicate compensatory immune responses that fail to counteract disease progression, highlighting their potential as supplementary biomarkers alongside DN1 B cell ratios for disease activity assessment in SSc. | [148] | |
| 2022 | Case study—can the appearance of natural alloantibodies without mutation or class-switching suggest a unique immune activation pathway in severe COVID-19? | In a lethal COVID-19 case, a de novo natural IgM lambda alloantibody emerged, targeting the M antigen of the MNS blood group on RBCs, without cross-reacting with SARS-CoV-2 antigens. This first report of a bystander natural alloantibody against RBCs underscores the extra-follicular humoral response in severe COVID-19. | [143] | |
| 2023 | How might understanding NSEs impact the development of future vaccine strategies and public health policies? | Live attenuated vaccines can lead to significant reductions in mortality and morbidity, while some non-live vaccines may increase these outcomes. Non-specific effects (NSEs) might stem from trained innate immunity, emergency granulopoiesis, and heterologous T cell immunity. These findings indicate that vaccine testing and regulation should account for NSEs. | [142] | |
| 2023 | Is there an association between the immune response triggered by different COVID-19 vaccines and the natural autoantibodies? | A statistically significant positive connection was observed between anti-HSP60, anti-HSP70, and anti-CS IgG titers and anti-SARS-CoV-2 IgG-positive serum levels, especially in mRNA vaccine recipients. | A positive correlation was found between anti-CS IgM nAAbs and the immune response, with elevated anti-CS IgM levels in cases of positive anti-SARS-CoV-2 IgG, IgA, and interferon-γ results. | [144] |
| 2023 | Is there an association between recent/aged antigenic triggers and the nAAb repertoire? | The study found statistically significant associations between SARS-CoV-2 vaccination (in terms of recent antigenic trigger) and increased levels of IgG natural autoantibodies (nAAbs). | Significant associations were found between anti-CS IgM levels and the presence of IgG antibodies specific to measles, mumps, and rubella, suggesting an interaction between nAAbs and antibodies from an “aged” (MMR vaccinations or infections) antigenic trigger. | [145] |
| 2023 | What unknown factors could be influencing the preferentialskewing of autoreactive VH4-34 antibodies toward the B1 cell population? | The study suggests that B cells with autoreactive B cell receptors may preferentially accumulate in the B1 cell pool or be excluded from the memory B cell pool. Alternatively, other factors might direct autoreactive VH4-34 antibodies toward the B1 population, with certain light chains reportedly suppressing inherent autoreactivity in these antibodies. | [177] | |
| 2023 | Can the insights into B-1 B cells and natural antibodies influence future therapeutic strategies for preventing or treating T1D? | B-1 B cell-derived GlcNAc-specific IgM binds β cell antigens, suppressing diabetogenic T cells and delaying T1D in recipients, suggesting a protective role in T1D. | [149] | |
| 2024 | Are there correlations between dcSSc-associated anti-Scl-70,anti-CS natural autoantibodies, and complement component C3 levels? | The negative correlation between serum natural autoantibodies (CS IgM) and complement component C3 in dcSSc suggests that natural autoantibodies may trigger C3 activation and, therefore, consumption, potentially leading to tissue damage. | [150] | |
| 2024 | How do nAAb levels change in healthy and Hashimoto thyroiditis patients throughout pregnancy? | Pregnant women with Hashimoto thyroiditis have elevated anti-Hsp60 and anti-Hsp70 IgM nAAbs from the first trimester onward, accompanied by lower anti-Hsp70 and Hsp60 IgG nAAb levels in the third trimester, suggesting a compensatory mechanism that may contribute to maternal tolerance towards the fetus. | [151] | |
References
- Cohen, I.R. The Cognitive Principle Challenges Clonal Selection. Immunol. Today 1992, 13, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Loh, L.; Gherardin, N.A.; Sant, S.; Grzelak, L.; Crawford, J.C.; Bird, N.L.; Koay, H.-F.; van de Sandt, C.E.; Moreira, M.L.; Lappas, M.; et al. Human Mucosal-Associated Invariant T Cells in Older Individuals Display Expanded TCRαβ Clonotypes with Potent Antimicrobial Responses. J. Immunol. 2020, 204, 1119–1133. [Google Scholar] [CrossRef] [PubMed]
- Kurioka, A.; Klenerman, P. Aging Unconventionally: Γδ T Cells, iNKT Cells, and MAIT Cells in Aging. Semin. Immunol. 2023, 69, 101816. [Google Scholar] [CrossRef] [PubMed]
- Rampoldi, F.; Donato, E.; Ullrich, L.; Deseke, M.; Janssen, A.; Demera, A.; Sandrock, I.; Bubke, A.; Juergens, A.-L.; Swallow, M.; et al. Γδ T Cells License Immature B Cells to Produce a Broad Range of Polyreactive Antibodies. Cell Rep. 2022, 39, 110854. [Google Scholar] [CrossRef]
- Carding, S.R.; Egan, P.J. Γδ T Cells: Functional Plasticity and Heterogeneity. Nat. Rev. Immunol. 2002, 2, 336–345. [Google Scholar] [CrossRef]
- Tough, D.F.; Sprent, J. Lifespan of γ/δ T Cells. J. Exp. Med. 1998, 187, 357–365. [Google Scholar] [CrossRef]
- Born, W.K.; Huang, Y.; Zeng, W.; Torres, R.M.; O’Brien, R.L. A Special Connection between Γδ T Cells and Natural Antibodies? Arch. Immunol. Ther. Exp. 2016, 64, 455–462. [Google Scholar] [CrossRef]
- Zhao, Y.; Lin, L.; Xiao, Z.; Li, M.; Wu, X.; Li, W.; Li, X.; Zhao, Q.; Wu, Y.; Zhang, H.; et al. Protective Role of Γδ T Cells in Different Pathogen Infections and Its Potential Clinical Application. J. Immunol. Res. 2018, 2018, 5081634. [Google Scholar] [CrossRef]
- Shiromizu, C.M.; Jancic, C.C. Γδ T Lymphocytes: An Effector Cell in Autoimmunity and Infection. Front. Immunol. 2018, 9, 2389. [Google Scholar] [CrossRef]
- Hu, Y.; Hu, Q.; Li, Y.; Lu, L.; Xiang, Z.; Yin, Z.; Kabelitz, D.; Wu, Y. Γδ T Cells: Origin and Fate, Subsets, Diseases and Immunotherapy. Signal Transduct. Target. Ther. 2023, 8, 1–38. [Google Scholar] [CrossRef]
- Latha, T.S.; Reddy, M.C.; Durbaka, P.V.R.; Rachamallu, A.; Pallu, R.; Lomada, D. Γδ T Cell-Mediated Immune Responses in Disease and Therapy. Front. Immunol. 2014, 5, 571. [Google Scholar] [CrossRef] [PubMed]
- Kabelitz, D.; Peters, C.; Wesch, D.; Oberg, H.-H. Regulatory Functions of Γδ T Cells. Int. Immunopharmacol. 2013, 16, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Lafont, V.; Sanchez, F.; Laprevotte, E.; Michaud, H.-A.; Gros, L.; Eliaou, J.-F.; Bonnefoy, N. Plasticity of Gamma Delta T Cells: Impact on the Anti-Tumor Response. Front. Immunol. 2014, 5, 622. [Google Scholar] [CrossRef] [PubMed]
- Saini, C.; Tarique, M.; Ramesh, V.; Khanna, N.; Sharma, A. γδ T Cells Are Associated with Inflammation and Immunopathogenesis of Leprosy Reactions. Immunol. Lett. 2018, 200, 55–65. [Google Scholar] [CrossRef]
- Ou, Q.; Power, R.; Griffin, M.D. Revisiting Regulatory T Cells as Modulators of Innate Immune Response and Inflammatory Diseases. Front. Immunol. 2023, 14, 1287465. [Google Scholar] [CrossRef]
- Krovi, S.H.; Gapin, L. Invariant Natural Killer T Cell Subsets—More Than Just Developmental Intermediates. Front. Immunol. 2018, 9, 1393. [Google Scholar] [CrossRef]
- Terabe, M.; Berzofsky, J.A. Tissue-Specific Roles of NKT Cells in Tumor Immunity. Front. Immunol. 2018, 9, 1838. [Google Scholar] [CrossRef]
- Middendorp, S.; Nieuwenhuis, E.E.S. NKT Cells in Mucosal Immunity. Mucosal Immunol. 2009, 2, 393–402. [Google Scholar] [CrossRef]
- Godfrey, D.I.; Kronenberg, M. Going Both Ways: Immune Regulation via CD1d-Dependent NKT Cells. J. Clin. Investig. 2004, 114, 1379–1388. [Google Scholar] [CrossRef]
- Eberl, G.; Brawand, P.; MacDonald, H.R. Selective Bystander Proliferation of Memory CD4+ and CD8+ T Cells Upon NK T or T Cell Activation. J. Immunol. 2000, 165, 4305–4311. [Google Scholar] [CrossRef]
- Roark, J.H.; Park, S.H.; Jayawardena, J.; Kavita, U.; Shannon, M.; Bendelac, A. CD1.1 Expression by Mouse Antigen-Presenting Cells and Marginal Zone B Cells. J. Immunol. 1998, 160, 3121–3127. [Google Scholar] [CrossRef] [PubMed]
- Exley, M.; Garcia, J.; Wilson, S.B.; Spada, F.; Gerdes, D.; Tahir, S.M.A.; Patton, K.T.; Blumberg, R.S.; Porcelli, S.; Chott, A.; et al. CD1d Structure and Regulation on Human Thymocytes, Peripheral Blood T Cells, B Cells and Monocytes. Immunology 2000, 100, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Brigl, M.; Brenner, M.B. CD1: Antigen Presentation and T Cell Function. Annu. Rev. Immunol. 2004, 22, 817–890. [Google Scholar] [CrossRef] [PubMed]
- Leadbetter, E.A.; Karlsson, M.C. Reading the Room: iNKT Cells Influence B Cell Responses. Mol. Immunol. 2020, 130, 49. [Google Scholar] [CrossRef]
- Campos, R.A.; Szczepanik, M.; Lisbonne, M.; Itakura, A.; Leite-de-Moraes, M.; Askenase, P.W. Invariant NKT Cells Rapidly Activated via Immunization with Diverse Contact Antigens Collaborate In Vitro with B-1 Cells to Initiate Contact Sensitivity1. J. Immunol. 2006, 177, 3686–3694. [Google Scholar] [CrossRef]
- Lobo, P.I.; Schlegal, K.H.; Vengal, J.; Okusa, M.D.; Pei, H. Naturally Occurring IgM Anti-Leukocyte Autoantibodies Inhibit T-Cell Activation and Chemotaxis. J. Clin. Immunol. 2010, 30, 31–36. [Google Scholar] [CrossRef]
- Lobo, P.I. Role of Natural Autoantibodies and Natural IgM Anti-Leucocyte Autoantibodies in Health and Disease. Front. Immunol. 2016, 7, 198. [Google Scholar] [CrossRef]
- Hayakawa, K.; Hardy, R.R.; Herzenberg, L.A.; Herzenberg, L.A. Progenitors for Ly-1 B Cells Are Distinct from Progenitors for Other B Cells. J. Exp. Med. 1985, 161, 1554–1568. [Google Scholar] [CrossRef]
- Hinks, T.S.C.; Zhang, X.-W. MAIT Cell Activation and Functions. Front. Immunol. 2020, 11, 1014. [Google Scholar] [CrossRef]
- Nel, I.; Bertrand, L.; Toubal, A.; Lehuen, A. MAIT Cells, Guardians of Skin and Mucosa? Mucosal Immunol. 2021, 14, 803–814. [Google Scholar] [CrossRef]
- Jeffery, H.C.; van Wilgenburg, B.; Kurioka, A.; Parekh, K.; Stirling, K.; Roberts, S.; Dutton, E.E.; Hunter, S.; Geh, D.; Braitch, M.K.; et al. Biliary Epithelium and Liver B Cells Exposed to Bacteria Activate Intrahepatic MAIT Cells through MR1. J. Hepatol. 2016, 64, 1118–1127. [Google Scholar] [CrossRef] [PubMed]
- Gold, M.C.; Cerri, S.; Smyk-Pearson, S.; Cansler, M.E.; Vogt, T.M.; Delepine, J.; Winata, E.; Swarbrick, G.M.; Chua, W.-J.; Yu, Y.Y.L.; et al. Human Mucosal Associated Invariant T Cells Detect Bacterially Infected Cells. PLoS Biol. 2010, 8, e1000407. [Google Scholar] [CrossRef] [PubMed]
- Kjer-Nielsen, L.; Patel, O.; Corbett, A.J.; Le Nours, J.; Meehan, B.; Liu, L.; Bhati, M.; Chen, Z.; Kostenko, L.; Reantragoon, R.; et al. MR1 Presents Microbial Vitamin B Metabolites to MAIT Cells. Nature 2012, 491, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Ko, E.-J.; Bhuyan, F.; Enyindah-Asonye, G.; Hunegnaw, R.; Helmold Hait, S.; Hogge, C.J.; Venzon, D.J.; Hoang, T.; Robert-Guroff, M. Mucosal-Associated Invariant T (MAIT) Cells Provide B-Cell Help in Vaccinated and Subsequently SIV-Infected Rhesus Macaques. Sci. Rep. 2020, 10, 10060. [Google Scholar] [CrossRef]
- Marzano, P.; Balin, S.; Terzoli, S.; Della Bella, S.; Cazzetta, V.; Piazza, R.; Sandrock, I.; Ravens, S.; Tan, L.; Prinz, I.; et al. Transcriptomic Profile of TNFhigh MAIT Cells Is Linked to B Cell Response Following SARS-CoV-2 Vaccination. Front. Immunol. 2023, 14, 1208662. [Google Scholar] [CrossRef]
- Chancellor, A.; Alan Simmons, R.; Khanolkar, R.C.; Nosi, V.; Beshirova, A.; Berloffa, G.; Colombo, R.; Karuppiah, V.; Pentier, J.M.; Tubb, V.; et al. Promiscuous Recognition of MR1 Drives Self-Reactive Mucosal-Associated Invariant T Cell Responses. J. Exp. Med. 2023, 220, e20221939. [Google Scholar] [CrossRef]
- Kaveri, S.V.; Silverman, G.J.; Bayry, J. Natural IgM in Immune Equilibrium and Harnessing Their Therapeutic Potential. J. Immunol. 2012, 188, 939–945. [Google Scholar] [CrossRef]
- Rothstein, T.L.; Quach, T.D. The Human Counterpart of Mouse B-1 Cells. Ann. N. Y. Acad. Sci. 2015, 1362, 143–152. [Google Scholar] [CrossRef]
- Lunderberg, J.M.; Dutta, S.; Collier, A.-R.Y.; Lee, J.-S.; Hsu, Y.-M.; Wang, Q.; Zheng, W.; Hao, S.; Zhang, H.; Feng, L.; et al. Pan-Neutralizing, Germline-Encoded Antibodies against SARS-CoV-2: Addressing the Long-Term Problem of Escape Variants. Front. Immunol. 2022, 13, 1032574. [Google Scholar] [CrossRef]
- Baumgarth, N. The Double Life of a B-1 Cell: Self-Reactivity Selects for Protective Effector Functions. Nat. Rev. Immunol. 2011, 11, 34–46. [Google Scholar] [CrossRef]
- Rothstein, T.L.; Griffin, D.O.; Holodick, N.E.; Quach, T.D.; Kaku, H. Human B-1 Cells Take the Stage. Ann. N. Y. Acad. Sci. 2013, 1285, 97–114. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Li, X.; Wu, J.; Zhang, S.; Zhu, L.; Chen, Q.; Fan, Y.; Wu, Z.; Xie, S.; Chen, Q.; et al. Pre-Existing Humoral Immunity to Low Pathogenic Human Coronaviruses Exhibits Limited Cross-Reactive Antibodies Response against SARS-CoV-2 in Children. Front. Immunol. 2022, 13, 1042406. [Google Scholar] [CrossRef] [PubMed]
- Stall, A.M.; Adams, S.; Herzenberg, L.A.; Kantor, A.B. Characteristics and Development of the Murine B-Lb (Ly-1 B Sister) Cell Population. Ann. N. Y. Acad. Sci. 1992, 651, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, S.M.; Perry, H.M.; Gonen, A.; Prohaska, T.A.; Srikakulapu, P.; Grewal, S.; Das, D.; McSkimming, C.; Taylor, A.M.; Tsimikas, S.; et al. B-1b Cells Secrete Atheroprotective IgM and Attenuate Atherosclerosis. Circ. Res. 2015, 117, e28–e39. [Google Scholar] [CrossRef]
- Srikakulapu, P.; Pattarabanjird, T.; Upadhye, A.; Bontha, S.V.; Osinski, V.; Marshall, M.A.; Garmey, J.; Deroissart, J.; Prohaska, T.A.; Witztum, J.L.; et al. B-1b Cells Have Unique Functional Traits Compared to B-1a Cells at Homeostasis and in Aged Hyperlipidemic Mice With Atherosclerosis. Front. Immunol. 2022, 13, 909475. [Google Scholar] [CrossRef]
- Vergani, S.; Muleta, K.G.; Da Silva, C.; Doyle, A.; Kristiansen, T.A.; Sodini, S.; Krausse, N.; Montano, G.; Kotarsky, K.; Nakawesi, J.; et al. A Self-Sustaining Layer of Early-Life-Origin B Cells Drives Steady-State IgA Responses in the Adult Gut. Immunity 2022, 55, 1829–1842.e6. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, C.; Yang, Q.; Kantor, A.B.; Chu, H.; Ghosn, E.E.; Qin, G.; Mazmanian, S.K.; Han, J.; Herzenberg, L.A. Distinct Mechanisms Define Murine B Cell Lineage Immunoglobulin Heavy Chain (IgH) Repertoires. eLife 2015, 4, e09083. [Google Scholar] [CrossRef]
- Cunningham, A.F.; Flores-Langarica, A.; Bobat, S.; Medina, C.C.D.; Cook, C.N.L.; Ross, E.A.; Lopez-Macias, C.; Henderson, I.R. B1b Cells Recognize Protective Antigens after Natural Infection and Vaccination. Front. Immunol. 2014, 5, 535. [Google Scholar] [CrossRef]
- Serruto, D.; Bottomley, M.J.; Ram, S.; Giuliani, M.M.; Rappuoli, R. The New Multicomponent Vaccine against Meningococcal Serogroup B, 4CMenB: Immunological, Functional and Structural Characterization of the Antigens. Vaccine 2012, 30, B87–B97. [Google Scholar] [CrossRef]
- Barbour, A.G. Antigenic variation of a relapsing fever borrelia species. Annu. Rev. Microbiol. 1990, 44, 155–171. [Google Scholar] [CrossRef]
- Foote, J.B.; Kearney, J.F. Generation of B Cell Memory to the Bacterial Polysaccharide α-1,3 Dextran1. J. Immunol. 2009, 183, 6359–6368. [Google Scholar] [CrossRef] [PubMed]
- Alugupalli, K.R.; Gerstein, R.M.; Chen, J.; Szomolanyi-Tsuda, E.; Woodland, R.T.; Leong, J.M. The Resolution of Relapsing Fever Borreliosis Requires IgM and Is Concurrent with Expansion of B1b Lymphocytes1. J. Immunol. 2003, 170, 3819–3827. [Google Scholar] [CrossRef] [PubMed]
- Alugupalli, K.R. A Distinct Role for B1b Lymphocytes in T Cell-Independent Immunity. In Specialization and Complementation of Humoral Immune Responses to Infection; Manser, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 105–130. ISBN 978-3-540-73900-5. [Google Scholar]
- Alugupalli, K.R.; Leong, J.M.; Woodland, R.T.; Muramatsu, M.; Honjo, T.; Gerstein, R.M. B1b Lymphocytes Confer T Cell-Independent Long-Lasting Immunity. Immunity 2004, 21, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Liu, H.; Wu, N.C.; Lee, C.-C.D.; Zhu, X.; Zhao, F.; Huang, D.; Yu, W.; Hua, Y.; Tien, H.; et al. Structural Basis of a Shared Antibody Response to SARS-CoV-2. Science 2020, 369, 1119–1123. [Google Scholar] [CrossRef]
- Shi, R.; Shan, C.; Duan, X.; Chen, Z.; Liu, P.; Song, J.; Song, T.; Bi, X.; Han, C.; Wu, L.; et al. A Human Neutralizing Antibody Targets the Receptor-Binding Site of SARS-CoV-2. Nature 2020, 584, 120–124. [Google Scholar] [CrossRef]
- Seydoux, E.; Homad, L.J.; MacCamy, A.J.; Parks, K.R.; Hurlburt, N.K.; Jennewein, M.F.; Akins, N.R.; Stuart, A.B.; Wan, Y.-H.; Feng, J.; et al. Analysis of a SARS-CoV-2-Infected Individual Reveals Development of Potent Neutralizing Antibodies with Limited Somatic Mutation. Immunity 2020, 53, 98–105.e5. [Google Scholar] [CrossRef]
- Robbiani, D.F.; Gaebler, C.; Muecksch, F.; Lorenzi, J.C.C.; Wang, Z.; Cho, A.; Agudelo, M.; Barnes, C.O.; Gazumyan, A.; Finkin, S.; et al. Convergent Antibody Responses to SARS-CoV-2 in Convalescent Individuals. Nature 2020, 584, 437–442. [Google Scholar] [CrossRef]
- Brouwer, P.J.M.; Caniels, T.G.; van der Straten, K.; Snitselaar, J.L.; Aldon, Y.; Bangaru, S.; Torres, J.L.; Okba, N.M.A.; Claireaux, M.; Kerster, G.; et al. Potent Neutralizing Antibodies from COVID-19 Patients Define Multiple Targets of Vulnerability. Science 2020, 369, 643–650. [Google Scholar] [CrossRef]
- Kreer, C.; Zehner, M.; Weber, T.; Ercanoglu, M.S.; Gieselmann, L.; Rohde, C.; Halwe, S.; Korenkov, M.; Schommers, P.; Vanshylla, K.; et al. Longitudinal Isolation of Potent Near-Germline SARS-CoV-2-Neutralizing Antibodies from COVID-19 Patients. Cell 2020, 182, 843–854.e12. [Google Scholar] [CrossRef]
- Vuyyuru, R.; Liu, H.; Manser, T.; Alugupalli, K.R. Characteristics of Borrelia Hermsii Infection in Human Hematopoietic Stem Cell-Engrafted Mice Mirror Those of Human Relapsing Fever. Proc. Natl. Acad. Sci. USA 2011, 108, 20707–20712. [Google Scholar] [CrossRef]
- Cohen, I.R. Real and Artificial Immune Systems: Computing the State of the Body. Nat. Rev. Immunol. 2007, 7, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.O.; Holodick, N.E.; Rothstein, T.L. Human B1 Cells in Umbilical Cord and Adult Peripheral Blood Express the Novel Phenotype CD20+CD27+CD43+CD70−. J. Exp. Med. 2011, 208, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Holodick, N.E.; Tumang, J.R.; Rothstein, T.L. Immunoglobulin Secretion by B1 Cells: Differential Intensity and IRF4-Dependence of Spontaneous IgM Secretion by Peritoneal and Splenic B1 Cells. Eur. J. Immunol. 2010, 40, 3007–3016. [Google Scholar] [CrossRef] [PubMed]
- Graf, R.; Seagal, J.; Otipoby, K.L.; Lam, K.-P.; Ayoub, S.; Zhang, B.; Sander, S.; Chu, V.T.; Rajewsky, K. BCR-Dependent Lineage Plasticity in Mature B Cells. Science 2019, 363, 748–753. [Google Scholar] [CrossRef]
- Khasbiullina, N.R.; Bovin, N.V. Hypotheses of the Origin of Natural Antibodies: A Glycobiologist’s Opinion. Biochem. Mosc. 2015, 80, 820–835. [Google Scholar] [CrossRef]
- HAMPE, C.S. Protective Role of Anti-Idiotypic Antibodies in Autoimmunity—Lessons for Type 1 Diabetes. Autoimmunity 2012, 45, 320–331. [Google Scholar] [CrossRef]
- Matejuk, A.; Beardall, M.; Xu, Y.; Tian, Q.; Phillips, D.; Alabyev, B.; Mannoor, K.; Chen, C. Exclusion of Natural Autoantibody-Producing B Cells from IgG Memory B Cell Compartment during T Cell-Dependent Immune Responses1. J. Immunol. 2009, 182, 7634–7643. [Google Scholar] [CrossRef]
- Baumgarth, N. B-1 Cell Heterogeneity and the Regulation of Natural and Antigen-Induced IgM Production. Front. Immunol. 2016, 7, 324. [Google Scholar] [CrossRef]
- Elkon, K.; Casali, P. Nature and Functions of Autoantibodies. Nat. Clin. Pract. Rheumatol. 2008, 4, 491–498. [Google Scholar] [CrossRef]
- Avrameas, S. Natural Autoantibodies: From ‘Horror Autotoxicus’ to ‘Gnothi Seauton’. Immunol. Today 1991, 12, 154–159. [Google Scholar] [CrossRef]
- Rauch, P.J.; Chudnovskiy, A.; Robbins, C.S.; Weber, G.F.; Etzrodt, M.; Hilgendorf, I.; Tiglao, E.; Figueiredo, J.-L.; Iwamoto, Y.; Theurl, I.; et al. Innate Response Activator B Cells Protect Against Microbial Sepsis. Science 2012, 335, 597–601. [Google Scholar] [CrossRef]
- Grönwall, C.; Vas, J.; Silverman, G. Protective Roles of Natural IgM Antibodies. Front. Immunol. 2012, 3, 66. [Google Scholar] [CrossRef]
- Gorevic, P.D. Rheumatoid Factor, Complement, and Mixed Cryoglobulinemia. J. Immunol. Res. 2012, 2012, 439018. [Google Scholar] [CrossRef]
- Charles, E.D.; Dustin, L.B. Hepatitis C Virus–Induced Cryoglobulinemia. Kidney Int. 2009, 76, 818–824. [Google Scholar] [CrossRef]
- Newkirk, M.M. Rheumatoid Factors: What Do They Tell Us? J. Rheumatol. 2002, 29, 2034–2040. [Google Scholar]
- Lobo, P.I.; Schlegel, K.H.; Bajwa, A.; Huang, L.; Kurmaeva, E.; Wang, B.; Ye, H.; Tedder, T.F.; Kinsey, G.R.; Okusa, M.D. Natural IgM Switches the Function of Lipopolysaccharide-Activated Murine Bone Marrow–Derived Dendritic Cells to a Regulatory Dendritic Cell That Suppresses Innate Inflammation. J. Immunol. 2015, 195, 5215–5226. [Google Scholar] [CrossRef]
- Lobo, P.I.; Bajwa, A.; Schlegel, K.H.; Vengal, J.; Lee, S.J.; Huang, L.; Ye, H.; Deshmukh, U.; Wang, T.; Pei, H.; et al. Natural IgM Anti-Leukocyte Autoantibodies Attenuate Excess Inflammation Mediated by Innate and Adaptive Immune Mechanisms Involving Th-17. J. Immunol. 2012, 188, 1675–1685. [Google Scholar] [CrossRef]
- Adib, M.; Ragimbeau, J.; Avrameas, S.; Ternynck, T. IgG Autoantibody Activity in Normal Mouse Serum Is Controlled by IgM. J. Immunol. 1990, 145, 3807–3813. [Google Scholar] [CrossRef]
- Lobo, P.I.; Sturgill, B.C.; Bolton, W.K. Cold-reactive alloantibodies and allograft malfunction occurring immediately posttransplant. Transplantation 1984, 37, 76. [Google Scholar] [CrossRef]
- Zhang, M.; Alicot, E.M.; Carroll, M.C. Human Natural IgM Can Induce Ischemia/Reperfusion Injury in a Murine Intestinal Model. Mol. Immunol. 2008, 45, 4036–4039. [Google Scholar] [CrossRef]
- Sidman, C.L.; Shultz, L.D.; Hardy, R.R.; Hayakawa, K.; Herzenberg, L.A. Production of Immunoglobulin Isotypes by Ly-1+ B Cells in Viable Motheaten and Normal Mice. Science 1986, 232, 1423–1425. [Google Scholar] [CrossRef]
- Solvason, N.; Lehuen, A.; Kearney, J.F. An Embryonic Source of Ly1 but Not Conventional B Cells. Int. Immunol. 1991, 3, 543–550. [Google Scholar] [CrossRef]
- Berneman, A.; Ternynck, T.; Avrameas, S. Natural Mouse IgG Reacts with Self Antigens Including Molecules Involved in the Immune Response. Eur. J. Immunol. 1992, 22, 625–633. [Google Scholar] [CrossRef]
- Hurez, V.; Kaveri, S.-V.; Kazatchkine, M.D. Expression and Control of the Natural Autoreactive IgG Repertoire in Normal Human Serum. Eur. J. Immunol. 1993, 23, 783–789. [Google Scholar] [CrossRef]
- Pereira, P.; Forni, L.; Larsson, E.-L.-L.; Cooper, M.; Heusser, C.; Coutinho, A. Autonomous Activation of B and T Cells in Antigen-Free Mice. Eur. J. Immunol. 1986, 16, 685–688. [Google Scholar] [CrossRef]
- Watts, R.A.; Isenberg, D.A. Autoantibodies and Antibacterial Antibodies: From Both Sides Now. Ann. Rheum. Dis. 1990, 49, 961–965. [Google Scholar] [CrossRef]
- Hamanova, M.; Chmelikova, M.; Nentwich, I.; Thon, V.; Lokaj, J. Anti-Gal IgM, IgA and IgG Natural Antibodies in Childhood. Immunol. Lett. 2015, 164, 40–43. [Google Scholar] [CrossRef]
- Bos, N.A.; Kimura, H.; Meeuwsen, C.G.; Visser, H.D.; Hazenberg, M.P.; Wostmann, B.S.; Pleasants, J.R.; Benner, R.; Marcus, D.M. Serum Immunoglobulin Levels and Naturally Occurring Antibodies against Carbohydrate Antigens in Germ-Free BALB/c Mice Fed Chemically Defined Ultrafiltered Diet. Eur. J. Immunol. 1989, 19, 2335–2339. [Google Scholar] [CrossRef]
- Nagele, E.P.; Han, M.; Acharya, N.K.; DeMarshall, C.; Kosciuk, M.C.; Nagele, R.G. Natural IgG Autoantibodies Are Abundant and Ubiquitous in Human Sera, and Their Number Is Influenced By Age, Gender, and Disease. PLoS ONE 2013, 8, e60726. [Google Scholar] [CrossRef]
- Beinart, D.; Ren, D.; Pi, C.; Poulton, S.; Holzknecht, Z.E.; Swanson, C.; Parker, W. Immunization Enhances the Natural Antibody Repertoire. EXCLI J. 2017, 16, 1018–1030. [Google Scholar] [CrossRef]
- Lacroix-Desmazes, S.; Mouthon, L.; Coutinho, A.; Kazatchkine, M.D. Analysis of the Natural Human IgG Antibody Repertoire: Life-Long Stability of Reactivities towards Self Antigens Contrasts with Age-Dependent Diversification of Reactivities against Bacterial Antigens. Eur. J. Immunol. 1995, 25, 2598–2604. [Google Scholar] [CrossRef]
- Palma, J.; Tokarz-Deptuła, B.; Deptuła, J.; Deptuła, W. Natural Antibodies—Facts Known and Unknown. Cent. Eur. J. Immunol. 2018, 43, 466–475. [Google Scholar] [CrossRef]
- Amital, H.; Shoenfeld, Y. Natural autoantibodies, heralding, protecting and inducing autoimmunity. In Autoantibodies; Elsevier Science: Amsterdam, The Netherlands, 2007; pp. 7–12. [Google Scholar] [CrossRef]
- Zorn, E.; See, S.B. Polyreactive Natural Antibodies in Transplantation. Curr. Opin. Organ Transplant. 2017, 22, 8–13. [Google Scholar] [CrossRef]
- Cohen, I.R. Autoantibody Repertoires, Natural Biomarkers, and System Controllers. Trends Immunol. 2013, 34, 620–625. [Google Scholar] [CrossRef]
- Czömpöly, T.; Olasz, K.; Simon, D.; Nyárády, Z.; Pálinkás, L.; Czirják, L.; Berki, T.; Németh, P. A Possible New Bridge between Innate and Adaptive Immunity: Are the Anti-Mitochondrial Citrate Synthase Autoantibodies Components of the Natural Antibody Network? Mol. Immunol. 2006, 43, 1761–1768. [Google Scholar] [CrossRef]
- Avrameas, S.; Alexopoulos, H.; Moutsopoulos, H.M. Natural Autoantibodies: An Undersugn Hero of the Immune System and Autoimmune Disorders—A Point of View. Front. Immunol. 2018, 9, 1320. [Google Scholar] [CrossRef]
- Tillman, D.M.; Jou, N.T.; Hill, R.J.; Marion, T.N. Both IgM and IgG Anti-DNA Antibodies Are the Products of Clonally Selective B Cell Stimulation in (NZB x NZW)F1 Mice. J. Exp. Med. 1992, 176, 761–779. [Google Scholar] [CrossRef]
- Ikematsu, H.; Kasaian, M.T.; Schettino, E.W.; Casali, P. Structural Analysis of the VH-D-JH Segments of Human Polyreactive IgG mAb. Evidence for Somatic Selection. J. Immunol. 1993, 151, 3604–3616. [Google Scholar] [CrossRef]
- Kasaian, M.T.; Ikematsu, H.; Balow, J.E.; Casali, P. Structure of the VH and VL Segments of Mono Reactive and Polyreactive IgA Autoantibodies to DNA in Patients with Systemic Lupus Erythematosus. J. Immunol. 1994, 152, 3137–3151. [Google Scholar] [CrossRef]
- Cunningham, M.W.; Hall, N.K.; Krisher, K.K.; Spanier, A.M. A Study of Anti-Group A Streptococcal Monoclonal Antibodies Cross-Reactive with Myosin. J. Immunol. 1986, 136, 293–298. [Google Scholar] [CrossRef]
- Danieli, M.G.; Antonelli, E.; Gammeri, L.; Longhi, E.; Cozzi, M.F.; Palmeri, D.; Gangemi, S.; Shoenfeld, Y. Intravenous Immunoglobulin as a Therapy for Autoimmune Conditions. Autoimmun. Rev. 2025, 24, 103710. [Google Scholar] [CrossRef]
- Schwab, I.; Nimmerjahn, F. Intravenous Immunoglobulin Therapy: How Does IgG Modulate the Immune System? Nat. Rev. Immunol. 2013, 13, 176–189. [Google Scholar] [CrossRef]
- Ephrem, A.; Chamat, S.; Miquel, C.; Fisson, S.; Mouthon, L.; Caligiuri, G.; Delignat, S.; Elluru, S.; Bayry, J.; Lacroix-Desmazes, S.; et al. Expansion of CD4+CD25+ Regulatory T Cells by Intravenous Immunoglobulin: A Critical Factor in Controlling Experimental Autoimmune Encephalomyelitis. Blood 2008, 111, 715–722. [Google Scholar] [CrossRef]
- Besredka, A. Les Antihémolysines Naturelles. Ann. De l’Institut Pasteur 1901, 15, 785–807. [Google Scholar]
- Landsteiner, K. The Nature and Specificity of Antibodies. In The Specificity of Serological Reactions; Harvard University Press: Boston, MA, USA, 1945; pp. 127–155. [Google Scholar]
- Burnet, F.M., Sir. The Clonal Selection Theory of Acquired Immunity; Cambridge University Press: Cambridge, UK, 1959. [Google Scholar]
- Jerne, N.K. Towards a Network Theory of the Immune System. Ann. Immunol. 1974, 125C, 373–389. [Google Scholar]
- Stewart, J. Immunoglobulins Did Not Arise in Evolution to Fight Infection. Immunol. Today 1992, 13, 396–399. [Google Scholar] [CrossRef]
- Avrameas, S.; Guilbert, B.; Dighiero, G. Natural Antibodies against Tubulin, Actin, Myoglobin, Thyroglobulin, Fetuin, Albumin and Transferrin Are Present in Normal Human Sera and Monoclonal Immunoglobulins from Multiple Myeloma and Waldenström’s Macroglobulinemia May Express Similar Antibody Specificities. Ann. Inst. Pasteur Immunol. 1981, 132, 231–236. [Google Scholar] [CrossRef]
- Hahn, B.H. Characteristics of Pathogenic Subpopulations of Antibodies to Dna. Arthritis Rheum. 1982, 25, 747–752. [Google Scholar] [CrossRef]
- Isenberg, D.A.; Madaio, M.P.; Reichlin, M.; Shoenfeld, Y.; Rauch, J.; Stollar, B.D.; Schwartz, R.S. Anti-Dna Antibody Idiotypes In Systemic Lupus Erythematosus. The Lancet. 1984, 324, 417–422. [Google Scholar] [CrossRef]
- Bakimer, R.; Fishman, P.; Blank, M.; Sredni, B.; Djaldetti, M.; Shoenfeld, Y. Induction of Primary Antiphospholipid Syndrome in Mice by Immunization with a Human Monoclonal Anticardiolipin Antibody (H-3). J. Clin. Invest. 1992, 89, 1558–1563. [Google Scholar] [CrossRef]
- Mendlovic, S.; Brocke, S.; Shoenfeld, Y.; Ben-Bassat, M.; Meshorer, A.; Bakimer, R.; Mozes, E. Induction of a Systemic Lupus Erythematosus-like Disease in Mice by a Common Human Anti-DNA Idiotype. Proc. Natl. Acad. Sci. USA 1988, 85, 2260–2264. [Google Scholar] [CrossRef] [PubMed]
- Reyneveld, G.I.; Savelkoul, H.F.J.; Parmentier, H.K. Current Understanding of Natural Antibodies and Exploring the Possibilities of Modulation Using Veterinary Models. A Review. Front. Immunol. 2020, 11, 2139. [Google Scholar] [CrossRef] [PubMed]
- Coutinho, A.; Kazatchkine, M.D.; Avrameas, S. Natural Autoantibodies. Curr. Opin. Immunol. 1995, 7, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Ochsenbein, A.F.; Fehr, T.; Lutz, C.; Suter, M.; Brombacher, F.; Hengartner, H.; Zinkernagel, R.M. Control of Early Viral and Bacterial Distribution and Disease by Natural Antibodies. Science 1999, 286, 2156–2159. [Google Scholar] [CrossRef]
- Quintana, F.J.; Cohen, I.R. The Natural Autoantibody Repertoire and Autoimmune Disease. Biomed. Pharmacother. 2004, 58, 276–281. [Google Scholar] [CrossRef]
- Németh, P.; Simon, D. Natural and Pathologic Autoantibodies. In Insights and Perspectives in Rheumatology; IntechOpen: Rijeka, Croatia, 2012; ISBN 978-953-307-846-5. [Google Scholar]
- Devalapalli, A.P.; Lesher, A.; Shieh, K.; Solow, J.S.; Everett, M.L.; Edala, A.S.; Whitt, P.; Long, R.R.; Newton, N.; Parker, W. Increased Levels of IgE and Autoreactive, Polyreactive IgG in Wild Rodents: Implications for the Hygiene Hypothesis. Scand. J. Immunol. 2006, 64, 125–136. [Google Scholar] [CrossRef]
- Perotti, M.; Ghidoli, N.; Altara, R.; Diotti, R.A.; Clementi, N.; De Marco, D.; Sassi, M.; Clementi, M.; Burioni, R.; Mancini, N. Hepatitis C Virus (HCV)-Driven Stimulation of Subfamily-Restricted Natural IgM Antibodies in Mixed Cryoglobulinemia. Autoimmun. Rev. 2008, 7, 468–472. [Google Scholar] [CrossRef]
- Varbiro, S.; Biro, A.; Cervenak, J.; Cervenak, L.; Singh, M.; Banhidy, F.; Sebestyen, A.; Füst, G.; Prohászka, Z. Human Anti-60 kD Heat Shock Protein Autoantibodies Are Characterized by Basic Features of Natural Autoantibodies. Acta Physiol. Hung. 2010, 97, 1–10. [Google Scholar] [CrossRef]
- Wu, J.; Li, L. Autoantibodies in Alzheimer’s Disease: Potential Biomarkers, Pathogenic Roles, and Therapeutic Implications. J. Biomed. Res. 2016, 30, 361–372. [Google Scholar] [CrossRef]
- Miteva, D.; Vasilev, G.V.; Velikova, T. Role of Specific Autoantibodies in Neurodegenerative Diseases: Pathogenic Antibodies or Promising Biomarkers for Diagnosis. Antibodies 2023, 12, 81. [Google Scholar] [CrossRef]
- Shim, S.-M.; Koh, Y.H.; Kim, J.-H.; Jeon, J.-P. A Combination of Multiple Autoantibodies Is Associated with the Risk of Alzheimer’s Disease and Cognitive Impairment. Sci. Rep. 2022, 12, 1312. [Google Scholar] [CrossRef] [PubMed]
- Ehtewish, H.; Mesleh, A.; Ponirakis, G.; Lennard, K.; Al Hamad, H.; Chandran, M.; Parray, A.; Abdesselem, H.; Wijten, P.; Decock, J.; et al. Profiling the Autoantibody Repertoire Reveals Autoantibodies Associated with Mild Cognitive Impairment and Dementia. Front. Neurol. 2023, 14, 1256745. [Google Scholar] [CrossRef]
- DeMarshall, C.; Goldwaser, E.L.; Sarkar, A.; Godsey, G.A.; Acharya, N.K.; Thayasivam, U.; Belinka, B.A.; Nagele, R.G. Autoantibodies as Diagnostic Biomarkers for the Detection and Subtyping of Multiple Sclerosis. J. Neuroimmunol. 2017, 309, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.-H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Béziat, V.; et al. Autoantibodies against Type I IFNs in Patients with Life-Threatening COVID-19. Science 2020, 370, eabd4585. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.; Kashyap, A.; Salvator, H.; Rosen, L.B.; Colby, D.; Ardeshir-Larijani, F.; Loehrer, P.J.; Ding, L.; Reyes, S.O.L.; Riminton, S.; et al. Anti–Interleukin-23 Autoantibodies in Adult-Onset Immunodeficiency. N. Engl. J. Med. 2024, 390, 1105–1117. [Google Scholar] [CrossRef]
- Batista, C.M.; Foti, L. Anti-SARS-CoV-2 and Anti-Cytokine Storm Neutralizing Antibody Therapies against COVID-19: Update, Challenges, and Perspectives. Int. Immunopharmacol. 2021, 99, 108036. [Google Scholar] [CrossRef]
- Graudal, N.A.; Svenson, M.; Tarp, U.; Garred, P.; Jurik, A.-G.; Bendtzen, K. Autoantibodies against Interleukin 1α in Rheumatoid Arthritis: Association with Long Term Radiographic Outcome. Ann. Rheum. Dis. 2002, 61, 598–602. [Google Scholar] [CrossRef]
- Wildbaum, G.; Nahir, M.A.; Karin, N. Beneficial Autoimmunity to Proinflammatory Mediators Restrains the Consequences of Self-Destructive Immunity. Immunity 2003, 19, 679–688. [Google Scholar] [CrossRef]
- von Stemann, J.H.; Rigas, A.S.; Thørner, L.W.; Rasmussen, D.G.K.; Pedersen, O.B.; Rostgaard, K.; Erikstrup, C.; Ullum, H.; Hansen, M.B. Prevalence and Correlation of Cytokine-Specific Autoantibodies with Epidemiological Factors and C-Reactive Protein in 8972 Healthy Individuals: Results from the Danish Blood Donor Study. PLoS ONE 2017, 12, e0179981. [Google Scholar] [CrossRef]
- von Stemann, J.H.; Dubois, F.; Saint-André, V.; Bondet, V.; Posseme, C.; Charbit, B.; Quintana-Murci, L.; Hansen, M.B.; Ostrowski, S.R.; Duffy, D.; et al. Cytokine Autoantibodies Alter Gene Expression Profiles of Healthy Donors. Eur. J. Immunol. 2025, 55, e202451211. [Google Scholar] [CrossRef]
- Gupta, S.; Tatouli, I.P.; Rosen, L.B.; Hasni, S.; Alevizos, I.; Manna, Z.G.; Rivera, J.; Jiang, C.; Siegel, R.M.; Holland, S.M.; et al. Distinct Functions of Autoantibodies Against Interferon in Systemic Lupus Erythematosus: A Comprehensive Analysis of Anticytokine Autoantibodies in Common Rheumatic Diseases. Arthritis Rheumatol. 2016, 68, 1677–1687. [Google Scholar] [CrossRef] [PubMed]
- Schrader, J.W.; Goding, J.W. Chapter 76—Autoantibodies Against Cytokines. In The Autoimmune Diseases, 5th ed.; Rose, N.R., Mackay, I.R., Eds.; Academic Press: Boston, MA, USA, 2014; pp. 1141–1158. ISBN 978-0-12-384929-8. [Google Scholar]
- Howe, H.S.; Leung, B.P.L. Anti-Cytokine Autoantibodies in Systemic Lupus Erythematosus. Cells 2020, 9, 72. [Google Scholar] [CrossRef] [PubMed]
- Panem, S.; Check, I.J.; Henriksen, D.; Vilcek, J. Antibodies to Alpha-Interferon in a Patient with Systemic Lupus Erythematosus. J. Immunol. 1982, 129, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Fosgerau, K.; Galle, P.; Hansen, T.; Albrechtsen, A.; Rieper, C.d.L.; Pedersen, B.K.; Larsen, L.K.; Thomsen, A.R.; Pedersen, O.; Hansen, M.B.; et al. Interleukin-6 Autoantibodies Are Involved in the Pathogenesis of a Subset of Type 2 Diabetes. J. Endocrinol. 2010, 204, 265–273. [Google Scholar] [CrossRef]
- Uthayakumar, D.; Paris, S.; Chapat, L.; Freyburger, L.; Poulet, H.; Luca, K.D. Non-Specific Effects of Vaccines Illustrated Through the BCG Example: From Observations to Demonstrations. Front. Immunol. 2018, 9, 2869. [Google Scholar] [CrossRef]
- Benn, C.S.; Amenyogbe, N.; Björkman, A.; Domínguez-Andrés, J.; Fish, E.N.; Flanagan, K.L.; Klein, S.L.; Kollmann, T.R.; Kyvik, K.O.; Netea, M.G.; et al. Implications of Non-Specific Effects for Testing, Approving, and Regulating Vaccines. Drug Saf. 2023, 46, 439–448. [Google Scholar] [CrossRef]
- Jeannet, R.; Descazeaud, A.; Daix, T.; Pauthier, H.; Pascal, V.; Hantz, S.; Cam, S.L.; Francois, B.; Feuillard, J.; Lafarge, X. De Novo Natural Anti-M Alloantibody Emergence in Severe Coronavirus Disease 2019. J. Infect. Public Health 2022, 15, 1455–1458. [Google Scholar] [CrossRef]
- Böröcz, K.; Kinyó, Á.; Simon, D.; Erdő-Bonyár, S.; Németh, P.; Berki, T. Complexity of the Immune Response Elicited by Different COVID-19 Vaccines, in the Light of Natural Autoantibodies and Immunomodulatory Therapies. Int. J. Mol. Sci. 2023, 24, 6439. [Google Scholar] [CrossRef]
- Szinger, D.; Berki, T.; Németh, P.; Erdo-Bonyar, S.; Simon, D.; Drenjančević, I.; Samardzic, S.; Zelić, M.; Sikora, M.; Požgain, A.; et al. Following Natural Autoantibodies: Further Immunoserological Evidence Regarding Their Silent Plasticity and Engagement in Immune Activation. Int. J. Mol. Sci. 2023, 24, 14961. [Google Scholar] [CrossRef]
- Czömpöly, T.; Simon, D.; Czirják, L.; Németh, P. Anti-Topoisomerase I Autoantibodies in Systemic Sclerosis. Autoimmun. Rev. 2009, 8, 692–696. [Google Scholar] [CrossRef]
- Böröcz, K.; Simon, D.; Erdő-Bonyár, S.; Kovács, K.T.; Tuba; Czirják, L.; Németh, P.; Berki, T. Relationship between Natural and Infection-Induced Antibodies in Systemic Autoimmune Diseases (SAD): SLE, SSc and RA. Clin. Exp. Immunol. 2021, 203, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.; Balogh, P.; Erdő-Bonyár, S.; Böröcz, K.; Minier, T.; Czirják, L.; Berki, T. Increased Frequency of Activated Switched Memory B Cells and Its Association With the Presence of Pulmonary Fibrosis in Diffuse Cutaneous Systemic Sclerosis Patients. Front. Immunol. 2021, 12, 686483. [Google Scholar] [CrossRef] [PubMed]
- New, J.S.; Dizon, B.L.P.; King, R.G.; Greenspan, N.S.; Kearney, J.F. B-1 B Cell–Derived Natural Antibodies against N-Acetyl-d-Glucosamine Suppress Autoimmune Diabetes Pathogenesis. J. Immunol. 2023, 211, 1320–1331. [Google Scholar] [CrossRef] [PubMed]
- Erdő-Bonyár, S.; Rapp, J.; Subicz, R.; Böröcz, K.; Szinger, D.; Filipánits, K.; Minier, T.; Kumánovics, G.; Czirják, L.; Berki, T.; et al. Disturbed Complement Receptor Expression Pattern of B Cells Is Enhanced by Toll-like Receptor CD180 Ligation in Diffuse Cutaneous Systemic Sclerosis. Int. J. Mol. Sci. 2024, 25, 9230. [Google Scholar] [CrossRef]
- Simon, D.; Erdő-Bonyár, S.; Böröcz, K.; Balázs, N.; Badawy, A.; Bajnok, A.; Nörenberg, J.; Serény-Litvai, T.; Várnagy, Á.; Kovács, K.; et al. Altered Levels of Natural Autoantibodies against Heat Shock Proteins in Pregnant Women with Hashimoto’s Thyroiditis. Int. J. Mol. Sci. 2024, 25, 1423. [Google Scholar] [CrossRef]
- Boonstra, M.; Bakker, J.A.; Grummels, A.; Ninaber, M.K.; Ajmone Marsan, N.; Wortel, C.M.; Huizinga, T.W.J.; Jordan, S.; Hoffman-Vold, A.-M.; Distler, O.; et al. Association of Anti–Topoisomerase I Antibodies of the IgM Isotype With Disease Progression in Anti–Topoisomerase I–Positive Systemic Sclerosis. Arthritis Rheumatol. 2020, 72, 1897–1904. [Google Scholar] [CrossRef]
- Simon, D.; Kacsándi, D.; Pusztai, A.; Soós, B.; Végh, E.; Kerekes, G.; Bodoki, M.; Szamosi, S.; Szűcs, G.; Prohászka, Z.; et al. Natural Autoantibodies in Biologic-Treated Rheumatoid Arthritis and Ankylosing Spondylitis Patients: Associations with Vascular Pathophysiology. Int. J. Mol. Sci. 2024, 25, 3429. [Google Scholar] [CrossRef]
- Holmes, D. Natural Autoantibodies Protect against T1DM. Nat. Rev. Endocrinol. 2016, 12, 560. [Google Scholar] [CrossRef]
- Nobrega, A.; Stransky, B.; Nicolas, N.; Coutinho, A. Regeneration of Natural Antibody Repertoire After Massive Ablation of Lymphoid System: Robust Selection Mechanisms Preserve Antigen Binding Specificities1. J. Immunol. 2002, 169, 2971–2978. [Google Scholar] [CrossRef]
- Neiman, M.; Hellström, C.; Just, D.; Mattsson, C.; Fagerberg, L.; Schuppe-Koistinen, I.; Gummesson, A.; Bergström, G.; Achour, A.; Sallinen, R.; et al. Individual and Stable Autoantibody Repertoires in Healthy Individuals. Autoimmunity 2019, 52, 1–11. [Google Scholar] [CrossRef]
- Kay, J.; Upchurch, K.S. ACR/EULAR 2010 Rheumatoid Arthritis Classification Criteria. Rheumatology 2012, 51, vi5–vi9. [Google Scholar] [CrossRef] [PubMed]
- Reijm, S.; Kissel, T.; Stoeken-Rijsbergen, G.; Slot, L.M.; Wortel, C.M.; van Dooren, H.J.; Levarht, N.E.W.; Kampstra, A.S.B.; Derksen, V.F.A.M.; Heer, P.O.; et al. Cross-Reactivity of IgM Anti-Modified Protein Antibodies in Rheumatoid Arthritis despite Limited Mutational Load. Arthritis Res. Ther. 2021, 23, 230. [Google Scholar] [CrossRef] [PubMed]
- Barbhaiya, M.; Zuily, S.; Naden, R.; Hendry, A.; Manneville, F.; Amigo, M.-C.; Amoura, Z.; Andrade, D.; Andreoli, L.; Artim-Esen, B.; et al. The 2023 ACR/EULAR Antiphospholipid Syndrome Classification Criteria. Arthritis Rheumatol. 2023, 75, 1687–1702. [Google Scholar] [CrossRef] [PubMed]
- Pignatelli, P.; Ettorre, E.; Menichelli, D.; Pani, A.; Violi, F.; Pastori, D. Seronegative Antiphospholipid Syndrome: Refining the Value of “Non-Criteria” Antibodies for Diagnosis and Clinical Management. Haematologica 2020, 105, 562–572. [Google Scholar] [CrossRef]
- Truglia, S.; Riitano, G.; Mancuso, S.; Recalchi, S.; Rapino, L.; Garufi, C.; Manganelli, V.; Garofalo, T.; Misasi, R.; Alessandri, C.; et al. Antibody Profiles in the Mosaic of ‘Seronegative’ APS Syndrome. Clin. Exp. Immunol. 2024, 218, 275–282. [Google Scholar] [CrossRef]
- Cabiedes, J.; Cabral, A.R.; López-Mendoza, A.T.; Cordero-Esperón, H.A.; Huerta, M.T.; Alarcón-Segovia, D. Characterization of Anti-Phosphatidylcholine Polyreactive Natural Autoantibodies from Normal Human Subjects. J. Autoimmun. 2002, 18, 181–190. [Google Scholar] [CrossRef]
- Merrill, J.T. Do Antiphospholipid Antibodies Develop for a Purpose? Curr. Rheumatol. Rep. 2006, 8, 109–113. [Google Scholar] [CrossRef]
- Domange Jordö, E.; Wermeling, F.; Chen, Y.; Karlsson, M.C.I. Scavenger Receptors as Regulators of Natural Antibody Responses and B Cell Activation in Autoimmunity. Mol. Immunol. 2011, 48, 1307–1318. [Google Scholar] [CrossRef]
- Von andenberg, P.; Döring, Y.; Modrow, S.; Lackner, K.J. Are Antiphospholipid Antibodies an Essential Requirement for an Effective Immune Response to Infections? Ann. N. Y. Acad. Sci. 2007, 1108, 578–583. [Google Scholar] [CrossRef]
- Lackner, K.J.; Müller-Calleja, N. Antiphospholipid Antibodies: Their Origin and Development. Antibodies 2016, 5, 15. [Google Scholar] [CrossRef]
- PIERANGELI, S.S.; HARRIS, E.N. Induction of Phospholipid-Binding Antibodies in Mice and Rabbits by Immunization with Human Β2 Glycoprotein 1 or Anticardiolipin Antibodies Alone. Clin. Exp. Immunol. 1993, 93, 269–272. [Google Scholar] [CrossRef] [PubMed]
- Volkov, M.; Kampstra, A.S.B.; van Schie, K.A.; Kawakami, A.; Tamai, M.; Kawashiri, S.; Maeda, T.; Huizinga, T.W.J.; Toes, R.E.M.; van der Woude, D. Evolution of Anti-Modified Protein Antibody Responses Can Be Driven by Consecutive Exposure to Different Post-Translational Modifications. Arthritis Res. Ther. 2021, 23, 298. [Google Scholar] [CrossRef] [PubMed]
- Manganelli, V.; Recalchi, S.; Capozzi, A.; Riitano, G.; Mattei, V.; Longo, A.; Di Franco, M.; Alessandri, C.; Bombardieri, M.; Valesini, G.; et al. Autophagy Induces Protein Carbamylation in Fibroblast-like Synoviocytes from Patients with Rheumatoid Arthritis. Rheumatology 2018, 57, 2032–2041. [Google Scholar] [CrossRef] [PubMed]
- Sorice, M.; Iannuccelli, C.; Manganelli, V.; Capozzi, A.; Alessandri, C.; Lococo, E.; Garofalo, T.; Di Franco, M.; Bombardieri, M.; Nerviani, A.; et al. Autophagy Generates Citrullinated Peptides in Human Synoviocytes: A Possible Trigger for Anti-Citrullinated Peptide Antibodies. Rheumatology 2016, 55, 1374–1385. [Google Scholar] [CrossRef]
- Riitano, G.; Recalchi, S.; Capozzi, A.; Manganelli, V.; Misasi, R.; Garofalo, T.; Sorice, M.; Longo, A. The Role of Autophagy as a Trigger of Post-Translational Modifications of Proteins and Extracellular Vesicles in the Pathogenesis of Rheumatoid Arthritis. Int. J. Mol. Sci. 2023, 24, 12764. [Google Scholar] [CrossRef]
- Wu, C.-Y.; Yang, H.-Y.; Luo, S.-F.; Lai, J.-H. From Rheumatoid Factor to Anti-Citrullinated Protein Antibodies and Anti-Carbamylated Protein Antibodies for Diagnosis and Prognosis Prediction in Patients with Rheumatoid Arthritis. Int. J. Mol. Sci. 2021, 22, 686. [Google Scholar] [CrossRef]
- Cao, W.; Li, J.; Yang, K.; Cao, D. An Overview of Autophagy: Mechanism, Regulation and Research Progress. Bull. Cancer (Paris) 2021, 108, 304–322. [Google Scholar] [CrossRef]
- Lacroix-Desmazes, S.; Kaveri, S.V.; Mouthon, L.; Ayouba, A.; Malanchère, E.; Coutinho, A.; Kazatchkine, M.D. Self-Reactive Antibodies (Natural Autoantibodies) in Healthy Individuals. J. Immunol. Methods 1998, 216, 117–137. [Google Scholar] [CrossRef]
- Czömpöly, T.; Olasz, K.; Nyárády, Z.; Simon, D.; Bovári, J.; Németh, P. Detailed Analyses of Antibodies Recognizing Mitochondrial Antigens Suggest Similar or Identical Mechanism for Production of Natural Antibodies and Natural Autoantibodies. Autoimmun. Rev. 2008, 7, 463–467. [Google Scholar] [CrossRef]
- Aziz, M.; Brenner, M.; Wang, P. Therapeutic Potential of B-1a Cells in COVID-19. Shock 2020, 54, 586. [Google Scholar] [CrossRef]
- Ray, M.E.; Rothstein, T.L. Human VH4-34 Antibodies Derived from B1 Cells Are More Frequently Autoreactive than VH4-34 Antibodies Derived from Memory Cells. Front. Immunol. 2023, 14, 1259827. [Google Scholar] [CrossRef]
| Category | nAAb Type/Target | Disease Context 1 | Key Insight | Source |
|---|---|---|---|---|
| Protective IgM nAAbs | IgM against apoptotic cells, oxidized self-antigens | SLE | Higher IgM levels are linked to milder disease and protection from nephritis and atherosclerosis. | [74] |
| Prognostic marker | IgM anti-topoisomerase I (Topo I) | SSc | The presence of IgM anti-Topo I alongside IgG anti-Topo I predicts more rapid skin/lung progression. | [146,152] |
| Inflammation correlate | IgM/IgG nAAbs against mitochondrial enzymes, HSPs | RA, AS | Baseline and therapy-modulated levels predict CRP, disease activity, and vascular health. | [153] |
| Disease risk marker | Autoantibodies (e.g., anti-type I IFNs) | T1D, APS-1 | The presence of neutralizing IFN autoantibodies correlated with protection from type 1 diabetes. | [154] |
| Early biomarkers | IgM nAAb “fingerprints” (from arrays) | T1D, RA, SLE | Autoantibody profiles may distinguish preclinical patients and predict autoimmune progression. | [116,155,156] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Böröcz, K.; Szinger, D.; Simon, D.; Berki, T.; Németh, P. Regulators and Conductors of Immunity: Natural Immune System in Health and Autoimmunity. Int. J. Mol. Sci. 2025, 26, 5413. https://doi.org/10.3390/ijms26115413
Böröcz K, Szinger D, Simon D, Berki T, Németh P. Regulators and Conductors of Immunity: Natural Immune System in Health and Autoimmunity. International Journal of Molecular Sciences. 2025; 26(11):5413. https://doi.org/10.3390/ijms26115413
Chicago/Turabian StyleBöröcz, Katalin, Dávid Szinger, Diána Simon, Timea Berki, and Péter Németh. 2025. "Regulators and Conductors of Immunity: Natural Immune System in Health and Autoimmunity" International Journal of Molecular Sciences 26, no. 11: 5413. https://doi.org/10.3390/ijms26115413
APA StyleBöröcz, K., Szinger, D., Simon, D., Berki, T., & Németh, P. (2025). Regulators and Conductors of Immunity: Natural Immune System in Health and Autoimmunity. International Journal of Molecular Sciences, 26(11), 5413. https://doi.org/10.3390/ijms26115413