
IL-6R Signaling Is Associated with PAD4 and Neutrophil Extracellular Trap Formation in Patients with STEMI
 , , and
, , and
Abstract

1. Introduction
2. Results
2.1. Baseline Characteristics
2.2. Investigations in Coronary Thrombi
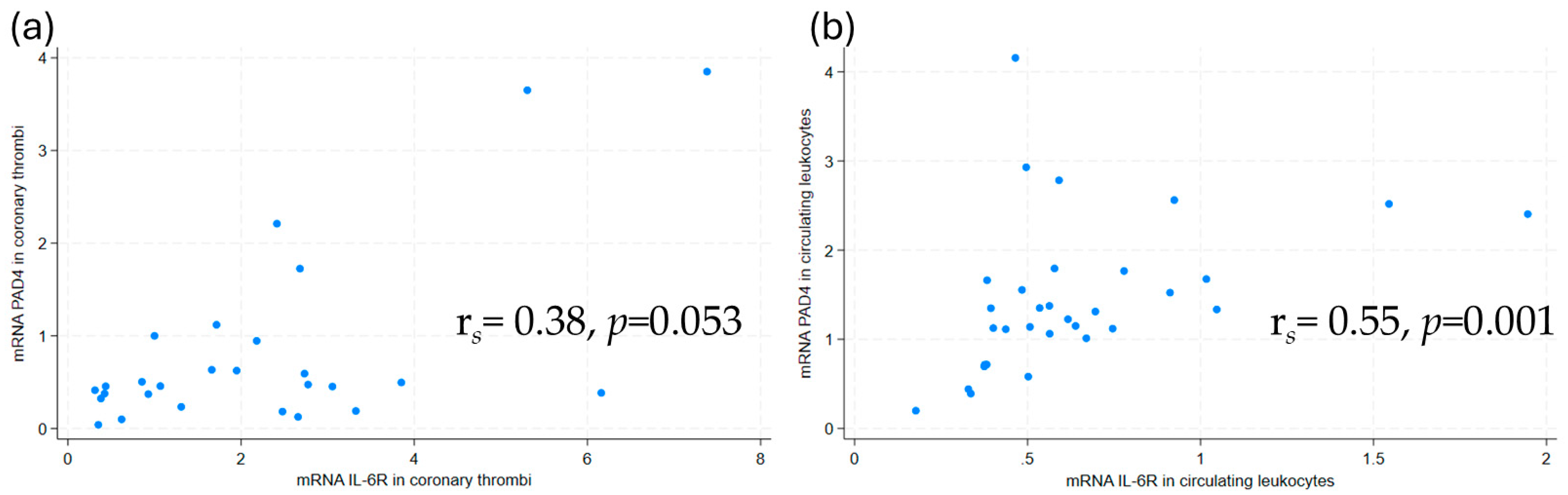
2.2.1. Associations Between PAD4, IL-6R, and IL-6 mRNA Levels
2.2.2. Associations Between PAD4 mRNA Levels and Troponin T
2.2.3. Immunofluorescence Investigations of NETosis
2.3. Investigations in Peripheral Blood
2.3.1. Associations Between PAD4 mRNA, IL-6, IL-6R, and NETs
2.3.2. Comparison of Coronary PAD4 mRNA and Circulating Markers
3. Discussion
Limitations
4. Materials and Methods
4.1. Study Design
4.2. Thrombi Preparation
4.3. Blood Sampling Protocol
4.4. Laboratory Analysis
4.5. Statistical Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| STEMI | ST-elevation myocardial infarction |
| Il-6 | Interleukin-6 |
| IL-6R | Interleukin-6 receptor |
| NETs | Neutrophil extracellular traps |
| PAD4 | Peptidylarginine deiminase 4 |
| dsDNA | Double-stranded deoxyribonucleic acid |
| H3Cit | Citrullinated histone H3 |
| MPO-DNA | Myeloperioxidase bound to deoxyribonucleic acid |
| NLRP3 | NOD-, LRR-, and pyrin domain-containing protein 3 |
| ASSAIL-MI | Assessing the effect of anti-IL-6 treatment in myocardial infarction |
| PCI | Percutaneous coronary intervention |
| FFPE | Formalin-fixed, paraffin-embedded |
| ELISA | Enzyme-linked immunosorbent assay |
References
- Tollefsen, I.M.; Shetelig, C.; Seljeflot, I.; Eritsland, J.; Hoffmann, P.; Andersen, G.O. High levels of interleukin-6 are associated with final infarct size and adverse clinical events in patients with STEMI. Open Heart 2021, 8, e001869. [Google Scholar] [CrossRef]
- Groot, H.E.; Al Ali, L.; van der Horst, I.C.C.; Schurer, R.A.J.; van der Werf, H.W.; Lipsic, E.; van Veldhuisen, D.J.; Karper, J.C.; van der Harst, P. Plasma interleukin 6 levels are associated with cardiac function after ST-elevation myocardial infarction. Clin. Res. Cardiol. 2019, 108, 612–621. [Google Scholar] [CrossRef]
- Libby, P. Targeting Inflammatory Pathways in Cardiovascular Disease: The Inflammasome, Interleukin-1, Interleukin-6 and Beyond. Cells 2021, 10, 951. [Google Scholar] [CrossRef]
- Broch, K.; Anstensrud, A.K.; Woxholt, S.; Sharma, K.; Tollefsen, I.M.; Bendz, B.; Aakhus, S.; Ueland, T.; Amundsen, B.H.; Damas, J.K.; et al. Randomized Trial of Interleukin-6 Receptor Inhibition in Patients With Acute ST-Segment Elevation Myocardial Infarction. J. Am. Coll. Cardiol. 2021, 77, 1845–1855. [Google Scholar] [CrossRef]
- Feng, Y.; Ye, D.; Wang, Z.; Pan, H.; Lu, X.; Wang, M.; Xu, Y.; Yu, J.; Zhang, J.; Zhao, M.; et al. The Role of Interleukin-6 Family Members in Cardiovascular Diseases. Front. Cardiovasc. Med. 2022, 9, 818890. [Google Scholar] [CrossRef]
- Kindberg, K.M.; Broch, K.; Andersen, G.O.; Anstensrud, A.K.; Akra, S.; Woxholt, S.; Tollefsen, I.M.; Ueland, T.; Amundsen, B.H.; Klow, N.E.; et al. Neutrophil Extracellular Traps in ST-Segment Elevation Myocardial Infarction: Reduced by Tocilizumab and Associated With Infarct Size. JACC Adv. 2024, 3, 101193. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Mangold, A.; Alias, S.; Scherz, T.; Hofbauer, M.; Jakowitsch, J.; Panzenbock, A.; Simon, D.; Laimer, D.; Bangert, C.; Kammerlander, A.; et al. Coronary neutrophil extracellular trap burden and deoxyribonuclease activity in ST-elevation acute coronary syndrome are predictors of ST-segment resolution and infarct size. Circ. Res. 2015, 116, 1182–1192. [Google Scholar] [CrossRef]
- Doring, Y.; Soehnlein, O.; Weber, C. Neutrophil Extracellular Traps in Atherosclerosis and Atherothrombosis. Circ. Res. 2017, 120, 736–743. [Google Scholar] [CrossRef]
- Nappi, F.; Bellomo, F.; Avtaar Singh, S.S. Worsening Thrombotic Complication of Atherosclerotic Plaques Due to Neutrophils Extracellular Traps: A Systematic Review. Biomedicines 2023, 11, 113. [Google Scholar] [CrossRef]
- Varju, I.; Longstaff, C.; Szabo, L.; Farkas, A.Z.; Varga-Szabo, V.J.; Tanka-Salamon, A.; Machovich, R.; Kolev, K. DNA, histones and neutrophil extracellular traps exert anti-fibrinolytic effects in a plasma environment. Thromb. Haemost. 2015, 113, 1289–1298. [Google Scholar] [CrossRef]
- Folco, E.J.; Mawson, T.L.; Vromman, A.; Bernardes-Souza, B.; Franck, G.; Persson, O.; Nakamura, M.; Newton, G.; Luscinskas, F.W.; Libby, P. Neutrophil Extracellular Traps Induce Endothelial Cell Activation and Tissue Factor Production Through Interleukin-1alpha and Cathepsin G. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 1901–1912. [Google Scholar] [CrossRef]
- Helseth, R.; Shetelig, C.; Andersen, G.O.; Langseth, M.S.; Limalanathan, S.; Opstad, T.B.; Arnesen, H.; Hoffmann, P.; Eritsland, J.; Seljeflot, I. Neutrophil Extracellular Trap Components Associate with Infarct Size, Ventricular Function, and Clinical Outcome in STEMI. Mediat. Inflamm. 2019, 2019, 7816491. [Google Scholar] [CrossRef]
- Blasco, A.; Coronado, M.J.; Vela, P.; Martin, P.; Solano, J.; Ramil, E.; Mesquida, A.; Santos, A.; Cozar, B.; Royuela, A.; et al. Prognostic Implications of Neutrophil Extracellular Traps in Coronary Thrombi of Patients with ST-Elevation Myocardial Infarction. Thromb. Haemost. 2022, 122, 1415–1428. [Google Scholar] [CrossRef]
- Novotny, J.; Chandraratne, S.; Weinberger, T.; Philippi, V.; Stark, K.; Ehrlich, A.; Pircher, J.; Konrad, I.; Oberdieck, P.; Titova, A.; et al. Histological comparison of arterial thrombi in mice and men and the influence of Cl-amidine on thrombus formation. PLoS ONE 2018, 13, e0190728. [Google Scholar] [CrossRef]
- Riegger, J.; Byrne, R.A.; Joner, M.; Chandraratne, S.; Gershlick, A.H.; Ten Berg, J.M.; Adriaenssens, T.; Guagliumi, G.; Godschalk, T.C.; Neumann, F.J.; et al. Histopathological evaluation of thrombus in patients presenting with stent thrombosis. A multicenter European study: A report of the prevention of late stent thrombosis by an interdisciplinary global European effort consortium. Eur. Heart J. 2016, 37, 1538–1549. [Google Scholar] [CrossRef]
- Fuchs, T.A.; Brill, A.; Duerschmied, D.; Schatzberg, D.; Monestier, M.; Myers, D.D., Jr.; Wrobleski, S.K.; Wakefield, T.W.; Hartwig, J.H.; Wagner, D.D. Extracellular DNA traps promote thrombosis. Proc. Natl. Acad. Sci. USA 2010, 107, 15880–15885. [Google Scholar] [CrossRef]
- Gould, T.J.; Vu, T.T.; Swystun, L.L.; Dwivedi, D.J.; Mai, S.H.; Weitz, J.I.; Liaw, P.C. Neutrophil extracellular traps promote thrombin generation through platelet-dependent and platelet-independent mechanisms. Arter. Thromb. Vasc. Biol. 2014, 34, 1977–1984. [Google Scholar] [CrossRef]
- Thiam, H.R.; Wong, S.L.; Qiu, R.; Kittisopikul, M.; Vahabikashi, A.; Goldman, A.E.; Goldman, R.D.; Wagner, D.D.; Waterman, C.M. NETosis proceeds by cytoskeleton and endomembrane disassembly and PAD4-mediated chromatin decondensation and nuclear envelope rupture. Proc. Natl. Acad. Sci. USA 2020, 117, 7326–7337. [Google Scholar] [CrossRef]
- Du, M.; Yang, W.; Schmull, S.; Gu, J.; Xue, S. Inhibition of peptidyl arginine deiminase-4 protects against myocardial infarction induced cardiac dysfunction. Int. Immunopharmacol. 2020, 78, 106055. [Google Scholar] [CrossRef]
- Nordeng, J.; Schandiz, H.; Solheim, S.; Akra, S.; Hoffman, P.; Roald, B.; Bendz, B.; Arnesen, H.; Helseth, R.; Seljeflot, I. The Inflammasome Signaling Pathway Is Actively Regulated and Related to Myocardial Damage in Coronary Thrombi from Patients with STEMI. Mediat. Inflamm. 2021, 2021, 5525917. [Google Scholar] [CrossRef] [PubMed]
- Thalin, C.; Hisada, Y.; Lundstrom, S.; Mackman, N.; Wallen, H. Neutrophil Extracellular Traps: Villains and Targets in Arterial, Venous, and Cancer-Associated Thrombosis. Arter. Thromb. Vasc. Biol. 2019, 39, 1724–1738. [Google Scholar] [CrossRef]
- Bonaventura, A.; Vecchie, A.; Abbate, A.; Montecucco, F. Neutrophil Extracellular Traps and Cardiovascular Diseases: An Update. Cells 2020, 9, 231. [Google Scholar] [CrossRef] [PubMed]
- Farkas, A.Z.; Farkas, V.J.; Gubucz, I.; Szabo, L.; Balint, K.; Tenekedjiev, K.; Nagy, A.I.; Sotonyi, P.; Hidi, L.; Nagy, Z.; et al. Neutrophil extracellular traps in thrombi retrieved during interventional treatment of ischemic arterial diseases. Thromb. Res. 2019, 175, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Longstaff, C.; Varju, I.; Sotonyi, P.; Szabo, L.; Krumrey, M.; Hoell, A.; Bota, A.; Varga, Z.; Komorowicz, E.; Kolev, K. Mechanical stability and fibrinolytic resistance of clots containing fibrin, DNA, and histones. J. Biol. Chem. 2013, 288, 6946–6956. [Google Scholar] [CrossRef]
- Yipp, B.G.; Kubes, P. NETosis: How vital is it? Blood 2013, 122, 2784–2794. [Google Scholar] [CrossRef]
- McGill, C.J.; Lu, R.J.; Benayoun, B.A. Protocol for analysis of mouse neutrophil NETosis by flow cytometry. STAR Protoc. 2021, 2, 100948. [Google Scholar] [CrossRef]
- Stakos, D.A.; Kambas, K.; Konstantinidis, T.; Mitroulis, I.; Apostolidou, E.; Arelaki, S.; Tsironidou, V.; Giatromanolaki, A.; Skendros, P.; Konstantinides, S.; et al. Expression of functional tissue factor by neutrophil extracellular traps in culprit artery of acute myocardial infarction. Eur. Heart J. 2015, 36, 1405–1414. [Google Scholar] [CrossRef]
- Heger, L.A.; Schommer, N.; Van Bruggen, S.; Sheehy, C.E.; Chan, W.; Wagner, D.D. Neutrophil NLRP3 promotes cardiac injury following acute myocardial infarction through IL-1beta production, VWF release and NET deposition in the myocardium. Sci. Rep. 2024, 14, 14524. [Google Scholar] [CrossRef]
- Munzer, P.; Negro, R.; Fukui, S.; di Meglio, L.; Aymonnier, K.; Chu, L.; Cherpokova, D.; Gutch, S.; Sorvillo, N.; Shi, L.; et al. NLRP3 Inflammasome Assembly in Neutrophils Is Supported by PAD4 and Promotes NETosis Under Sterile Conditions. Front. Immunol. 2021, 12, 683803. [Google Scholar] [CrossRef]
- Yahagi, A.; Saika, T.; Hirano, H.; Takai-Imamura, M.; Tsuji, F.; Aono, H.; Iseki, M.; Morita, Y.; Igarashi, H.; Saeki, Y.; et al. IL-6-PAD4 axis in the earliest phase of arthritis in knock-in gp130F759 mice, a model for rheumatoid arthritis. RMD Open 2019, 5, e000853. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Yu, H.; Yang, X.; Zhu, Y.; Bai, X.; Wang, R.; Cao, Y.; Xu, H.; Luo, H.; Lu, L.; et al. Neutrophil extracellular traps released by neutrophils impair revascularization and vascular remodeling after stroke. Nat. Commun. 2020, 11, 2488. [Google Scholar] [CrossRef]
- Yang, K.; Gao, R.; Chen, H.; Hu, J.; Zhang, P.; Wei, X.; Shi, J.; Chen, Y.; Zhang, L.; Chen, J.; et al. Myocardial reperfusion injury exacerbation due to ALDH2 deficiency is mediated by neutrophil extracellular traps and prevented by leukotriene C4 inhibition. Eur. Heart J. 2024, 45, 1662–1680. [Google Scholar] [CrossRef]
- Hausenloy, D.J.; Yellon, D.M. Myocardial ischemia-reperfusion injury: A neglected therapeutic target. J. Clin. Investig. 2013, 123, 92–100. [Google Scholar] [CrossRef]
- Yellon, D.M.; Hausenloy, D.J. Myocardial reperfusion injury. N. Engl. J. Med. 2007, 357, 1121–1135. [Google Scholar] [CrossRef]
- Shah, M.; Yellon, D.M.; Davidson, S.M. The Role of Extracellular DNA and Histones in Ischaemia-Reperfusion Injury of the Myocardium. Cardiovasc. Drugs Ther. 2020, 34, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Saisorn, W.; Santiworakul, C.; Phuengmaung, P.; Siripen, N.; Rianthavorn, P.; Leelahavanichkul, A. Extracellular traps in peripheral blood mononuclear cell fraction in childhood-onset systemic lupus erythematosus. Sci. Rep. 2024, 14, 23177. [Google Scholar] [CrossRef]
- Paues Goranson, S.; Thalin, C.; Lundstrom, A.; Hallstrom, L.; Lasselin, J.; Wallen, H.; Soop, A.; Mobarrez, F. Circulating H3Cit is elevated in a human model of endotoxemia and can be detected bound to microvesicles. Sci. Rep. 2018, 8, 12641. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Li, Y.; Sun, R.; Hu, H.; Liu, Y.; Herrmann, M.; Zhao, Y.; Munoz, L.E. Receptor-Mediated NETosis on Neutrophils. Front. Immunol. 2021, 12, 775267. [Google Scholar] [CrossRef]
- Petretto, A.; Bruschi, M.; Pratesi, F.; Croia, C.; Candiano, G.; Ghiggeri, G.; Migliorini, P. Neutrophil extracellular traps (NET) induced by different stimuli: A comparative proteomic analysis. PLoS ONE 2019, 14, e0218946. [Google Scholar] [CrossRef]
- Kessenbrock, K.; Krumbholz, M.; Schonermarck, U.; Back, W.; Gross, W.L.; Werb, Z.; Grone, H.J.; Brinkmann, V.; Jenne, D.E. Netting neutrophils in autoimmune small-vessel vasculitis. Nat. Med. 2009, 15, 623–625. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Total n = 33 | |
|---|---|
| Age, years | 60 (±11) |
| Females | 3 (9%) |
| Current or previous smoker | 27 (84%) |
| BMI, kg/m2 | 27.7 (23.4, 28.6) |
| sBP, mmHg | 127 (±31) |
| dBP, mmHg | 82 (±19) |
| Heart rate | 70 (65, 90) |
| Prior myocardial infarction | 1 (3%) |
| Prior hypertension | 11 (33%) |
| Prior diabetes mellitus type 2 | 4 (12%) |
| Prior medication: | |
| Acetylsalicylic acid | 6 (18%) |
| P2Y12-inhibitor | 2 (6%) |
| Anticoagulation | 3 (9%) |
| RAAS inhibitors | 5 (15%) |
| Betablocker | 4 (12%) |
| Aldosterone antagonist | 0 (0%) |
| Statins | 6 (18%) |
| Symptom start to PCI, min | 152 (122, 343) |
| Culprit artery: | |
| LAD | 16 (48%) |
| Cx | 6 (18%) |
| RCA | 11 (33%) |
| Troponin T peak, µg/L | 3434 (1250, 6967) |
| CRP, mg/L | 2.71 (0.96, 6.03) |
| Gene Expression in Thrombi | Gene Expression in Circulating Leukocytes | Circulating Levels | |
|---|---|---|---|
| IL-6 | √ | √ | √ |
| IL-6R | √ | √ | √ |
| PAD4 | √ | √ | √ |
| dsDNA | x | x | √ |
| MPO-DNA | x | x | √ |
| H3Cit | x | x | √ |
| Univariate | Multivariate | |||||
|---|---|---|---|---|---|---|
| β | CI | p-value | β | CI | p-value | |
| Association with PAD4 mRNA in coronary thrombi | ||||||
| IL-6R mRNA | 0.34 | 0.16–0.51 | 0.001 | 0.34 | 0.16–0.53 | 0.001 * |
| Circulating peak troponin T, quartile q4 vs. 1–3 | 1.0 | 0.20–1.80 | 0.016 | 1.15 | 0.27–2.04 | 0.013 * |
| Association with PAD4 mRNA in circulating leukocytes | ||||||
| IL-6R mRNA | 0.98 | 0.18–1.77 | 0.017 | 0.92 | 0.07–1.77 | 0.036 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kindberg, K.M.; Nordeng, J.; Langseth, M.S.; Schandiz, H.; Roald, B.; Solheim, S.; Seljeflot, I.; Stokke, M.K.; Helseth, R. IL-6R Signaling Is Associated with PAD4 and Neutrophil Extracellular Trap Formation in Patients with STEMI. Int. J. Mol. Sci. 2025, 26, 5348. https://doi.org/10.3390/ijms26115348
Kindberg KM, Nordeng J, Langseth MS, Schandiz H, Roald B, Solheim S, Seljeflot I, Stokke MK, Helseth R. IL-6R Signaling Is Associated with PAD4 and Neutrophil Extracellular Trap Formation in Patients with STEMI. International Journal of Molecular Sciences. 2025; 26(11):5348. https://doi.org/10.3390/ijms26115348
Chicago/Turabian StyleKindberg, Kristine Mørk, Jostein Nordeng, Miriam Sjåstad Langseth, Hossein Schandiz, Borghild Roald, Svein Solheim, Ingebjørg Seljeflot, Mathis Korseberg Stokke, and Ragnhild Helseth. 2025. "IL-6R Signaling Is Associated with PAD4 and Neutrophil Extracellular Trap Formation in Patients with STEMI" International Journal of Molecular Sciences 26, no. 11: 5348. https://doi.org/10.3390/ijms26115348
APA StyleKindberg, K. M., Nordeng, J., Langseth, M. S., Schandiz, H., Roald, B., Solheim, S., Seljeflot, I., Stokke, M. K., & Helseth, R. (2025). IL-6R Signaling Is Associated with PAD4 and Neutrophil Extracellular Trap Formation in Patients with STEMI. International Journal of Molecular Sciences, 26(11), 5348. https://doi.org/10.3390/ijms26115348