
Network Pharmacology and Molecular Docking-Based Approach to Explore Potential Bioactive Compounds from Kaempferia parviflora on Chemokine Signaling Pathways in the Treatment of Psoriasis Disease

 , and
, and
Abstract
1. Introduction
2. Results
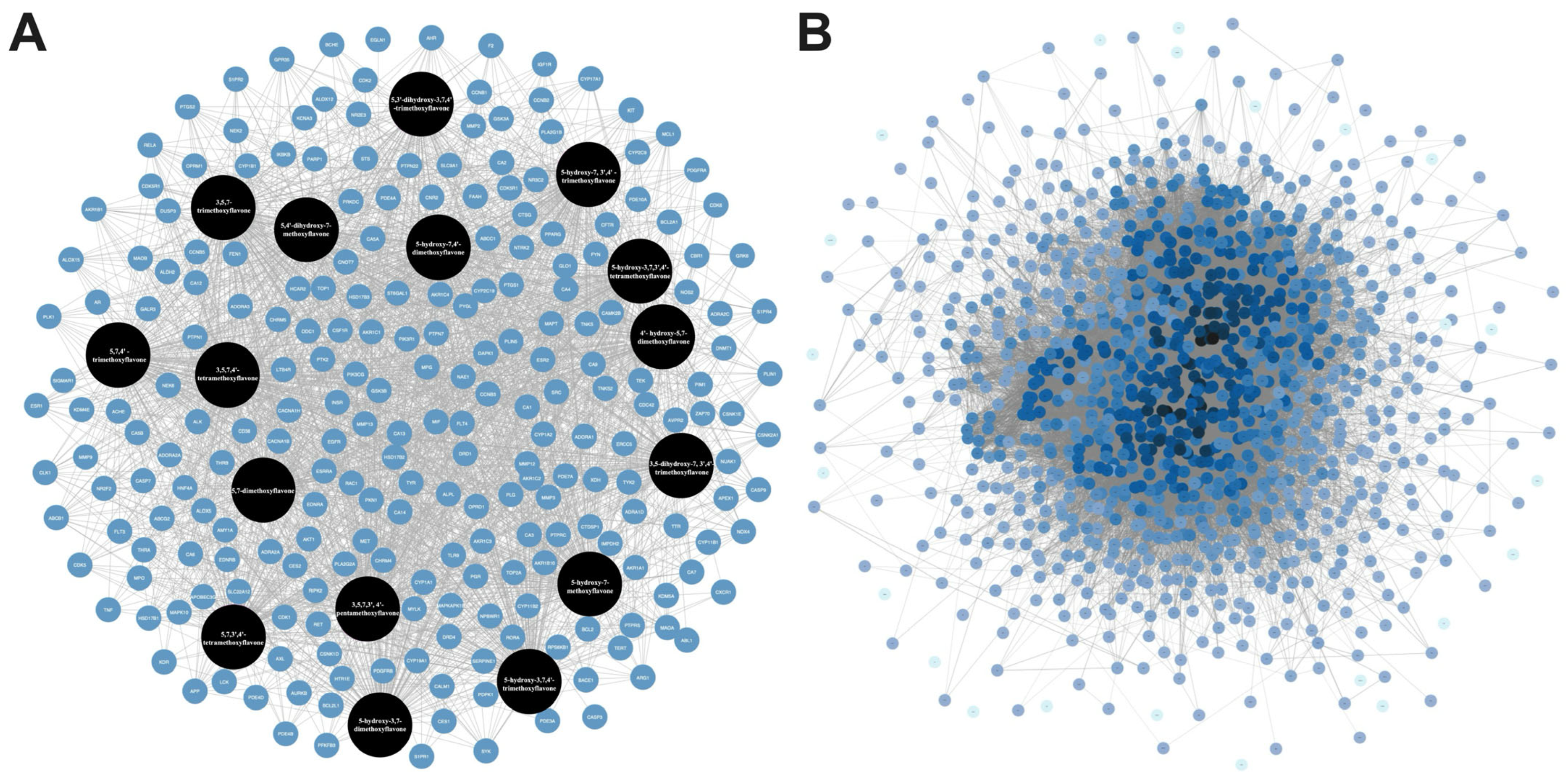
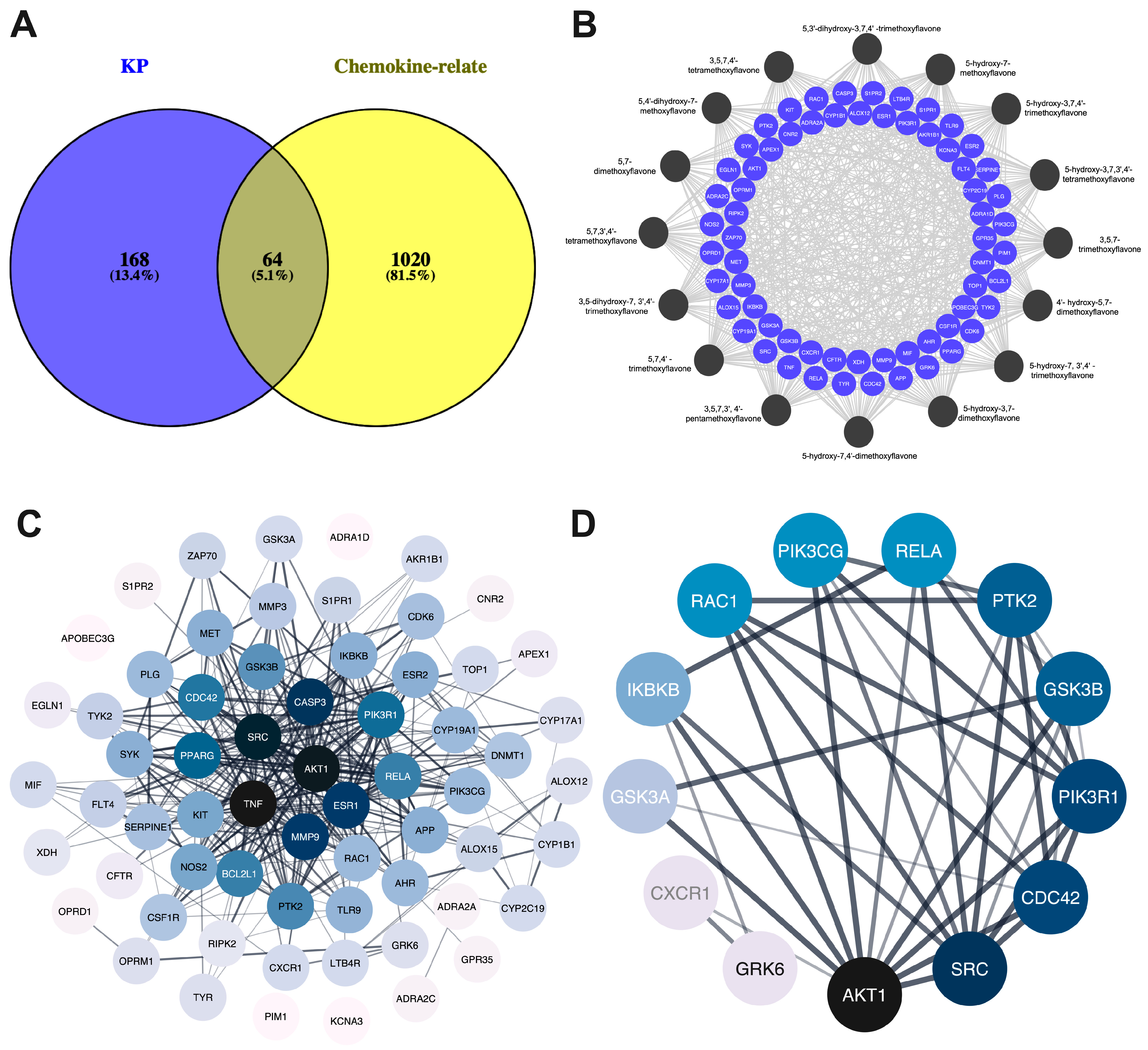
2.1. Bioactive Components, Target Prediction of KP, and Screening of Psoriasis-Related Chemokine Target Genes
2.2. PPI Network of Target Genes
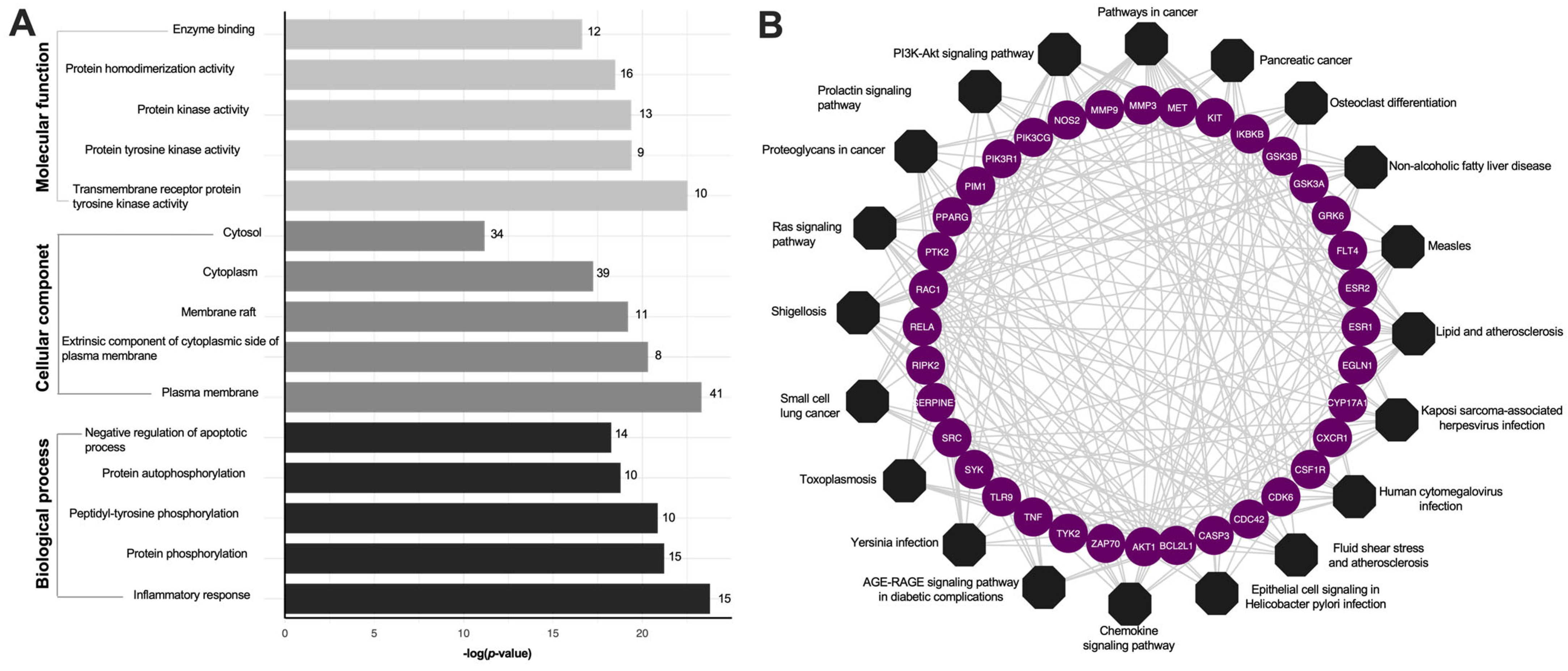
2.3. Gene Ontology (GO) and KEGG Pathway Enrichment Analyses
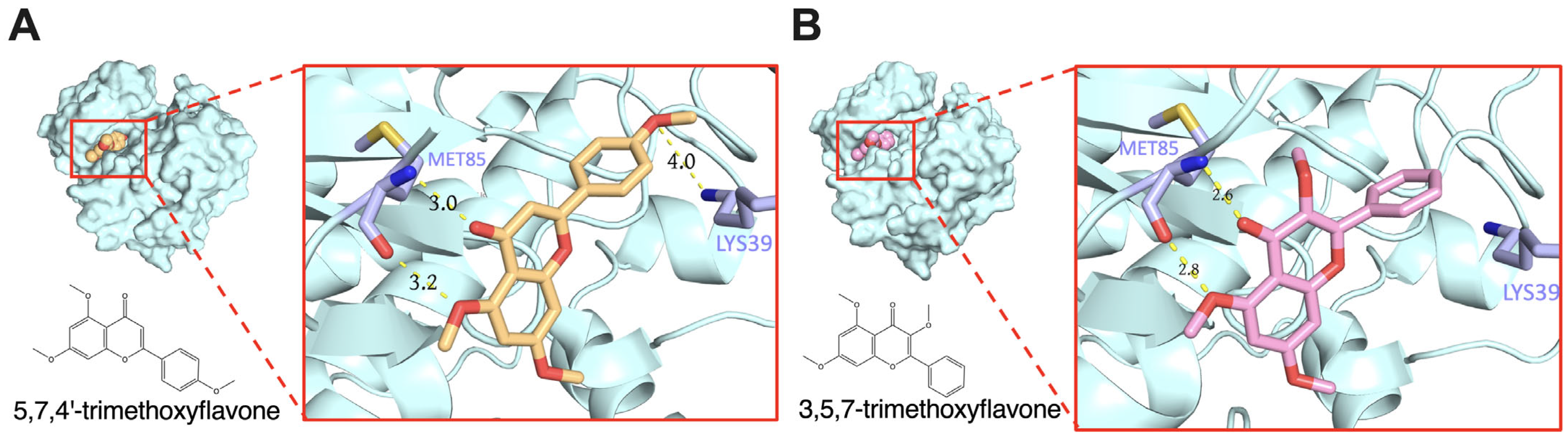
2.4. In Silico Analysis of Bioactive Compounds in KP on Psoriasis-Related Chemokine Target Genes
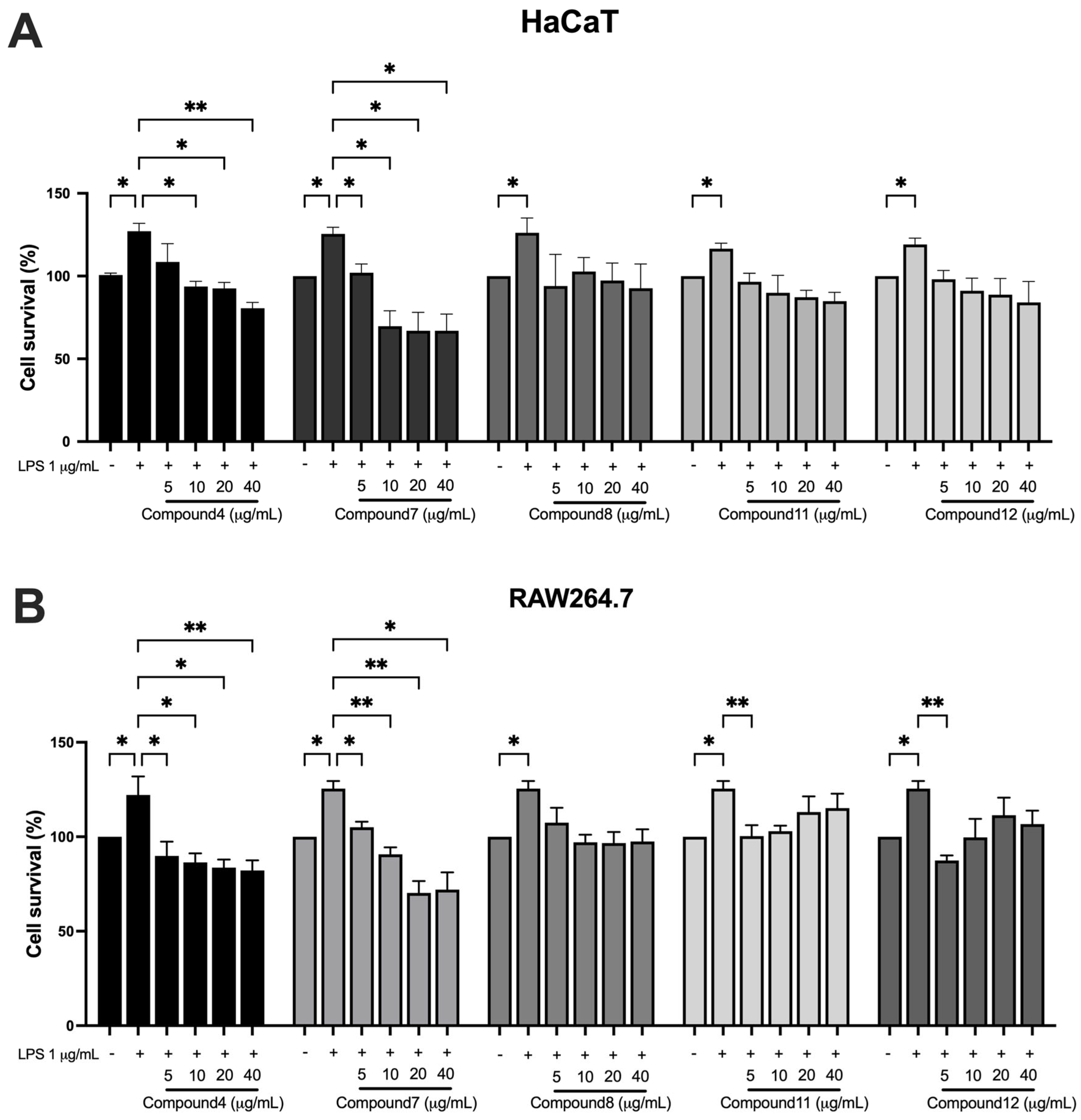
2.5. Anti-Proliferation Effects of Bioactive Compounds in KP on HaCaT and RAW264.7 Cells
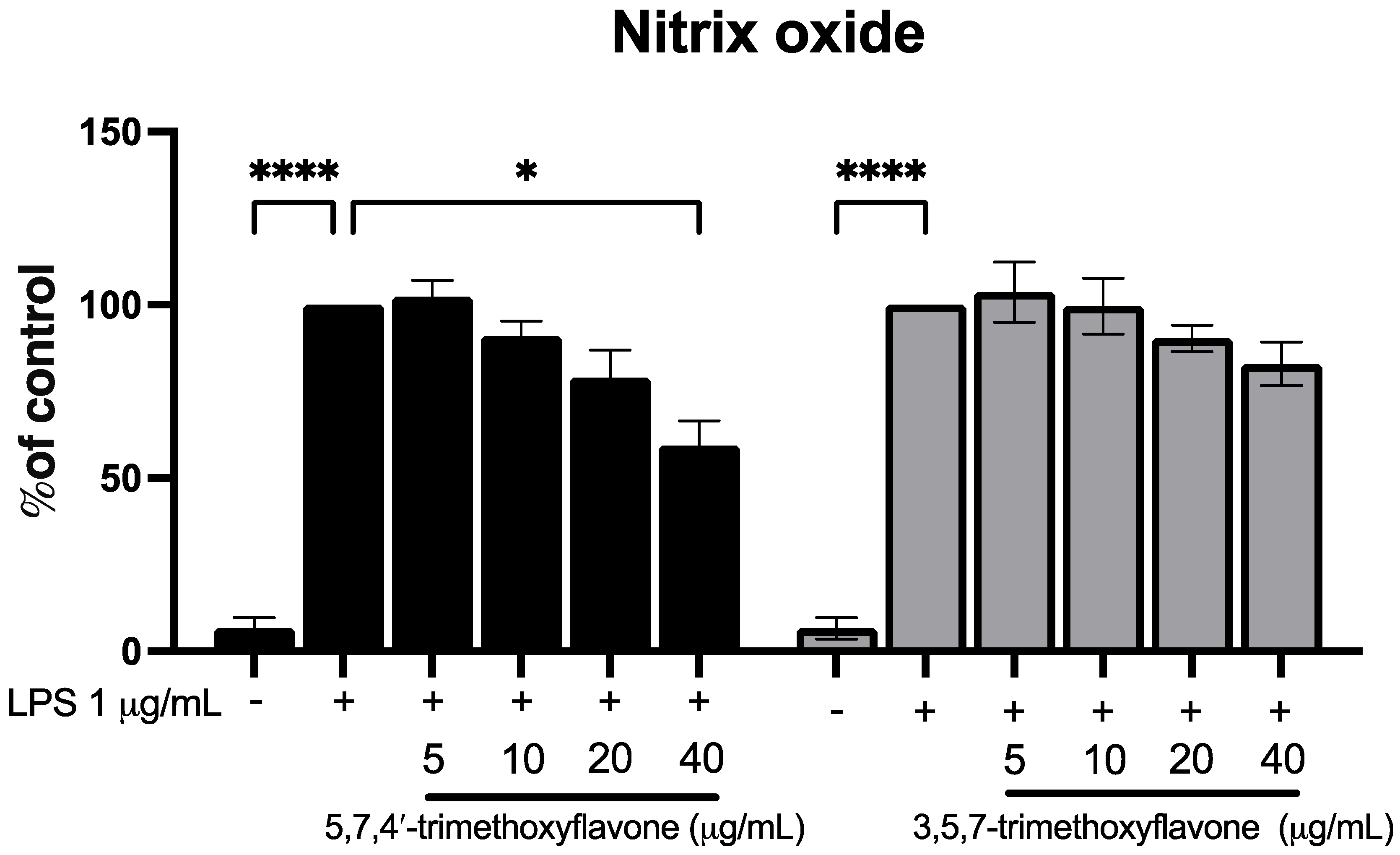
2.6. Effects of Bioactive Compounds in KP on RAW264.7 Cell Nitric Oxide Production and HaCaT Cell Migration
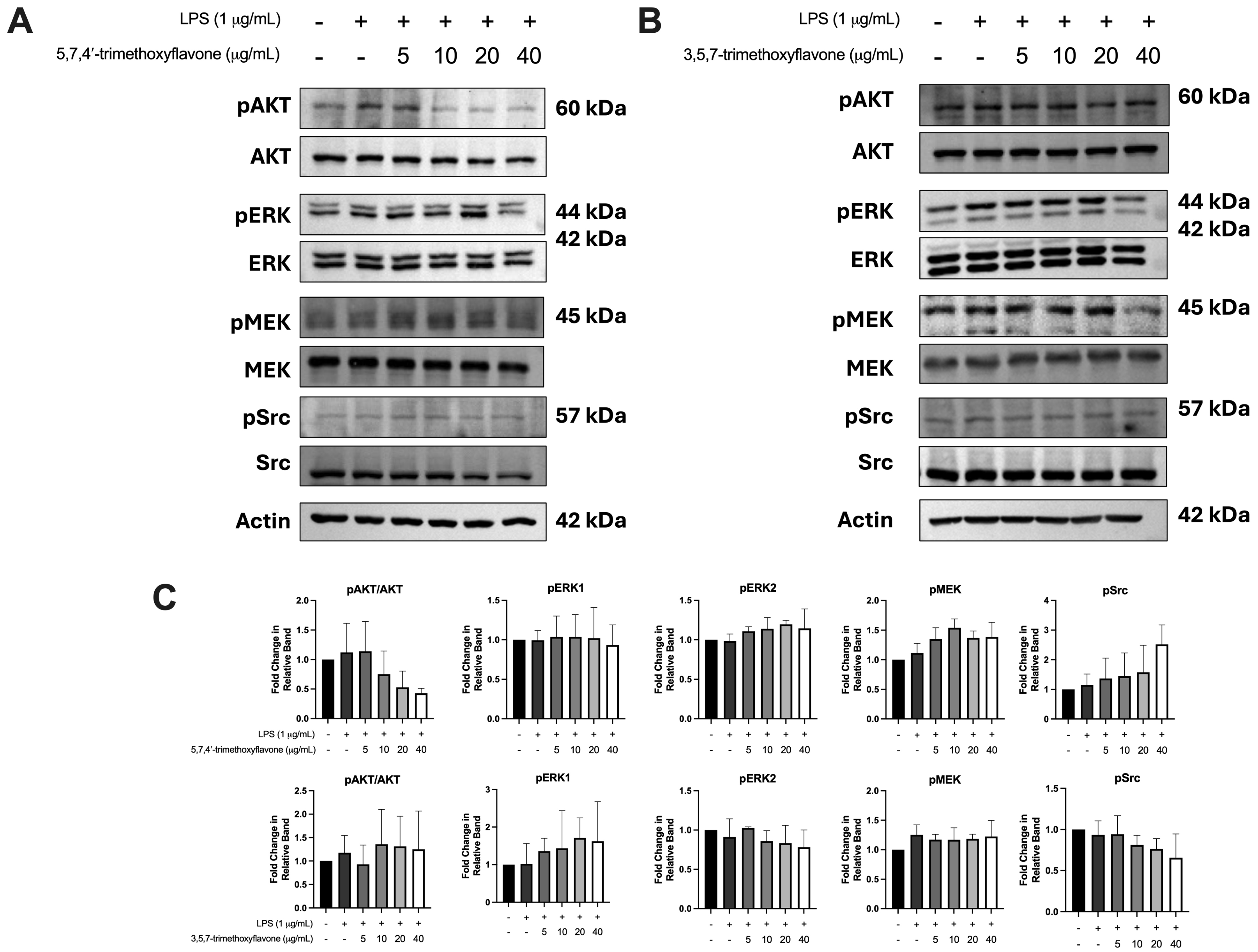
2.7. Effect of Bioactive Compounds in KP on Protein Expressions of Chemokine Signaling Pathways
3. Discussion
4. Materials and Methods
4.1. Screening the Bioactive Compounds and Target Prediction of KP
4.2. Screening for Psoriasis-Related Chemokine and Potential Target Genes for KP
4.3. GO Enrichment and KEGG Pathway Analysis
4.4. Screening of Core Targets and Protein–Protein Interaction Network Construction
4.5. Network Construction
4.6. Molecular Docking
4.7. Chemicals and Reagents
4.8. Cell Culture
4.9. Cell Proliferation
4.10. Cell Migration Assay
4.11. Western Blotting
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Christophers, E. Psoriasis: Clinical Spectrum and Classification; Future Medicine: London, UK, 2011. [Google Scholar]
- Michalek, I.M.; Loring, B.; John, S.M. Global Report on Psoriasis; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Li, J.; Yu, M.; Wang, Y.-w.; Zhang, J.-a.; Ju, M.; Chen, K.; Jiang, Y.; Li, M.; Chen, X.-S. Prevalence of psoriasis and associated risk factors in China: Protocol of a nationwide, population-based, cross-sectional study. BMJ Open 2019, 9, e027685. [Google Scholar] [CrossRef]
- Honma, M.; Hayashi, K. Psoriasis: Recent progress in molecular-targeted therapies. J. Dermatol. 2021, 48, 761–777. [Google Scholar] [CrossRef]
- Patel, A.B.; Tsilioni, I.; Weng, Z.; Theoharides, T.C. TNF stimulates IL-6, CXCL 8 and VEGF secretion from human keratinocytes via activation of mTOR, inhibited by tetramethoxyluteolin. Exp. Dermatol. 2018, 27, 135–143. [Google Scholar] [CrossRef]
- Buerger, C.; Shirsath, N.; Lang, V.; Berard, A.; Diehl, S.; Kaufmann, R.; Boehncke, W.-H.; Wolf, P. Inflammation dependent mTORC1 signaling interferes with the switch from keratinocyte proliferation to differentiation. PLoS ONE 2017, 12, e0180853. [Google Scholar] [CrossRef]
- Bonigen, J.; Raynaud-Donzel, C.; Hureaux, J.; Kramkimel, N.; Blom, A.; Jeudy, G.; Breton, A.; Hubiche, T.; Bedane, C.; Legoupil, D. Anti-PD1-induced psoriasis: A study of 21 patients. J. Eur. Acad. Dermatol. Venereol. JEADV 2017, 31, e254–e257. [Google Scholar] [CrossRef]
- Kim, D.-W.; Park, S.-K.; Woo, S.-H.; Yun, S.-K.; Kim, H.-U.; Park, J. New-onset psoriasis induced by rituximab therapy for non-Hodgkin lymphoma in a child. Eur. J. Dermatol. 2016, 26, 190–191. [Google Scholar] [CrossRef]
- Onishi, R.M.; Gaffen, S.L. Interleukin-17 and its target genes: Mechanisms of interleukin-17 function in disease. Immunology 2010, 129, 311–321. [Google Scholar] [CrossRef]
- Zdanowska, N.; Kasprowicz-Furmańczyk, M.; Placek, W.; Owczarczyk-Saczonek, A. The Role of Chemokines in Psoriasis-An Overview. Medicina 2021, 57, 754. [Google Scholar] [CrossRef]
- Nast, A.; Smith, C.; Spuls, P.I.; Avila Valle, G.; Bata-Csörgö, Z.; Boonen, H.; De Jong, E.; Garcia-Doval, I.; Gisondi, P.; Kaur-Knudsen, D.; et al. EuroGuiDerm Guideline on the systemic treatment of Psoriasis vulgaris—Part 1: Treatment and monitoring recommendations. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 2461–2498. [Google Scholar] [CrossRef]
- Zachariae, H. Methotrexate side-effects. Br. J. Dermatol. 1990, 122 (Suppl. 36), 127–133. [Google Scholar] [CrossRef]
- Schön, M.P. Advances in psoriasis treatment. Lancet 2005, 366, 1333–1335. [Google Scholar] [CrossRef]
- Gangadevi, V.; Thatikonda, S.; Pooladanda, V.; Devabattula, G.; Godugu, C. Selenium nanoparticles produce a beneficial effect in psoriasis by reducing epidermal hyperproliferation and inflammation. J. Nanobiotechnol. 2021, 19, 101. [Google Scholar] [CrossRef]
- Hopkins, A.L. Network pharmacology. Nat. Biotechnol. 2007, 25, 1110–1111. [Google Scholar] [CrossRef]
- Chandran, U.; Mehendale, N.; Patil, S.; Chaguturu, R.; Patwardhan, B. Network pharmacology. Innov. Approaches Drug Discov. 2017, 127–164. [Google Scholar] [CrossRef]
- Kumar, P.; Ghosh, I. Application of In Silico Drug Repurposing in Infectious Diseases. In In Silico Drug Design; Elsevier: Amsterdam, The Netherlands, 2019; pp. 427–462. [Google Scholar]
- Pujol, A.; Mosca, R.; Farrés, J.; Aloy, P. Unveiling the role of network and systems biology in drug discovery. Trends Pharmacol. Sci. 2010, 31, 115–123. [Google Scholar] [CrossRef]
- Zhou, Z.; Chen, B.; Chen, S.; Lin, M.; Chen, Y.; Jin, S.; Chen, W.; Zhang, Y. Applications of Network Pharmacology in Traditional Chinese Medicine Research. Evid.-Based Complement. Altern. Med. 2020, 2020, 1646905. [Google Scholar] [CrossRef]
- Zhang, R.; Zhu, X.; Bai, H.; Ning, K. Network Pharmacology Databases for Traditional Chinese Medicine: Review and Assessment. Front. Pharmacol. 2019, 10, 123. [Google Scholar] [CrossRef]
- Noor, F.; Tahir Ul Qamar, M.; Ashfaq, U.A.; Albutti, A.; Alwashmi, A.S.S.; Aljasir, M.A. Network Pharmacology Approach for Medicinal Plants: Review and Assessment. Pharmaceuticals 2022, 15, 572. [Google Scholar] [CrossRef]
- Gertsch, J. Botanical drugs, synergy, and network pharmacology: Forth and back to intelligent mixtures. Planta Med. 2011, 77, 1086–1098. [Google Scholar] [CrossRef]
- Gong, K.; Guo, W.; Du, K.; Wang, F.; Li, M.; Guo, J. Mechanism of Huoluo Xiaoling Dan in the Treatment of Psoriasis Based on Network Pharmacology and Molecular Docking. Evid.-Based Complement. Altern. Med. 2022, 2022, 7053613. [Google Scholar] [CrossRef]
- Sripanidkulchai, B.; Promthep, K.; Tuntiyasawasdikul, S.; Tabboon, P.; Areemit, R. Supplementation of Kaempferia parviflora Extract Enhances Physical Fitness and Modulates Parameters of Heart Rate Variability in Adolescent Student-Athletes: A Randomized, Double-Blind, Placebo-Controlled Clinical Study. J. Diet. Suppl. 2022, 19, 149–167. [Google Scholar] [CrossRef]
- Okabe, Y.; Shimada, T.; Horikawa, T.; Kinoshita, K.; Koyama, K.; Ichinose, K.; Aburada, M.; Takahashi, K. Suppression of adipocyte hypertrophy by polymethoxyflavonoids isolated from Kaempferia parviflora. Phytomedicine 2014, 21, 800–806. [Google Scholar] [CrossRef]
- Tewtrakul, S.; Subhadhirasakul, S.; Kummee, S. Anti-allergic activity of compounds from Kaempferia parviflora. J. Ethnopharmacol. 2008, 116, 191–193. [Google Scholar] [CrossRef]
- Takuathung, M.N.; Potikanond, S.; Sookkhee, S.; Mungkornasawakul, P.; Jearanaikulvanich, T.; Chinda, K.; Wikan, N.; Nimlamool, W. Anti-psoriatic and anti-inflammatory effects of Kaempferia parviflora in keratinocytes and macrophage cells. Biomed. Pharmacother. 2021, 143, 112229. [Google Scholar] [CrossRef]
- Sutthanut, K.; Sripanidkulchai, B.; Yenjai, C.; Jay, M. Simultaneous identification and quantitation of 11 flavonoid constituents in Kaempferia parviflora by gas chromatography. J. Chromatogr. A 2007, 1143, 227–233. [Google Scholar] [CrossRef]
- Sae-Wong, C.; Matsuda, H.; Tewtrakul, S.; Tansakul, P.; Nakamura, S.; Nomura, Y.; Yoshikawa, M. Suppressive effects of methoxyflavonoids isolated from Kaempferia parviflora on inducible nitric oxide synthase (iNOS) expression in RAW 264.7 cells. J. Ethnopharmacol. 2011, 136, 488–495. [Google Scholar] [CrossRef]
- Chen, D.; Li, H.; Li, W.; Feng, S.; Deng, D. Kaempferia parviflora and its methoxyflavones: Chemistry and biological activities. Evid.-Based Complement. Altern. Med. 2018, 2018, 4057456. [Google Scholar] [CrossRef]
- Oliveros, J.C.; VENNY. An Interactive Tool for Comparing Lists with Venn Diagrams. 2007. Available online: http://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 1 November 2024).
- Li, L.; Lu, J.; Liu, J.; Wu, J.; Zhang, X.; Meng, Y.; Wu, X.; Tai, Z.; Zhu, Q.; Chen, Z. Immune cells in the epithelial immune microenvironment of psoriasis: Emerging therapeutic targets. Front. Immunol. 2023, 14, 1340677. [Google Scholar] [CrossRef]
- Wang, H.; Chen, W.; Lie, C.; Zhang, Y.; Li, J.; Meng, J.; Zhang, N. ARG1 and CXCL2 are potential biomarkers target for psoriasis patients. Mol. Pain 2022, 18, 17448069221128423. [Google Scholar] [CrossRef]
- Ru, J.; Li, P.; Wang, J.; Zhou, W.; Li, B.; Huang, C.; Li, P.; Guo, Z.; Tao, W.; Yang, Y.; et al. TCMSP: A database of systems pharmacology for drug discovery from herbal medicines. J. Cheminform. 2014, 6, 13. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, Y.; Cui, L.; Shi, Y.; Guo, C. Advances in the pathogenesis of psoriasis: From keratinocyte perspective. Cell Death Dis. 2022, 13, 81. [Google Scholar] [CrossRef]
- Hu, X.; Li, J.; Fu, M.; Zhao, X.; Wang, W. The JAK/STAT signaling pathway: From bench to clinic. Signal Transduct. Target. Ther. 2021, 6, 402. [Google Scholar] [CrossRef] [PubMed]
- Sundarrajan, S.; Nandakumar, M.P.; Prabhu, D.; Jeyaraman, J.; Arumugam, M. Conformational insights into the inhibitory mechanism of phyto-compounds against Src kinase family members implicated in psoriasis. J. Biomol. Struct. Dyn. 2020, 38, 1398–1414. [Google Scholar] [CrossRef] [PubMed]
- Mercurio, L.; Albanesi, C.; Madonna, S. Recent Updates on the Involvement of PI3K/AKT/mTOR Molecular Cascade in the Pathogenesis of Hyperproliferative Skin Disorders. Front. Med. 2021, 8, 665647. [Google Scholar] [CrossRef]
- Teng, Y.; Fan, Y.; Ma, J.; Lu, W.; Liu, N.; Chen, Y.; Pan, W.; Tao, X. The PI3K/Akt Pathway: Emerging Roles in Skin Homeostasis and a Group of Non-Malignant Skin Disorders. Cells 2021, 10, 1219. [Google Scholar] [CrossRef]
- Li, Y.; Wang, D.; Ge, H.; Güngör, C.; Gong, X.; Chen, Y. Cytoskeletal and Cytoskeleton-Associated Proteins: Key Regulators of Cancer Stem Cell Properties. Pharmaceuticals 2022, 15, 1369. [Google Scholar] [CrossRef]
- Ongchai, S.; Chiranthanut, N.; Tangyuenyong, S.; Viriyakhasem, N.; Kongdang, P. Kaempferia parviflora Extract Alleviated Rat Arthritis, Exerted Chondroprotective Properties In Vitro, and Reduced Expression of Genes Associated with Inflammatory Arthritis. Molecules 2021, 26, 1527. [Google Scholar] [CrossRef]
- Kongdang, P.; Jaitham, R.; Thonghoi, S.; Kuensaen, C.; Pradit, W.; Ongchai, S. Ethanolic extract of Kaempferia parviflora interrupts the mechanisms-associated rheumatoid arthritis in SW982 culture model via p38/STAT1 and STAT3 pathways. Phytomedicine 2019, 59, 152755. [Google Scholar] [CrossRef]
- Phung, H.M.; Lee, S.; Hong, S.; Lee, S.; Jung, K.; Kang, K.S. Protective Effect of Polymethoxyflavones Isolated from Kaempferia parviflora against TNF-α-Induced Human Dermal Fibroblast Damage. Antioxidants 2021, 10, 1609. [Google Scholar] [CrossRef]
- Kobayashi, H.; Suzuki, R.; Sato, K.; Ogami, T.; Tomozawa, H.; Tsubata, M.; Ichinose, K.; Aburada, M.; Ochiai, W.; Sugiyama, K.; et al. Effect of Kaempferia parviflora extract on knee osteoarthritis. J. Nat. Med. 2018, 72, 136–144. [Google Scholar] [CrossRef]
- Lee, S.; Jang, T.; Kim, K.H.; Kang, K.S. Improvement of Damage in Human Dermal Fibroblasts by 3,5,7-Trimethoxyflavone from Black Ginger (Kaempferia parviflora). Antioxidants 2022, 11, 425. [Google Scholar] [CrossRef] [PubMed]
- Tewtrakul, S.; Subhadhirasakul, S. Effects of compounds from Kaempferia parviflora on nitric oxide, prostaglandin E2 and tumor necrosis factor-alpha productions in RAW264.7 macrophage cells. J. Ethnopharmacol. 2008, 120, 81–84. [Google Scholar] [CrossRef]
- Horigome, S.; Yoshida, I.; Ito, S.; Inohana, S.; Fushimi, K.; Nagai, T.; Yamaguchi, A.; Fujita, K.; Satoyama, T.; Katsuda, S.-i.; et al. Inhibitory effects of Kaempferia parviflora extract on monocyte adhesion and cellular reactive oxygen species production in human umbilical vein endothelial cells. Eur. J. Nutr. 2017, 56, 949–964. [Google Scholar] [CrossRef]
- Temviriyanukul, P.; Chansawhang, A.; Karinchai, J.; Phochantachinda, S.; Buranasinsup, S.; Inthachat, W.; Pitchakarn, P.; Chantong, B. Kaempferia parviflora Extracts Protect Neural Stem Cells from Amyloid Peptide-Mediated Inflammation in Co-Culture Model with Microglia. Nutrients 2023, 15, 1098. [Google Scholar] [CrossRef] [PubMed]
- Klinngam, W.; Rungkamoltip, P.; Thongin, S.; Joothamongkhon, J.; Khumkhrong, P.; Khongkow, M.; Namdee, K.; Tepaamorndech, S.; Chaikul, P.; Kanlayavattanakul, M.; et al. Polymethoxyflavones from Kaempferia parviflora ameliorate skin aging in primary human dermal fibroblasts and ex vivo human skin. Biomed. Pharmacother. 2022, 145, 112461. [Google Scholar] [CrossRef]
- Chaisuwan, V.; Dajanta, K.; Srikaeo, K. Effects of extraction methods on antioxidants and methoxyflavones of Kaempferia parviflora. Food Res. 2022, 6, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Xu, M.; Hu, H.; Zhang, Q.; Yu, D.; Cai, M.; Geng, X.; Zhang, H.; Zhang, Y.; Guo, M. 5,7,4′-Trimethoxyflavone triggers cancer cell PD-L1 ubiquitin–proteasome degradation and facilitates antitumor immunity by targeting HRD1. MedComm 2024, 5, e611. [Google Scholar] [CrossRef]
- Wu, R.; Li, P.; Hao, B.; Fredimoses, M.; Ge, Y.; Zhou, Y.; Tang, L.; Li, Y.; Liu, H.; Janson, V. Design, synthesis, and biological evaluation of novel 5, 7, 4′-trimethoxyflavone sulfonamide-based derivatives as highly potent inhibitors of LRPPRC/STAT3/CDK1. Bioorg. Chem. 2024, 153, 107878. [Google Scholar] [CrossRef]
- Lee, M.H.; Han, A.R.; Jang, M.; Choi, H.K.; Lee, S.Y.; Kim, K.T.; Lim, T.G. Antiskin Inflammatory Activity of Black Ginger (Kaempferia parviflora) through Antioxidative Activity. Oxidative Med. Cell Longev. 2018, 2018, 5967150. [Google Scholar] [CrossRef]
- Tewtrakul, S.; Subhadhirasakul, S.; Karalai, C.; Ponglimanont, C.; Cheenpracha, S. Anti-inflammatory effects of compounds from Kaempferia parviflora and Boesenbergia pandurata. Food Chem. 2009, 115, 534–538. [Google Scholar] [CrossRef]
- Bocheńska, K.; Smolińska, E.; Moskot, M.; Jakóbkiewicz-Banecka, J.; Gabig-Cimińska, M. Models in the Research Process of Psoriasis. Int. J. Mol. Sci. 2017, 18, 2514. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, S.; Tagawa, T.; Awa, R.; Ogasawara, J.; Kuwahara, H.; Fukuhara, I. Polymethoxyflavone purified from Kaempferia parviflora reduces visceral fat in Japanese overweight individuals: A randomised, double-blind, placebo-controlled study. Food Funct. 2021, 12, 1603–1613. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Frisch, A. Gaussian 09W Reference; Gaussian Inc.: Wallingford, CT, USA, 2009; Volume 470, 25p. [Google Scholar]
- Hehre, W.J.; Stewart, R.F.; Pople, J.A. Self-consistent molecular-orbital methods. I. Use of Gaussian expansions of Slater-type atomic orbitals. J. Chem. Phys. 1969, 51, 2657–2664. [Google Scholar] [CrossRef]
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K. Improvements to the APBS biomolecular solvation software suite. Protein Sci. 2018, 27, 112–128. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Visualizer, D. Discovery Studio Visualizer, v21. 1.0. 20298; Accelrys Software Inc.: San Diego, CA, USA, 2005.
- Schrödinger, L. The PyMOL Molecular Graphics System, Version 2.5.4; Schrödinger, LLC.: New York, NY, USA, 2015.
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef] [PubMed]
- Porwal, A.; Kundu, G.C.; Bhagwat, G.; Butti, R. Polyherbal formulation Anoac-H suppresses the expression of RANTES and VEGF for the management of bleeding hemorrhoids and fistula. Mol. Med. Rep. 2021, 24, 736. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compounds | Molecular Formula | Lipinski Rules | ||||
|---|---|---|---|---|---|---|---|
| MW (<500) | HBA (<10) | HBD (5) | MlogP (<4.15) | TPSA (Å2) (<140) | |||
| 1 | 5-hydroxy-7-methoxyflavone | C16H12O4 | 268.26 | 4 | 1 | 1.33 | 59.67 |
| 2 | 5,7-dimethoxyflavone | C17H14O4 | 282.29 | 4 | 0 | 1.57 | 48.67 |
| 3 | 5-hydroxy-7,4′-dimethoxyflavone | C17H14O5 | 298.29 | 5 | 1 | 1.01 | 68.90 |
| 4 | 5,7,4′-trimethoxyflavone | C18H16O5 | 312.32 | 5 | 0 | 1.25 | 57.90 |
| 5 | 5,7,3′,4′-tetramethoxyflavone | C19H18O6 | 342.34 | 6 | 0 | 0.94 | 67.13 |
| 6 | 5-hydroxy-3,7-dimethoxyflavone | C17H14O5 | 298.29 | 5 | 1 | 1.01 | 68.90 |
| 7 | 3,5,7-trimethoxyflavone | C18H16O5 | 312.32 | 5 | 0 | 1.25 | 57.90 |
| 8 | 5-hydroxy-3,7,4′-trimethoxyflavone | C18H16O6 | 328.32 | 6 | 1 | 0.7 | 78.13 |
| 9 | 3,5,7,4′-tetramethoxyflavone | C19H18O6 | 342.34 | 6 | 0 | 0.94 | 67.13 |
| 10 | 5,3′-dihydroxy-3,7,4′-trimethoxyflavone | C18H16O7 | 344.32 | 7 | 2 | 0.17 | 98.36 |
| 11 | 5-hydroxy-3,7,3′,4′-tetramethoxyflavone | C19H18O7 | 358.34 | 7 | 1 | 0.4 | 87.36 |
| 12 | 3,5,7,3′,4′-pentamethoxyflavone | C20H20O7 | 372.37 | 7 | 0 | 0.63 | 76.36 |
| 13 | 3,5-dihydroxy-7,3′,4′-trimethoxyflavone | C18H16O7 | 344.32 | 7 | 2 | 0.17 | 98 36 |
| 14 | 5,4′-dihydroxy-7-methoxyflavone | C16H12O5 | 284.26 | 5 | 2 | 0.77 | 79.90 |
| 15 | 5-hydroxy-7,3′,4′-trimethoxyflavone | C18H16O6 | 328.32 | 6 | 1 | 0.7 | 78.13 |
| 16 | 4′-hydroxy-5,7-dimethoxyflavone | C17H14O5 | 298.29 | 5 | 1 | 1.01 | 68.90 |
| Proteins | Compounds | Binding Energy (kcal/mol) | Hydrogen Bonds |
|---|---|---|---|
| SRC (PDB: 2BDJ) | 5,7,4′-trimethoxyflavone | −5.71 | LYS39, MET85 |
| 3,5,7-trimethoxyflavone | −5.54 | MET85 | |
| 5-hydroxy-3,7,4′-trimethoxyflavone | −6.03 | LYS39, TYR84, MET85 | |
| 5-hydroxy-3,7,3′,4′-tetramethoxyflavone | −6.54 | LYS39, MET85 | |
| 3,5,7,3′,4′-pentamethoxyflavone | −6.40 | LYS39, MET85 | |
| AKT1 (PDB: 3O96) | 5,7,4′-trimethoxyflavone | −5.88 | LYS267, ASN52 |
| 3,5,7-trimethoxyflavone | −6.33 | LYS267 | |
| 5-hydroxy-3,7,4′-trimethoxyflavone | −5.85 | LYS267 | |
| 5-hydroxy-3,7,3′,4′-tetramethoxyflavone | −6.02 | LYS267 | |
| 3,5,7,3′,4′-pentamethoxyflavone | −5.83 | LYS267 | |
| PIK3R1 (PDB: 5Fi4) | 5,7,4′-trimethoxyflavone | −6.11 | VAL850 |
| 3,5,7-trimethoxyflavone | −5.82 | VAL850 | |
| 5-hydroxy-3,7,4′-trimethoxyflavone | −6.00 | VAL850 | |
| 5-hydroxy-3,7,3′,4′-tetramethoxyflavone | −6.33 | LYS801, VAL850, SER853, ASP932 | |
| 3,5,7,3′,4′-pentamethoxyflavone | −6.66 | LYS801, VAL850, ASP932 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sakuludomkan, C.; Khowsathit, J.; Thippraphan, P.; Koonrungsesomboon, N.; Takuathung, M.N.; Taychaworaditsakul, W. Network Pharmacology and Molecular Docking-Based Approach to Explore Potential Bioactive Compounds from Kaempferia parviflora on Chemokine Signaling Pathways in the Treatment of Psoriasis Disease. Int. J. Mol. Sci. 2025, 26, 5243. https://doi.org/10.3390/ijms26115243
Sakuludomkan C, Khowsathit J, Thippraphan P, Koonrungsesomboon N, Takuathung MN, Taychaworaditsakul W. Network Pharmacology and Molecular Docking-Based Approach to Explore Potential Bioactive Compounds from Kaempferia parviflora on Chemokine Signaling Pathways in the Treatment of Psoriasis Disease. International Journal of Molecular Sciences. 2025; 26(11):5243. https://doi.org/10.3390/ijms26115243
Chicago/Turabian StyleSakuludomkan, Chotiwit, Jittasak Khowsathit, Pilaiporn Thippraphan, Nut Koonrungsesomboon, Mingkwan Na Takuathung, and Weerakit Taychaworaditsakul. 2025. "Network Pharmacology and Molecular Docking-Based Approach to Explore Potential Bioactive Compounds from Kaempferia parviflora on Chemokine Signaling Pathways in the Treatment of Psoriasis Disease" International Journal of Molecular Sciences 26, no. 11: 5243. https://doi.org/10.3390/ijms26115243
APA StyleSakuludomkan, C., Khowsathit, J., Thippraphan, P., Koonrungsesomboon, N., Takuathung, M. N., & Taychaworaditsakul, W. (2025). Network Pharmacology and Molecular Docking-Based Approach to Explore Potential Bioactive Compounds from Kaempferia parviflora on Chemokine Signaling Pathways in the Treatment of Psoriasis Disease. International Journal of Molecular Sciences, 26(11), 5243. https://doi.org/10.3390/ijms26115243