
TPP-Based Nanovesicles Kill MDR Neuroblastoma Cells and Induce Moderate ROS Increase, While Exerting Low Toxicity Towards Primary Cell Cultures: An In Vitro Study

 ,
,  ,
,  ,
,  , ,
, ,  , ,
, ,  and
and 
Abstract

1. Introduction
2. Results and Discussion
2.1. 1,1-(1,12-Dodecanediyl)-bis-[1,1,1]-triphenyl-phosphonium Di-Bromide (BPPB)
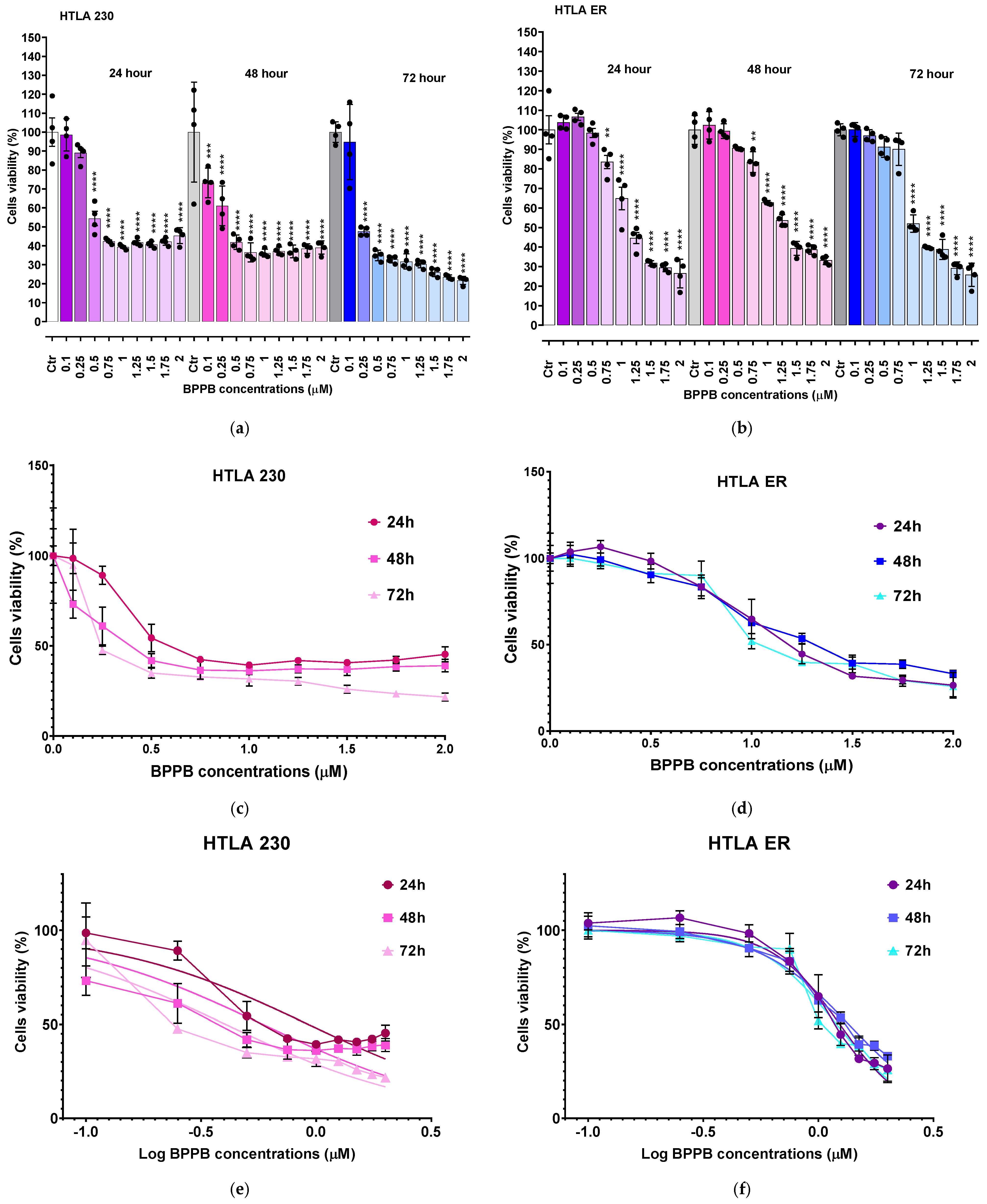
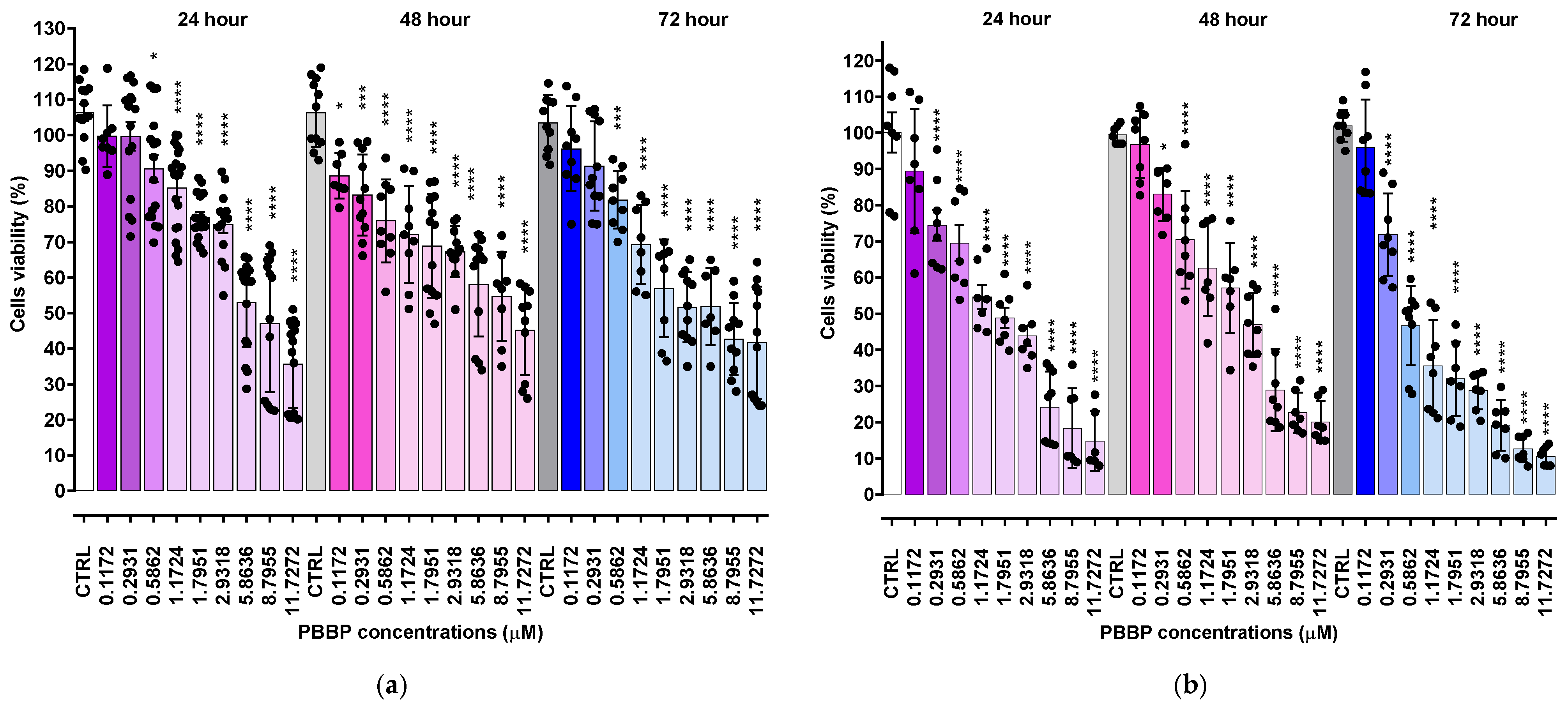
2.2. Cytotoxic Effects of BPPB Towards HTLA 230 and HTLA ER NB Cells
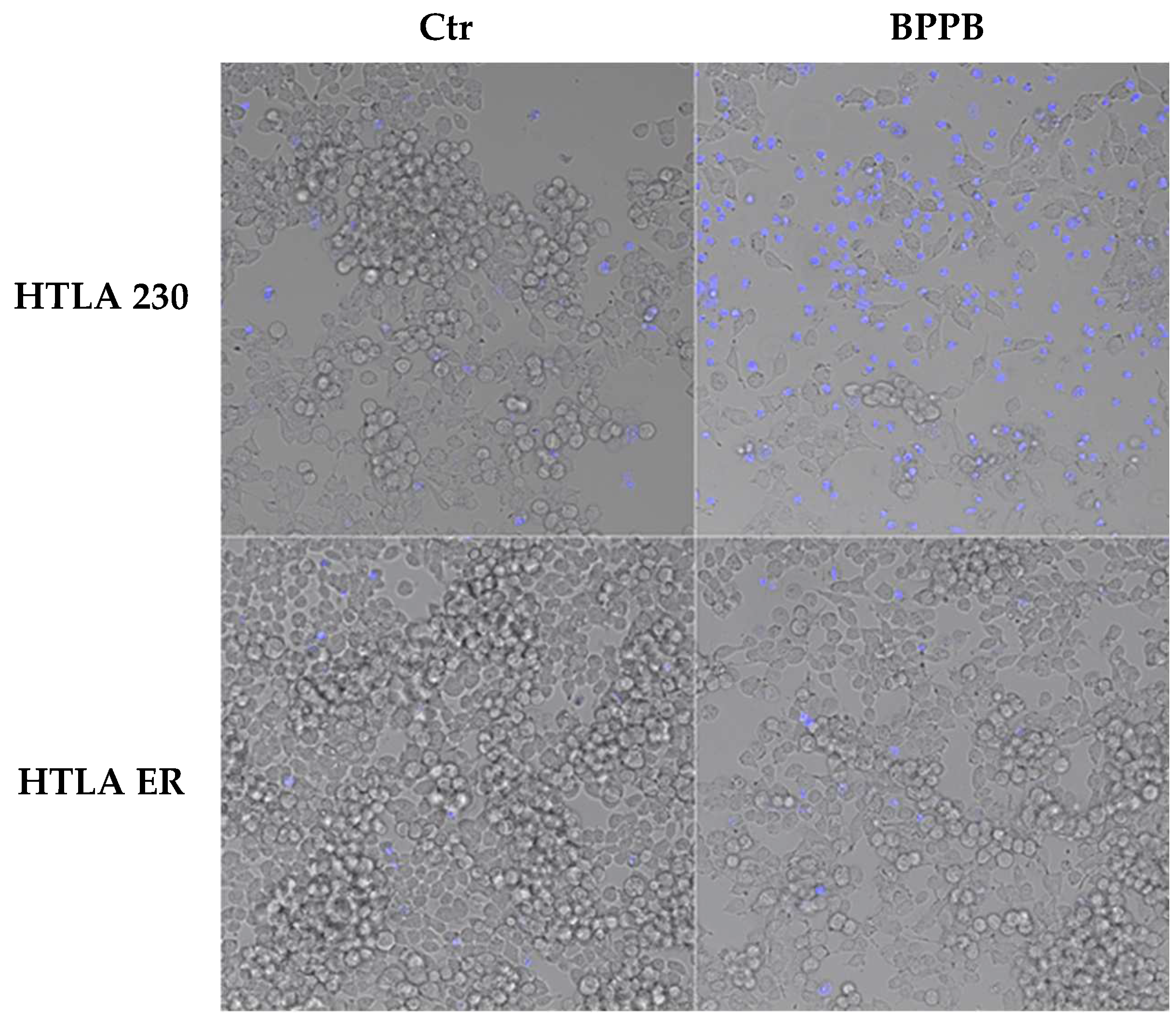
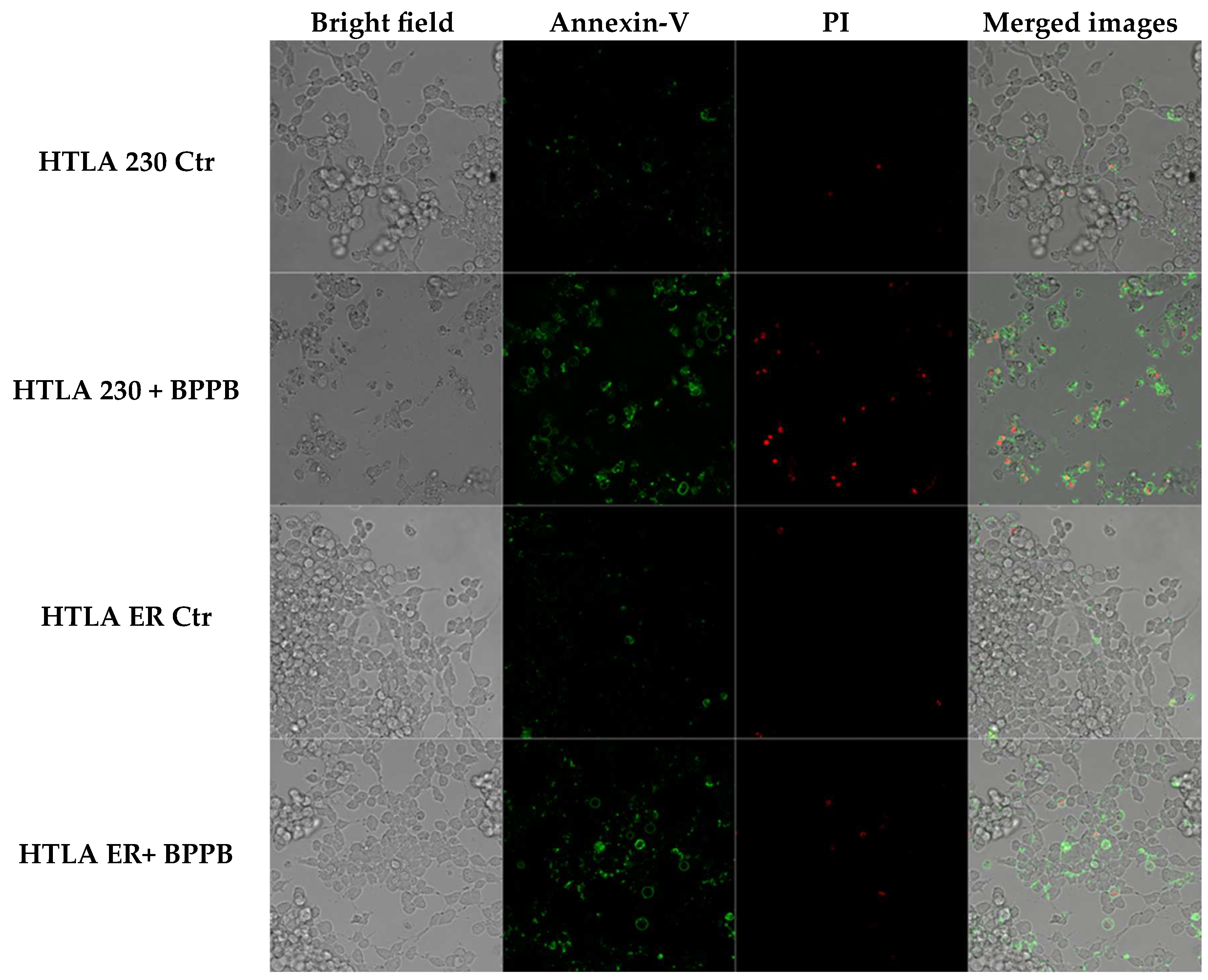
2.2.1. BPPB Induces Early and Late Apoptosis in Neuroblastoma Cells
2.2.2. Correlation Study
Correlation Between BPPB Cytotoxic Effects and BPPB Concentrations
Correlation Between BPPB Cytotoxic Effects and Exposure Time
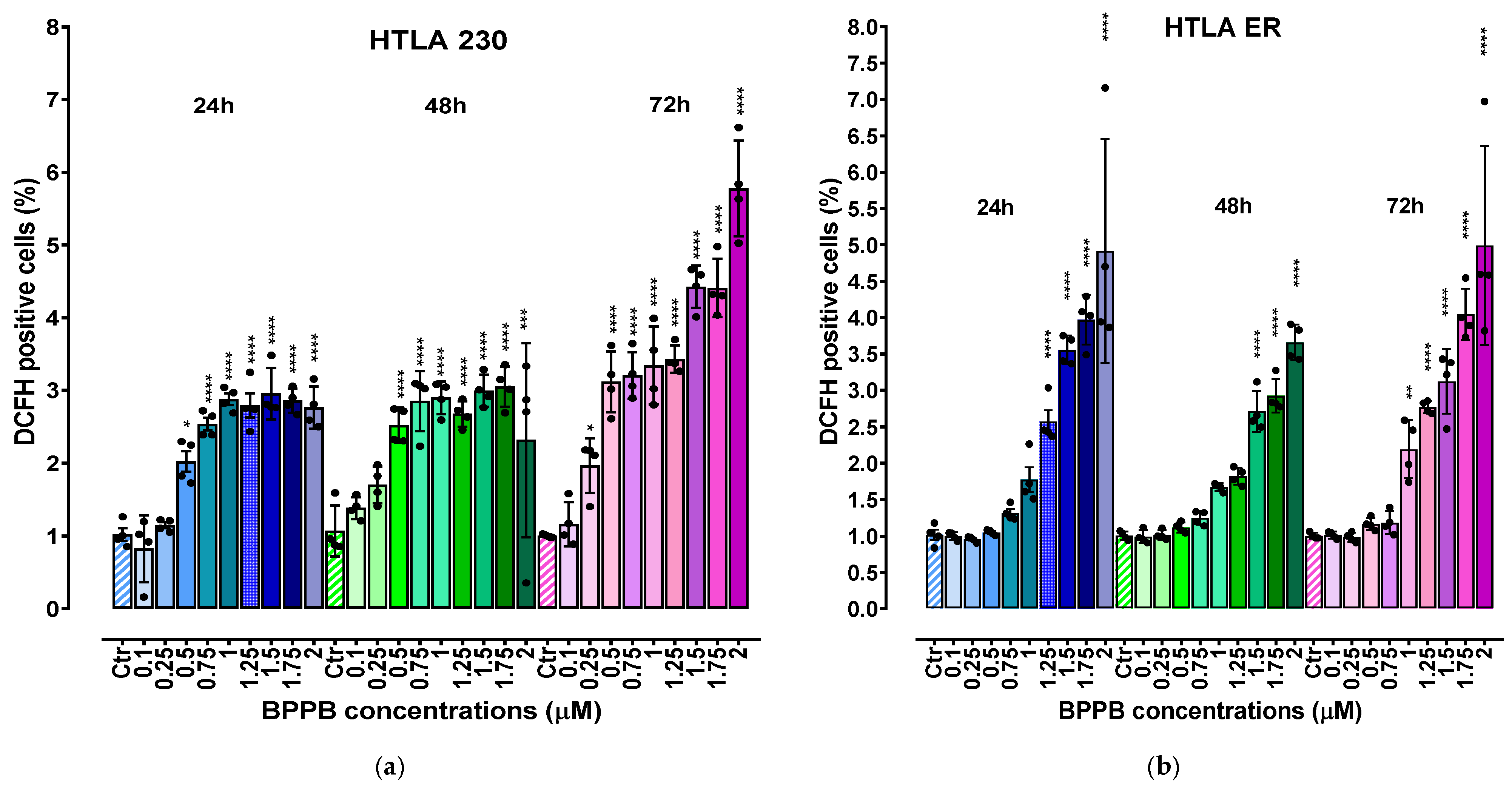
2.3. BPPB Induces ROS Overproduction in HTLA 230 and HTLA ER Cells: Concentration- and Time-Dependent Experiments
2.3.1. Correlation Between ROS Overproduction and BPPB Concentrations
2.3.2. Correlation Between ROS Overproduction and Exposure Time
2.3.3. Correlation Between BPPB Cytotoxic Effects (% Cell Viability) and BPPB-Induced ROS Production
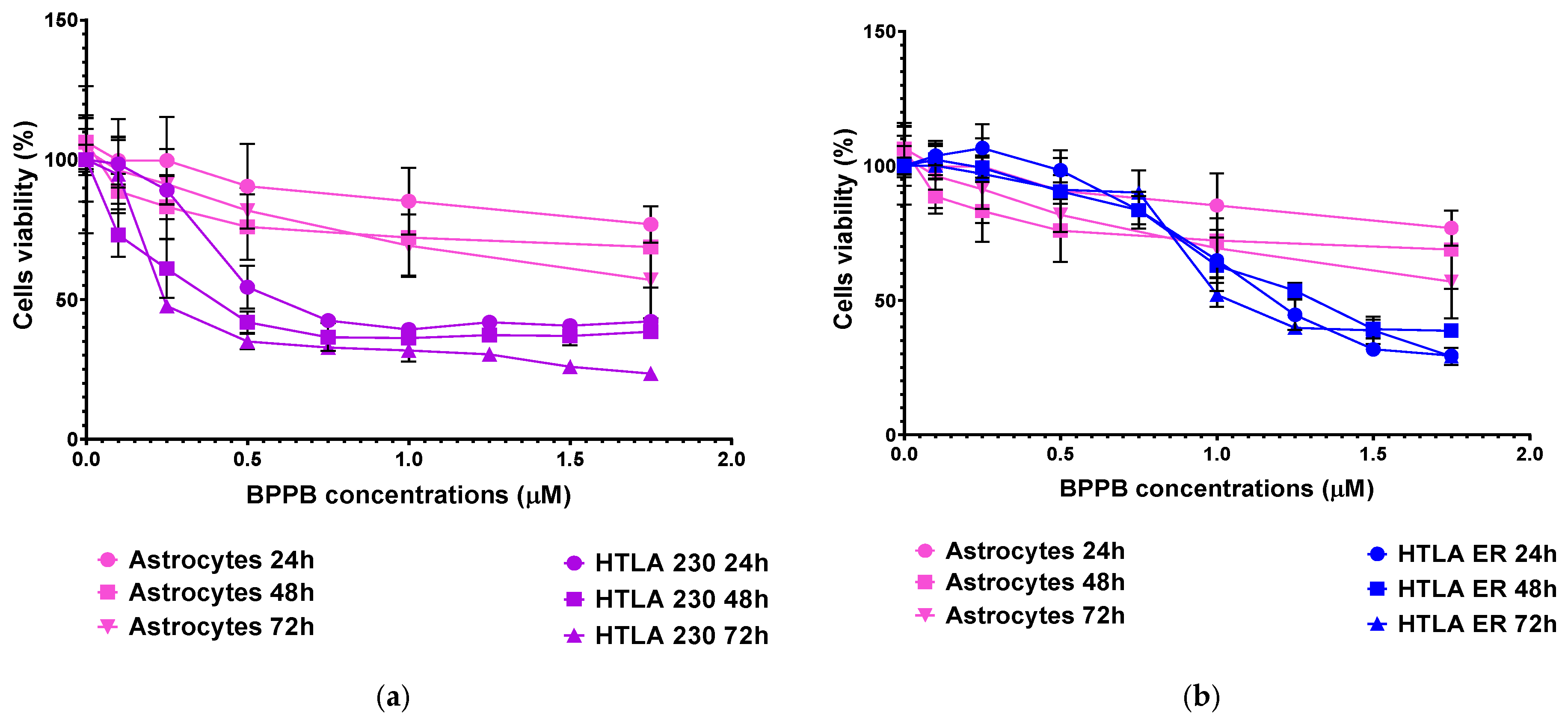
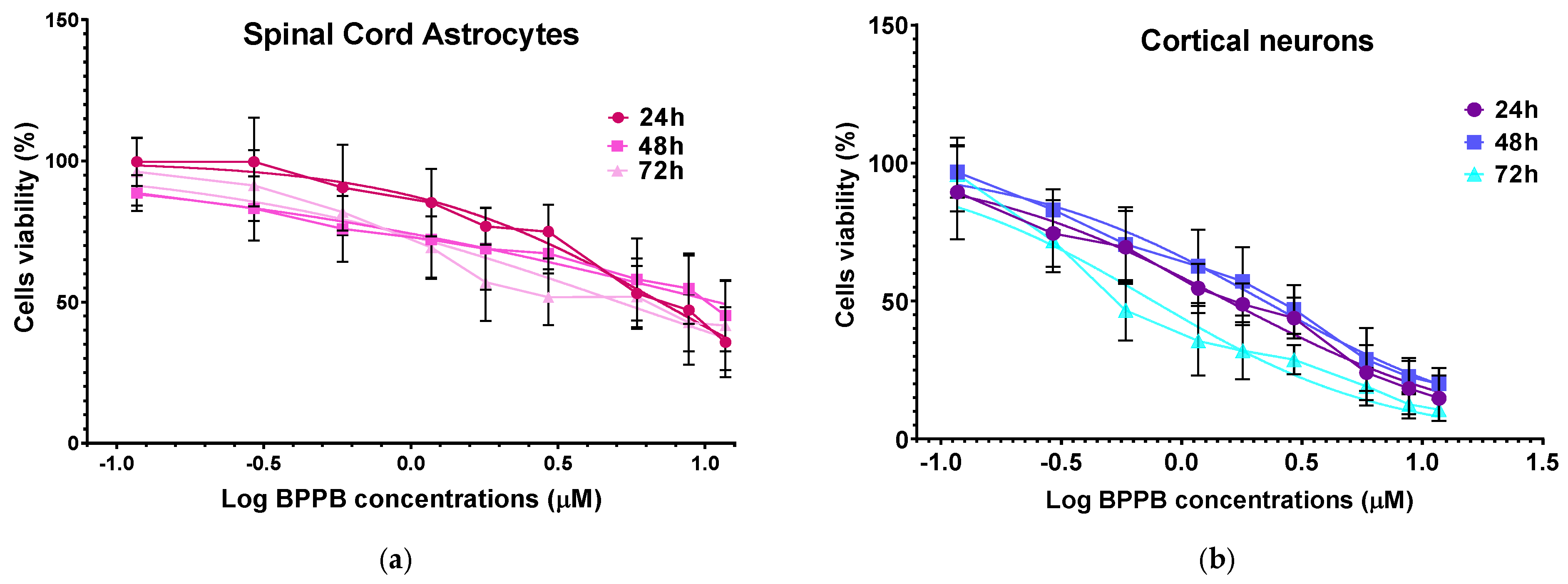
2.4. Concentration- and Time-Dependent Cytotoxic Effects of BPPB on Astrocyte and Neuron Primary Cell Cultures
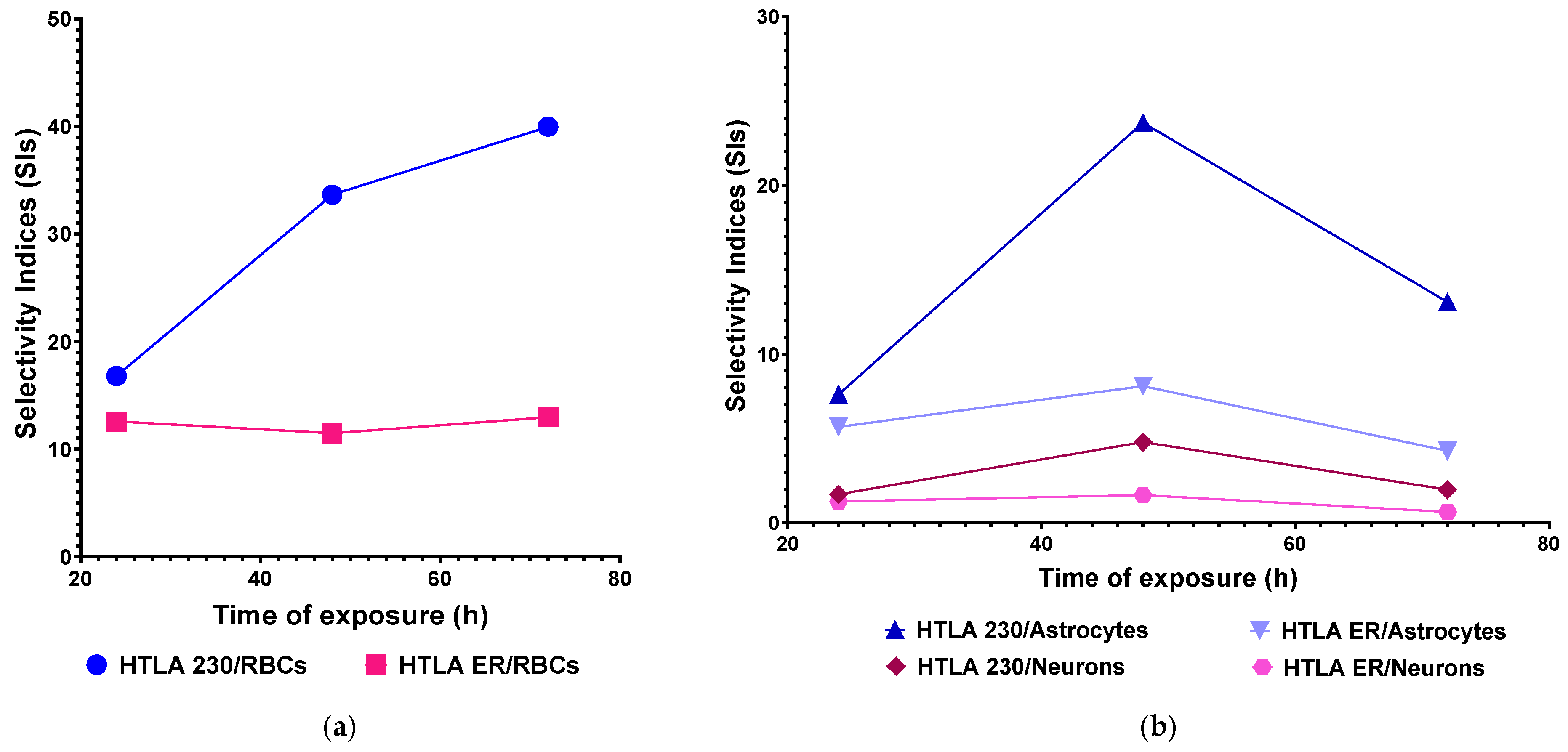
2.5. Selectivity Index
2.6. Correlation Between BPPB Cytotoxic Effects and BPPB Concentrations or Exposure Time on Astrocytes and Neuron Primary Cell Cultures
3. Materials and Methods
3.1. Chemicals and Instruments
3.2. In Vitro BPPB Cytotoxicity Evaluation on NB Cells
3.2.1. Cells and Culture Conditions
3.2.2. Treatments
3.2.3. Cell Viability Assay
3.2.4. DAPI Staining
3.2.5. Annexin-V and Propidium Iodide Staining
3.3. Detection of Hydrogen Peroxide (H2O2) Production
3.4. In Vitro Dose- and Time-Dependent Cytotoxicity of BPPB Against Spinal Cord Astrocytes and Cortical Neurons’ Primary Cell Cultures
3.4.1. Spinal Cord Astrocyte Primary Cell Cultures
3.4.2. Cortical Neuron Primary Cell Cultures
3.4.3. MTT Cytotoxicity Assay
3.5. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sainero-Alcolado, L.; Sjöberg Bexelius, T.; Santopolo, G.; Yuan, Y.; Liaño-Pons, J.; Arsenian-Henriksson, M. Defining Neuroblastoma: From Origin to Precision Medicine. Neuro-Oncology 2024, 26, 2174–2192. [Google Scholar] [CrossRef] [PubMed]
- Valter, K.; Zhivotovsky, B.; Gogvadze, V. Cell Death-Based Treatment of Neuroblastoma. Cell Death Dis. 2018, 9, 113. [Google Scholar] [CrossRef]
- Shusterman, S.; George, R.E. Neuroblastoma. In Oncology of Infancy and Childhood; Elsevier: Amsterdam, The Netherlands, 2009; pp. 507–540. [Google Scholar]
- Maris, J.M.; Mosse, Y.P.; Bradfield, J.P.; Hou, C.; Monni, S.; Scott, R.H.; Asgharzadeh, S.; Attiyeh, E.F.; Diskin, S.J.; Laudenslager, M.; et al. Chromosome 6p22 Locus Associated with Clinically Aggressive Neuroblastoma. N. Engl. J. Med. 2008, 358, 2585–2593. [Google Scholar] [CrossRef] [PubMed]
- Zafar, A.; Wang, W.; Liu, G.; Wang, X.; Xian, W.; McKeon, F.; Foster, J.; Zhou, J.; Zhang, R. Molecular Targeting Therapies for Neuroblastoma: Progress and Challenges. Med. Res. Rev. 2021, 41, 961–1021. [Google Scholar] [CrossRef] [PubMed]
- Bhoopathi, P.; Mannangatti, P.; Emdad, L.; Das, S.K.; Fisher, P.B. The Quest to Develop an Effective Therapy for Neuroblastoma. J. Cell. Physiol. 2021, 236, 7775–7791. [Google Scholar] [CrossRef]
- Speleman, F.; Park, J.R.; Henderson, T.O. Neuroblastoma: A Tough Nut to Crack. Am. Soc. Clin. Oncol. Educ. Book 2016, 36, e548–e557. [Google Scholar] [CrossRef]
- Li, Y.J.; Lei, Y.H.; Yao, N.; Wang, C.R.; Hu, N.; Ye, W.C.; Zhang, D.M.; Chen, Z.S. Autophagy and Multidrug Resistance in Cancer. Chin. J. Cancer 2017, 36, 52. [Google Scholar] [CrossRef]
- Brusnakov, M.; Golovchenko, O.; Velihina, Y.; Liavynets, O.; Zhirnov, V.; Brovarets, V. Evaluation of Anticancer Activity of 1,3-Oxazol-4-ylphosphonium Salts in Vitro. ChemMedChem 2022, 17, e202200319. [Google Scholar] [CrossRef]
- Gerges, A.; Canning, U. Neuroblastoma and Its Target Therapies: A Medicinal Chemistry Review. ChemMedChem 2024, 19, e202300535. [Google Scholar] [CrossRef]
- Alfei, S.; Giannoni, P.; Signorello, M.G.; Torazza, C.; Zuccari, G.; Athanassopoulos, C.M.; Domenicotti, C.; Marengo, B. The Remarkable and Selective In Vitro Cytotoxicity of Synthesized Bola-Amphiphilic Nanovesicles on Etoposide-Sensitive and -Resistant Neuroblastoma Cells. Nanomaterials 2024, 14, 1505. [Google Scholar] [CrossRef]
- Alfei, S.; Zuccari, G.; Athanassopoulos, C.M.; Domenicotti, C.; Marengo, B. Strongly ROS-Correlated, Time-Dependent, and Selective Antiproliferative Effects of Synthesized Nano Vesicles on BRAF Mutant Melanoma Cells and Their Hyaluronic Acid-Based Hydrogel Formulation. Int. J. Mol. Sci. 2024, 25, 10071. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Torazza, C.; Bacchetti, F.; Signorello, M.G.; Passalacqua, M.; Domenicotti, C.; Marengo, B. Tri-Phenyl-Phosphonium-Based Nano Vesicles: A New In Vitro Nanomolar-Active Weapon to Eradicate PLX-Resistant Melanoma Cells. Int. J. Mol. Sci. 2025, 26, 3227. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Zuccari, G.; Bacchetti, F.; Torazza, C.; Milanese, M.; Siciliano, C.; Athanassopoulos, C.M.; Piatti, G.; Schito, A.M. Synthesized Bis-Triphenyl Phosphonium-Based Nano Vesicles Have Potent and Selective Antibacterial Effects on Several Clinically Relevant Superbugs. Nanomaterials 2024, 14, 1351. [Google Scholar] [CrossRef]
- Malich, G.; Markovic, B.; Winder, C. The Sensitivity and Specificity of the MTS Tetrazolium Assay for Detecting the in Vitro Cytotoxicity of 20 Chemicals Using Human Cell Lines. Toxicology 1997, 124, 179–192. [Google Scholar] [CrossRef] [PubMed]
- Colla, R.; Izzotti, A.; De Ciucis, C.; Fenoglio, D.; Ravera, S.; Speciale, A.; Ricciarelli, R.; Furfaro, A.L.; Pulliero, A.; Passalacqua, M.; et al. Glutathione-Mediated Antioxidant Response and Aerobic Metabolism: Two Crucial Factors Involved in Determining the Multi-Drug Resistance of High-Risk Neuroblastoma. Oncotarget 2016, 7, 70715–70737. [Google Scholar] [CrossRef]
- Almutairi, B.; Ali, D.; Alyami, N.; Alothman, N.S.; Alakhtani, S.; Alarifi, S. Tantalum Doped TiO2 Nanoparticles Induced Cytotoxicity and DNA Damage through ROS Generation in Human Neuroblastoma Cells. J. King Saud. Univ.-Sci. 2021, 33, 101546. [Google Scholar] [CrossRef]
- Sambale, F.; Wagner, S.; Stahl, F.; Khaydarov, R.R.; Scheper, T.; Bahnemann, D. Investigations of the Toxic Effect of Silver Nanoparticles on Mammalian Cell Lines. J. Nanomater. 2015, 2015, 136765. [Google Scholar] [CrossRef]
- Haase, A.; Mantion, A.; Graf, P.; Plendl, J.; Thuenemann, A.F.; Meier, W.; Taubert, A.; Luch, A. A Novel Type of Silver Nanoparticles and Their Advantages in Toxicity Testing in Cell Culture Systems. Arch. Toxicol. 2012, 86. [Google Scholar] [CrossRef]
- Rosa, A.; Pollastro, F.; Sogos, V.; Piras, F. Comparative Evaluation of Cytotoxic and Apoptotic Effects of Natural Compounds in SH-SY5Y Neuroblastoma Cells in Relation to Their Physicochemical Properties. Molecules 2025, 30, 1742. [Google Scholar] [CrossRef]
- Skrzypczak, N.; Pyta, K.; Ruszkowski, P.; Mikołajczak, P.; Kucińska, M.; Murias, M.; Gdaniec, M.; Bartl, F.; Przybylski, P. Anticancer Activity and Toxicity of New Quaternary Ammonium Geldanamycin Derivative Salts and Their Mixtures with Potentiators. J. Enzyme Inhib. Med. Chem. 2021, 36, 1898–1904. [Google Scholar] [CrossRef]
- Nene, L.C.; Magadla, A.; Nyokong, T. Enhanced Mitochondria Destruction on MCF-7 and HeLa Cell Lines in Vitro Using Triphenyl-Phosphonium-Labelled Phthalocyanines in Ultrasound-Assisted Photodynamic Therapy Activity. J. Photochem. Photobiol. B 2022, 235, 112553. [Google Scholar] [CrossRef] [PubMed]
- Koshibu-Koizumi, J.; Akazawa, M.; Iwamoto, T.; Takasaki, M.; Mizuno, F.; Kobayashi, R.; Abe, A.; Tomoda, A.; Hamatake, M.; Ishida, R. Antitumor Activity of a Phenoxazine Compound, 2-Amino-4, 4α-Dihydro-4α,7-Dimethyl-3H-Phenoxazine-3-One against Human B Cell and T Cell Lymphoblastoid Cell Lines: Induction of Mixed Types of Cell Death, Apoptosis, and Necrosis. J. Cancer Res. Clin. Oncol. 2002, 128, 363–368. [Google Scholar] [CrossRef]
- Marengo, B.; De Ciucis, C.G.; Ricciarelli, R.; Furfaro, A.L.; Colla, R.; Canepa, E.; Traverso, N.; Marinari, U.M.; Pronzato, M.A.; Domenicotti, C. P38MAPK Inhibition: A New Combined Approach to Reduce Neuroblastoma Resistance under Etoposide Treatment. Cell Death Dis. 2013, 4, e589. [Google Scholar] [CrossRef]
- Nazarparvar-Noshadi, M.; Ezzati Nazhad Dolatabadi, J.; Rasoulzadeh, Y.; Mohammadian, Y.; Shanehbandi, D. Apoptosis and DNA Damage Induced by Silica Nanoparticles and Formaldehyde in Human Lung Epithelial Cells. Environ. Sci. Pollut. Res. 2020, 27, 18592–18601. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Lu, B.; Gan, J.; Wang, S.; Jiang, X.; Li, H. Apoptosis Detection: A Purpose-Dependent Approach Selection. Cell Cycle 2021, 20, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Crowley, L.C.; Marfell, B.J.; Scott, A.P.; Waterhouse, N.J. Quantitation of Apoptosis and Necrosis by Annexin V Binding, Propidium Iodide Uptake, and Flow Cytometry. Cold Spring Harb. Protoc. 2016, 2016, pdb–prot087288. [Google Scholar] [CrossRef]
- Wlodkowic, D.; Telford, W.; Skommer, J.; Darzynkiewicz, Z. Apoptosis and Beyond: Cytometry in Studies of Programmed Cell Death. Methods Cell Biol. 2011, 103, 55–98. [Google Scholar]
- Sawai, H.; Domae, N. Discrimination between Primary Necrosis and Apoptosis by Necrostatin-1 in Annexin V-Positive/Propidium Iodide-Negative Cells. Biochem. Biophys. Res. Commun. 2011, 411, 569–573. [Google Scholar] [CrossRef]
- Kumar, R.; Saneja, A.; Panda, A.K. An Annexin V-FITC—Propidium Iodide-Based Method for Detecting Apoptosis in a Non-Small Cell Lung Cancer Cell Line. In Methods in Molecular Biology; Springers: Berlin/Heidelberg, Germany, 2021; Volume 2279. [Google Scholar]
- Liu, T.; Zhu, W.; Yang, X.; Chen, L.; Yang, R.; Hua, Z.; Li, G. Detection of Apoptosis Based on the Interaction between Annexin V and Phosphatidylserine. Anal. Chem. 2009, 81, 2410–2413. [Google Scholar] [CrossRef]
- Wallberg, F.; Tenev, T.; Meier, P. Analysis of Apoptosis and Necroptosis by Fluorescence-Activated Cell Sorting. Cold Spring Harb. Protoc. 2016, 2016, pdb–prot087387. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics; McGraw-Hill Book Company: New York, NY, USA, 1960. [Google Scholar]
- Zach Bobbitt Statology. What Is a Good R-Squared Value? Available online: https://www.statology.org/good-r-squared-value/#:~:text=For%20example%2C%20in%20scientific%20studies%2C%20the%20R-squared%20may,if%20there%20is%20extreme%20variability%20in%20the%20dataset (accessed on 22 January 2025).
- Valutazione Della Bontà Di Adattamento per Modelli Non Lineari: Metodi, Parametri e Considerazioni Pratiche. Available online: https://www.geeksforgeeks.org/evaluating-goodness-of-fit-for-nonlinear-models-methods-metrics-and-practical-considerations/ (accessed on 22 January 2025).
- Ceccacci, F.; Sennato, S.; Rossi, E.; Proroga, R.; Sarti, S.; Diociaiuti, M.; Casciardi, S.; Mussi, V.; Ciogli, A.; Bordi, F.; et al. Aggregation Behaviour of Triphenylphosphonium Bolaamphiphiles. J. Colloid. Interface Sci. 2018, 531, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Marengo, B.; Domenicotti, C. Polyester-Based Dendrimer Nanoparticles Combined with Etoposide Have an Improved Cytotoxic and Pro-Oxidant Effect on Human Neuroblastoma Cells. Antioxidants 2020, 9, 50. [Google Scholar] [CrossRef] [PubMed]
- Kciuk, M.; Gielecińska, A.; Mujwar, S.; Kołat, D.; Kałuzińska-Kołat, Ż.; Celik, I.; Kontek, R. Doxorubicin—An Agent with Multiple Mechanisms of Anticancer Activity. Cells 2023, 12, 659. [Google Scholar] [CrossRef]
- MOHIUDDIN, M.; KASAHARA, K. Cisplatin Activates the Growth Inhibitory Signaling Pathways by Enhancing the Production of Reactive Oxygen Species in Non-Small Cell Lung Cancer Carrying an EGFR Exon 19 Deletion. Cancer Genom.-Proteom. 2021, 18, 471–486. [Google Scholar] [CrossRef]
- Trachootham, D.; Lu, W.; Ogasawara, M.A.; Valle, N.R.D.; Huang, P. Redox Regulation of Cell Survival. Antioxid. Redox Signal. 2008, 10, 1343–1374. [Google Scholar] [CrossRef]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of Reactive Oxygen Species (ROS) in Apoptosis Induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef]
- Ruijter, N.; Soeteman-Hernández, L.G.; Carrière, M.; Boyles, M.; McLean, P.; Catalán, J.; Katsumiti, A.; Cabellos, J.; Delpivo, C.; Sánchez Jiménez, A.; et al. The State of the Art and Challenges of In Vitro Methods for Human Hazard Assessment of Nanomaterials in the Context of Safe-by-Design. Nanomaterials 2023, 13, 472. [Google Scholar] [CrossRef]
- Ruijter, N.; van der Zee, M.; Katsumiti, A.; Boyles, M.; Cassee, F.R.; Braakhuis, H. Improving the Dichloro-Dihydro-Fluorescein (DCFH) Assay for the Assessment of Intracellular Reactive Oxygen Species Formation by Nanomaterials. NanoImpact 2024, 35, 100521. [Google Scholar] [CrossRef]
- Zeng, Y.; Cao, Y.; Ren, S.; Zhang, C.; Liu, J.; Liu, K.; Wang, Y.; Chen, H.; Zhou, F.; Yang, X.; et al. Responsive ROS-Augmented Prodrug Hybridization Nanoassemblies for Multidimensionally Synergitic Treatment of Hepatocellular Carcinoma in Cascade Assaults. Adv. Sci. 2025, 2501420. [Google Scholar] [CrossRef]
- Bauer, D.; Werth, F.; Nguyen, H.A.; Kiecker, F.; Eberle, J. Critical Role of Reactive Oxygen Species (ROS) for Synergistic Enhancement of Apoptosis by Vemurafenib and the Potassium Channel Inhibitor TRAM-34 in Melanoma Cells. Cell Death Dis. 2017, 8, e2594. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Xiong, J.; Wang, Y.; Liang, G.; Xu, Y.; Lin, F. Quantitative Prediction of Hemolytic Toxicity for Small Molecules and Their Potential Hemolytic Fragments by Machine Learning and Recursive Fragmentation Methods. J. Chem. Inf. Model. 2020, 60, 3231–3245. [Google Scholar] [CrossRef] [PubMed]
- Nunes, B.; Cagide, F.; Fernandes, C.; Borges, A.; Borges, F.; Simões, M. Efficacy of Novel Quaternary Ammonium and Phosphonium Salts Differing in Cation Type and Alkyl Chain Length against Antibiotic-Resistant Staphylococcus Aureus. Int. J. Mol. Sci. 2023, 25, 504. [Google Scholar] [CrossRef]
- Valenti, G.E.; Alfei, S.; Caviglia, D.; Domenicotti, C.; Marengo, B. Antimicrobial Peptides and Cationic Nanoparticles: A Broad-Spectrum Weapon to Fight Multi-Drug Resistance Not Only in Bacteria. Int. J. Mol. Sci. 2022, 23, 6108. [Google Scholar] [CrossRef] [PubMed]
- Lange, S.C.; Bak, L.K.; Waagepetersen, H.S.; Schousboe, A.; Norenberg, M.D. Primary Cultures of Astrocytes: Their Value in Understanding Astrocytes in Health and Disease. Neurochem. Res. 2012, 37, 2569–2588. [Google Scholar] [CrossRef]
- Akere, M.T.; Zajac, K.K.; Bretz, J.D.; Madhavaram, A.R.; Horton, A.C.; Schiefer, I.T. Real-Time Analysis of Neuronal Cell Cultures for CNS Drug Discovery. Brain Sci. 2024, 14, 770. [Google Scholar] [CrossRef]
- Bexiga, M.G.; Varela, J.A.; Wang, F.; Fenaroli, F.; Salvati, A.; Lynch, I.; Simpson, J.C.; Dawson, K.A. Cationic Nanoparticles Induce Caspase 3-, 7- and 9-Mediated Cytotoxicity in a Human Astrocytoma Cell Line. Nanotoxicology 2011, 5, 557–567. [Google Scholar] [CrossRef]
- Navarro, S.; Boix, E.; Cuchillo, C.M.; Nogués, M.V. Eosinophil-Induced Neurotoxicity: The Role of Eosinophil Cationic Protein/RNase 3. J. Neuroimmunol. 2010, 227, 60–70. [Google Scholar] [CrossRef]
- Aoshima, Y.; Hokama, R.; Sou, K.; Sarker, S.R.; Iida, K.; Nakamura, H.; Inoue, T.; Takeoka, S. Cationic Amino Acid Based Lipids as Effective Nonviral Gene Delivery Vectors for Primary Cultured Neurons. ACS Chem. Neurosci. 2013, 4, 1514–1519. [Google Scholar] [CrossRef]
- Pei, Y.; Peng, J.; Behl, M.; Sipes, N.S.; Shockley, K.R.; Rao, M.S.; Tice, R.R.; Zeng, X. Comparative Neurotoxicity Screening in Human IPSC-Derived Neural Stem Cells, Neurons and Astrocytes. Brain Res. 2016, 1638, 57–73. [Google Scholar] [CrossRef]
- Vidal, F.; Vásquez, P.; Cayumán, F.; Díaz, C.; Fuentealba, J.; Aguayo, L.; Yévenes, G.; Alderete, J.; Guzmán, L. Prevention of Synaptic Alterations and Neurotoxic Effects of PAMAM Dendrimers by Surface Functionalization. Nanomaterials 2017, 8, 7. [Google Scholar] [CrossRef]
- Albertazzi, L.; Gherardini, L.; Brondi, M.; Sulis Sato, S.; Bifone, A.; Pizzorusso, T.; Ratto, G.M.; Bardi, G. In Vivo Distribution and Toxicity of PAMAM Dendrimers in the Central Nervous System Depend on Their Surface Chemistry. Mol. Pharm. 2013, 10, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of Cell Viability by the MTT Assay. Cold Spring Harb. Protoc. 2018, 2018, pdb–prot095505. [Google Scholar] [CrossRef]
- Borges, J.M.P.; de Jesus, L.B.; dos Santos Souza, C.; da Silva, V.D.A.; Costa, S.L.; de Fátima Dias Costa, M.; El-Bachá, R.S. Astrocyte Reaction to Catechol-Induced Cytotoxicity Relies on the Contact with Microglia Before Isolation. Neurotox. Res. 2022, 40, 973–994. [Google Scholar] [CrossRef] [PubMed]
- Krzywik, J.; Mozga, W.; Aminpour, M.; Janczak, J.; Maj, E.; Wietrzyk, J.; Tuszyński, J.A.; Huczyński, A. Synthesis, Antiproliferative Activity and Molecular Docking Studies of Novel Doubly Modified Colchicine Amides and Sulfonamides as Anticancer Agents. Molecules 2020, 25, 1789. [Google Scholar] [CrossRef]
- Peña-Morán, O.; Villarreal, M.; Álvarez-Berber, L.; Meneses-Acosta, A.; Rodríguez-López, V. Cytotoxicity, Post-Treatment Recovery, and Selectivity Analysis of Naturally Occurring Podophyllotoxins from Bursera Fagaroides Var. Fagaroides on Breast Cancer Cell Lines. Molecules 2016, 21, 1013. [Google Scholar] [CrossRef]
- Chandrasekhar, B.; Gor, R.; Ramalingam, S.; Thiagarajan, A.; Sohn, H.; Madhavan, T. Repurposing FDA-Approved Compounds to Target JAK2 for Colon Cancer Treatment. Discov. Oncol. 2024, 15, 226. [Google Scholar] [CrossRef] [PubMed]
- Gürer, G.; Gursoy-Ozdemir, Y.; Erdemli, E.; Can, A.; Dalkara, T. Astrocytes Are More Resistant to Focal Cerebral Ischemia Than Neurons and Die by a Delayed Necrosis. Brain Pathol. 2009, 19, 630–641. [Google Scholar] [CrossRef]
- Marengo, B.; Monti, P.; Miele, M.; Menichini, P.; Ottaggio, L.; Foggetti, G.; Pulliero, A.; Izzotti, A.; Speciale, A.; Garbarino, O.; et al. Etoposide-Resistance in a Neuroblastoma Model Cell Line Is Associated with 13q14.3 Mono-Allelic Deletion and MiRNA-15a/16-1 down-Regulation. Sci. Rep. 2018, 8, 13762. [Google Scholar] [CrossRef]
- Pezzolo, A.; Parodi, F.; Corrias, M.V.; Cinti, R.; Gambini, C.; Pistoia, V. Tumor Origin of Endothelial Cells in Human Neuroblastoma. J. Clin. Oncol. 2007, 25, 376–383. [Google Scholar] [CrossRef]
- Karlsson, J.; Øra, I.; Pörn-Ares, I.; Påhlman, S. Arsenic Trioxide-Induced Death of Neuroblastoma Cells Involves Activation of Bax and Does Not Require P53. Clin. Cancer Res. 2004, 10, 3179–3188. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Marengo, B.; Zuccari, G.; Turrini, F.; Domenicotti, C. Dendrimer Nanodevices and Gallic Acid as Novel Strategies to Fight Chemoresistance in Neuroblastoma Cells. Nanomaterials 2020, 10, 1243. [Google Scholar] [CrossRef]
- Marini, C.; Cossu, V.; Kumar, M.; Milanese, M.; Cortese, K.; Bruno, S.; Bellese, G.; Carta, S.; Zerbo, R.A.; Torazza, C.; et al. The Role of Endoplasmic Reticulum in the Differential Endurance against Redox Stress in Cortical and Spinal Astrocytes from the Newborn SOD1G93A Mouse Model of Amyotrophic Lateral Sclerosis. Antioxidants 2021, 10, 1392. [Google Scholar] [CrossRef] [PubMed]
- Provenzano, F.; Nyberg, S.; Giunti, D.; Torazza, C.; Parodi, B.; Bonifacino, T.; Usai, C.; Kerlero de Rosbo, N.; Milanese, M.; Uccelli, A.; et al. Micro-RNAs Shuttled by Extracellular Vesicles Secreted from Mesenchymal Stem Cells Dampen Astrocyte Pathological Activation and Support Neuroprotection in In-Vitro Models of ALS. Cells 2022, 11, 3923. [Google Scholar] [CrossRef] [PubMed]
- Torazza, C.; Provenzano, F.; Gallia, E.; Cerminara, M.; Balbi, M.; Bonifacino, T.; Tessitore, S.; Ravera, S.; Usai, C.; Musante, I.; et al. Genetic Downregulation of the Metabotropic Glutamate Receptor Type 5 Dampens the Reactive and Neurotoxic Phenotype of Adult ALS Astrocytes. Cells 2023, 12, 1952. [Google Scholar] [CrossRef]
- Brofiga, M.; Losacco, S.; Poggio, F.; Zerbo, R.A.; Milanese, M.; Massobrio, P.; Burlando, B. Multiple Neuron Clusters on Micro-Electrode Arrays as an in Vitro Model of Brain Network. Sci. Rep. 2023, 13, 15604. [Google Scholar] [CrossRef]
- Bacchetti, F.; Schito, A.M.; Milanese, M.; Castellaro, S.; Alfei, S. Anti Gram-Positive Bacteria Activity of Synthetic Quaternary Ammonium Lipid and Its Precursor Phosphonium Salt. Int. J. Mol. Sci. 2024, 25, 2761. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure Time (h) | IC50 HTLA 230 (µM) | IC50 HTLA ER (µM) |
|---|---|---|
| 24 | 0.9257 ± 0.1637 | 1.2390 ± 0.0610 |
| 48 | 0.4623 ± 0.1031 | 1.3540 ± 0.0520 |
| 72 | 0.3892 ± 0.0836 | 1.1990 ± 0.0695 |
| Exposure Time (h) | Primary Spinal Cord Astrocytes (µM) | Primary Cortical Neurons (µM) |
|---|---|---|
| 24 | 7.044 ± 0.4760 | 1.5610 ± 0.2775 |
| 48 | 10.9700 ± 5.0935 | 2.2080 ± 0.3365 |
| 72 | 5.0950 ± 1.2240 | 0.7631 ± 0.1270 |
| IC50 (µM) | IC50 24 h (µM) | IC50 48 h (µM) | IC50 72 h (µM) | HC50 Experiment Time (µM) |
|---|---|---|---|---|
| RBCs * | N.A.Q. | N.A.Q. | N.A.Q. | 15.56 ± 12.13 |
| Spinal cord astrocytes | 7.044 ± 0.9805 | 10.9700 ± 5.0935 | 5.0950 ± 1.2240 | ---- |
| Cortical neurons | 1.5610 ± 0.2775 | 2.2080 ± 0.3365 | 0.7631 ± 0.1270 | ---- |
| HTLA 230 | 0.9257 ± 0.1637 | 0.4623 ± 0.1031 | 0.3892 ± 0.0836 | ---- |
| HTLA ER | 1.2390 ± 0.0610 | 1.3540 ± 0.0520 | 1.1990 ± 0.0695 | ---- |
| Cells | SI 24 h a | SI 48 h a | SI 72 h a |
|---|---|---|---|
| RBCs * | 16.81 | 33.66 | 39.98 |
| RBCs ** | 12.56 | 11.49 | 12.98 |
| Spinal cord astrocytes * | 7.61 | 23.73 | 13.09 |
| Spinal cord astrocytes ** | 5.69 | 8.10 | 4.25 |
| Cortical Neurons * | 1.69 | 4.78 | 1.96 |
| Cortical Neurons ** | 1.26 | 1.63 | 0.64 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfei, S.; Torazza, C.; Bacchetti, F.; Milanese, M.; Passalacqua, M.; Khaledizadeh, E.; Vernazza, S.; Domenicotti, C.; Marengo, B. TPP-Based Nanovesicles Kill MDR Neuroblastoma Cells and Induce Moderate ROS Increase, While Exerting Low Toxicity Towards Primary Cell Cultures: An In Vitro Study. Int. J. Mol. Sci. 2025, 26, 4991. https://doi.org/10.3390/ijms26114991
Alfei S, Torazza C, Bacchetti F, Milanese M, Passalacqua M, Khaledizadeh E, Vernazza S, Domenicotti C, Marengo B. TPP-Based Nanovesicles Kill MDR Neuroblastoma Cells and Induce Moderate ROS Increase, While Exerting Low Toxicity Towards Primary Cell Cultures: An In Vitro Study. International Journal of Molecular Sciences. 2025; 26(11):4991. https://doi.org/10.3390/ijms26114991
Chicago/Turabian StyleAlfei, Silvana, Carola Torazza, Francesca Bacchetti, Marco Milanese, Mario Passalacqua, Elaheh Khaledizadeh, Stefania Vernazza, Cinzia Domenicotti, and Barbara Marengo. 2025. "TPP-Based Nanovesicles Kill MDR Neuroblastoma Cells and Induce Moderate ROS Increase, While Exerting Low Toxicity Towards Primary Cell Cultures: An In Vitro Study" International Journal of Molecular Sciences 26, no. 11: 4991. https://doi.org/10.3390/ijms26114991
APA StyleAlfei, S., Torazza, C., Bacchetti, F., Milanese, M., Passalacqua, M., Khaledizadeh, E., Vernazza, S., Domenicotti, C., & Marengo, B. (2025). TPP-Based Nanovesicles Kill MDR Neuroblastoma Cells and Induce Moderate ROS Increase, While Exerting Low Toxicity Towards Primary Cell Cultures: An In Vitro Study. International Journal of Molecular Sciences, 26(11), 4991. https://doi.org/10.3390/ijms26114991