

Evaluating the Chemical Composition and Antitumor Activity of Origanum vulgare ssp. hirtum Essential Oil in a Preclinical Colon Cancer Model
 , , , , ,
, , , , ,  and
and
Abstract
1. Introduction
2. Results
2.1. Characterization of the Chemical Composition of Origanum vulgare ssp. hirtum Essential Oil
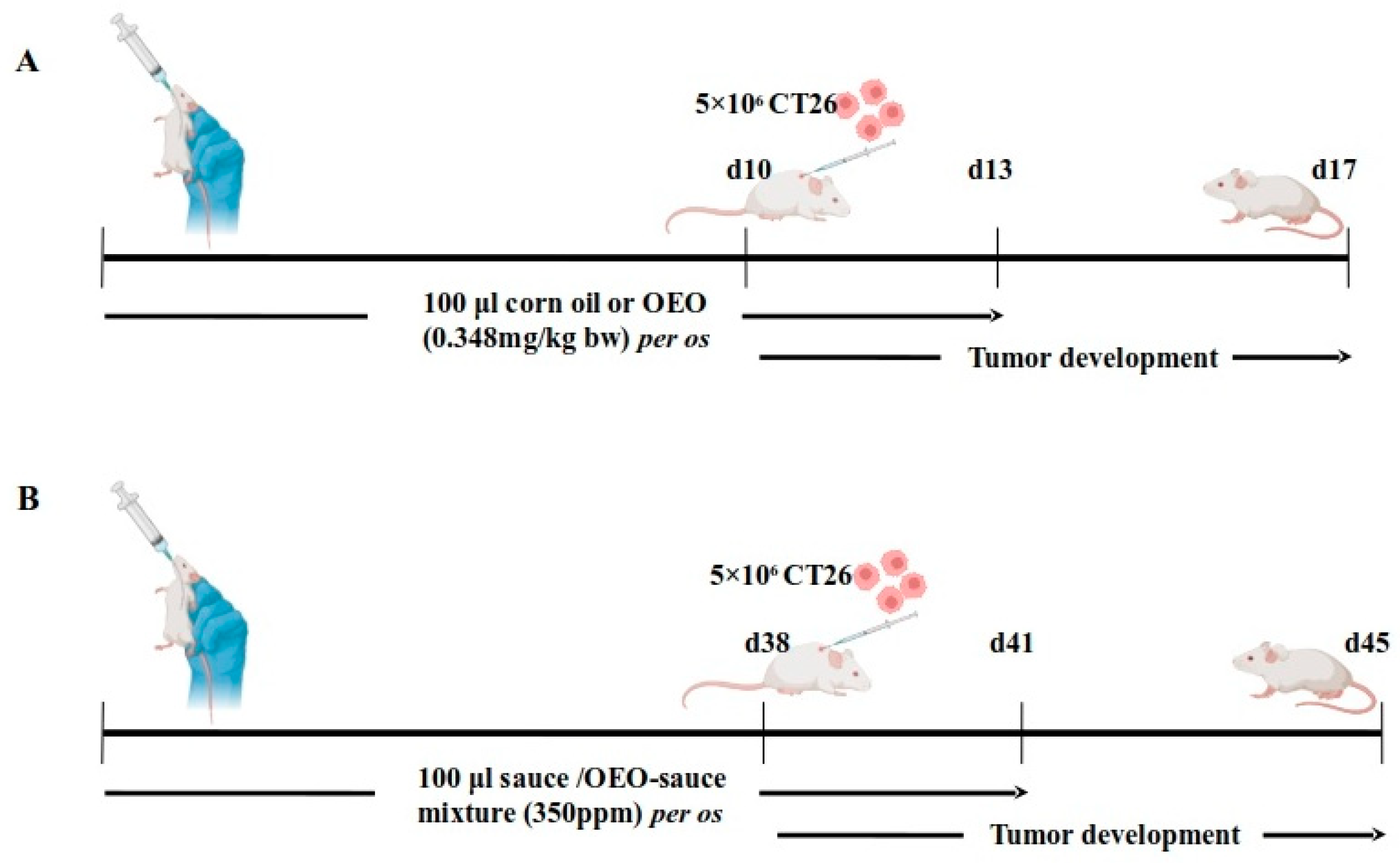
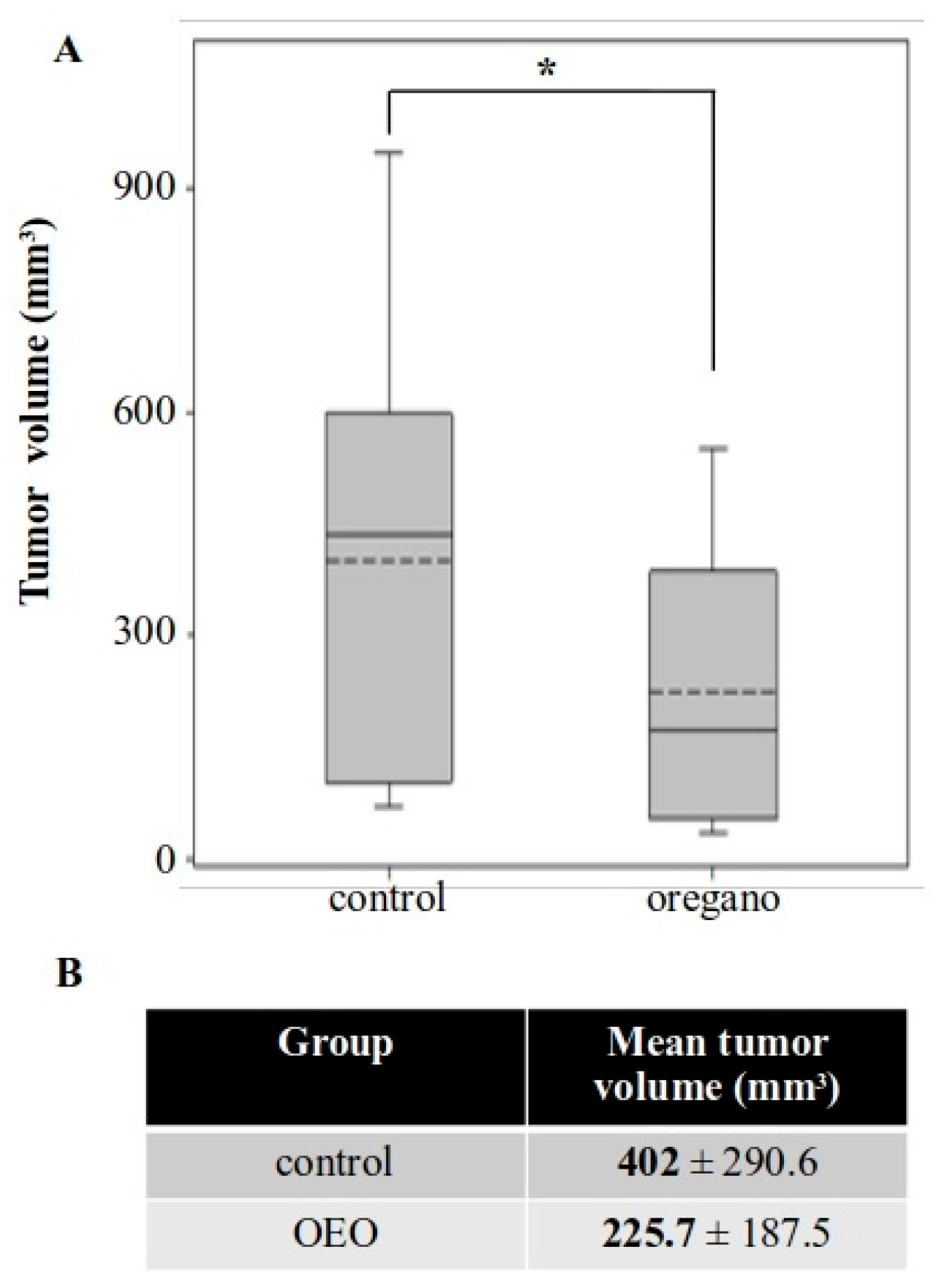
2.2. Tumor Growth Inhibition Following Consumption of Origanum vulgare ssp. hirtum Essential Oil
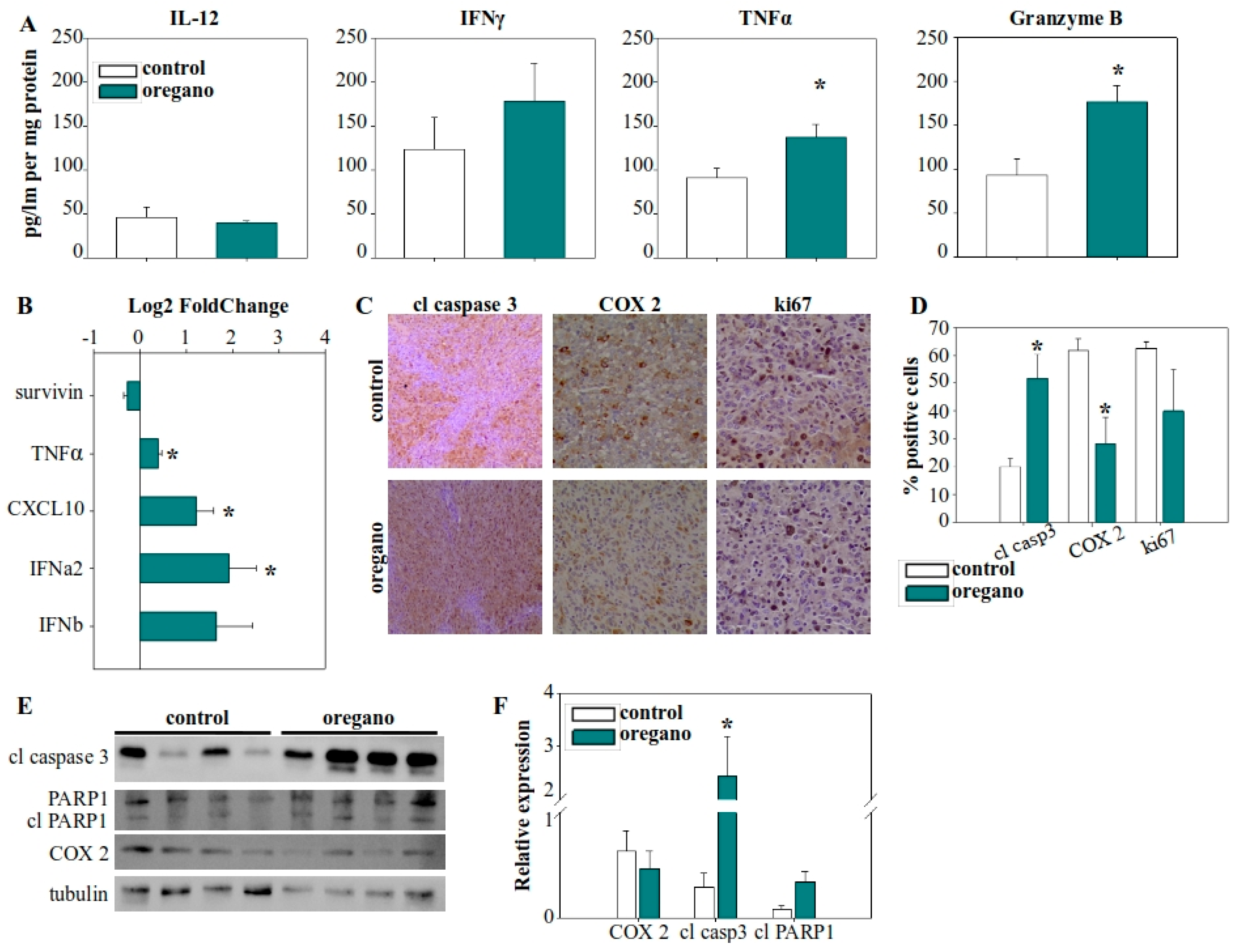
2.3. Expression Analysis of Cytokines and Tumor-Associated Molecules Following Administration of the Essential Oil
2.4. Sustained Growth Inhibitory Effect of Oregano Essential Oil Following Long-Term Prophylactic Administration of Low Essential Oil Concentration
2.5. Impact of Low-Dose Oregano Emulsion on the Expression of Immunomodulatory Molecules in the Tumor: Comparable to High Concentrations of Essential Oil
3. Discussion
4. Materials and Methods
4.1. Solvents
4.2. Extraction of Essential Oil and Preparation of Oregano Mixtures
4.3. Preparation of Standards and Samples
4.4. Liquid Chromatography Tandem Mass Spectrometry (LC-MS/MS) Conditions
4.5. Total Phenolic Content (TPC) and Total Flavonoid Content (TFC)
4.6. Total Condensed Tannins Content (TCTC)
4.7. Total Monoterpenoid Content (TMC)
4.8. Cell Lines and Culture
4.9. Animals and Tumor Models
4.10. Western Blot
4.11. Enzyme-Linked Immunosorbent Assay (ELISA)
4.12. Real-Time qPCR
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IFN | interferon |
| TNF | tumor necrosis factor |
| UPLC-MS/MS | ultra-performance liquid chromatography–mass spectrometry |
| ESI | electrospray ionization |
| LoD | limit of detection |
| LoQ | limit of quantification |
| TPC | total phenolic content |
| TFC | total flavonoid content |
| TCTC | total condensed tannins content |
| TMC | total monoterpenoid content |
| UPLC-ESI-QqQ | UPLC-electrospray ionization-triple quadrupole |
| IL | interleukin |
| NK | natural killer |
| CXCL | chemokine (C-X-C motif) ligand |
| Cox | cyclooxygenase |
| GC-MS | gas chromatography–mass spectrometry |
| ppm | parts per million |
| ppb | parts per billion |
| MCE | mixed cellulose esters |
| MRM | multiple reactions monitoring |
| PVDF | polyvinylidene fluoride |
| ELISA | enzyme-linked immunosorbent assay |
| qPCR | quantitative polymerase chain reaction |
References
- Aboukhalid, K.; Al Faiz, C.; Douaik, A.; Bakha, M.; Kursa, K.; Agacka-Mołdoch, M.; Machon, N.; Tomi, F.; Lamiri, A. Influence of Environmental Factors on Essential Oil Variability in Origanum compactum Benth. Growing Wild in Morocco. Chem. Biodivers. 2017, 14, e1700158. [Google Scholar] [CrossRef] [PubMed]
- Soltani, S.; Shakeri, A.; Iranshahi, M.; Boozari, M. A Review of the Phytochemistry and Antimicrobial Properties of Origanum vulgare L. and Subspecies. Iran. J. Pharm. Res. IJPR 2021, 20, 268–285. [Google Scholar] [CrossRef] [PubMed]
- Ličina, B.Z.; Stefanović, O.D.; Vasić, S.M.; Radojević, I.D.; Dekić, M.S.; Čomić, L.R. Biological activities of the extracts from wild growing Origanum vulgare L. Food Control 2013, 33, 498–504. [Google Scholar] [CrossRef]
- Mitropoulou, G.; Oreopoulou, A.; Papavassilopoulou, E.; Vamvakias, M.; Panas, P.; Fragias, S.; Kourkoutas, Y. Origanum vulgare ssp. hirtum Essential Oil as a Natural Intrinsic Hurdle against Common Spoilage and Pathogenic Microbes of Concern in Tomato Juice. Appl. Microbiol. 2021, 1, 1–10. [Google Scholar] [CrossRef]
- Kolypetri, S.; Kostoglou, D.; Nikolaou, A.; Kourkoutas, Y.; Giaouris, E. Chemical Composition, Antibacterial and Antibiofilm Actions of Oregano (Origanum vulgare subsp. hirtum) Essential Oil against Salmonella typhimurium and Listeria monocytogenes. Foods 2023, 12, 2893. [Google Scholar] [CrossRef]
- Spyridopoulou, K.; Fitsiou, E.; Bouloukosta, E.; Tiptiri-Kourpeti, A.; Vamvakias, M.; Oreopoulou, A.; Papavassilopoulou, E.; Pappa, A.; Chlichlia, K. Extraction, Chemical Composition, and Anticancer Potential of Origanum onites L. Essential Oil. Molecules 2019, 24, 2612. [Google Scholar] [CrossRef]
- Di Liberto, D.; Iacuzzi, N.; Pratelli, G.; Porrello, A.; Maggio, A.; La Bella, S.; De Blasio, A.; Notaro, A.; D’Anneo, A.; Emanuele, S.; et al. Cytotoxic Effect Induced by Sicilian Oregano Essential Oil in Human Breast Cancer Cells. Cells 2023, 12, 2733. [Google Scholar] [CrossRef]
- Arafat, K.; Al-Azawi, A.M.; Sulaiman, S.; Attoub, S. Exploring the Anticancer Potential of Origanum majorana Essential Oil Monoterpenes Alone and in Combination against Non-Small Cell Lung Cancer. Nutrients 2023, 15, 5010. [Google Scholar] [CrossRef]
- Tomsuk, Ö.; Kuete, V.; Sivas, H.; Kürkçüoğlu, M. Effects of essential oil of Origanum onites and its major component carvacrol on the expression of toxicity pathway genes in HepG2 cells. BMC Complement. Med. Ther. 2024, 24, 265. [Google Scholar] [CrossRef]
- Kamenova, K.; Iliev, I.; Prancheva, A.; Tuleshkov, P.; Rusanov, K.; Atanassov, I.; Petrov, P.D. Hydroxypropyl Cellulose Hydrogel Containing Origanum vulgare ssp. hirtum Essential-Oil-Loaded Polymeric Micelles for Enhanced Treatment of Melanoma. Gels 2024, 10, 627. [Google Scholar] [CrossRef]
- Arafat, K.; Sulaiman, S.; Al-Azawi, A.M.; Yasin, J.; Sugathan, S.; Nemmar, A.; Karam, S.; Attoub, S. Origanum majorana essential oil decreases lung tumor growth and metastasis in vitro and in vivo. Biomed. Pharmacother. 2022, 155, 113762. [Google Scholar] [CrossRef] [PubMed]
- Dias, D.A.; Urban, S.; Roessner, U. A historical overview of natural products in drug discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef] [PubMed]
- Catalano, A.; Iacopetta, D.; Ceramella, J.; Scumaci, D.; Giuzio, F.; Saturnino, C.; Aquaro, S.; Rosano, C.; Sinicropi, M.S. Multidrug Resistance (MDR): A Widespread Phenomenon in Pharmacological Therapies. Molecules 2022, 27, 616. [Google Scholar] [CrossRef]
- Cragg, G.M. Paclitaxel (Taxol): A success story with valuable lessons for natural product drug discovery and development. Med. Res. Rev. 1998, 18, 315–331. [Google Scholar] [CrossRef]
- Morgan, R.D.; McNeish, I.A.; Cook, A.D.; James, E.C.; Lord, R.; Dark, G.; Glasspool, R.M.; Krell, J.; Parkinson, C.; Poole, C.J.; et al. Objective responses to first-line neoadjuvant carboplatin–paclitaxel regimens for ovarian, fallopian tube, or primary peritoneal carcinoma (ICON8): Post-hoc exploratory analysis of a randomised, phase 3 trial. Lancet Oncol. 2021, 22, 277–288. [Google Scholar] [CrossRef]
- Xin, M.; Wang, Y.; Ren, Q.; Guo, Y. Formononetin and metformin act synergistically to inhibit growth of MCF-7 breast cancer cells in vitro. Biomed. Pharmacother. 2019, 109, 2084–2089. [Google Scholar] [CrossRef]
- Wei, T.; Xiaojun, X.; Peilong, C. Magnoflorine improves sensitivity to doxorubicin (DOX) of breast cancer cells via inducing apoptosis and autophagy through AKT/mTOR and p38 signaling pathways. Biomed. Pharmacother. 2020, 121, 109139. [Google Scholar] [CrossRef]
- Yang, M.-D.; Sun, Y.; Zhou, W.-J.; Xie, X.-Z.; Zhou, Q.-M.; Lu, Y.-Y.; Su, S.-B. Resveratrol Enhances Inhibition Effects of Cisplatin on Cell Migration and Invasion and Tumor Growth in Breast Cancer MDA-MB-231 Cell Models In Vivo and In Vitro. Molecules 2021, 26, 2204. [Google Scholar] [CrossRef]
- Sampaio, L.A.; Pina, L.T.S.; Serafini, M.R.; Tavares, D.D.S.; Guimarães, A.G. Antitumor Effects of Carvacrol and Thymol: A Systematic Review. Front. Pharmacol. 2021, 12, 702487. [Google Scholar] [CrossRef]
- Jin, H.; Leng, Q.; Zhang, C.; Zhu, Y.; Wang, J. P-cymene prevent high-fat diet-associated colorectal cancer by improving the structure of intestinal flora. J. Cancer 2021, 12, 4355–4361. [Google Scholar] [CrossRef]
- Acikgul, F.C.; Duran, N.; Kutlu, T.; Ay, E.; Tek, E.; Bayraktar, S. The therapeutic potential and molecular mechanism of Alpha-pinene, Gamma-terpinene, and P-cymene against melanoma cells. Heliyon 2024, 10, e36223. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.I.M.; Nascimento, E.A.; Silva, C.J.; Silva, T.G.; Aguiar, J.S. Anticancer activity of monoterpenes: A systematic review. Mol. Biol. Rep. 2021, 48, 5775–5785. [Google Scholar] [CrossRef] [PubMed]
- Machado, T.Q.; da Fonseca, A.C.C.; Duarte, A.B.S.; Robbs, B.K.; de Sousa, D.P. A Narrative Review of the Antitumor Activity of Monoterpenes from Essential Oils: An Update. Biomed Res. Int. 2022, 2022, 6317201. [Google Scholar] [CrossRef] [PubMed]
- Jafari Khorsand, G.; Morshedloo, M.R.; Mumivand, H.; Emami Bistgani, Z.; Maggi, F.; Khademi, A. Natural diversity in phenolic components and antioxidant properties of oregano (Origanum vulgare L.) accessions, grown under the same conditions. Sci. Rep. 2022, 12, 5813. [Google Scholar] [CrossRef]
- Betlej, I.; Żurek, N.; Cebulak, T.; Kapusta, I.; Balawejder, M.; Kiełtyka-Dadasiewicz, A.; Jaworski, S.; Lange, A.; Kutwin, M.; Krochmal-Marczak, B.; et al. Evaluation of Chemical Profile and Biological Properties of Extracts of Different Origanum vulgare Cultivars Growing in Poland. Int. J. Mol. Sci. 2024, 25, 9417. [Google Scholar] [CrossRef]
- Wahle, K.W.J.; Brown, I.; Rotondo, D.; Heys, S.D. Plant phenolics in the prevention and treatment of cancer. Adv. Exp. Med. Biol. 2010, 698, 36–51. [Google Scholar] [CrossRef]
- Abotaleb, M.; Liskova, A.; Kubatka, P.; Büsselberg, D. Therapeutic Potential of Plant Phenolic Acids in the Treatment of Cancer. Biomolecules 2020, 10, 221. [Google Scholar] [CrossRef]
- Ververis, A.; Ioannou, K.; Kyriakou, S.; Violaki, N.; Panayiotidis, M.I.; Plioukas, M.; Christodoulou, K. Sideritis scardica Extracts Demonstrate Neuroprotective Activity against Aβ(25-35) Toxicity. Plants 2023, 12, 1716. [Google Scholar] [CrossRef]
- Kyriakou, S.; Michailidou, K.; Amery, T.; Stewart, K.; Winyard, P.G.; Trafalis, D.T.; Franco, R.; Pappa, A.; Panayiotidis, M.I. Polyphenolics, glucosinolates and isothiocyanates profiling of aerial parts of Nasturtium officinale (Watercress). Front. Plant Sci. 2022, 13, 998755. [Google Scholar] [CrossRef]
- von Schomberg, R. A Vision of Responsible Research and Innovation. In Responsible Innovation; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 51–74. ISBN 9781118551424. [Google Scholar]
- Nisar, M.F.; Khadim, M.; Rafiq, M.; Chen, J.; Yang, Y.; Wan, C.C. Pharmacological Properties and Health Benefits of Eugenol: A Comprehensive Review. Oxid. Med. Cell. Longev. 2021, 2021, 2497354. [Google Scholar] [CrossRef]
- Issa, H.; Loubaki, L.; Al Amri, A.; Zibara, K.; Almutairi, M.H.; Rouabhia, M.; Semlali, A. Eugenol as a potential adjuvant therapy for gingival squamous cell carcinoma. Sci. Rep. 2024, 14, 10958. [Google Scholar] [CrossRef] [PubMed]
- Zari, A.T.; Zari, T.A.; Hakeem, K.R. Anticancer Properties of Eugenol: A Review. Molecules 2021, 26, 7407. [Google Scholar] [CrossRef] [PubMed]
- Padhy, I.; Paul, P.; Sharma, T.; Banerjee, S.; Mondal, A. Molecular Mechanisms of Action of Eugenol in Cancer: Recent Trends and Advancement. Life 2022, 12, 1795. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.; Taine, E.G.; Meng, D.; Cui, T.; Tan, W. Chlorogenic Acid: A Systematic Review on the Biological Functions, Mechanistic Actions, and Therapeutic Potentials. Nutrients 2024, 16, 924. [Google Scholar] [CrossRef]
- Gupta, A.; Atanasov, A.G.; Li, Y.; Kumar, N.; Bishayee, A. Chlorogenic acid for cancer prevention and therapy: Current status on efficacy and mechanisms of action. Pharmacol. Res. 2022, 186, 106505. [Google Scholar] [CrossRef]
- Murai, T.; Matsuda, S. The Chemopreventive Effects of Chlorogenic Acids, Phenolic Compounds in Coffee, against Inflammation, Cancer, and Neurological Diseases. Molecules 2023, 28, 2381. [Google Scholar] [CrossRef]
- Huang, W.; Lin, W.; Chen, B.; Zhang, J.; Gao, P.; Fan, Y.; Lin, Y.; Wei, P. NFAT and NF-κB dynamically co-regulate TCR and CAR signaling responses in human T cells. Cell Rep. 2023, 42, 112663. [Google Scholar] [CrossRef]
- Xue, N.; Zhou, Q.; Ji, M.; Jin, J.; Lai, F.; Chen, J.; Zhang, M.; Jia, J.; Yang, H.; Zhang, J.; et al. Chlorogenic acid inhibits glioblastoma growth through repolarizating macrophage from M2 to M1 phenotype. Sci. Rep. 2017, 7, 39011. [Google Scholar] [CrossRef]
- Bampidis, V.; Azimonti, G.; Bastos, M.d.L.; Christensen, H.; Kouba, M.; Kos Durjava, M.; López-Alonso, M.; López Puente, S.; Marcon, F.; Mayo, B.; et al. Safety and efficacy of an essential oil of Origanum vulgare ssp. hirtum (Link) leetsw. for all poultry species. EFSA J. Eur. Food Saf. Auth. 2019, 17, e05653. [Google Scholar] [CrossRef]
- Bampidis, V.; Azimonti, G.; Bastos, M.d.L.; Christensen, H.; Kouba, M.; Kos Durjava, M.; López-Alonso, M.; López Puente, S.; Marcon, F.; Mayo, B.; et al. Safety and efficacy of an essential oil from Origanum vulgare ssp. hirtum (Link) Ietsw. for all animal species. EFSA J. Eur. Food Saf. Auth. 2019, 17, e05909. [Google Scholar] [CrossRef]
- Simos, Y.V.; Zerikiotis, S.; Lekkas, P.; Athinodorou, A.-M.; Zachariou, C.; Tzima, C.; Assariotakis, A.; Peschos, D.; Tsamis, K.; Halabalaki, M.; et al. Oral Supplementation with Hydroxytyrosol Synthesized Using Genetically Modified Escherichia coli Strains and Essential Oils Mixture: A Pilot Study on the Safety and Biological Activity. Microorganisms 2023, 11, 770. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef] [PubMed]
- Mashimo, M.; Onishi, M.; Uno, A.; Tanimichi, A.; Nobeyama, A.; Mori, M.; Yamada, S.; Negi, S.; Bu, X.; Kato, J.; et al. The 89-kDa PARP1 cleavage fragment serves as a cytoplasmic PAR carrier to induce AIF-mediated apoptosis. J. Biol. Chem. 2021, 296, 100046. [Google Scholar] [CrossRef]
- Raskov, H.; Orhan, A.; Christensen, J.P.; Gögenur, I. Cytotoxic CD8(+) T cells in cancer and cancer immunotherapy. Br. J. Cancer 2021, 124, 359–367. [Google Scholar] [CrossRef]
- Rousalova, I.; Krepela, E. Granzyme B-induced apoptosis in cancer cells and its regulation (review). Int. J. Oncol. 2010, 37, 1361–1378. [Google Scholar] [CrossRef]
- Voskoboinik, I.; Whisstock, J.C.; Trapani, J.A. Perforin and granzymes: Function, dysfunction and human pathology. Nat. Rev. Immunol. 2015, 15, 388–400. [Google Scholar] [CrossRef]
- Zöphel, D.; Angenendt, A.; Kaschek, L.; Ravichandran, K.; Hof, C.; Janku, S.; Hoth, M.; Lis, A. Faster cytotoxicity with age: Increased perforin and granzyme levels in cytotoxic CD8(+) T cells boost cancer cell elimination. Aging Cell 2022, 21, e13668. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, J.; Bi, E.; Zhao, Y.; Qin, T.; Wang, Y.; Wang, A.; Gao, S.; Yi, Q.; Wang, S. TNF-α enhances Th9 cell differentiation and antitumor immunity via TNFR2-dependent pathways. J. Immunother. Cancer 2019, 7, 28. [Google Scholar] [CrossRef]
- Jorgovanovic, D.; Song, M.; Wang, L.; Zhang, Y. Roles of IFN-γ in tumor progression and regression: A review. Biomark. Res. 2020, 8, 49. [Google Scholar] [CrossRef]
- Tilsed, C.M.; Principe, N.; Kidman, J.; Chin, W.L.; Orozco Morales, M.L.; Zemek, R.M.; Chee, J.; Islam, R.; Fear, V.S.; Forbes, C.; et al. CD4(+) T cells drive an inflammatory, TNF-α/IFN-rich tumor microenvironment responsive to chemotherapy. Cell Rep. 2022, 41, 111874. [Google Scholar] [CrossRef]
- Castro, F.; Cardoso, A.P.; Gonçalves, R.M.; Serre, K.; Oliveira, M.J. Interferon-gamma at the crossroads of tumor immune surveillance or evasion. Front. Immunol. 2018, 9, 847. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Zhu, B.; Chen, D. Type I interferon-mediated tumor immunity and its role in immunotherapy. Cell. Mol. Life Sci. 2022, 79, 191. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Wang, W.; Xu, H.; Liang, Y.; Ding, J.; Lv, M.; Ren, B.; Peng, H.; Fu, Y.-X.; Zhu, M. Metabolic reprograming mediated by tumor cell-intrinsic type I IFN signaling is required for CD47-SIRPα blockade efficacy. Nat. Commun. 2024, 15, 5759. [Google Scholar] [CrossRef]
- Mödl, B.; Moritsch, S.; Zwolanek, D.; Eferl, R. Type I and II interferon signaling in colorectal cancer liver metastasis. Cytokine 2023, 161, 156075. [Google Scholar] [CrossRef]
- Wang, C.-L.; Ho, A.-S.; Chang, C.-C.; Sie, Z.-L.; Peng, C.-L.; Chang, J.; Cheng, C.-C. Radiotherapy enhances CXCR3(high)CD8(+) T cell activation through inducing IFNγ-mediated CXCL10 and ICAM-1 expression in lung cancer cells. Cancer Immunol. Immunother. 2023, 72, 1865–1880. [Google Scholar] [CrossRef]
- Majumder, S.; Bhattacharjee, S.; Paul Chowdhury, B.; Majumdar, S. CXCL10 is critical for the generation of protective CD8 T cell response induced by antigen pulsed CpG-ODN activated dendritic cells. PLoS ONE 2012, 7, e48727. [Google Scholar] [CrossRef]
- Cappuyns, S.; Philips, G.; Vandecaveye, V.; Boeckx, B.; Schepers, R.; Van Brussel, T.; Arijs, I.; Mechels, A.; Bassez, A.; Lodi, F.; et al. PD-1(-) CD45RA(+) effector-memory CD8 T cells and CXCL10(+) macrophages are associated with response to atezolizumab plus bevacizumab in advanced hepatocellular carcinoma. Nat. Commun. 2023, 14, 7825. [Google Scholar] [CrossRef]
- Lim, R.J.; Salehi-Rad, R.; Tran, L.M.; Oh, M.S.; Dumitras, C.; Crosson, W.P.; Li, R.; Patel, T.S.; Man, S.; Yean, C.E.; et al. CXCL9/10-engineered dendritic cells promote T cell activation and enhance immune checkpoint blockade for lung cancer. Cell reports. Med. 2024, 5, 101479. [Google Scholar] [CrossRef]
- Cheng, C.-C.; Chang, J.; Ho, A.-S.; Sie, Z.-L.; Peng, C.-L.; Wang, C.-L.; Dev, K.; Chang, C.-C. Tumor-intrinsic IFNα and CXCL10 are critical for immunotherapeutic efficacy by recruiting and activating T lymphocytes in tumor microenvironment. Cancer Immunol. Immunother. 2024, 73, 175. [Google Scholar] [CrossRef]
- Kyriakou, S.; Tragkola, V.; Paraskevaidis, I.; Plioukas, M.; Trafalis, D.T.; Franco, R.; Pappa, A.; Panayiotidis, M.I. Chemical Characterization and Biological Evaluation of Epilobium parviflorum Extracts in an In Vitro Model of Human Malignant Melanoma. Plants 2023, 12, 1590. [Google Scholar] [CrossRef]
- Spyridopoulou, K.; Kyriakou, S.; Nomikou, A.; Roupas, A.; Ermogenous, A.; Karamanoli, K.; Moyankova, D.; Djilianov, D.; Galanis, A.; Panayiotidis, M.I.; et al. Chemical Profiling, Antiproliferative and Antimigratory Capacity of Haberlea rhodopensis Extracts in an In Vitro Platform of Various Human Cancer Cell Lines. Antioxidants 2022, 11, 2305. [Google Scholar] [CrossRef] [PubMed]
- Ververis, A.; Kyriakou, S.; Paraskeva, H.; Panayiotidis, M.I.; Plioukas, M.; Christodoulou, K. Chemical Characterization and Assessment of the Neuroprotective Potential of Euphrasia officinalis. Int. J. Mol. Sci. 2024, 25, 12902. [Google Scholar] [CrossRef] [PubMed]
- Łukowski, A.; Jagiełło, R.; Robakowski, P.; Adamczyk, D.; Karolewski, P. Adaptation of a simple method to determine the total terpenoid content in needles of coniferous trees. Plant Sci. 2022, 314, 111090. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Phenolic Content (TPC) (μg of gallic acid eq/g of dry extract) | 25,600.65 ± 148.25 |
| Total Flavonoid Content (TFC) (μg of catechin eq/g of dry extract) | 17,899.24 ± 235.58 |
| Total Condensed Tannins Content (TCTC) (μg of catechin equivalents/g of dry extract) | 568.10 ± 15.88 |
| Total Monoterpenoid Content (TMC) (μg of linalool equivalents/g of dry extract) | 110.47 ± 9.43 |
| Origanum vulgare ssp. hirtum | |
|---|---|
| Compound | Quantity (ng/g of Dry Extract) |
| Benzoic acid derivatives | |
| m-hydroxy benzoic acid | 63.16 ± 2.47 |
| Protocatechuic acid | 22.04 ± 0.13 |
| Vanillin | 2.65 ± 0.16 |
| p-hydroxy benzaldehyde | 9.49 ± 0.77 |
| Gallic acid derivatives | |
| Gallic acid | 85.71 ± 2.32 |
| Ethyl gallate | 127.04 ± 6.14 |
| Cinnamic acid derivatives | |
| Ferulic acid | 38.98 ± 2.14 |
| Caffeic acid | 68.37 ± 4.96 |
| Dihydro caffeic acid | 123.47 ± 6.88 |
| Chlorogenic acid | 262.55 ± 10.55 |
| Coumarin derivatives | |
| Coumarin | 32.76 ± 2.86 |
| m-hydroxycoumarin | 87.94 ± 6.54 |
| Phenolic derivative | |
| Eugenol | 1060.20 ± 23.21 |
| Furanocoumarin derivative | |
| Xanthotoxol | 0.14 ± 0.01 |
| Flavanone derivatives | |
| 4′-methoxyflavanone | 49.79 ± 2.21 |
| Naringin | 19.39 ± 1.21 |
| Flavonol derivatives | |
| Isorhamnetin | 25.51 ± 1.36 |
| Quercetin-3-O-rhamnoside | 6.12 ± 0.08 |
| Myricetin-3-O-galactoside | 22.55 ± 1.36 |
| Myricetin-3-O-rhamnoside | 9.08 ± 0.42 |
| Kaempferol | 3429.59 ± 89.5 |
| Procyanidin | |
| Procyanidin-B2 | 11.33 ± 0.06 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aindelis, G.; Spyridopoulou, K.; Kyriakou, S.; Tiptiri-Kourpeti, A.; Panayiotidis, M.I.; Pappa, A.; Chlichlia, K. Evaluating the Chemical Composition and Antitumor Activity of Origanum vulgare ssp. hirtum Essential Oil in a Preclinical Colon Cancer Model. Int. J. Mol. Sci. 2025, 26, 4737. https://doi.org/10.3390/ijms26104737
Aindelis G, Spyridopoulou K, Kyriakou S, Tiptiri-Kourpeti A, Panayiotidis MI, Pappa A, Chlichlia K. Evaluating the Chemical Composition and Antitumor Activity of Origanum vulgare ssp. hirtum Essential Oil in a Preclinical Colon Cancer Model. International Journal of Molecular Sciences. 2025; 26(10):4737. https://doi.org/10.3390/ijms26104737
Chicago/Turabian StyleAindelis, Georgios, Katerina Spyridopoulou, Sotiris Kyriakou, Angeliki Tiptiri-Kourpeti, Mihalis I. Panayiotidis, Aglaia Pappa, and Katerina Chlichlia. 2025. "Evaluating the Chemical Composition and Antitumor Activity of Origanum vulgare ssp. hirtum Essential Oil in a Preclinical Colon Cancer Model" International Journal of Molecular Sciences 26, no. 10: 4737. https://doi.org/10.3390/ijms26104737
APA StyleAindelis, G., Spyridopoulou, K., Kyriakou, S., Tiptiri-Kourpeti, A., Panayiotidis, M. I., Pappa, A., & Chlichlia, K. (2025). Evaluating the Chemical Composition and Antitumor Activity of Origanum vulgare ssp. hirtum Essential Oil in a Preclinical Colon Cancer Model. International Journal of Molecular Sciences, 26(10), 4737. https://doi.org/10.3390/ijms26104737