
The Role of Intermittent Fasting in the Activation of Autophagy Processes in the Context of Cancer Diseases



Abstract

1. Introduction
2. Autophagy—Key Mechanism of Cellular Homeostasis
2.1. Types of Autophagy
- -
- Microautophagy involves the direct engulfment of small portions of the cytoplasm by lysosomes. This process facilitates nutrient recycling and supports cellular metabolic balance [12].
- -
- Chaperone-mediated autophagy (CMA) is a highly selective mechanism in which chaperone proteins (e.g., Hsp70) target specific substrates to lysosomes via the lysosomal membrane protein LAMP2A. This type of autophagy plays a crucial role in the removal of misfolded proteins and the maintenance of proteostatic homeostasis [13].
- -
- Macroautophagy is the most extensively studied and complex form of autophagy. It involves the encapsulation of damaged organelles and proteins within autophagosomal membranes, followed by their delivery to lysosomes for degradation. Macroautophagy is essential for cellular stress responses and intracellular resource recycling [14].
2.2. Autophagy in Non-Cancerous Disease Processes
2.3. Autophagy in Cancer Diseases
Autophagy as a Mechanism Supporting Cancer Progression
2.4. Pharmacological Modulation of Autophagy in Cancer Therapy
2.4.1. Autophagy Inhibition as a Therapeutic Strategy
2.4.2. The Induction of Autophagy as a Therapeutic Strategy
3. Intermittent Fasting
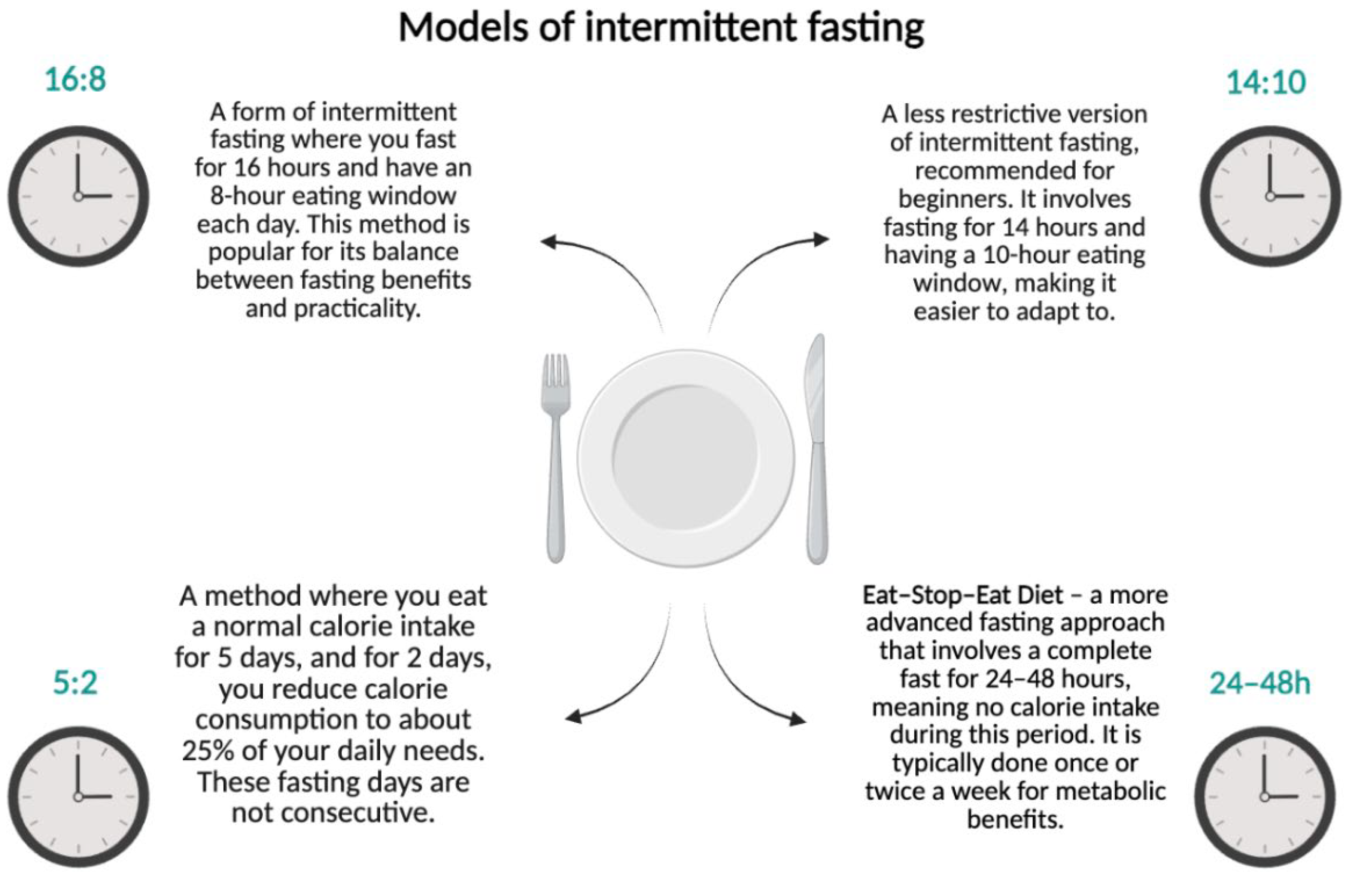
3.1. Types of Intermittent Fasting
- 16:8—16 h of fasting with an 8 h eating window;
- 14:10—a less restrictive form recommended for beginners;
- 5:2—consuming a standard number of calories for 5 days and reducing intake to 25% of daily requirements for 2 days;
- Eat–Stop–Eat—complete fasting for 24–48 h.
3.2. The Impact of Intermittent Fasting on the Body
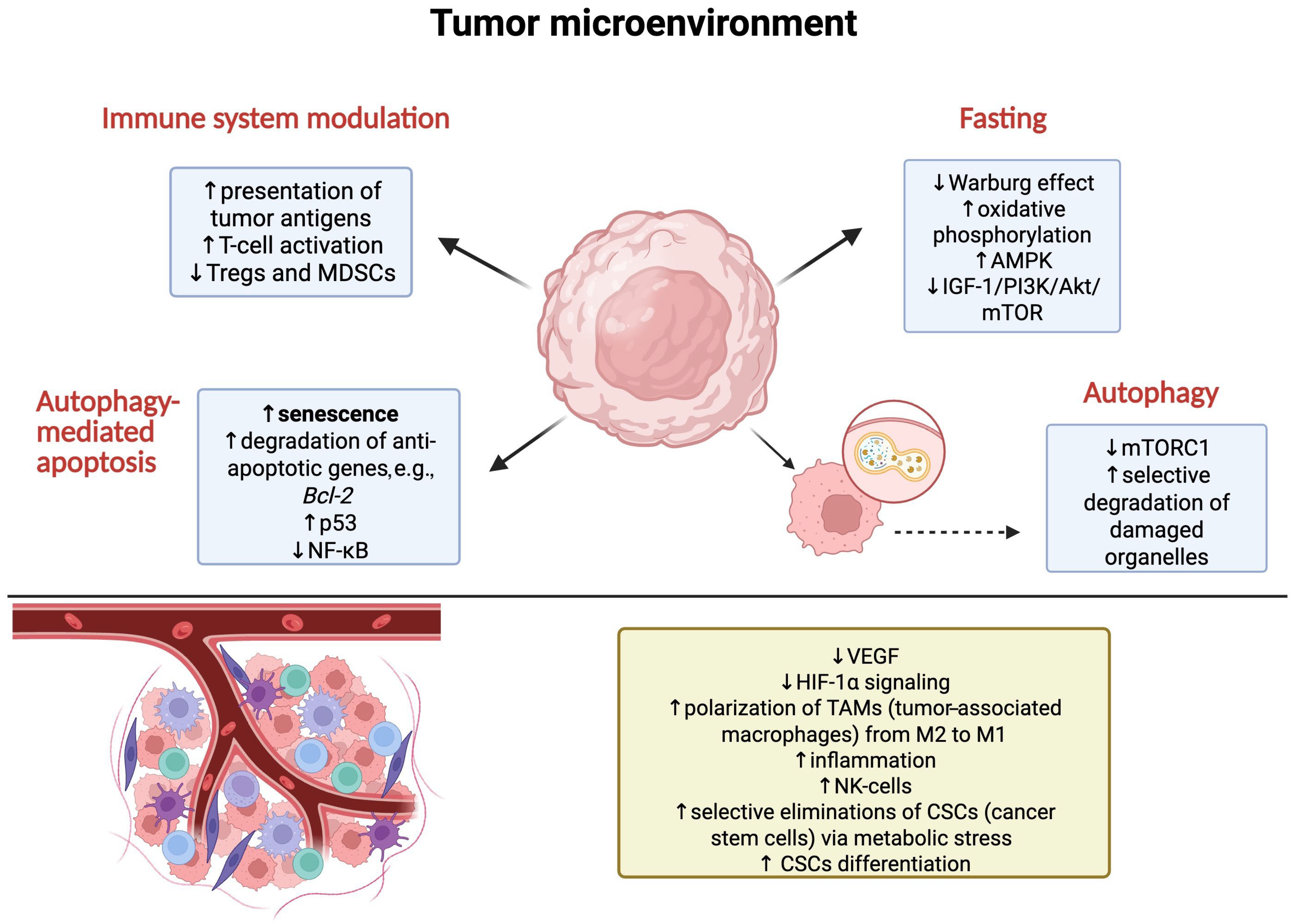
3.3. Impact of Autophagy Induced by Intermittent Fasting on Cancer
3.4. Potential Risks of Intermittent Fasting in Cancer Patients
Risk of Weight Loss and Cachexia
- Body weakness—loss of muscle mass impairs function and reduces the patient’s quality of life.
- Increased susceptibility to infections—malnutrition weakens the immune system, which increases the risk of complications.
- Decreased therapy effectiveness—cachectic patients have a poorer tolerance to chemotherapy and radiotherapy, which may lead to treatment limitations.Although IF can regulate cancer cell metabolism, its use in patients with advanced cancer remains controversial. The lack of conclusive clinical evidence suggesting benefits of this strategy in patients with cachexia means that further research is needed [7,126].
4. Conclusions
5. Materials and Methods
Literature Search Strategy
- (autophagy[Title/Abstract] OR “Autophagy”[MeSH Terms]) AND (intermittent fasting[Title/Abstract] OR “Intermittent Fasting”[MeSH Terms]) AND (cancer[Title/Abstract] OR tumor[Title/Abstract] OR neoplasm[Title/Abstract])
- (autophagy[Title/Abstract]) AND (fasting mimicking diet[Title/Abstract]) AND (cancer[Title/Abstract])
- (autophagy[Title/Abstract]) AND (caloric restriction[Title/Abstract]) AND (neoplasm[Title/Abstract])
- (intermittent fasting[Title/Abstract]) AND (tumor metabolism[Title/Abstract]) AND (autophagy[Title/Abstract])
- (fasting[Title/Abstract]) AND (chemosensitization[Title/Abstract]) AND (autophagy[Title/Abstract]) AND (cancer[Title/Abstract])
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 3-MA | 3-methyladenine |
| Akt | protein kinase B (PKB) |
| Ambra1 | autophagy and Beclin 1 regulator 1 |
| AMPK | 5′AMP-activated protein kinase |
| ATG5/7/12 | autophagy gene 5/7/12 |
| Bcl2 | B-cell lymphoma 2 |
| BECN2 | Beclin 2 |
| BECN1 | Beclin 1 |
| BPC11 | biphosphinic paladacycle complex 11 |
| BRAF | B-Raf proto-oncogene, serine/threonine kinase |
| cAMP | cyclic adenosine monophosphate |
| CMA | chaperone-mediated autophagy |
| CR | caloric restriction |
| CRP | C-reactive protein |
| CSCs | cancer stem cells |
| FMD | fasting-mimicking diet |
| GLUT1/2 | glucose transporter ½ |
| HDAC | histone deacetylase |
| HDL | high-density lipoprotein |
| HIF-1α | hypoxia-inducible factor 1-alpha |
| IF | intermittent fasting |
| IGF-1 | insulin-like growth factor 1 |
| JAK1/STAT | Janus kinase 1/signal transducer and activator of transcription |
| LAMP2A | lysosome-associated membrane protein 2A |
| LDL | low-density lipoprotein |
| MCL1 | myeloid cell leukemia 1 |
| MDM2 | mouse double minute 2 homolog |
| MDSC | myeloid-derived suppressor cells |
| MHC II | major histocompatibility complex class II |
| mTOR | mammalian target of rapamycin |
| mTORC1 | mammalian target of rapamycin complex 1 |
| MYC | MYC proto-oncogene, bHLH transcription factor |
| NAFLD | non-alcoholic fatty liver disease |
| NGFR | nerve growth factor receptor |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NK-cells | natural killer cells |
| PF | periodic fasting |
| p53 | tumor protein p53 |
| PI3K | phosphoinositide 3-kinase |
| PINK1 | PTEN-induced kinase 1 |
| PKA | protein kinase A |
| RND3 | Rho family GTPase 3 (RhoE) |
| ROS | reactive oxygen species |
| TAM | tumor-associated macrophage |
| TFEB | transcription factor EB |
| TNF-α | tumor necrosis factor alpha |
| TPT1 | tumor protein, translationally controlled 1 |
| Tregs | regulatory T cells |
| TRF | time-restricted feeding |
| ULK1 | Unc-51-like autophagy activating kinase 1 |
| UCP1 | uncoupling protein 1 |
| VEGF | vascular endothelial growth factor |
| WHO | World Health Organization |
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global Cancer Statistics 2022: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Bizuayehu, H.M.; Ahmed, K.Y.; Kibret, G.D.; Dadi, A.F.; Belachew, S.A.; Bagade, T.; Tegegne, T.K.; Venchiarutti, R.L.; Kibret, K.T.; Hailegebireal, A.H.; et al. Global Disparities of Cancer and Its Projected Burden in 2050. JAMA Netw. Open 2024, 7, e2443198. [Google Scholar] [CrossRef] [PubMed]
- Piña-Sánchez, P.; Chávez-González, A.; Ruiz-Tachiquín, M.; Vadillo, E.; Monroy-García, A.; Montesinos, J.J.; Grajales, R.; de la Barrera, M.G.; Mayani, H. Cancer Biology, Epidemiology, and Treatment in the 21st Century: Current Status and Future Challenges From a Biomedical Perspective. Cancer Control. 2021, 28, 10732748211038735. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Xia, Z.; Xia, W.; Jiang, P. Metabolic Reprogramming, Sensing, and Cancer Therapy. Cell Rep. 2024, 43, 115064. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef]
- Safdie, F.M.; Dorff, T.; Quinn, D.; Fontana, L.; Wei, M.; Lee, C.; Cohen, P.; Longo, V.D. Fasting and Cancer Treatment in Humans: A Case Series Report. Aging 2009, 1, 988–1007. [Google Scholar] [CrossRef]
- Fatima, G.; Mehdi, A.A.; Fedacko, J.; Hadi, N.; Magomedova, A.; Mehdi, A. Fasting as Cancer Treatment: Myth or Breakthrough in Oncology. Cureus 2025, 17, e81395. [Google Scholar] [CrossRef]
- Liu, S.; Yao, S.; Yang, H.; Liu, S.; Wang, Y. Autophagy: Regulator of Cell Death. Cell Death Dis. 2023, 14, 648. [Google Scholar] [CrossRef]
- Holczer, M.; Hajdú, B.; Lőrincz, T.; Szarka, A.; Bánhegyi, G.; Kapuy, O. Fine-Tuning of AMPK–ULK1–mTORC1 Regulatory Triangle Is Crucial for Autophagy Oscillation. Sci. Rep. 2020, 10, 17803. [Google Scholar] [CrossRef]
- Glick, D.; Barth, S.; Macleod, K.F. Autophagy: Cellular and Molecular Mechanisms. J. Pathol. 2010, 221, 3–12. [Google Scholar] [CrossRef]
- Intermittent Fasting: Cellular Cleansing to Improve Health? Available online: https://www.pasteur.fr/en/research-journal/news/intermittent-fasting-cellular-cleansing-improve-health (accessed on 12 May 2025).
- Kul, S.; Savaş, E.; Öztürk, Z.A.; Karadağ, G. Does Ramadan Fasting Alter Body Weight and Blood Lipids and Fasting Blood Glucose in a Healthy Population? A Meta-Analysis. J. Relig. Health 2014, 53, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Aksungar, F.B.; Topkaya, A.E.; Akyildiz, M. Interleukin-6, C-Reactive Protein and Biochemical Parameters during Prolonged Intermittent Fasting. Ann. Nutr. Metab. 2007, 51, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Sadeghirad, B.; Motaghipisheh, S.; Kolahdooz, F.; Zahedi, M.J.; Haghdoost, A.A. Islamic Fasting and Weight Loss: A Systematic Review and Meta-Analysis. Public Health Nutr. 2014, 17, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Virgilio, L.; Silva-Lucero, M.-C.; Flores-Morelos, D.-S.; Gallardo-Nieto, J.; Lopez-Toledo, G.; Abarca-Fernandez, A.-M.; Zacapala-Gómez, A.-E.; Luna-Muñoz, J.; Montiel-Sosa, F.; Soto-Rojas, L.O.; et al. Autophagy: A Key Regulator of Homeostasis and Disease: An Overview of Molecular Mechanisms and Modulators. Cells 2022, 11, 2262. [Google Scholar] [CrossRef]
- Yun, C.W.; Jeon, J.; Go, G.; Lee, J.H.; Lee, S.H. The Dual Role of Autophagy in Cancer Development and a Therapeutic Strategy for Cancer by Targeting Autophagy. Int. J. Mol. Sci. 2020, 22, 179. [Google Scholar] [CrossRef]
- Furmli, S.; Elmasry, R.; Ramos, M.; Fung, J. Therapeutic Use of Intermittent Fasting for People with Type 2 Diabetes as an Alternative to Insulin. BMJ Case Rep. 2018, 2018, bcr2017221854. [Google Scholar] [CrossRef]
- Hara, T.; Nakamura, K.; Matsui, M.; Yamamoto, A.; Nakahara, Y.; Suzuki-Migishima, R.; Yokoyama, M.; Mishima, K.; Saito, I.; Okano, H.; et al. Suppression of Basal Autophagy in Neural Cells Causes Neurodegenerative Disease in Mice. Nature 2006, 441, 885–889. [Google Scholar] [CrossRef]
- Li, X.; He, S.; Ma, B. Autophagy and Autophagy-Related Proteins in Cancer. Mol. Cancer 2020, 19, 12. [Google Scholar] [CrossRef]
- Lee, J.-H.; Yu, W.H.; Kumar, A.; Lee, S.; Mohan, P.S.; Peterhoff, C.M.; Wolfe, D.M.; Martinez-Vicente, M.; Massey, A.C.; Sovak, G.; et al. Lysosomal Proteolysis and Autophagy Require Presenilin 1 and Are Disrupted by Alzheimer-Related PS1 Mutations. Cell 2010, 141, 1146–1158. [Google Scholar] [CrossRef]
- Bai, B.; Wang, X.; Li, Y.; Chen, P.-C.; Yu, K.; Dey, K.K.; Yarbro, J.M.; Han, X.; Lutz, B.M.; Rao, S.; et al. Deep Multilayer Brain Proteomics Identifies Molecular Networks in Alzheimer’s Disease Progression. Neuron 2020, 106, 700. [Google Scholar] [CrossRef]
- Sciarretta, S.; Volpe, M.; Sadoshima, J. mTOR Signaling in Cardiac Physiology and Disease. Circ. Res. 2014, 114, 549–564. [Google Scholar] [CrossRef] [PubMed]
- Sciarretta, S.; Maejima, Y.; Zablocki, D.; Sadoshima, J. The Role of Autophagy in the Heart. Annu. Rev. Physiol. 2018, 80, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Ryter, S.W.; Koo, J.K.; Choi, A.M.K. Molecular Regulation of Autophagy and Its Implications for Metabolic Diseases. Curr. Opin. Clin. Nutr. Metab. Care 2014, 17, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.-M.; Lim, H.; Hur, K.Y.; Quan, W.; Lee, H.-Y.; Cheon, H.; Ryu, D.; Koo, S.-H.; Kim, H.L.; Kim, J.; et al. Systemic Autophagy Insufficiency Compromises Adaptation to Metabolic Stress and Facilitates Progression from Obesity to Diabetes. Nat. Commun. 2014, 5, 4934. [Google Scholar] [CrossRef] [PubMed]
- Cairó, M.; Villarroya, J. The Role of Autophagy in Brown and Beige Adipose Tissue Plasticity. J. Physiol. Biochem. 2020, 76, 213–226. [Google Scholar] [CrossRef]
- Masiero, E.; Agatea, L.; Mammucari, C.; Blaauw, B.; Loro, E.; Komatsu, M.; Metzger, D.; Reggiani, C.; Schiaffino, S.; Sandri, M. Autophagy Is Required to Maintain Muscle Mass. Cell Metab. 2009, 10, 507–515. [Google Scholar] [CrossRef]
- Shapiro, I.M.; Layfield, R.; Lotz, M.; Settembre, C.; Whitehouse, C. Boning up on Autophagy: The Role of Autophagy in Skeletal Biology. Autophagy 2014, 10, 7–19. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Rao, S.; Tortola, L.; Perlot, T.; Wirnsberger, G.; Novatchkova, M.; Nitsch, R.; Sykacek, P.; Frank, L.; Schramek, D.; Komnenovic, V.; et al. A Dual Role for Autophagy in a Murine Model of Lung Cancer. Nat. Commun. 2014, 5, 3056. [Google Scholar] [CrossRef]
- Faubert, B.; Solmonson, A.; DeBerardinis, R.J. Metabolic Reprogramming and Cancer Progression. Science 2020, 368, eaaw5473. [Google Scholar] [CrossRef]
- Di Leo, L.; Bodemeyer, V.; Bosisio, F.M.; Claps, G.; Carretta, M.; Rizza, S.; Faienza, F.; Frias, A.; Khan, S.; Bordi, M.; et al. Loss of Ambra1 Promotes Melanoma Growth and Invasion. Nat. Commun. 2021, 12, 2550. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, L.D.; Young, A.R.J.; Pérez-Mancera, P.A.; Nimmervoll, B.; Jaulim, A.; Chen, H.-C.; McIntyre, D.J.O.; Brais, R.; Ricketts, T.; Pacey, S.; et al. A Novel Atg5-shRNA Mouse Model Enables Temporal Control of Autophagy in Vivo. Autophagy 2018, 14, 1256–1266. [Google Scholar] [CrossRef]
- Takamura, A.; Komatsu, M.; Hara, T.; Sakamoto, A.; Kishi, C.; Waguri, S.; Eishi, Y.; Hino, O.; Tanaka, K.; Mizushima, N. Autophagy-Deficient Mice Develop Multiple Liver Tumors. Genes Dev. 2011, 25, 795–800. [Google Scholar] [CrossRef]
- Kim, S.; Domon-Dell, C.; Kang, J.; Chung, D.H.; Freund, J.-N.; Evers, B.M. Down-Regulation of the Tumor Suppressor PTEN by the Tumor Necrosis Factor-Alpha/Nuclear Factor-kappaB (NF-kappaB)-Inducing Kinase/NF-kappaB Pathway Is Linked to a Default IkappaB-Alpha Autoregulatory Loop. J. Biol. Chem. 2004, 279, 4285–4291. [Google Scholar] [CrossRef]
- Rosenfeldt, M.T.; O’Prey, J.; Lindsay, C.R.; Nixon, C.; Roth, S.; Sansom, O.J.; Ryan, K.M. Loss of Autophagy Affects Melanoma Development in a Manner Dependent on PTEN Status. Cell Death Differ. 2021, 28, 1437–1439. [Google Scholar] [CrossRef]
- Miller, D.R.; Thorburn, A. Autophagy and Organelle Homeostasis in Cancer. Dev. Cell 2021, 56, 906–918. [Google Scholar] [CrossRef]
- Gomes, L.R.; Menck, C.F.M.; Cuervo, A.M. Chaperone-Mediated Autophagy Prevents Cellular Transformation by Regulating MYC Proteasomal Degradation. Autophagy 2017, 13, 928–940. [Google Scholar] [CrossRef]
- Bonhoure, A.; Vallentin, A.; Martin, M.; Senff-Ribeiro, A.; Amson, R.; Telerman, A.; Vidal, M. Acetylation of Translationally Controlled Tumor Protein Promotes Its Degradation through Chaperone-Mediated Autophagy. Eur. J. Cell Biol. 2017, 96, 83–98. [Google Scholar] [CrossRef]
- Lu, T.-L.; Huang, G.-J.; Wang, H.-J.; Chen, J.-L.; Hsu, H.-P.; Lu, T.-J. Hispolon Promotes MDM2 Downregulation through Chaperone-Mediated Autophagy. Biochem. Biophys. Res. Commun. 2010, 398, 26–31. [Google Scholar] [CrossRef]
- Schneider, J.L.; Villarroya, J.; Diaz-Carretero, A.; Patel, B.; Urbanska, A.M.; Thi, M.M.; Villarroya, F.; Santambrogio, L.; Cuervo, A.M. Loss of Hepatic Chaperone-Mediated Autophagy Accelerates Proteostasis Failure in Aging. Aging Cell 2015, 14, 249–264. [Google Scholar] [CrossRef]
- Palikaras, K.; Lionaki, E.; Tavernarakis, N. Mechanisms of Mitophagy in Cellular Homeostasis, Physiology and Pathology. Nat. Cell Biol. 2018, 20, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa-Ishimoto, Y.; Hwang, S.; Cadwell, K. Autophagy and Inflammation. Annu. Rev. Immunol. 2018, 36, 73–101. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Pietrocola, F.; Bravo-San Pedro, J.M.; Amaravadi, R.K.; Baehrecke, E.H.; Cecconi, F.; Codogno, P.; Debnath, J.; Gewirtz, D.A.; Karantza, V.; et al. Autophagy in Malignant Transformation and Cancer Progression. EMBO J. 2015, 34, 856–880. [Google Scholar] [CrossRef] [PubMed]
- Kon, M.; Kiffin, R.; Koga, H.; Chapochnick, J.; Macian, F.; Varticovski, L.; Cuervo, A.M. Chaperone-Mediated Autophagy Is Required for Tumor Growth. Sci. Transl. Med. 2011, 3, 109ra117. [Google Scholar] [CrossRef]
- Zhou, J.; Yang, J.; Fan, X.; Hu, S.; Zhou, F.; Dong, J.; Zhang, S.; Shang, Y.; Jiang, X.; Guo, H.; et al. Chaperone-Mediated Autophagy Regulates Proliferation by Targeting RND3 in Gastric Cancer. Autophagy 2016, 12, 515–528. [Google Scholar] [CrossRef]
- Suzuki, J.; Nakajima, W.; Suzuki, H.; Asano, Y.; Tanaka, N. Chaperone-Mediated Autophagy Promotes Lung Cancer Cell Survival through Selective Stabilization of the pro-Survival Protein, MCL1. Biochem. Biophys. Res. Commun. 2017, 482, 1334–1340. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Petroni, G.; Amaravadi, R.K.; Baehrecke, E.H.; Ballabio, A.; Boya, P.; Bravo-San Pedro, J.M.; Cadwell, K.; Cecconi, F.; Choi, A.M.K.; et al. Autophagy in Major Human Diseases. EMBO J. 2021, 40, e108863. [Google Scholar] [CrossRef]
- Gundamaraju, R.; Lu, W.; Paul, M.K.; Jha, N.K.; Gupta, P.K.; Ojha, S.; Chattopadhyay, I.; Rao, P.V.; Ghavami, S. Autophagy and EMT in Cancer and Metastasis: Who Controls Whom? Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166431. [Google Scholar] [CrossRef]
- Sharifi, M.N.; Mowers, E.E.; Drake, L.E.; Collier, C.; Chen, H.; Zamora, M.; Mui, S.; Macleod, K.F. Autophagy Promotes Focal Adhesion Disassembly and Cell Motility of Metastatic Tumor Cells through the Direct Interaction of Paxillin with LC3. Cell Rep. 2016, 15, 1660–1672. [Google Scholar] [CrossRef]
- Liang, X.H.; Jackson, S.; Seaman, M.; Brown, K.; Kempkes, B.; Hibshoosh, H.; Levine, B. Induction of Autophagy and Inhibition of Tumorigenesis by Beclin 1. Nature 1999, 402, 672–676. [Google Scholar] [CrossRef]
- Yue, Z.; Jin, S.; Yang, C.; Levine, A.J.; Heintz, N. Beclin 1, an Autophagy Gene Essential for Early Embryonic Development, Is a Haploinsufficient Tumor Suppressor. Proc. Natl. Acad. Sci. USA 2003, 100, 15077–15082. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, T.; Knauer, H.; Schauer, A.; Büttner, S.; Ruckenstuhl, C.; Carmona-Gutierrez, D.; Ring, J.; Schroeder, S.; Magnes, C.; Antonacci, L.; et al. Induction of Autophagy by Spermidine Promotes Longevity. Nat. Cell Biol. 2009, 11, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- White, E. Deconvoluting the Context-Dependent Role for Autophagy in Cancer. Nat. Rev. Cancer 2012, 12, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Mathew, R.; White, E. Autophagy, Stress, and Cancer Metabolism: What Doesn’t Kill You Makes You Stronger. Cold Spring Harb. Symp. Quant. Biol. 2011, 76, 389–396. [Google Scholar] [CrossRef]
- Amaravadi, R.K.; Kimmelman, A.C.; Debnath, J. Targeting Autophagy in Cancer: Recent Advances and Future Directions. Cancer Discov. 2019, 9, 1167–1181. [Google Scholar] [CrossRef]
- Bishop, E.; Bradshaw, T.D. Autophagy Modulation: A Prudent Approach in Cancer Treatment? Cancer Chemother. Pharmacol. 2018, 82, 913–922. [Google Scholar] [CrossRef]
- Chen, J.-L.; Wu, X.; Yin, D.; Jia, X.-H.; Chen, X.; Gu, Z.-Y.; Zhu, X.-M. Autophagy Inhibitors for Cancer Therapy: Small Molecules and Nanomedicines. Pharmacol. Ther. 2023, 249, 108485. [Google Scholar] [CrossRef]
- Grasso, S.; Pereira, G.J.S.; Palmeira-Dos-Santos, C.; Calgarotto, A.K.; Martínez-Lacaci, I.; Ferragut, J.A.; Smaili, S.S.; Bincoletto, C. Autophagy Regulates Selumetinib (AZD6244) Induced-Apoptosis in Colorectal Cancer Cells. Eur. J. Med. Chem. 2016, 122, 611–618. [Google Scholar] [CrossRef]
- Mauthe, M.; Orhon, I.; Rocchi, C.; Zhou, X.; Luhr, M.; Hijlkema, K.-J.; Coppes, R.P.; Engedal, N.; Mari, M.; Reggiori, F. Chloroquine Inhibits Autophagic Flux by Decreasing Autophagosome-Lysosome Fusion. Autophagy 2018, 14, 1435–1455. [Google Scholar] [CrossRef]
- Mauvezin, C.; Neufeld, T.P. Bafilomycin A1 Disrupts Autophagic Flux by Inhibiting Both V-ATPase-Dependent Acidification and Ca-P60A/SERCA-Dependent Autophagosome-Lysosome Fusion. Autophagy 2015, 11, 1437–1438. [Google Scholar] [CrossRef]
- Wu, Y.-T.; Tan, H.-L.; Shui, G.; Bauvy, C.; Huang, Q.; Wenk, M.R.; Ong, C.-N.; Codogno, P.; Shen, H.-M. Dual Role of 3-Methyladenine in Modulation of Autophagy via Different Temporal Patterns of Inhibition on Class I and III Phosphoinositide 3-Kinase. J. Biol. Chem. 2010, 285, 10850–10861. [Google Scholar] [CrossRef] [PubMed]
- Gigli, R.; Pereira, G.J.S.; Antunes, F.; Bechara, A.; Garcia, D.M.; Spindola, D.G.; Jasiulionis, M.G.; Caires, A.C.F.; Smaili, S.S.; Bincoletto, C. The Biphosphinic Paladacycle Complex Induces Melanoma Cell Death through Lysosomal–Mitochondrial Axis Modulation and Impaired Autophagy. Eur. J. Med. Chem. 2016, 107, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, H.; Liu, C.; Yuan, X. Role of ROS-mediated Autophagy in Melanoma (Review). Mol. Med. Rep. 2022, 26, 303. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.-Z.; Sun, J.-F.; Chen, H. The Role of Endoplasmic Reticulum Stress in Melanoma. Int. J. Dermatol. Venereol. 2023, 6, 150–156. [Google Scholar] [CrossRef]
- Tao, H.; Qian, P.; Lu, J.; Guo, Y.; Zhu, H.; Wang, F. Autophagy Inhibition Enhances Radiosensitivity of Eca-109 Cells via the Mitochondrial Apoptosis Pathway. Int. J. Oncol. 2018, 52, 1853–1862. [Google Scholar] [CrossRef]
- Liang, B.; Kong, D.; Liu, Y.; Liang, N.; He, M.; Ma, S.; Liu, X. Autophagy Inhibition Plays the Synergetic Killing Roles with Radiation in the Multi-Drug Resistant SKVCR Ovarian Cancer Cells. Radiat. Oncol. 2012, 7, 213. [Google Scholar] [CrossRef]
- Chen, Y.; Li, X.; Guo, L.; Wu, X.; He, C.; Zhang, S.; Xiao, Y.; Yang, Y.; Hao, D. Combining Radiation with Autophagy Inhibition Enhances Suppression of Tumor Growth and Angiogenesis in Esophageal Cancer. Mol. Med. Rep. 2015, 12, 1645–1652. [Google Scholar] [CrossRef]
- Liu, E.Y.; Xu, N.; O’Prey, J.; Lao, L.Y.; Joshi, S.; Long, J.S.; O’Prey, M.; Croft, D.R.; Beaumatin, F.; Baudot, A.D.; et al. Loss of Autophagy Causes a Synthetic Lethal Deficiency in DNA Repair. Proc. Natl. Acad. Sci. USA 2015, 112, 773–778. [Google Scholar] [CrossRef]
- Yao, C.; Wu, S.; Kong, J.; Sun, Y.; Bai, Y.; Zhu, R.; Li, Z.; Sun, W.; Zheng, L. Angiogenesis in Hepatocellular Carcinoma: Mechanisms and Anti-Angiogenic Therapies. Cancer Biol. Med. 2023, 20, 25–43. [Google Scholar] [CrossRef]
- Jalouli, M. Emerging Role of Hypoxia-Inducible Factors (HIFs) in Modulating Autophagy: Perspectives on Cancer Therapy. Int. J. Mol. Sci. 2025, 26, 1752. [Google Scholar] [CrossRef]
- Benjamin, D.; Colombi, M.; Moroni, C.; Hall, M.N. Rapamycin Passes the Torch: A New Generation of mTOR Inhibitors. Nat. Rev. Drug Discov. 2011, 10, 868–880. [Google Scholar] [CrossRef] [PubMed]
- Zaytseva, Y.Y.; Valentino, J.D.; Gulhati, P.; Evers, B.M. mTOR Inhibitors in Cancer Therapy. Cancer Lett. 2012, 319, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Deleyto-Seldas, N.; Efeyan, A. The mTOR–Autophagy Axis and the Control of Metabolism. Front. Cell Dev. Biol. 2021, 9, 655731. [Google Scholar] [CrossRef]
- Buckley, C.E.; O’Brien, R.M.; Nugent, T.S.; Donlon, N.E.; O’Connell, F.; Reynolds, J.V.; Hafeez, A.; O’Ríordáin, D.S.; Hannon, R.A.; Neary, P.; et al. Metformin Is a Metabolic Modulator and Radiosensitiser in Rectal Cancer. Front. Oncol. 2023, 13, 1216911. [Google Scholar] [CrossRef]
- Sato, A.; Sunayama, J.; Okada, M.; Watanabe, E.; Seino, S.; Shibuya, K.; Suzuki, K.; Narita, Y.; Shibui, S.; Kayama, T.; et al. Glioma-Initiating Cell Elimination by Metformin Activation of FOXO3 via AMPK. Stem Cells Transl. Med. 2012, 1, 811–824. [Google Scholar] [CrossRef]
- Lee, J.W.; Park, S.; Takahashi, Y.; Wang, H.-G. The Association of AMPK with ULK1 Regulates Autophagy. PLoS ONE 2010, 5, e15394. [Google Scholar] [CrossRef]
- Shchors, K.; Massaras, A.; Hanahan, D. Dual Targeting of the Autophagic Regulatory Circuitry in Gliomas with Repurposed Drugs Elicits Cell-Lethal Autophagy and Therapeutic Benefit. Cancer Cell 2015, 28, 456–471. [Google Scholar] [CrossRef]
- Wong, P.-M.; Puente, C.; Ganley, I.G.; Jiang, X. The ULK1 Complex. Autophagy 2013, 9, 124–137. [Google Scholar] [CrossRef]
- García-López, D.; Zaragoza-Ojeda, M.; Eguía-Aguilar, P.; Arenas-Huertero, F. Endoplasmic Reticulum Stress in Gliomas: Exploiting a Dual-Effect Dysfunction through Chemical Pharmaceutical Compounds and Natural Derivatives for Therapeutical Uses. Int. J. Mol. Sci. 2024, 25, 4078. [Google Scholar] [CrossRef]
- Yang, R.; Dong, H.; Jia, S.; Yang, Z. Resveratrol as a Modulatory of Apoptosis and Autophagy in Cancer Therapy. Clin. Transl. Oncol. 2022, 24, 1219–1230. [Google Scholar] [CrossRef]
- Li, J.; Fan, Y.; Zhang, Y.; Liu, Y.; Yu, Y.; Ma, M. Resveratrol Induces Autophagy and Apoptosis in Non-Small-Cell Lung Cancer Cells by Activating the NGFR-AMPK-mTOR Pathway. Nutrients 2022, 14, 2413. [Google Scholar] [CrossRef] [PubMed]
- Pietrocola, F.; Lachkar, S.; Enot, D.P.; Niso-Santano, M.; Bravo-San Pedro, J.M.; Sica, V.; Izzo, V.; Maiuri, M.C.; Madeo, F.; Mariño, G.; et al. Spermidine Induces Autophagy by Inhibiting the Acetyltransferase EP300. Cell Death Differ. 2015, 22, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Shu, F.; Xiao, H.; Li, Q.-N.; Ren, X.-S.; Liu, Z.-G.; Hu, B.-W.; Wang, H.-S.; Wang, H.; Jiang, G.-M. Epigenetic and Post-Translational Modifications in Autophagy: Biological Functions and Therapeutic Targets. Signal Transduct. Target. Ther. 2023, 8, 32. [Google Scholar] [CrossRef]
- Prasher, P.; Sharma, M.; Singh, S.K.; Gulati, M.; Chellappan, D.K.; Rajput, R.; Gupta, G.; Ydyrys, A.; Kulbayeva, M.; Abdull Razis, A.F.; et al. Spermidine as a Promising Anticancer Agent: Recent Advances and Newer Insights on Its Molecular Mechanisms. Front. Chem. 2023, 11, 1164477. [Google Scholar] [CrossRef]
- Schwarz, C.; Stekovic, S.; Wirth, M.; Benson, G.; Royer, P.; Sigrist, S.J.; Pieber, T.; Dammbrueck, C.; Magnes, C.; Eisenberg, T.; et al. Safety and Tolerability of Spermidine Supplementation in Mice and Older Adults with Subjective Cognitive Decline. Aging 2018, 10, 19–33. [Google Scholar] [CrossRef]
- Moro, T.; Tinsley, G.; Bianco, A.; Marcolin, G.; Pacelli, Q.F.; Battaglia, G.; Palma, A.; Gentil, P.; Neri, M.; Paoli, A. Effects of Eight Weeks of Time-Restricted Feeding (16/8) on Basal Metabolism, Maximal Strength, Body Composition, Inflammation, and Cardiovascular Risk Factors in Resistance-Trained Males. J. Transl. Med. 2016, 14, 290. [Google Scholar] [CrossRef]
- Wei, M.; Brandhorst, S.; Shelehchi, M.; Mirzaei, H.; Cheng, C.W.; Budniak, J.; Groshen, S.; Mack, W.J.; Guen, E.; Di Biase, S.; et al. Fasting-Mimicking Diet and Markers/Risk Factors for Aging, Diabetes, Cancer, and Cardiovascular Disease. Sci. Transl. Med. 2017, 9, eaai8700. [Google Scholar] [CrossRef]
- Chaix, A.; Zarrinpar, A.; Miu, P.; Panda, S. Time-Restricted Feeding Is a Preventative and Therapeutic Intervention against Diverse Nutritional Challenges. Cell Metab. 2014, 20, 991–1005. [Google Scholar] [CrossRef]
- Carter, S.; Clifton, P.M.; Keogh, J.B. The Effects of Intermittent Compared to Continuous Energy Restriction on Glycaemic Control in Type 2 Diabetes; a Pragmatic Pilot Trial. Diabetes Res. Clin. Pract. 2016, 122, 106–112. [Google Scholar] [CrossRef]
- Belkacemi, L.; Selselet-Attou, G.; Hupkens, E.; Nguidjoe, E.; Louchami, K.; Sener, A.; Malaisse, W.J. Intermittent Fasting Modulation of the Diabetic Syndrome in Streptozotocin-Injected Rats. Int. J. Endocrinol. 2012, 2012, 962012. [Google Scholar] [CrossRef]
- Anson, R.M.; Guo, Z.; de Cabo, R.; Iyun, T.; Rios, M.; Hagepanos, A.; Ingram, D.K.; Lane, M.A.; Mattson, M.P. Intermittent Fasting Dissociates Beneficial Effects of Dietary Restriction on Glucose Metabolism and Neuronal Resistance to Injury from Calorie Intake. Proc. Natl. Acad. Sci. USA 2003, 100, 6216–6220. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, I.; Higami, Y.; Utsuyama, M.; Tuchiya, T.; Komatsu, T.; Chiba, T.; Yamaza, H. Life Span Extension by Reduction in Growth Hormone-Insulin-Like Growth Factor-1 Axis in a Transgenic Rat Model. Am. J. Pathol. 2002, 160, 2259–2265. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Kroemer, G. Biological Functions of Autophagy Genes: A Disease Perspective. Cell 2019, 176, 11–42. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kundu, M.; Viollet, B.; Guan, K.-L. AMPK and mTOR Regulate Autophagy through Direct Phosphorylation of Ulk1. Nat. Cell Biol. 2011, 13, 132–141. [Google Scholar] [CrossRef]
- Liu, Q.; Guan, J.-Z.; Sun, Y.; Le, Z.; Zhang, P.; Yu, D.; Liu, Y. Insulin-like Growth Factor 1 Receptor-Mediated Cell Survival in Hypoxia Depends on the Promotion of Autophagy via Suppression of the PI3K/Akt/mTOR Signaling Pathway. Mol. Med. Rep. 2017, 15, 2136–2142. [Google Scholar] [CrossRef]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of Metabolism and Mitochondrial Homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef]
- Kaushik, S.; Cuervo, A.M. Chaperone-Mediated Autophagy: A Unique Way to Enter the Lysosome World. Trends Cell Biol. 2012, 22, 407–417. [Google Scholar] [CrossRef]
- Pickles, S.; Vigié, P.; Youle, R.J. The Art of Mitochondrial Maintenance. Curr. Biol. 2018, 28, R170–R185. [Google Scholar] [CrossRef]
- Lee, C.; Raffaghello, L.; Brandhorst, S.; Safdie, F.M.; Bianchi, G.; Martin-Montalvo, A.; Pistoia, V.; Wei, M.; Hwang, S.; Merlino, A.; et al. Fasting Cycles Retard Growth of Tumors and Sensitize a Range of Cancer Cell Types to Chemotherapy. Sci. Transl. Med. 2012, 4, ra27–ra124. [Google Scholar] [CrossRef]
- Kroemer, G.; Mariño, G.; Levine, B. Autophagy and the Integrated Stress Response. Mol. Cell 2010, 40, 280–293. [Google Scholar] [CrossRef]
- Bianchi, G.; Martella, R.; Ravera, S.; Marini, C.; Capitanio, S.; Orengo, A.; Emionite, L.; Lavarello, C.; Amaro, A.; Petretto, A.; et al. Fasting Induces Anti-Warburg Effect That Increases Respiration but Reduces ATP-Synthesis to Promote Apoptosis in Colon Cancer Models. Oncotarget 2015, 6, 11806–11819. [Google Scholar] [CrossRef] [PubMed]
- Antunes, F.; Erustes, A.G.; Costa, A.J.; Nascimento, A.C.; Bincoletto, C.; Ureshino, R.P.; Pereira, G.J.S.; Smaili, S.S. Autophagy and Intermittent Fasting: The Connection for Cancer Therapy? Clinics 2018, 73, e814s. [Google Scholar] [CrossRef] [PubMed]
- D’Arcy, M.S. Cell Death: A Review of the Major Forms of Apoptosis, Necrosis and Autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef] [PubMed]
- de Cabo, R.; Mattson, M.P. Effects of Intermittent Fasting on Health, Aging, and Disease. N. Engl. J. Med. 2019, 381, 2541–2551. [Google Scholar] [CrossRef]
- Longo, V.D.; Mattson, M.P. Fasting: Molecular Mechanisms and Clinical Applications. Cell Metab. 2014, 19, 181–192. [Google Scholar] [CrossRef]
- Intermittent Fasting (IF) Reduces Tumor Metastasis Via Ras/MAPK, PI3K/AKT/mTOR, Wnt/β-Catenin, and HGF/c-Met Pathways. Available online: https://www.scirp.org/journal/paperinformation?paperid=138288 (accessed on 6 March 2025).
- Vidoni, C.; Ferraresi, A.; Esposito, A.; Maheshwari, C.; Dhanasekaran, D.N.; Mollace, V.; Isidoro, C. Calorie Restriction for Cancer Prevention and Therapy: Mechanisms, Expectations, and Efficacy. J. Cancer Prev. 2021, 26, 224–236. [Google Scholar] [CrossRef]
- Rubinsztein, D.C.; Mariño, G.; Kroemer, G. Autophagy and Aging. Cell 2011, 146, 682–695. [Google Scholar] [CrossRef]
- Alirezaei, M.; Kemball, C.C.; Flynn, C.T.; Wood, M.R.; Whitton, J.L.; Kiosses, W.B. Short-Term Fasting Induces Profound Neuronal Autophagy. Autophagy 2010, 6, 702–710. [Google Scholar] [CrossRef]
- Raffaghello, L.; Lee, C.; Safdie, F.M.; Wei, M.; Madia, F.; Bianchi, G.; Longo, V.D. Starvation-Dependent Differential Stress Resistance Protects Normal but Not Cancer Cells against High-Dose Chemotherapy. Proc. Natl. Acad. Sci. USA 2008, 105, 8215–8220. [Google Scholar] [CrossRef]
- Thomas, J.A.; Antonelli, J.A.; Lloyd, J.C.; Masko, E.M.; Poulton, S.H.; Phillips, T.E.; Pollak, M.; Freedland, S.J. Effect of Intermittent Fasting on Prostate Cancer Tumor Growth in a Mouse Model. Prostate Cancer Prostatic Dis. 2010, 13, 350–355. [Google Scholar] [CrossRef]
- Lo Re, O.; Panebianco, C.; Porto, S.; Cervi, C.; Rappa, F.; Di Biase, S.; Caraglia, M.; Pazienza, V.; Vinciguerra, M. Fasting Inhibits Hepatic Stellate Cells Activation and Potentiates Anti-Cancer Activity of Sorafenib in Hepatocellular Cancer Cells. J. Cell Physiol. 2018, 233, 1202–1212. [Google Scholar] [CrossRef] [PubMed]
- Madeo, F.; Pietrocola, F.; Eisenberg, T.; Kroemer, G. Caloric Restriction Mimetics: Towards a Molecular Definition. Nat. Rev. Drug Discov. 2014, 13, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Wang, H.; He, Z.; Chen, X.; Wu, Q.; Chen, W.; Sun, Z.; Weng, M.; Zhu, M.; Ma, D.; et al. Fasting Inhibits Colorectal Cancer Growth by Reducing M2 Polarization of Tumor-Associated Macrophages. Oncotarget 2017, 8, 74649–74660. [Google Scholar] [CrossRef] [PubMed]
- Pietrocola, F.; Pol, J.; Kroemer, G. Fasting Improves Anticancer Immunosurveillance via Autophagy Induction in Malignant Cells. Cell Cycle 2016, 15, 3327–3328. [Google Scholar] [CrossRef]
- Cheng, C.-W.; Adams, G.B.; Perin, L.; Wei, M.; Zhou, X.; Lam, B.S.; Da Sacco, S.; Mirisola, M.; Quinn, D.I.; Dorff, T.B.; et al. Prolonged Fasting Reduces IGF-1/PKA to Promote Hematopoietic-Stem-Cell-Based Regeneration and Reverse Immunosuppression. Cell Stem Cell 2014, 14, 810–823. [Google Scholar] [CrossRef]
- Brandhorst, S.; Choi, I.Y.; Wei, M.; Cheng, C.W.; Sedrakyan, S.; Navarrete, G.; Dubeau, L.; Yap, L.P.; Park, R.; Vinciguerra, M.; et al. A Periodic Diet That Mimics Fasting Promotes Multi-System Regeneration, Enhanced Cognitive Performance, and Healthspan. Cell Metab. 2015, 22, 86–99. [Google Scholar] [CrossRef]
- de Groot, S.; Pijl, H.; van der Hoeven, J.J.M.; Kroep, J.R. Effects of Short-Term Fasting on Cancer Treatment. J. Exp. Clin. Cancer Res. 2019, 38, 209. [Google Scholar] [CrossRef]
- Rieger, J.; Bähr, O.; Maurer, G.D.; Hattingen, E.; Franz, K.; Brucker, D.; Walenta, S.; Kämmerer, U.; Coy, J.F.; Weller, M.; et al. ERGO: A Pilot Study of Ketogenic Diet in Recurrent Glioblastoma. Int. J. Oncol. 2014, 44, 1843–1852. [Google Scholar] [CrossRef]
- Redding, A.; Santarossa, S.; Murphy, D.; Udumula, M.P.; Munkarah, A.; Hijaz, M.; Rattan, R. A Patient Perspective on Applying Intermittent Fasting in Gynecologic Cancer. BMC Res. Notes 2023, 16, 190. [Google Scholar] [CrossRef]
- Schreck, K.C.; Hsu, F.-C.; Berrington, A.; Henry-Barron, B.; Vizthum, D.; Blair, L.; Kossoff, E.H.; Easter, L.; Whitlow, C.T.; Barker, P.B.; et al. Feasibility and Biological Activity of a Ketogenic/Intermittent-Fasting Diet in Patients With Glioma. Neurology 2021, 97, e953–e963. [Google Scholar] [CrossRef]
- Vernieri, C.; Fucà, G.; Ligorio, F.; Huber, V.; Vingiani, A.; Iannelli, F.; Raimondi, A.; Rinchai, D.; Frigè, G.; Belfiore, A.; et al. Fasting-Mimicking Diet Is Safe and Reshapes Metabolism and Antitumor Immunity in Patients with Cancer. Cancer Discov. 2022, 12, 90–107. [Google Scholar] [CrossRef] [PubMed]
- O’Flanagan, C.H.; Smith, L.A.; McDonell, S.B.; Hursting, S.D. When Less May Be More: Calorie Restriction and Response to Cancer Therapy. BMC Med. 2017, 15, 106. [Google Scholar] [CrossRef] [PubMed]
- Baracos, V.E.; Martin, L.; Korc, M.; Guttridge, D.C.; Fearon, K.C.H. Cancer-Associated Cachexia. Nat. Rev. Dis. Primers. 2018, 4, 17105. [Google Scholar] [CrossRef] [PubMed]
- Kalam, F.; James, D.L.; Li, Y.R.; Coleman, M.F.; Kiesel, V.A.; Cespedes Feliciano, E.M.; Hursting, S.D.; Sears, D.D.; Kleckner, A.S. Intermittent Fasting Interventions to Leverage Metabolic and Circadian Mechanisms for Cancer Treatment and Supportive Care Outcomes. JNCI Monogr. 2023, 2023, 84–103. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Primary Therapy Strategy | Diet Type | Therapeutic Effect | Study Population | Number of Participants | Study |
|---|---|---|---|---|---|---|
| various types | various standard types—chemotherapy, radiotherapy, surgery | intermittent fasting (2 days per week) | reduction in IGF-1 levels, improved quality of life | patients with multiple cancer types | 10 | [6] |
| breast cancer | no therapy before surgery | fasting-mimicking diet (FMD) for 5 days per month | reduction in glucose and insulin levels, potential autophagy activation | women with breast cancer in the preoperative period | 34 | [119] |
| glioma | radiotherapy | intermittent fasting (50% calorie restriction every other day) | disease stabilization, improved response to radiotherapy | patients with glioma | 8 | [120] |
| gynecological cancers | chemotherapy—carboplatin, paclitaxel | intermittent fasting (16:8) | improved well-being and reduction in side effects of oncological therapy | women with gynecological cancers | 47 | [121] |
| glioma | radiotherapy; chemotherapy—temozolomide | ketogenic diet/intermittent fasting | in some cases, slowed tumor progression | patients with glioma | 25 | [122] |
| various cancer types | various standard treatments | caloric restriction and time-restricted feeding (TRF) | lowered blood glucose (by 18.6%), insulin (by 50.7%), and IGF-1 (by 30.3%); increased activation of CD8+ T cells | patients with multiple cancer types | 101 | [123] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wolska, W.; Gutowska, I.; Wszołek, A.; Żwierełło, W. The Role of Intermittent Fasting in the Activation of Autophagy Processes in the Context of Cancer Diseases. Int. J. Mol. Sci. 2025, 26, 4742. https://doi.org/10.3390/ijms26104742
Wolska W, Gutowska I, Wszołek A, Żwierełło W. The Role of Intermittent Fasting in the Activation of Autophagy Processes in the Context of Cancer Diseases. International Journal of Molecular Sciences. 2025; 26(10):4742. https://doi.org/10.3390/ijms26104742
Chicago/Turabian StyleWolska, Waleria, Izabela Gutowska, Agata Wszołek, and Wojciech Żwierełło. 2025. "The Role of Intermittent Fasting in the Activation of Autophagy Processes in the Context of Cancer Diseases" International Journal of Molecular Sciences 26, no. 10: 4742. https://doi.org/10.3390/ijms26104742
APA StyleWolska, W., Gutowska, I., Wszołek, A., & Żwierełło, W. (2025). The Role of Intermittent Fasting in the Activation of Autophagy Processes in the Context of Cancer Diseases. International Journal of Molecular Sciences, 26(10), 4742. https://doi.org/10.3390/ijms26104742