

Integrated Transcriptomic and Metabolomic Analysis of Exogenous NAA Effects on Maize Seedling Root Systems under Potassium Deficiency

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Root Growth and Development
2.2. Root Growth and K+ Uptake
2.3. The Transcriptomics of 90-21-3 and D937 Response to Exogenous NAA under K-Deficiency
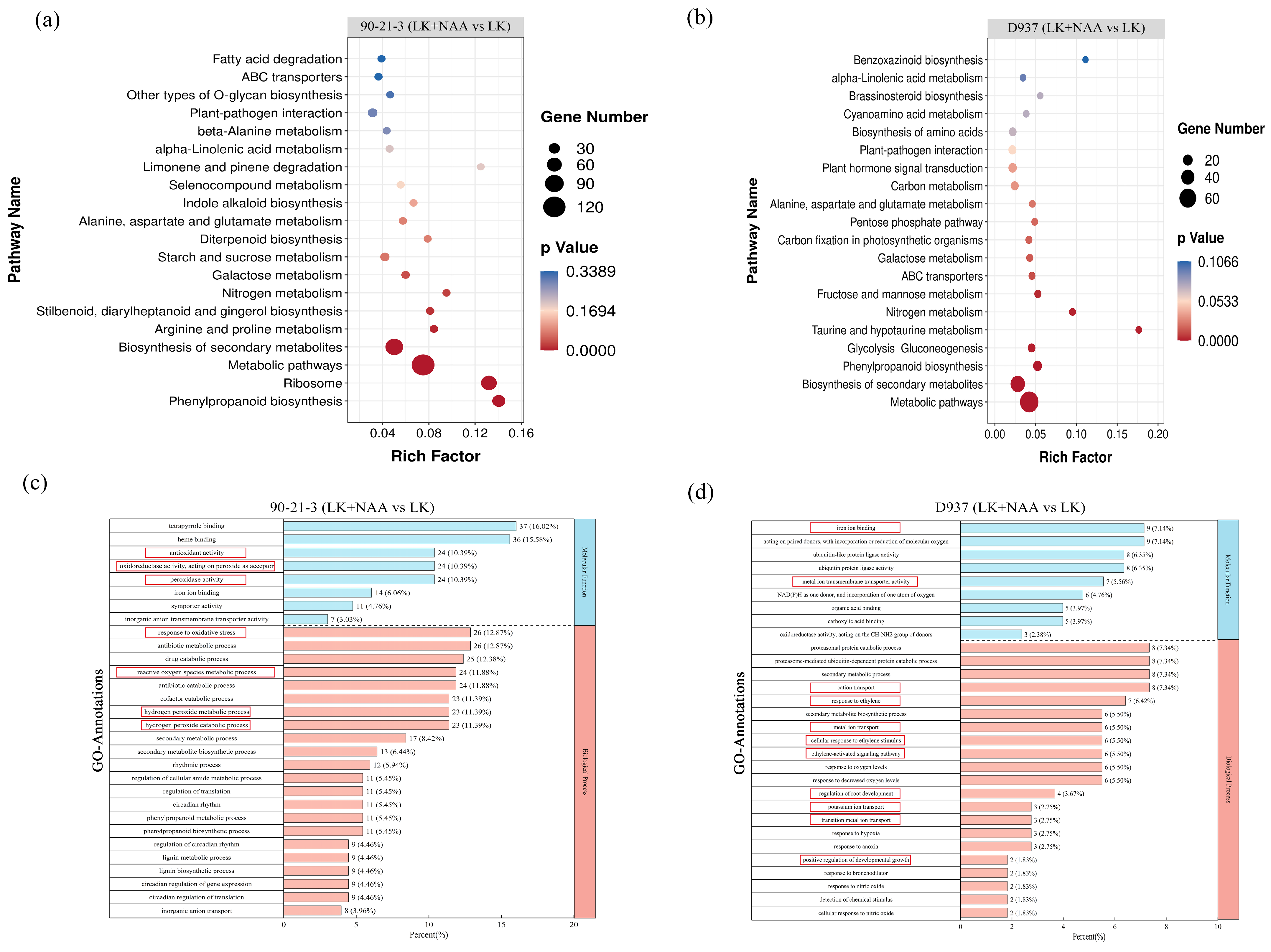
2.4. Enrichment Analysis of Differentially Expressed Genes
2.5. Quantitative Real-Time PCR Validation
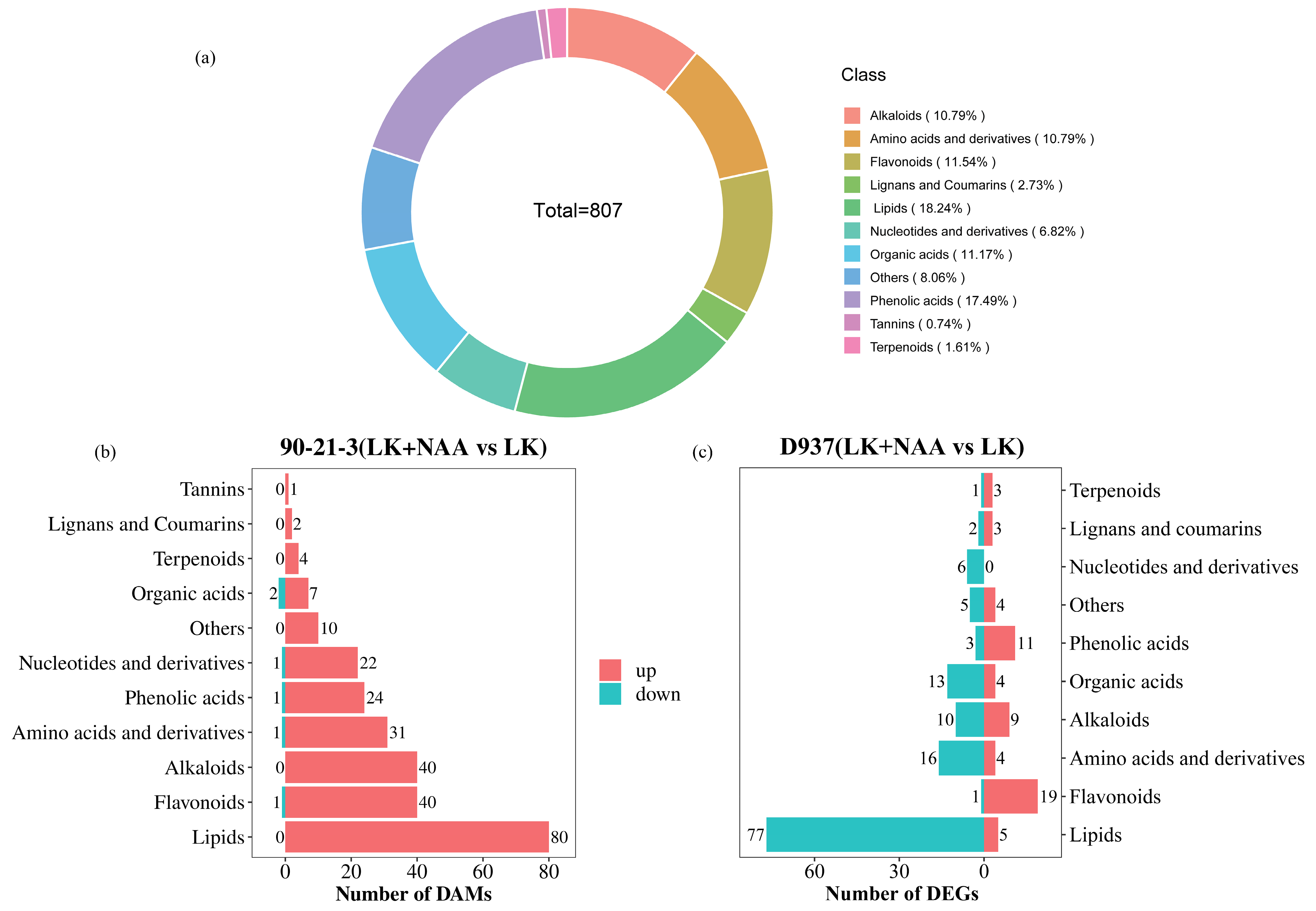
2.6. Metabolomic Analysis of 90-21-3 and D937 in Response to Exogenous NAA under K-Deficiency
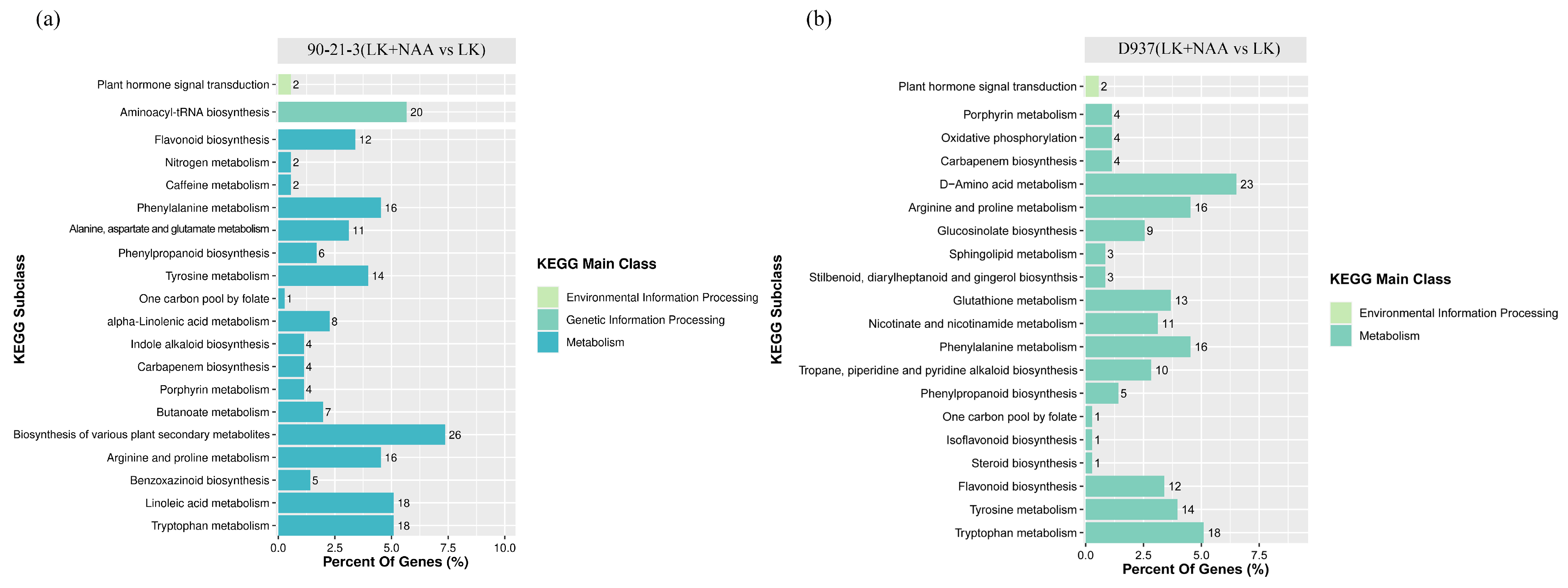
2.7. Enrichment Analysis of Differentially Accumulated Metabolites
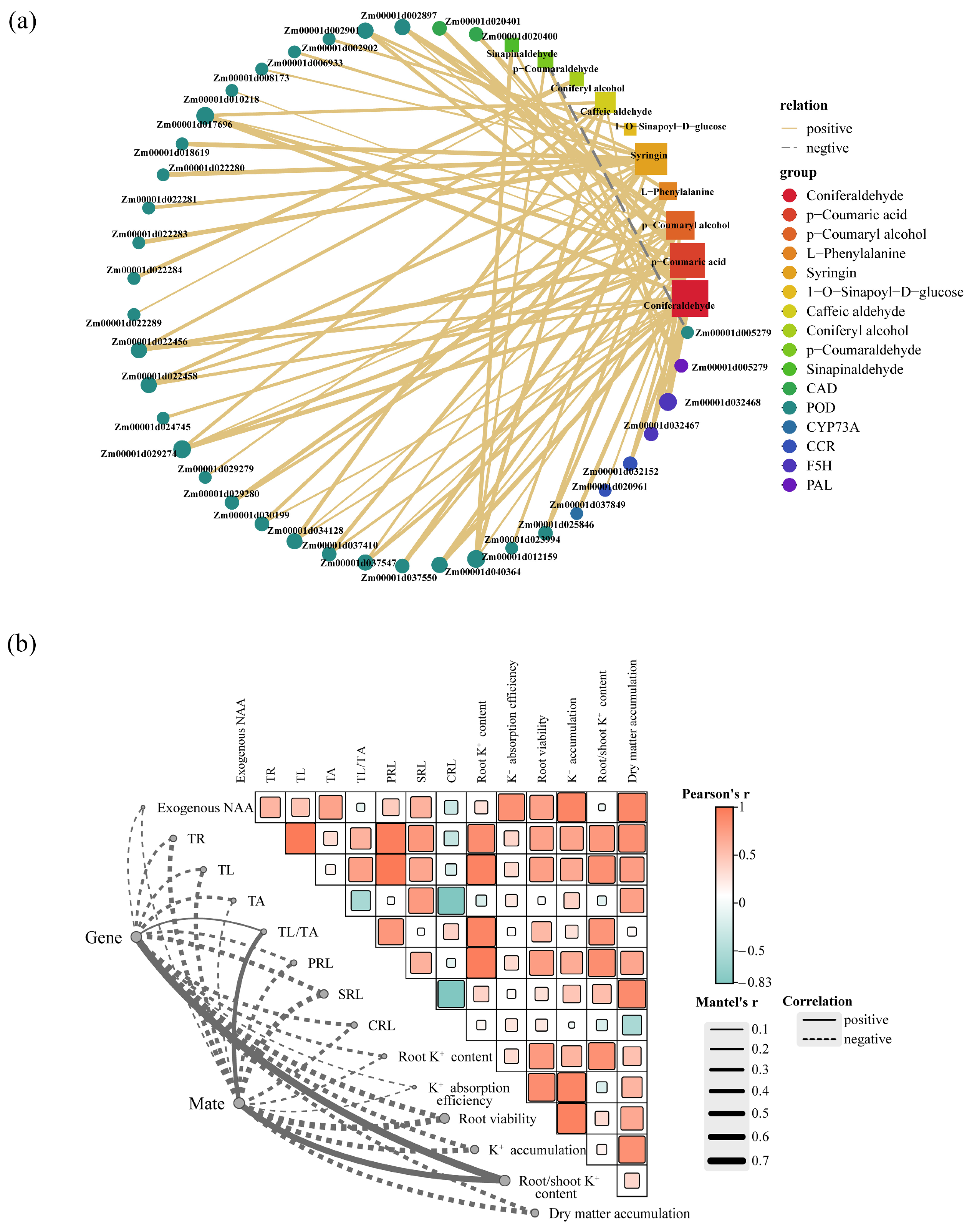
2.8. Combined Analysis of Transcriptome and Metabolome
3. Discussion
3.1. Effects of Exogenous NAA on Maize Root Growth and K+ Uptake under K-Deficiency
3.2. Effects of Exogenous NAA on Gene Expression and Metabolite Accumulation in Maize Roots
3.3. Regulation of Exogenous NAA on Phenylpropanoid Biosynthesis Pathway under K-Deficiency
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Plant Sampling
4.3. Potassium Content Determination
4.4. Root Viability
4.5. Transcriptome Determination and Analysis
4.6. Metabolome Determination and Analysis
4.7. qPCR
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, R.; Vishwakarma, K.; Hossen, M.S.; Kumar, V.; Shackira, A.M.; Puthur, J.T.; Abdi, G.; Sarraf, M.; Hasanuzzaman, M. Potassium in Plants: Growth Regulation, Signaling, and Environmental Stress Tolerance. Plant Physiol. Biochem. 2022, 172, 56–69. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Rahman, M.M.; Ghosh, T.K.; Kabir, A.H.; Abdelrahman, M.; Khan, M.A.R.; Mochida, K.; Tran, L.P. Potassium in Plant Physiological Adaptation to Abiotic Stresses. Plant Physiol. Biochem. 2022, 186, 279–289. [Google Scholar] [CrossRef]
- Schroeder, J.I.; Ward, J.M.; Gassmann, W. Perspectives on the Physiology and Structure of Inward-rectifying K+ Channels in Higher Plants: Biophysical Implications for K+ Uptake. Annu. Rev. Biophys. Biomol. Struct. 1994, 23, 441–471. [Google Scholar] [CrossRef]
- Maathuis, F.J. Physiological Functions of Mineral Macronutrients. Curr. Opin. Plant Biol. 2009, 12, 250–258. [Google Scholar] [CrossRef]
- Rodríguez-Navarro, A. Potassium Transport in Fungi and Plants. Biochim. Biophys. Acta (BBA) Biomembr. 2000, 1469, 1–30. [Google Scholar] [CrossRef]
- Lynch, J.P. Root Phenes for Enhanced Soil Exploration and Phosphorus Acquisition: Tools for Future Crops. Plant Physiol. 2011, 156, 1041–1049. [Google Scholar] [CrossRef]
- Xu, G.; Fan, X.; Miller, A.J. Plant Nitrogen Assimilation and Use Efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef]
- White, P.J.; Bell, M.J.; Djalovic, I.; Hinsinger, P.; Rengel, Z. Potassium Use Efficiency of Plants. In Improving Potassium Recommendations for Agricultural Crops; Murrell, T.S., Mikkelsen, R.L., Sulewski, G., Norton, R., Thompson, M.L., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 119–145. [Google Scholar]
- Mei, Y.; Jia, W.J.; Chu, Y.J.; Xue, H.W. Arabidopsis Phosphatidylinositol Monophosphate 5-kinase 2 is Involved in Root Gravitropism Through Regulation of Polar Auxin Transport by Affecting the Cycling of PIN Proteins. Cell Res. 2012, 22, 581–597. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, W.-H. Potassium Transport and Signaling in Higher Plants. Annu. Rev. Plant Biol. 2013, 64, 451–476. [Google Scholar] [CrossRef]
- Walker, D.J.; Leigh, R.A.; Miller, A.J. Potassium Homeostasis in Vacuolate Plant Cells. Proc. Natl. Acad. Sci. USA 1996, 93, 10510–10514. [Google Scholar] [CrossRef]
- Tang, R.-J.; Zhao, F.-G.; Yang, Y.; Wang, C.; Li, K.; Kleist, T.J.; Lemaux, P.G.; Luan, S. A Calcium Signalling Network Activates Vacuolar K+ Remobilization to Enable Plant Adaptation to Low-K Environments. Nat. Plants 2020, 6, 384–393. [Google Scholar] [CrossRef]
- Sustr, M.; Soukup, A.; Tylova, E. Potassium in Root Growth and Development. Plants 2019, 8, 435. [Google Scholar] [CrossRef]
- Wang, C.; Chen, H.F.; Hao, Q.N.; Sha, A.H.; Shan, Z.H.; Chen, L.M.; Zhou, R.; Zhi, H.J.; Zhou, X.A. Transcript Profile of the Response of Two Soybean Genotypes to Potassium Deficiency. PLoS ONE 2012, 7, e39856. [Google Scholar] [CrossRef]
- Ruan, L.; Zhang, J.B.; Xin, X.L.; Zhang, C.Z.; Ma, D.H.; Chen, L.; Zhao, B.Z. Comparative Analysis of Potassium Deficiency-Responsive Transcriptomes in Low Potassium Susceptible and Tolerant Wheat (Triticum aestivum L.). Sci. Rep. 2015, 5, 10090. [Google Scholar] [CrossRef]
- Pasternak, T.P.; Steinmacher, D. Plant Growth Regulation in Cell and Tissue Culture In Vitro. Plants 2024, 13, 327. [Google Scholar] [CrossRef]
- Roychoudhry, S.; Kepinski, S. Auxin in Root Development. Cold Spring Harb. Perspect. Biol. 2021, 14, a039933. [Google Scholar] [CrossRef]
- Edelmann, H.G. Plant Root Development: Is the Classical Theory for Auxin-regulated Root Growth False? Protoplasma 2022, 259, 823–832. [Google Scholar] [CrossRef]
- Claussen, M.; Lüthe, H.; Blatt, M.; Böttger, M. Auxin-induced Growth and Its Linkage to Potassium Channels. Planta 1997, 201, 227–234. [Google Scholar] [CrossRef]
- Song, Z.; Wu, X.; Gao, Y.; Cui, X.; Jiao, F.; Chen, X.; Li, Y. Genome-wide Analysis of the HAK Potassium Transporter Gene Family Reveals Asymmetrical Evolution in Tobacco (Nicotiana tabacum). Genome 2019, 62, 267–278. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, M.-L.; Ma, T.-L.; Wang, Y. Phosphorylation of ARF2 Relieves Its Repression of Transcription of the K+ Transporter Gene HAK5 in Response to Low Potassium Stress. Plant Cell 2016, 28, 3005–3019. [Google Scholar] [CrossRef]
- Li, W.; Xu, G.; Alli, A.; Yu, L. Plant HAK/KUP/KT K+ Transporters: Function and Regulation. Semin. Cell Dev. Biol. 2017, 74, 133–141. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, Q.; Jin, R.; Zhao, P.; Zhu, X.; Wang, J.; Yu, Y.; Tang, Z. The Role of IAA in Regulating Root Architecture of Sweetpotato (Ipomoea batatas [L.] Lam) in Response to Potassium Deficiency Stress. Plants 2023, 12, 1779. [Google Scholar] [CrossRef]
- Zhao, X.-h.; Yu, H.-q.; Wen, J.; Wang, X.-g.; Du, Q.; Wang, J.; Wang, Q. Response of Root Morphology, Physiology and Endogenous Hormones in Maize (Zea mays L.) to Potassium Deficiency. J. Integr. Agric. 2016, 15, 785–794. [Google Scholar] [CrossRef]
- Yang, L.; Luo, S.; Wu, Z.; Rong, X.; Han, Y. Low Nitrogen Stress Stimulated Nitrate Uptake Rate Modulated by Auxin in Brassica napus L. J. Soil Sci. Plant Nutr. 2022, 22, 3500–3506. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, B.; Zhao, Y.; Zhang, J.; Li, Z. Auxin and GA Signaling Play Important Roles in the Maize Response to Phosphate Deficiency. Plant Sci. 2019, 283, 177–188. [Google Scholar] [CrossRef]
- Hu, S.; Zhang, M.; Yang, Y.; Xuan, W.; Zou, Z.; Arkorful, E.; Chen, Y.; Ma, Q.; Jeyaraj, A.; Chen, X.; et al. A Novel Insight into Nitrogen and Auxin Signaling in Lateral Root Formation in Tea Plant [Camellia sinensis (L.) O. Kuntze]. BMC Plant Biol. 2020, 20, 232. [Google Scholar] [CrossRef]
- Nacry, P.; Canivenc, G.; Muller, B.; Azmi, A.; Van Onckelen, H.; Rossignol, M.; Doumas, P. A Role for Auxin Redistribution in the Responses of the Root System Architecture to Phosphate Starvation in Arabidopsis. Plant Physiol. 2005, 138, 2061–2074. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Shen, J.; Zhang, F.; Rengel, Z. Interactive Effects of Phosphorus Deficiency and Exogenous Auxin on Root Morphological and Physiological Traits in White Lupin (Lupinus albus L.). Sci. China Life Sci. 2013, 56, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Overvoorde, P.; Fukaki, H.; Beeckman, T. Auxin Control of Root Development. Cold Spring Harb. Perspect. Biol. 2010, 2, a001537. [Google Scholar] [CrossRef]
- Grieneisen, V.A.; Xu, J.; Marée, A.F.M.; Hogeweg, P.; Scheres, B. Auxin Transport is Sufficient to Generate a Maximum and Gradient Guiding Root Growth. Nature 2007, 449, 1008–1013. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Scheres, B. Lateral Root Formation and the Multiple Roles of Auxin. J. Exp. Bot. 2017, 69, 155–167. [Google Scholar] [CrossRef]
- Rigas, S.; Debrosses, G.; Haralampidis, K.; Vicente-Agullo, F.; Feldmann, K.A.; Grabov, A.; Dolan, L.; Hatzopoulos, P. TRH1 Encodes a Potassium Transporter Required for Tip Growth in Arabidopsis Root Hairs. Plant Cell 2001, 13, 139–151. [Google Scholar] [CrossRef]
- Cao, Y.; Glass, A.D.; Crawford, N.M. Ammonium Inhibition of Arabidopsis Root Growth Can be Reversed by Potassium and by Auxin Resistance Mutations aux1, axr1, and axr2. Plant Physiol. 1993, 102, 983–989. [Google Scholar] [CrossRef][Green Version]
- Li, J.; Wu, W.-H.; Wang, Y. Potassium Channel AKT1 is Involved in the Auxin-mediated Root Growth Inhibition in Arabidopsis Response to Low K+ stress. J. Integr. Plant Biol. 2017, 59, 895–909. [Google Scholar] [CrossRef]
- Chen, J.; Gabelman, W.H. Morphological and Physiological Characteristics of Tomato Roots Associated with Potassium-acquisition Efficiency. Sci. Hortic. 2000, 83, 213–225. [Google Scholar] [CrossRef]
- Song, W.; Xue, R.; Song, Y.; Bi, Y.; Liang, Z.; Meng, L.; Dong, C.; Wang, C.; Liu, G.; Dong, J.; et al. Differential Response of First-Order Lateral Root Elongation to Low Potassium Involves Nitric Oxide in Two Tobacco Cultivars. J. Plant Growth Regul. 2018, 37, 114–127. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, R.; Zhao, S.; Lu, C.; Zhu, Z.; Li, H. Transporter NRT1.5/NPF7.3 Suppresses Primary Root Growth under Low K+ Stress by Regulating the Degradation of PIN-FORMED2. BMC Plant Biol. 2022, 22, 330. [Google Scholar] [CrossRef]
- Song, W.; Liu, S.; Meng, L.; Xue, R.; Wang, C.; Liu, G.; Dong, C.; Wang, S.; Dong, J.; Zhang, Y. Potassium Deficiency Inhibits Lateral Root Development in Tobacco Seedlings by Changing Auxin Distribution. Plant Soil 2015, 396, 163–173. [Google Scholar] [CrossRef]
- Viana, W.G.; Scharwies, J.D.; Dinneny, J.R. Deconstructing the Root System of Grasses Through an Exploration of Development, Anatomy and Function. Plant Cell Environ. 2022, 45, 602–619. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Agullo, F.; Rigas, S.; Desbrosses, G.; Dolan, L.; Hatzopoulos, P.; Grabov, A. Potassium Carrier TRH1 is Required for Auxin Transport in Arabidopsis Roots. Plant J. 2004, 40, 523–535. [Google Scholar] [CrossRef] [PubMed]
- Gruber, B.D.; Giehl, R.F.H.; Friedel, S.; von Wirén, N. Plasticity of the Arabidopsis Root System under Nutrient Deficiencies. Plant Physiol. 2013, 163, 161–179. [Google Scholar] [CrossRef]
- Pasternak, T.; Palme, K.; Pérez-Pérez, J.M. Role of Reactive Oxygen Species in the Modulation of Auxin Flux and Root Development in Arabidopsis thaliana. Plant J. 2023, 114, 83–95. [Google Scholar] [CrossRef]
- Jung, J.-Y.; Shin, R.; Schachtman, D.P. Ethylene Mediates Response and Tolerance to Potassium Deprivation in Arabidopsis. Plant Cell 2009, 21, 607–621. [Google Scholar] [CrossRef]
- Liu, M.; Yu, Y.; Jin, R.; Zhao, P.; Zhang, Q.; Zhu, X.; Wang, J.; Tang, Z. Exogenous IAA Enhances Low Potassium Tolerance of Sweet Potato by Regulating Root Response Strategy. Arch. Agron. Soil Sci. 2024, 70, 1–15. [Google Scholar] [CrossRef]
- Baskar, V.; Venkatesh, R.; Ramalingam, S. Flavonoids (Antioxidants Systems) in Higher Plants and Their Response to Stresses. In Antioxidants and Antioxidant Enzymes in Higher Plants; Gupta, D.K., Palma, J.M., Corpas, F.J., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 253–268. [Google Scholar]
- Shomali, A.; Das, S.; Arif, N.; Sarraf, M.; Zahra, N.; Yadav, V.; Aliniaeifard, S.; Chauhan, D.K.; Hasanuzzaman, M. Diverse Physiological Roles of Flavonoids in Plant Environmental Stress Responses and Tolerance. Plants 2022, 11, 3158. [Google Scholar] [CrossRef]
- Ismail, H.; Dragišic Maksimovic, J.; Maksimovic, V.; Shabala, L.; Živanovic, B.D.; Tian, Y.; Jacobsen, S.-E.; Shabala, S. Rutin, a Flavonoid with Antioxidant Activity, Improves Plant Salinity Tolerance by Regulating K+ Retention and Na+ Exclusion from Leaf Mesophyll in Quinoa and Broad Beans. Funct. Plant Biol. 2016, 43, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Cui, D.; Ye, Z.H. Secondary Cell Wall Biosynthesis. New Phytol. 2018, 221, 1703–1723. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Liu, F.; Xing, H.; Mao, K.; Chen, G.; Guo, Q.; Chen, J. Correlation Analysis of Lignin Accumulation and Expression of Key Genes Involved in Lignin Biosynthesis of Ramie (Boehmeria nivea). Genes 2019, 10, 389. [Google Scholar] [CrossRef]
- Piquemal, J.; Lapierre, C.; Myton, K.; O’connell, A.; Schuch, W.; Grima-pettenati, J.; Boudet, A.-M. Down-regulation of Cinnamoyl-CoA Reductase Induces Significant Changes of Lignin Profiles in Transgenic Tobacco Plants. Plant J. 1998, 13, 71–83. [Google Scholar] [CrossRef]
- Fan, L.; Linker, R.; Gepstein, S.; Tanimoto, E.; Yamamoto, R.; Neumann, P.M. Progressive Inhibition by Water Deficit of Cell Wall Extensibility and Growth Along the Elongation Zone of Maize Roots is Related to Increased Lignin Metabolism and Progressive Stelar Accumulation of Wall Phenolics. Plant Physiol. 2006, 140, 603–612. [Google Scholar] [CrossRef]
- García, J.R.; Anderson, N.; Le-Feuvre, R.; Iturra, C.; Elissetche, J.; Chapple, C.; Valenzuela, S. Rescue of Syringyl Lignin and Sinapate Ester Biosynthesis in Arabidopsis thaliana by a Coniferaldehyde 5-hydroxylase from Eucalyptus Globulus. Plant Cell Rep. 2014, 33, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Jones, L.; Ennos, A.R.; Turner, S.R. Cloning and Characterization of Irregular Xylem4 (irx4): A Severely Lignin-deficient Mutant of Arabidopsis. Plant J. 2001, 26, 205–216. [Google Scholar] [CrossRef]
- Wang, W.; Li, Y.; Cai, C.; Zhu, Q. Auxin Response Factors Fine-tune Lignin Biosynthesis in Response to Mechanical Bending in Bamboo. New Phytol. 2024, 241, 1161–1176. [Google Scholar] [CrossRef]
- Khadr, A.; Wang, G.-L.; Wang, Y.-H.; Zhang, R.-R.; Wang, X.-R.; Xu, Z.-S.; Tian, Y.-S.; Xiong, A.-S. Effects of Auxin (Indole-3-butyric Acid) on Growth Characteristics, Lignification, and Expression Profiles of Genes Involved in Lignin Biosynthesis in Carrot Taproot. Peer J. 2020, 8, e10492. [Google Scholar] [CrossRef]
- Qu, G.; Peng, D.; Yu, Z.; Chen, X.; Cheng, X.; Yang, Y.; Ye, T.; Lv, Q.; Ji, W.; Deng, X.; et al. Advances in the Role of Auxin for Transcriptional Regulation of Lignin Biosynthesis. Funct. Plant Biol. 2021, 48, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-C.; Chen, L.-M.; Liu, Z.-H. Rapid Effect of Copper on Lignin Biosynthesis in Soybean Roots. Plant Sci. 2005, 168, 855–861. [Google Scholar] [CrossRef]
- Ali, M.B.; Singh, N.; Shohael, A.M.; Hahn, E.J.; Paek, K.-Y. Phenolics Metabolism and Lignin Synthesis in Root Suspension Cultures of Panax Ginseng in Response to Copper Stress. Plant Sci. 2006, 171, 147–154. [Google Scholar] [CrossRef]
- Wang, H.; Jiang, C.; Wang, C.; Yang, Y.; Yang, L.; Gao, X.; Zhang, H. Antisense Expression of the Fasciclin-like Arabinogalactan Protein FLA6 gene in Populus Inhibits Expression of Its Homologous Genes and Alters Stem Biomechanics and Cell Wall Composition in Transgenic Trees. J. Exp. Bot. 2015, 66, 1291–1302. [Google Scholar] [CrossRef]
- Minjian, C.; Haiqiu, Y.; Hongkui, Y.; Chunji, J. Difference in Tolerance to Potassium Deficiency Between Two Maize Inbred Lines. Plant Prod. Sci. 2007, 10, 42–46. [Google Scholar] [CrossRef]
- Du, Q.; Zhao, X.-h.; Jiang, C.; Wang, X.; Han, Y.; Wang, J.; Yu, H. Effect of Potassium Deficiency on Root Growth and Nutrient Uptake in Maize (Zea mays L.). Agric. Sci. 2017, 08, 1263–1277. [Google Scholar] [CrossRef]
- Pedroza-Garcia, J.A.; Eekhout, T.; Achon, I.; Nisa, M.-U.; Coussens, G.; Vercauteren, I.; Van den Daele, H.; Pauwels, L.; Van Lijsebettens, M.; Raynaud, C.; et al. Maize ATR Safeguards Genome Stability During Kernel Development to Prevent Early Endosperm Endocycle Onset and Cell Death. Plant Cell 2021, 33, 2662–2684. [Google Scholar] [CrossRef] [PubMed]
- He, A.-L.; Niu, S.-Q.; Zhao, Q.; Li, Y.-S.; Gou, J.-Y.; Gao, H.-J.; Suo, S.-Z.; Zhang, J.-L. Induced Salt Tolerance of Perennial Ryegrass by a Novel Bacterium Strain from the Rhizosphere of a Desert Shrub Haloxylon ammodendron. Int. J. Mol. Sci. 2018, 19, 469. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Hameed, I.; Cao, D.D.; He, D.L.; Yang, P.F. Integrated Omics Analyses Identify Key Pathways Involved in Petiole Rigidity Formation in Sacred Lotus. Int. J. Mol. Sci. 2020, 21, 5087. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.H.; Guo, H.X.; Zhang, L.; Tang, Z.M.; Yu, X.M.; Wu, J.F.; Zeng, F.C. Metabolome and Transcriptome Association Analysis Reveals Dynamic Regulation of Purine Metabolism and Flavonoid Synthesis in Transdifferentiation during Somatic Embryogenesis in Cotton. Int. J. Mol. Sci. 2019, 20, 2070. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, D.; Zhang, Y.; Dong, Q.; Wang, K.; Zhang, H.; Du, Q.; Wang, J.; Wang, X.; Yu, H.; Zhao, X. Integrated Transcriptomic and Metabolomic Analysis of Exogenous NAA Effects on Maize Seedling Root Systems under Potassium Deficiency. Int. J. Mol. Sci. 2024, 25, 3366. https://doi.org/10.3390/ijms25063366
Zhou D, Zhang Y, Dong Q, Wang K, Zhang H, Du Q, Wang J, Wang X, Yu H, Zhao X. Integrated Transcriptomic and Metabolomic Analysis of Exogenous NAA Effects on Maize Seedling Root Systems under Potassium Deficiency. International Journal of Molecular Sciences. 2024; 25(6):3366. https://doi.org/10.3390/ijms25063366
Chicago/Turabian StyleZhou, Dongying, Yuanchun Zhang, Qiqi Dong, Kai Wang, He Zhang, Qi Du, Jing Wang, Xiaoguang Wang, Haiqiu Yu, and Xinhua Zhao. 2024. "Integrated Transcriptomic and Metabolomic Analysis of Exogenous NAA Effects on Maize Seedling Root Systems under Potassium Deficiency" International Journal of Molecular Sciences 25, no. 6: 3366. https://doi.org/10.3390/ijms25063366
APA StyleZhou, D., Zhang, Y., Dong, Q., Wang, K., Zhang, H., Du, Q., Wang, J., Wang, X., Yu, H., & Zhao, X. (2024). Integrated Transcriptomic and Metabolomic Analysis of Exogenous NAA Effects on Maize Seedling Root Systems under Potassium Deficiency. International Journal of Molecular Sciences, 25(6), 3366. https://doi.org/10.3390/ijms25063366