

T Cell Responses in Pregnant Women Who Received mRNA-Based Vaccination to Prevent COVID-19 Revealed Unknown Exposure to the Natural Infection and Numerous SARS-CoV-2-Specific CD4- CD8- Double Negative T Cells and Regulatory T Cells

 ,
,  and
and
Abstract
1. Introduction
2. Results
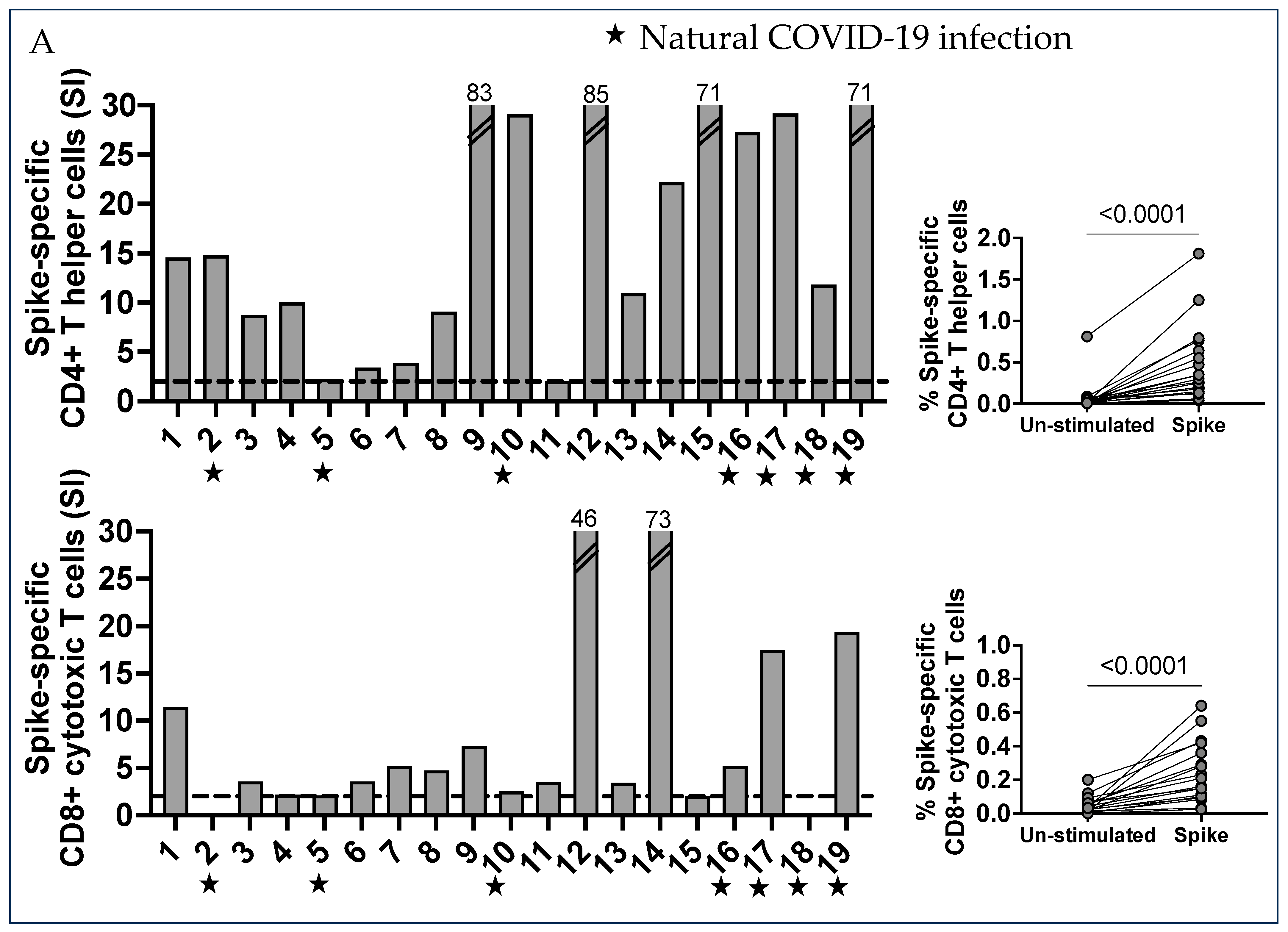
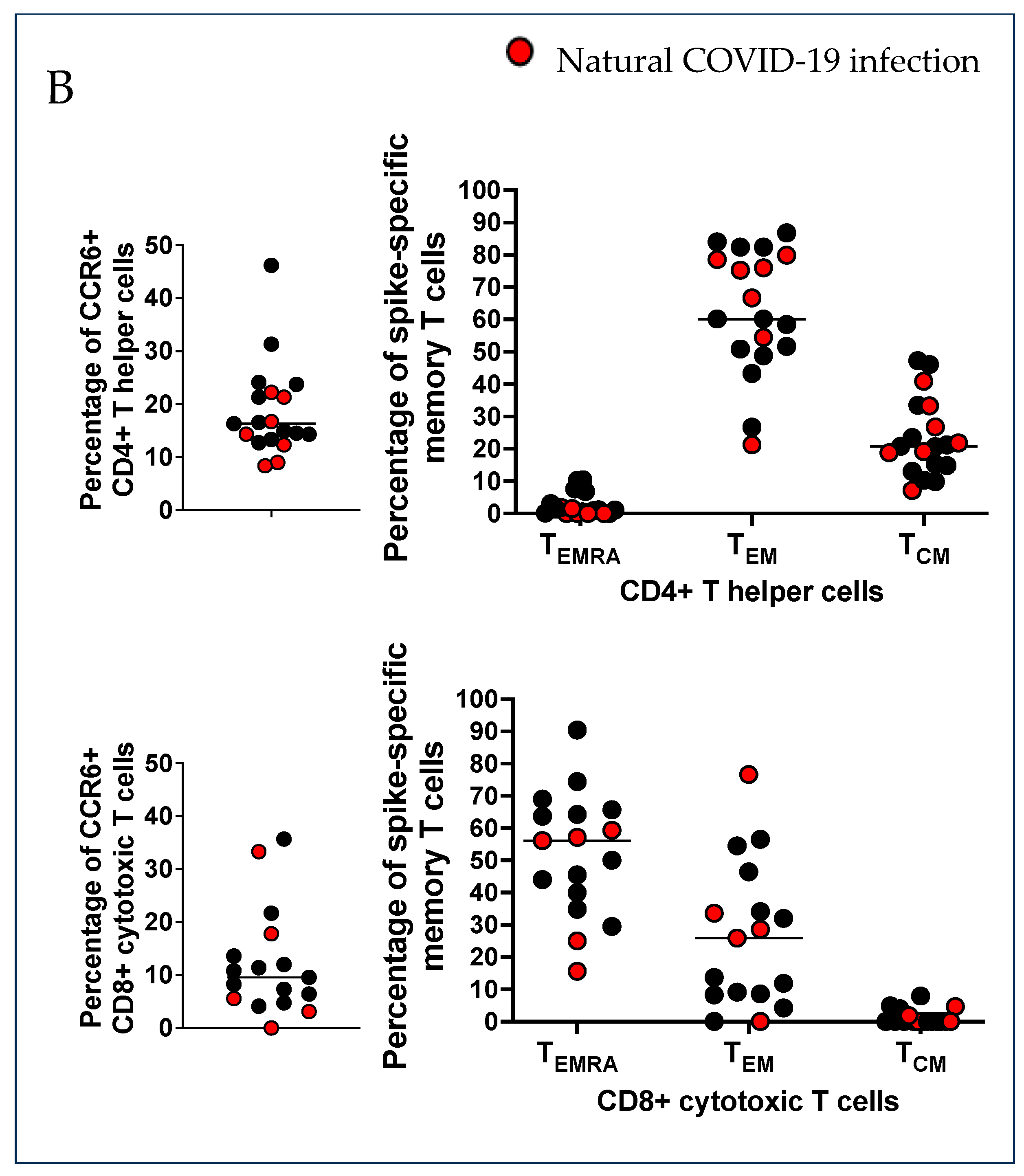
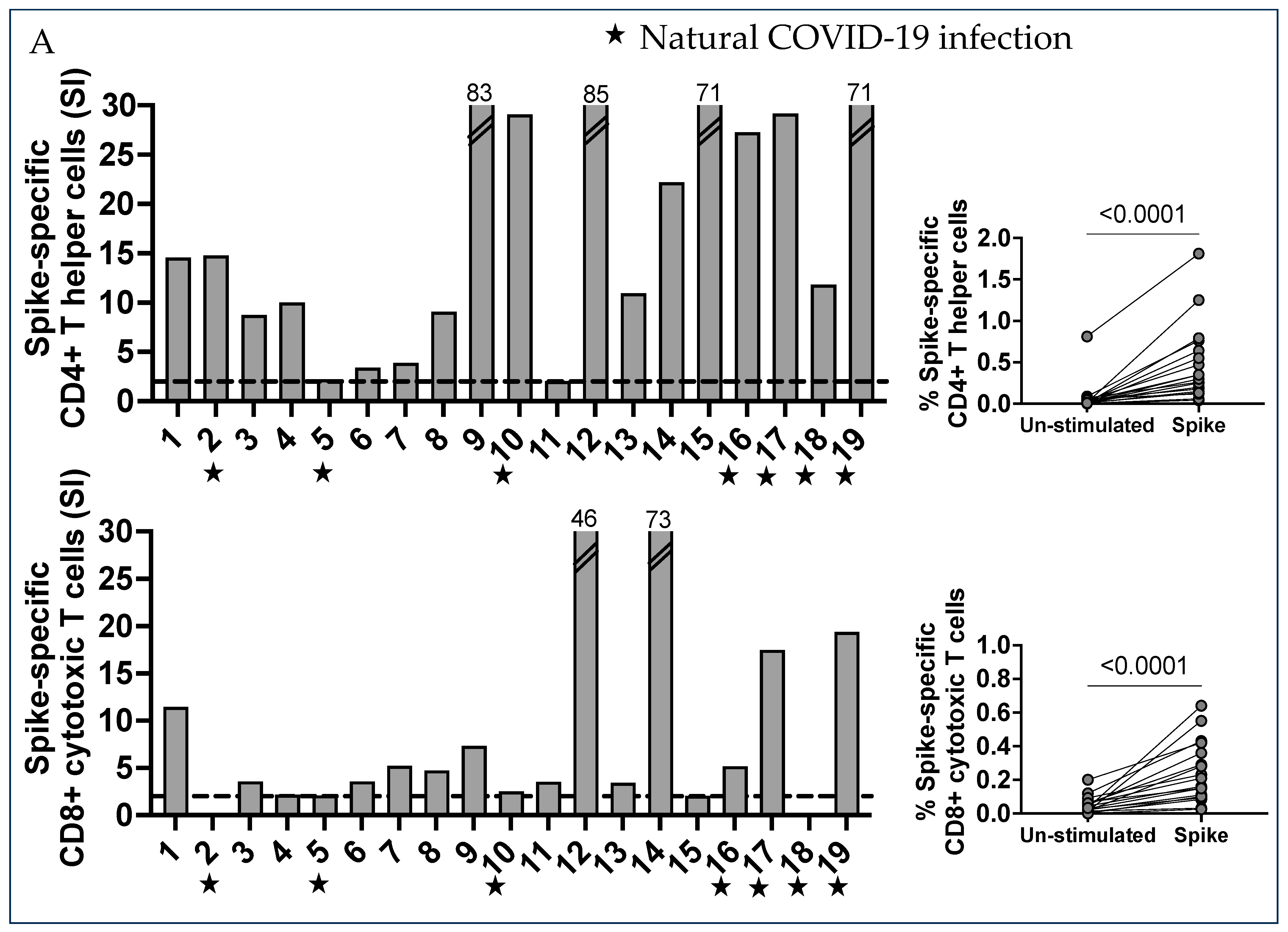
2.1. Characterization of SARS-CoV-2 Spike-Specific CD4+ and CD8+ T Cells and Their Memory Phenotype in Vaccinated Pregnant Women
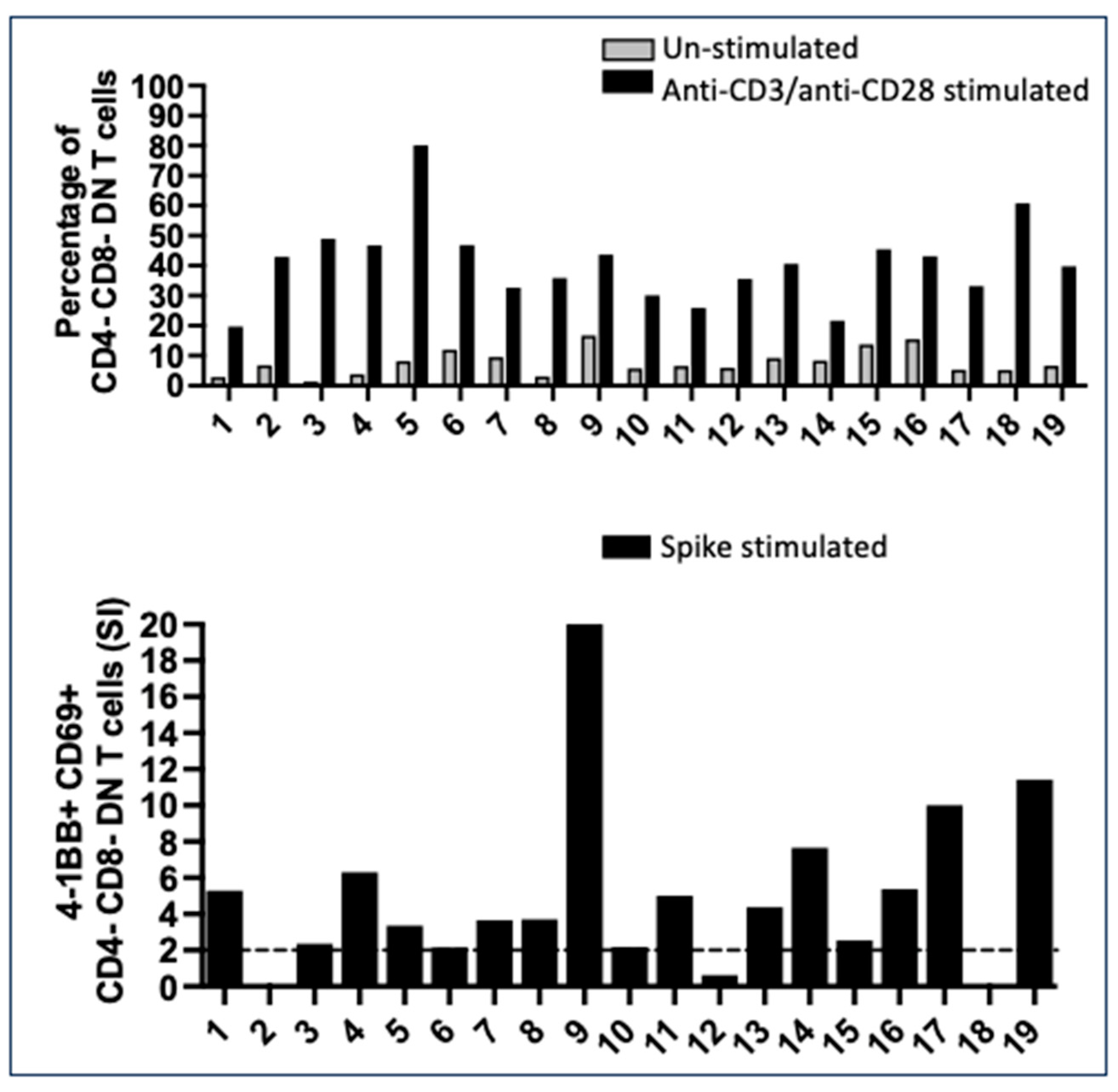
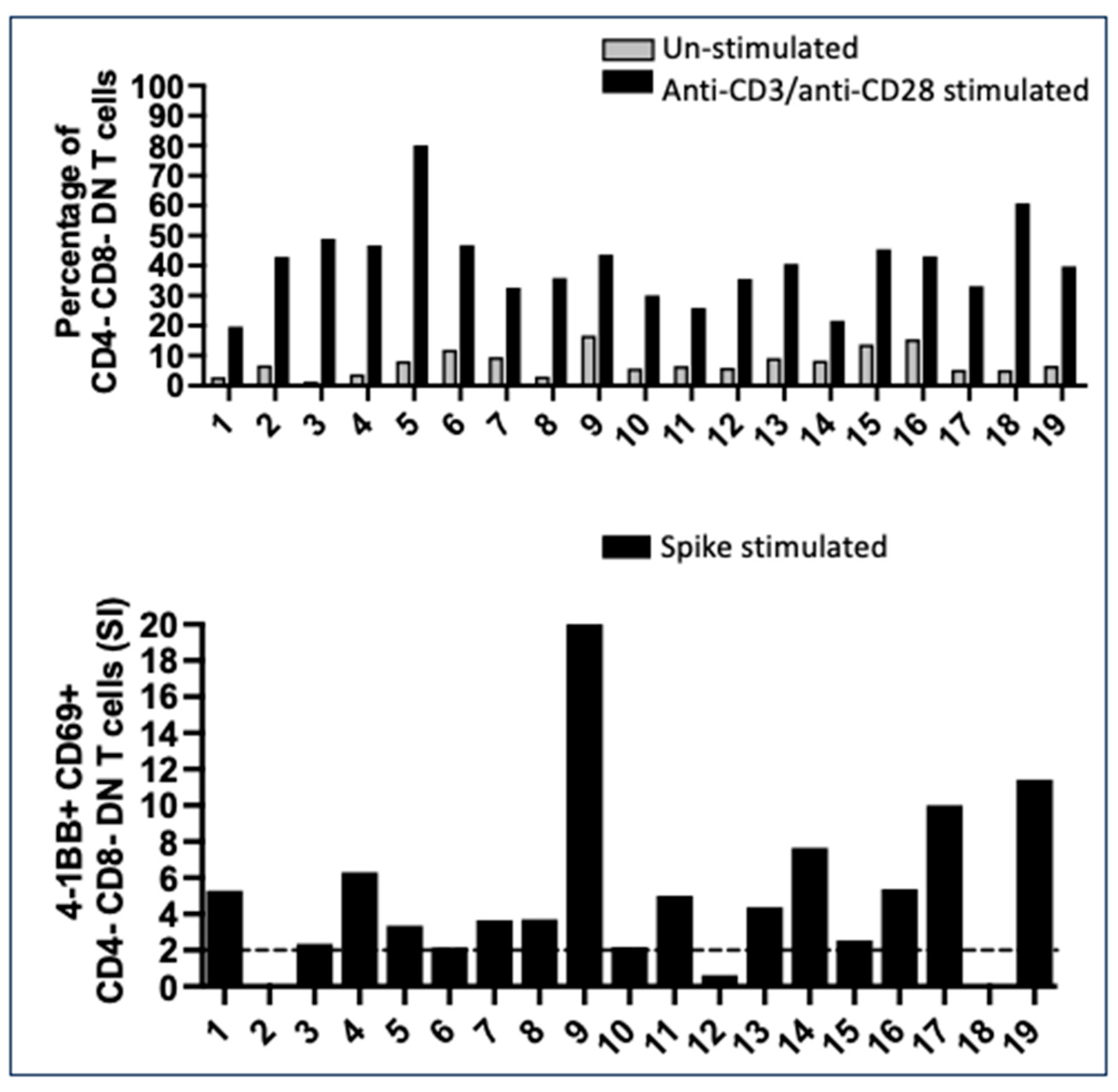
2.2. Numerous CD4- CD8- Double Negative T Cells with a Mature T Cell Receptor (TcR) Recognized Spike Peptides
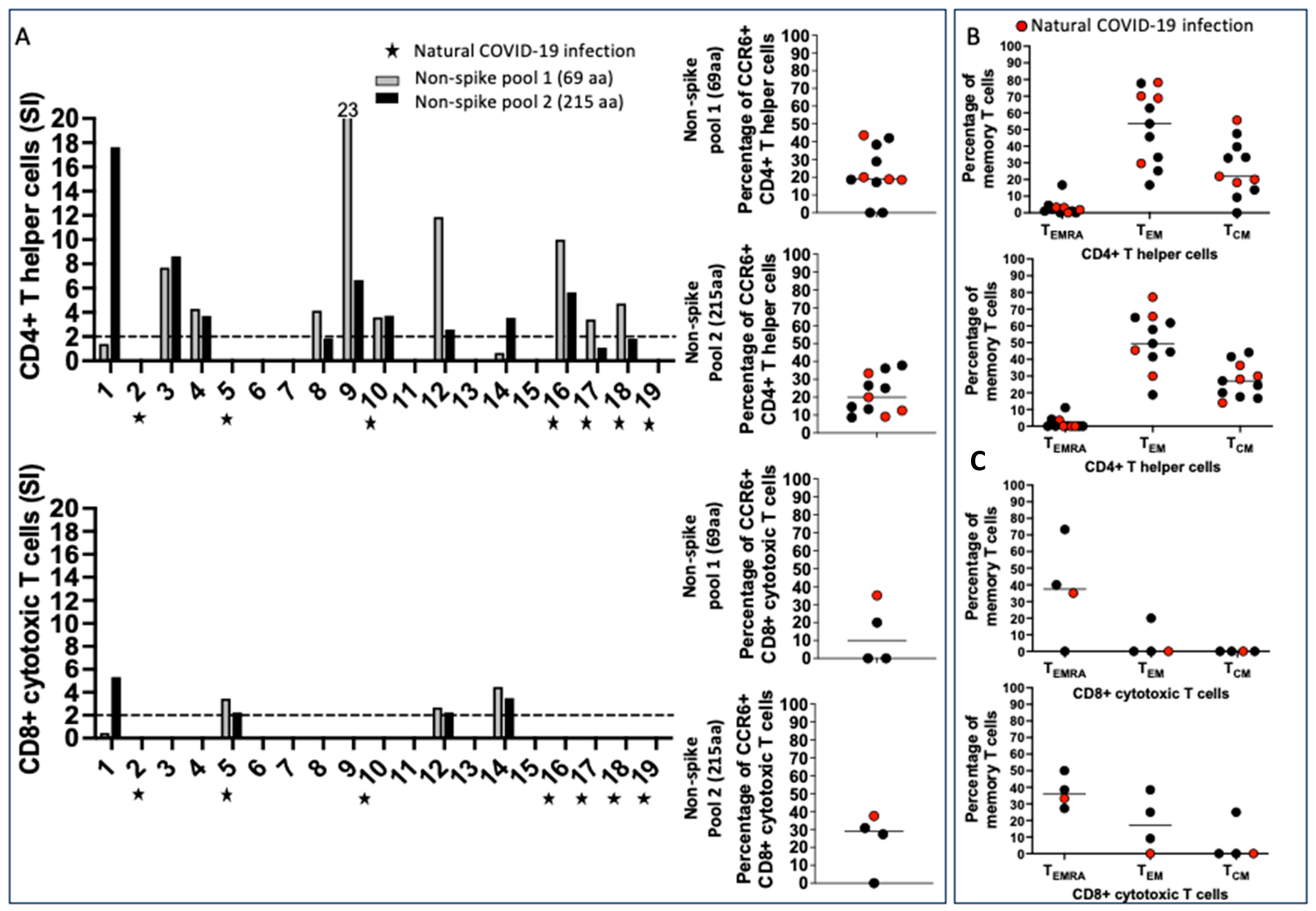
2.3. SARS-CoV-2 Non-Spike-Specific T Cells Were Found in Some Vaccinated Pregnant Subjects Who Were Not Aware of Previous COVID-19 Infection
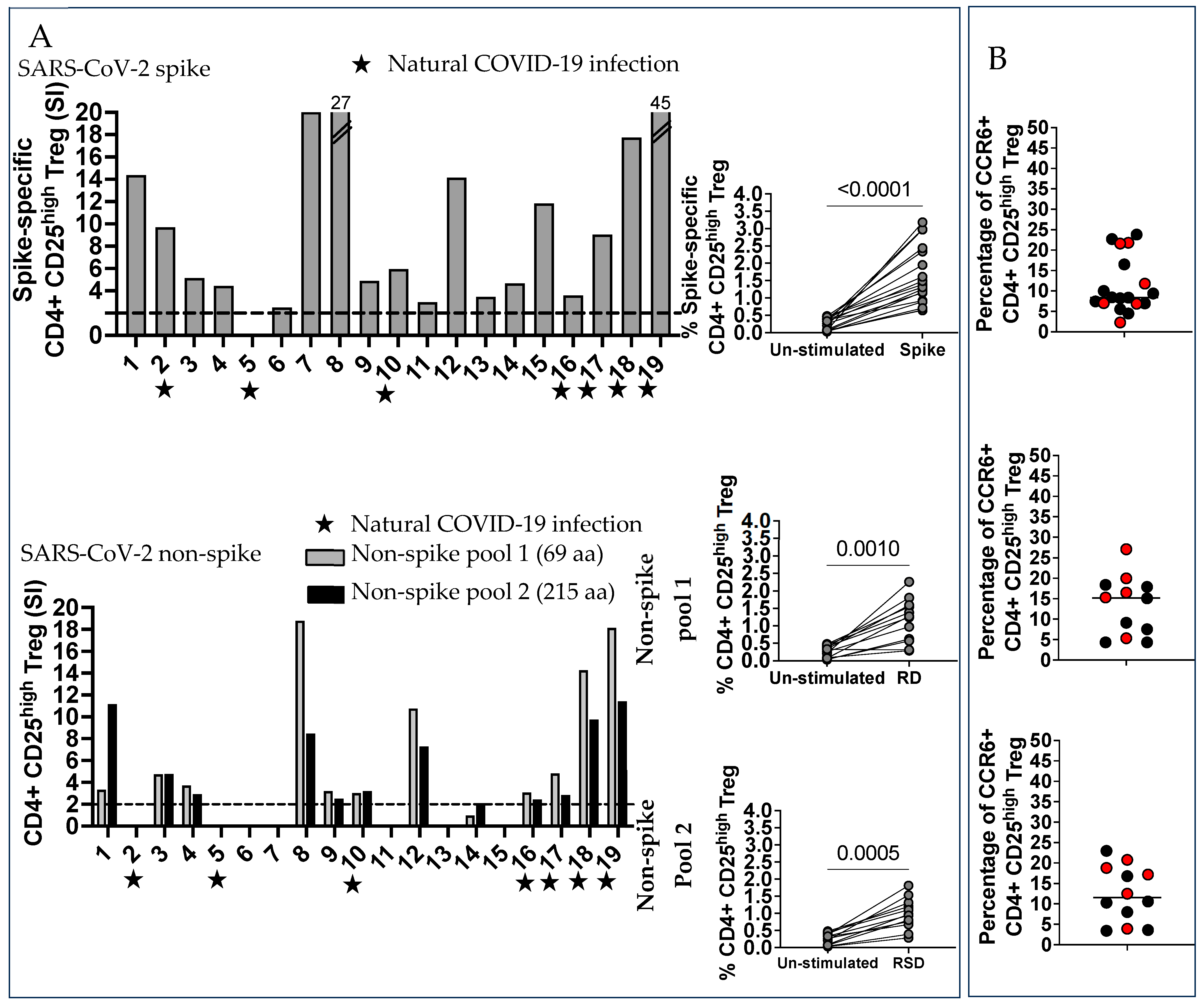
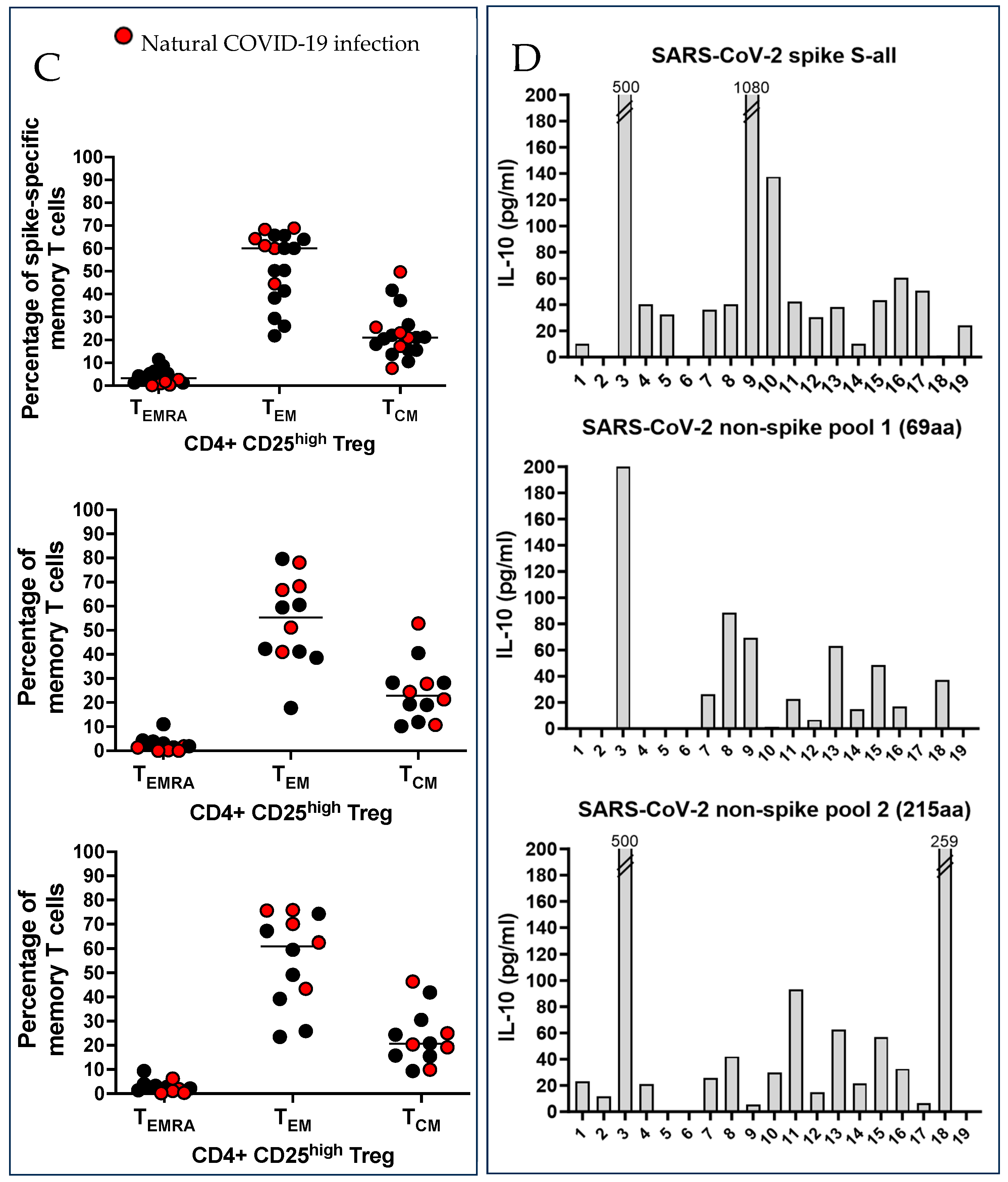
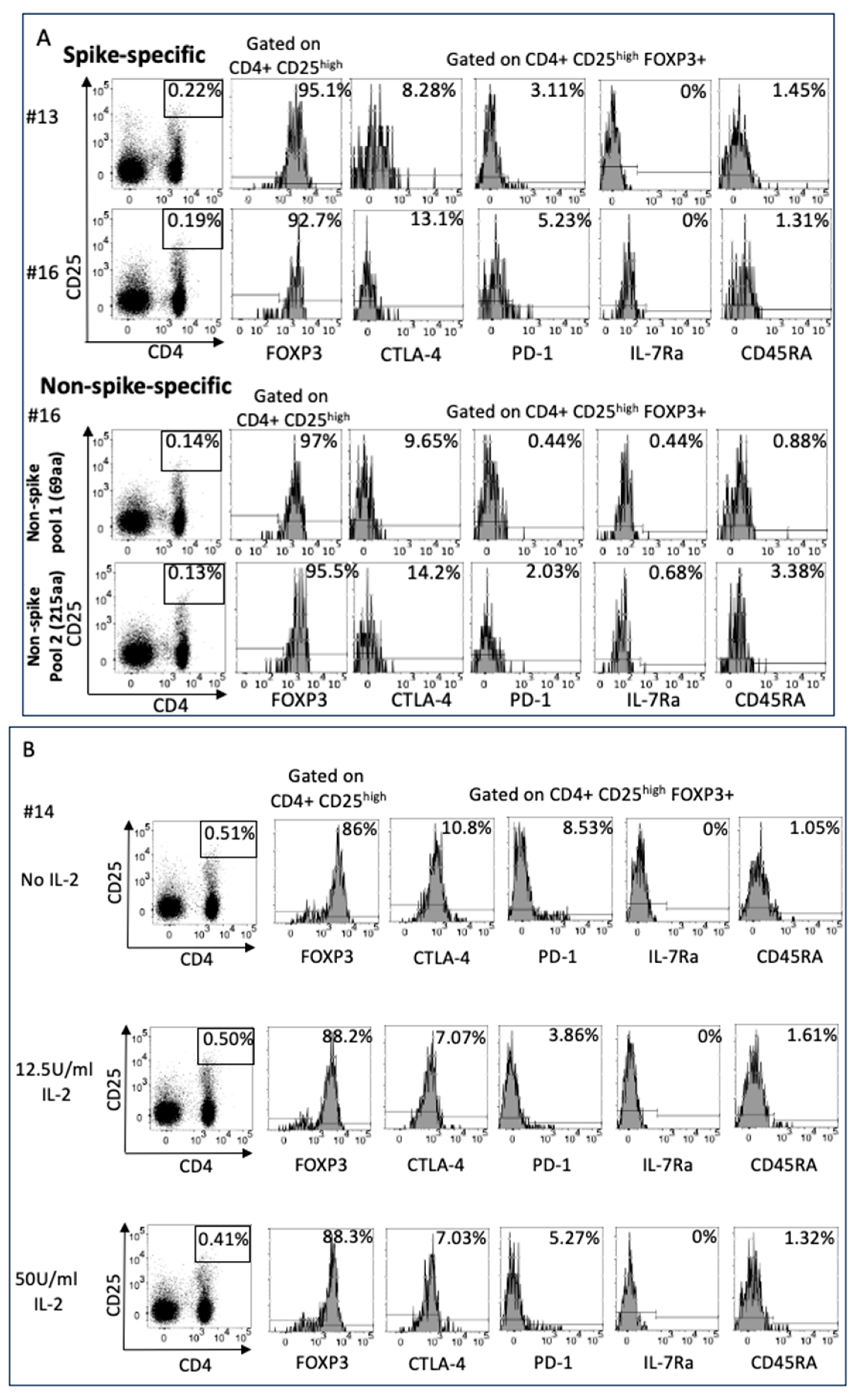
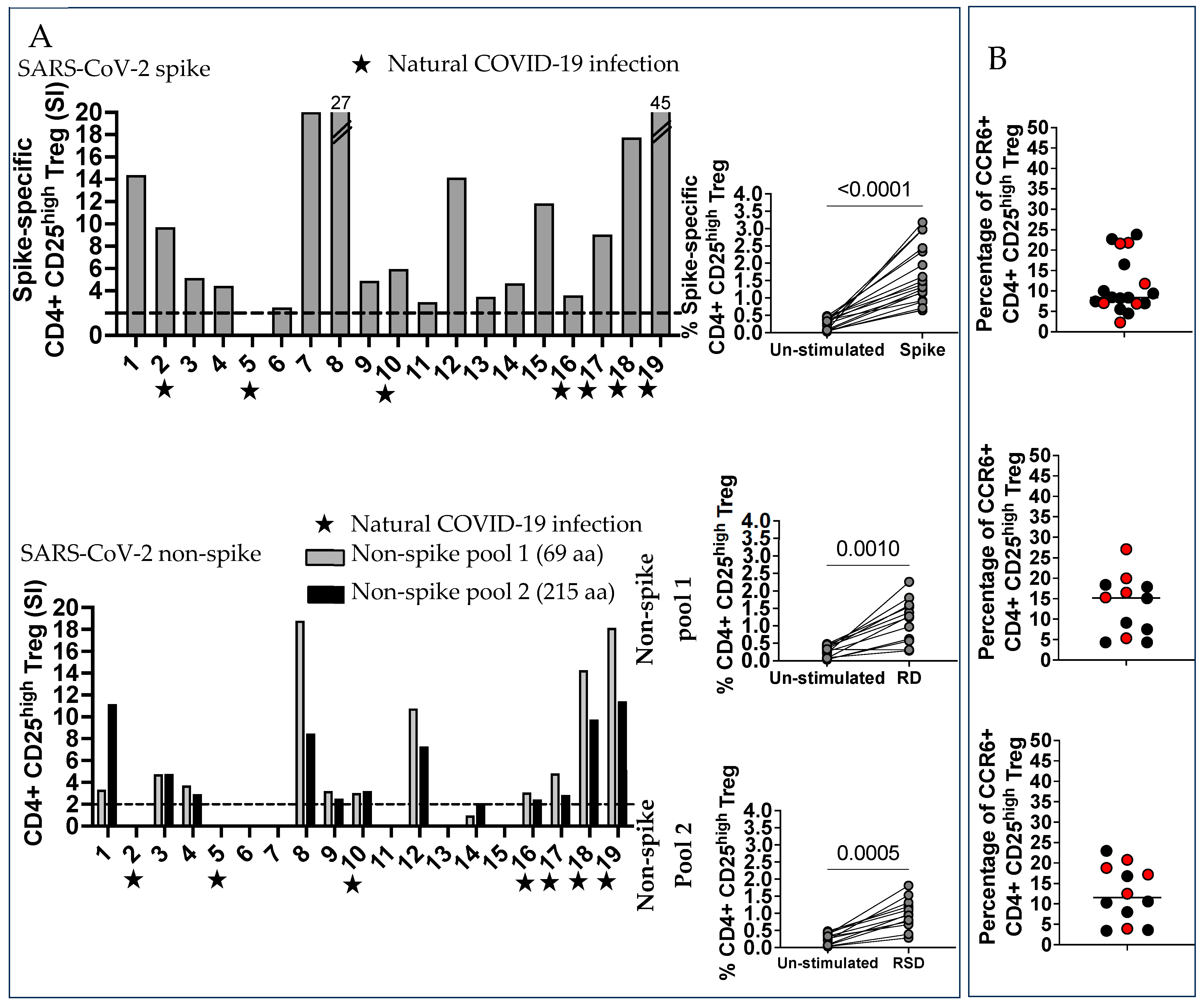
2.4. SARS-CoV-2 Spike and Non-Spike-Specific Treg Were Numerous in Vaccinated Pregnant Women
3. Discussion
4. Materials and Methods
4.1. Study Population
4.2. SARS-CoV-2 Peptides Mega Pool
4.3. AIM Assay
4.4. Phenotype of SARS-CoV-2 Spike and Non-Spike-Specific Treg
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ellington, S.; Strid, P.; Tong, V.T.; Woodworth, K.; Galang, R.R.; Zambrano, L.D.; Nahabedian, J.; Anderson, K.; Gilboa, S.M. Characteristics of Women of Reproductive Age with Laboratory-Confirmed SARS-CoV-2 Infection by Pregnancy Status—United States, January 22–June 7, 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 769–775. [Google Scholar] [CrossRef]
- Delahoy, M.J.; Whitaker, M.; O’Halloran, A.; Chai, S.J.; Kirley, P.D.; Alden, N.; Kawasaki, B.; Meek, J.; Yousey-Hindes, K.; Anderson, E.J.; et al. Characteristics and Maternal and Birth Outcomes of Hospitalized Pregnant Women with Laboratory-Confirmed COVID-19—COVID-NET, 13 States, March 1–August 22, 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 1347–1354. [Google Scholar] [CrossRef] [PubMed]
- Red-Horse, K.; Zhou, Y.; Genbacev, O.; Prakobphol, A.; Foulk, R.; McMaster, M.; Fisher, S.J. Trophoblast differentiation during embryo implantation and formation of the maternal fetal interface. J. Clin. Investig. 2004, 114, 744–754. [Google Scholar] [CrossRef] [PubMed]
- Moffett, A.; Loke, C. Immunology of placentation in ehtherian mammals. Nat. Rev. Immunol. 2006, 6, 584–594. [Google Scholar] [CrossRef]
- Mold, J.E.; McCune, G.M. Immunological tolerance during fetal development: From mouse to man. Adv. Immunol. 2012, 115, 73–111. [Google Scholar]
- Erlebacher, A. Immunology of the maternal-fetal interface. Annu. Rev. Immunol. 2013, 31, 387–411. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.; Kumar, J.; Lin, G.; Behnamfar, N.; Hisieh, L.; Shimizu, C.; Tremoulet, A.H.; Burns, J.C.; Linden, J. Pediatric tolerogenic DCs expressing CD4 and immunoglobulin-like transcript receptor (ILT)-4 secrete IL-10 in response to Fc and adenosine. Eur. J. Immunol. 2018, 48, 482–491. [Google Scholar] [CrossRef]
- Hsieh, L.E.; Grifoni, A.; Dave, H.; Wang, J.; Johnson, D.; Zellner, J.; Sidney, J.; Chambers, C.; Franco, A. SARS-CoV-2-specific T cell responses and immune regulation in infected pregnant women. J. Reprod. Immunol. 2022, 149, 103464–103471. [Google Scholar] [CrossRef]
- Amodio, G.; Mugione, A.; Sanchez, A.M.; Vigano’, P.; Candiani, M.; Somigliana, E.; Roncarolo, M.G.; Panina-Bordignon, P.; Gregori, S. HLA-G expressing DC-10 and CD4+ T cells accumulate in human decidua during pregnancy. Hum. Immunol. 2013, 74, 406–411. [Google Scholar] [CrossRef]
- Amodio, G.; Comi, M.; Tomasoni, D.; Gianolini, M.E.; Rizzo, R.; LeMaoult, J.L.; Roncarolo, M.G.; Gregori, S. HLA-G expression levels influence the tolerogenic activity of human DC-10. Cell Ther. Immunother. 2015, 100, 548. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Vignali, D.A.; Rudensky, A.Y.; Niec, R.E.; Waldmann, H. The plasticity and stability of regulatory T cells. Nat. Rev. Immunol. 2013, 13, 461–467. [Google Scholar] [CrossRef]
- Harb, H.; Benamar, M.; Lai, P.S.; Contini, P.; Griffith, J.W.; Crestani, E.; Schmitz-Abe, K.; Chen, Q.; Fong, J.; Marri, L.; et al. Notch4 signaling limits regulatory T-cell-mediated tissue repair and promotes severe lung inflammtion in viral infections. Immunity 2021, 54, 1186–1199. [Google Scholar] [CrossRef] [PubMed]
- Franco, A.; Song, J.; Chambers, C.; Sette, A.; Grifoni, A. SARS-CoV-2 spike-specific regulatory T cells (Treg) expand and develop memory in vaccine recipients suggesting a role for immune regulation in preventing severe symptoms in COVID-19. Autoimmunity 2023, 56, 2259133. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Jiang, X.; Day, X.; Li, B. The dymamic role of FOXP3+ Tregs and their potential therapeutic applications during SARS-CoV-2 infection. Front. Immunol. 2022, 13, 916411–916433. [Google Scholar] [CrossRef] [PubMed]
- Muyayalo, K.; Huang, D.-H.; Zhao, S.-J.; Xie, T.; Mor, G.; Liao, A.-H. COVID-19 and Treg/Th17 imbalance: Potential relationship to pregnancy outcomes. Am. J. Reprod. Immunol. 2020, 85, e13304. [Google Scholar] [CrossRef] [PubMed]
- Jarjour, N.N.; Masopust, D.; Jameson, S.C. T cell memory: Understanding COVID-19. Immunity 2021, 54, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Mateus, J.; Coelho, C.H.; Dan, J.M.; Moderbacher, C.R.; Gálvez, R.I.; Cortes, F.H.; Grifoni, A.; Tarke, A.; Chang, J.; et al. Humoral and cellular immune memory to four COVID-19 vaccines. Cell 2022, 185, 2434–2451. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Kalafat, E.; Blakeway, H.; Townsend, R.; O’brien, P.; Morris, E.; Draycott, T.; Thangaratinam, S.; Le Doare, K.; Ladhani, S.; et al. Systematic review and meta-analysis of the effectiveness and perinatal outcomes of COVID-19 vaccination in pregnancy. Nat. Commun. 2022, 13, 2414. [Google Scholar] [CrossRef]
- Grifoni, A.; Sidney, J.; Zhang, Y.; Scheuermann, R.H.; Peters, B.; Sette, A. A Sequence Homology and Bioinformatic Approach Can Predict Candidate Targets for Immune Responses to SARS-CoV-2. Cell Host Microbe 2020, 27, 671–680.e2. [Google Scholar] [CrossRef]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef]
- Mateus, J.; Grifoni, A.; Tarke, A.; Sidney, J.; Ramirez, S.I.; Dan, J.M.; Burger, Z.C.; Rawlings, S.A.; Smith, D.M.; Phillips, E.; et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science 2020, 370, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Weiskopf, D.; Schmitz, K.S.; Raadsen, M.P.; Grifoni, A.; Okba, N.M.; Endeman, H.; Van Den Akker, J.P.C.; Molenkamp, R.; Koopmans, M.P.G.; Van Gorp, E.C.M.; et al. Phenotype and kinetics of SARS-CoV-2-specific T cells in COVID-19 patients with acute respiratory distress syndrome. Sci. Immunol. 2020, 5, eabd2071. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.D.; Wang, E.; Garrigan, E.; Goodwin, B.; Sutherland, A.; Tarke, A.; Chang, J.; Galvez, R.I.; Mateus, J.; Ramirez, S.I.; et al. Development of a T cell-based immunodiagnostic system to effectively distinguish SARS-CoV-2 infection and COVID-19 vaccination status. Cell Host Microbe 2022, 30, 388–399. [Google Scholar] [CrossRef]
- Zielinski, C.; Mele, F.; Aschenner, D.; Jarrossay, D.; Ronchi, F.; Gattorno, M.; Monticelli, S.; Lanzavecchia, A.; Sallusto, F. Pathogen-induced human Th17 cells produce IFN-gamma or IL-10 and are regulated by IL-1beta. Nature 2012, 484, 514–518. [Google Scholar] [CrossRef]
- Duhen, T.; Duhen, R.; Lanzavecchia, A.; Sallusto, F.; Campbell, D.J. Functionally distinct subsets of human FOXP3+ Treg cells that phenotypically mirror effector Th cells. Blood 2012, 119, 4430–4440. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S.; Mikami, N.; Wing, J.B.; Tanaka, A.; Ichiyama, K.; Ohkura, N. Regulatory T cells and human diseases. Annu. Rev. Immunol. 2020, 38, 541–566. [Google Scholar] [CrossRef]
- Perry, J.A.; Shallberg, L.; Clark, J.T.; Gullicksrud, J.A.; DeLong, J.H.; Douglas, B.B.; Hart, A.P.; Lanzar, Z.; O’dea, K.; Konradt, C.; et al. PD-L1-PD-1 interactions limit effector regulatory T cell populations at homeostasis and during infection. Nat. Immunol. 2022, 23, 743–756. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Robey, E.A.; Fowlkes, B.J.; Gordon, J.W.; Kioussis, D.; von Boemer, H.; Ramsdell, F.; Axel, R. Thymic selection in CD8 transgenic mice supports an instructive model for commitment to a CD4 or CD8 lineage. Cell 1991, 64, 99–107. [Google Scholar] [CrossRef]
- Hsieh, L.E.; Grifoni, A.; Shimizu, C.; Tremoulet, A.H.; Dummer, K.B.; Burns, J.C.; Sette, A.; Franco, A. T cells in Multisystem Inflammatory Syndrome in children have a predominant CD4+ T helper response to SARS-CoV-2 and numerous virus-specific CD4- CD8- double negative T cells. Int. J. Mol. Sci. 2022, 23, 7219–7233. [Google Scholar] [CrossRef]
- Moro, A.; Gao, Z.; Wang, L.; Yu, A.; Hsiung, S.; Ban, Y.; Yan, A.; Sologon, C.M.; Chen, X.S.; Malek, T.R. Dynamic transcriptional activity and chromatin remodeling of regulatory T cells after varied duration of interleukin-2 receptor signaling. Nat. Immunol. 2022, 23, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Loretelli, C.; Abdelsalam, A.; D’Addio, F.; Nasr, M.B.; Assi, E.; Usuelli, V.; Maestroni, A.; Seelam, A.J.; Ippolito, E.; Maggio, S.D.; et al. PD-1 blockade counteracts post-COVID-19 immune abnormalities and stimulates the anti-SARS-CoV-2 immune response. JCI Insight 2021, 6, i146701. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A.; Sidney, J.; Vita, R.; Peters, B.; Crotty, S.; Weiskopf, D.; Sette, A. SARS-CoV-2 human T cell epitopes: Adaptive immune response against COVID-19. Cell Host Microbe 2021, 29, 1076–1092. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subject Number | Weeds Gestation at Sample Collection | Age | Ethnicity | Natural Infection | Latest COVID-I9 Vaccine |
|---|---|---|---|---|---|
| 1 | 10 | 35 | Hispanic/White | N | 12/2022, Moderna Bivalent |
| 2 | 24 | 34 | Non-hispanic/White | Y | 1/2023, Moderna Bivalent |
| 3 | 25 | 35 | Non-hispanic/White | N | 12/2022, Pfizer Bivalent |
| 4 | 27 | 28 | Non-hispanic/White | N | 12/2022, Moderna Bivalent |
| 5 | 19 | 39 | Non-hispanic/White | Y | 1/2023, Pfizer Bivalent |
| 6 | 17 | 40 | Non-hispanic/White | N | 12/2022, Moderna Bivalent |
| 7 | 26 | 36 | Non-hispanic/White | N | 12/2022, Pfizer Bivalent |
| 8 | 17 | 31 | Non-hispanic/White | N | 1/2023, Pfizer Bivalent |
| 9 | 20 | 31 | Non-hispanic/White | N | 2/2023, Pfizer Bivalent |
| 10 | 23 | 28 | Non-hispanic/White | Y | 2/2023, Pfizer Bivalent |
| 11 | 16 | 28 | Non-hispanic/Cancasian and Asian | N | 1/2023, Moderna Bivalent |
| 12 | 26 | 34 | Non-hispanic/White | N | 2/2023, Moderna Bivalent |
| 13 | 26 | 42 | Non-hispanic/White | N | 4/2023, Pfizer Bivalent |
| 14 | 23 | 30 | Non-hispanicWhite | N | 5/2023, Moderna Bivalent |
| 15 | 34 | 32 | Non-hispanic/White | N | 8/2023, Pfizer Bivalent |
| 16 | 24 | 32 | Non-Hispanic/Asian | Y | 9/2023, Pfizer Bivalent |
| 17 | 23 | 37 | Non-hispanic/White | Y | 9/2023, Pfizer Bivalent |
| 18 | 21 | 34 | Non-hispanic/White | Y | 10/2023, Moderna |
| 19 | 8 | 40 | Non-hispanic/White | Y | 9/2023, Moderna |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chambers, C.D.; Song, J.; da Silva Antunes, R.; Sette, A.; Franco, A. T Cell Responses in Pregnant Women Who Received mRNA-Based Vaccination to Prevent COVID-19 Revealed Unknown Exposure to the Natural Infection and Numerous SARS-CoV-2-Specific CD4- CD8- Double Negative T Cells and Regulatory T Cells. Int. J. Mol. Sci. 2024, 25, 2031. https://doi.org/10.3390/ijms25042031
Chambers CD, Song J, da Silva Antunes R, Sette A, Franco A. T Cell Responses in Pregnant Women Who Received mRNA-Based Vaccination to Prevent COVID-19 Revealed Unknown Exposure to the Natural Infection and Numerous SARS-CoV-2-Specific CD4- CD8- Double Negative T Cells and Regulatory T Cells. International Journal of Molecular Sciences. 2024; 25(4):2031. https://doi.org/10.3390/ijms25042031
Chicago/Turabian StyleChambers, Christina D., Jaeyoon Song, Ricardo da Silva Antunes, Alessandro Sette, and Alessandra Franco. 2024. "T Cell Responses in Pregnant Women Who Received mRNA-Based Vaccination to Prevent COVID-19 Revealed Unknown Exposure to the Natural Infection and Numerous SARS-CoV-2-Specific CD4- CD8- Double Negative T Cells and Regulatory T Cells" International Journal of Molecular Sciences 25, no. 4: 2031. https://doi.org/10.3390/ijms25042031
APA StyleChambers, C. D., Song, J., da Silva Antunes, R., Sette, A., & Franco, A. (2024). T Cell Responses in Pregnant Women Who Received mRNA-Based Vaccination to Prevent COVID-19 Revealed Unknown Exposure to the Natural Infection and Numerous SARS-CoV-2-Specific CD4- CD8- Double Negative T Cells and Regulatory T Cells. International Journal of Molecular Sciences, 25(4), 2031. https://doi.org/10.3390/ijms25042031