
Structural Analysis of Breast-Milk αS1-Casein: An α-Helical Conformation Is Required for TLR4-Stimulation



Abstract

1. Introduction
2. Results and Discussion
2.1. In Silico and Experimental Structure Analysis of Human αS1-Casein
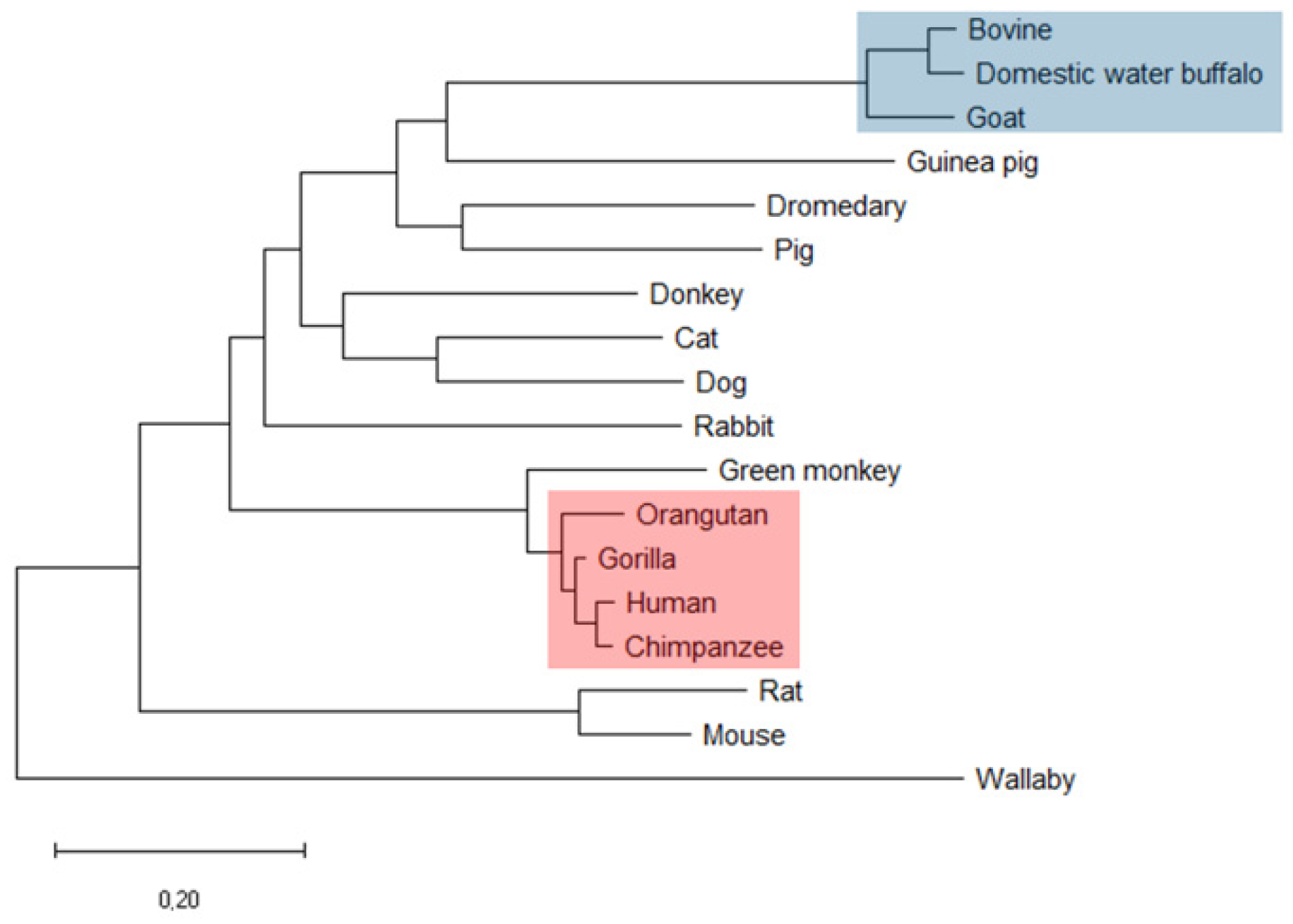
2.1.1. Amino Acid Sequence Analysis and Phylogenetic Relationships
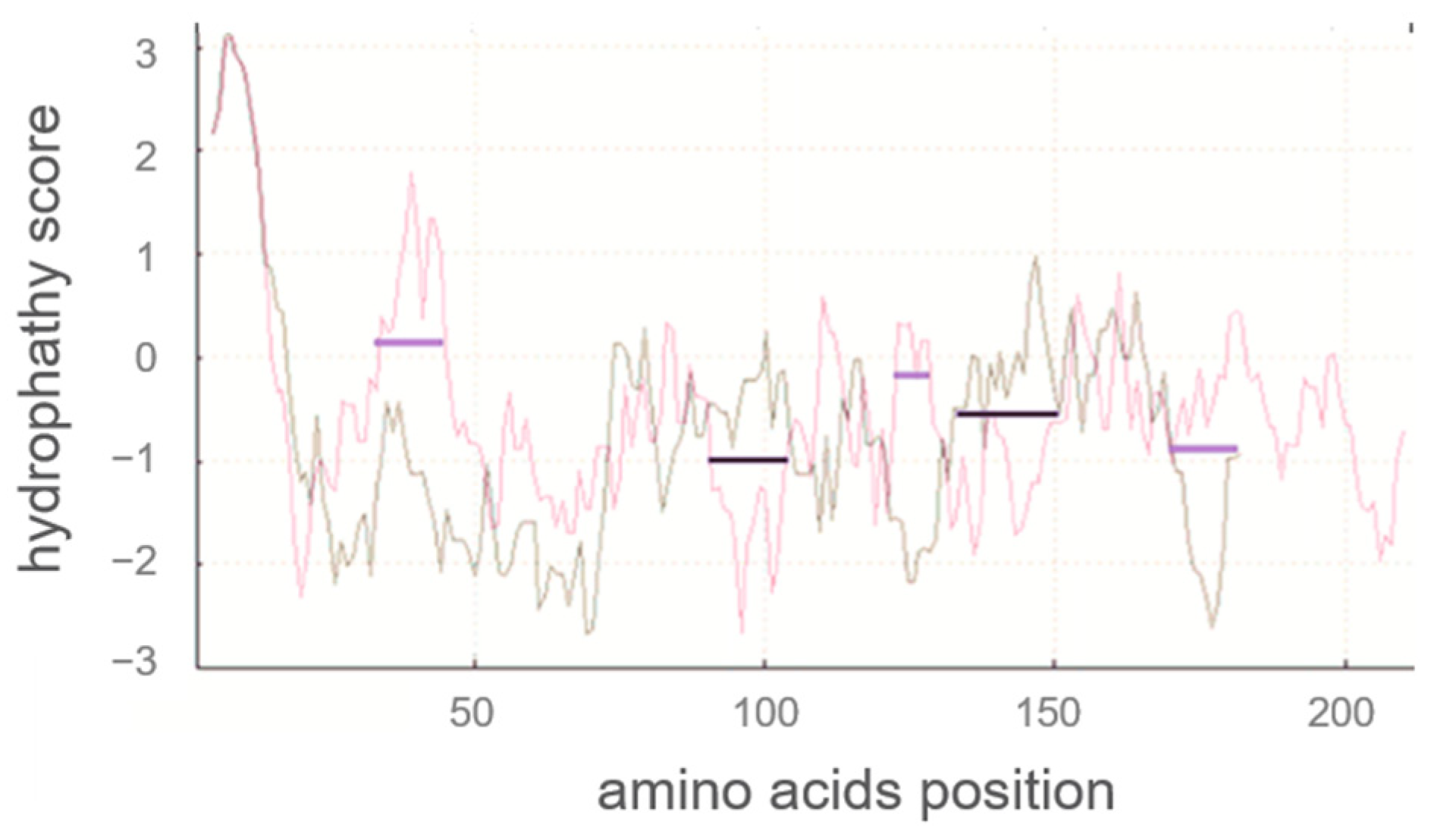
2.1.2. Hydrophobicity Analysis of Human αS1-Casein and Comparison with Bovine αS1-Casein
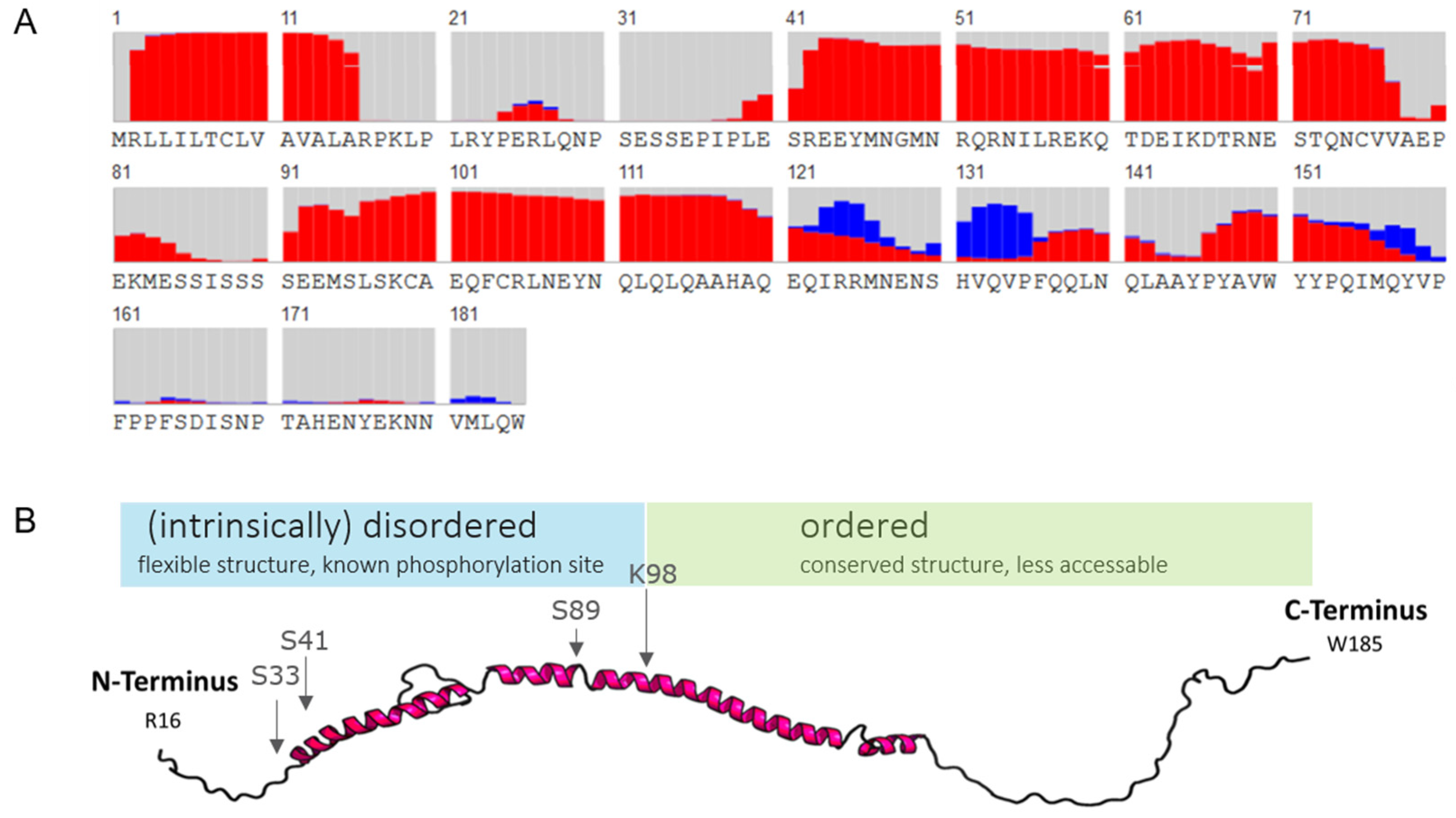
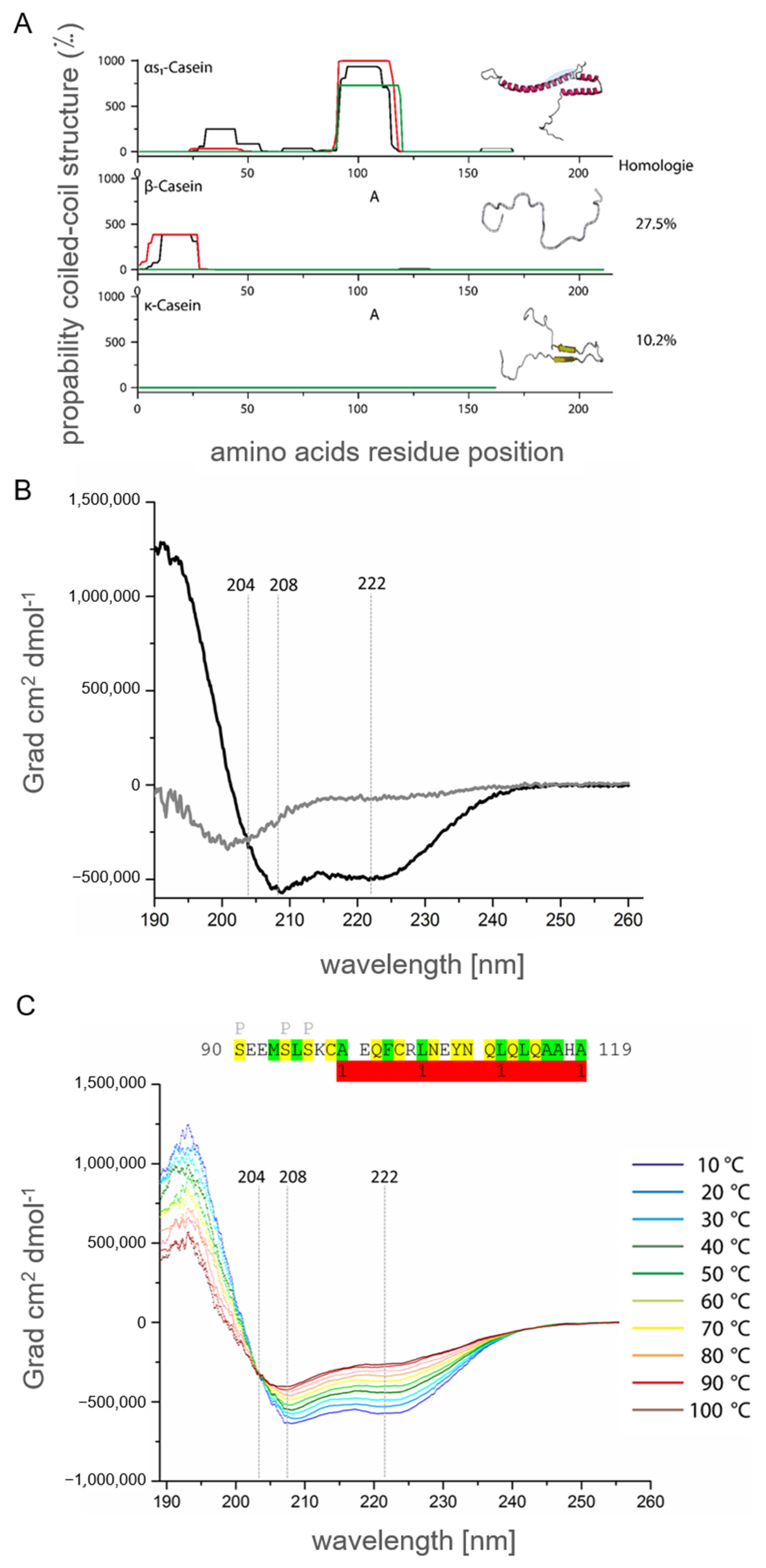
2.1.3. In Silico Secondary Structure Analysis of Human αS1-Casein
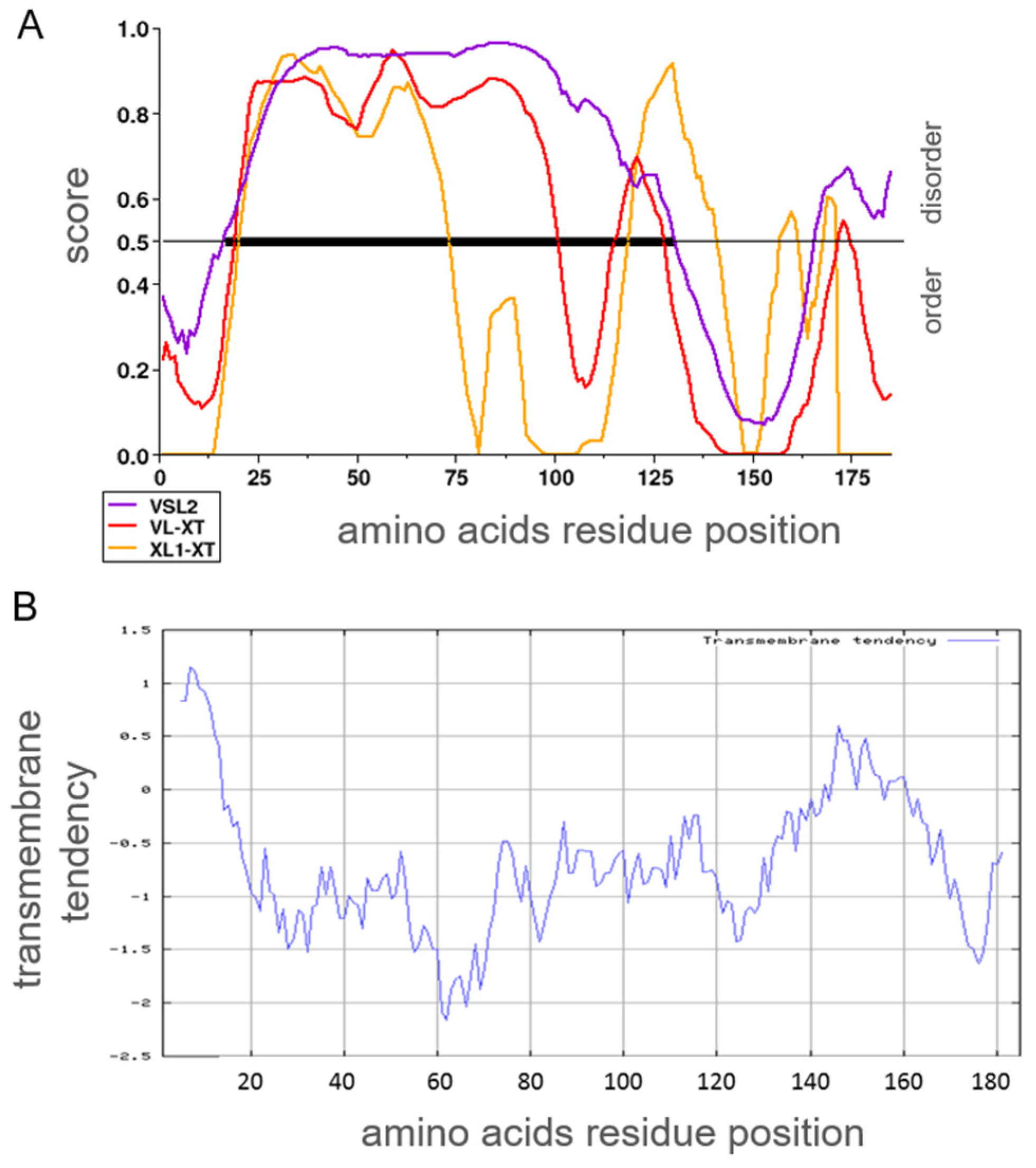
2.1.4. Prediction and Probability for Intrinsically Disordered Regions or Transmembrane Domains
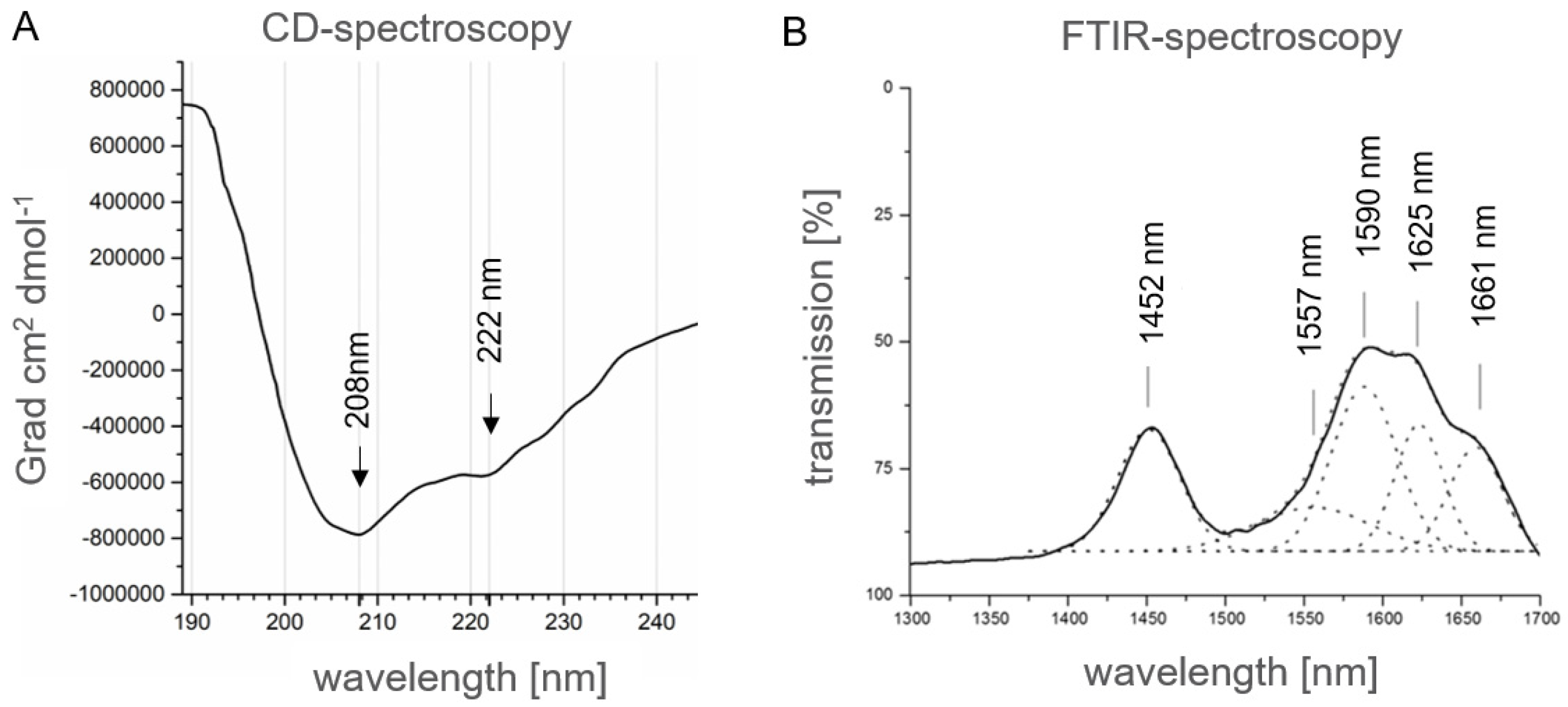
2.1.5. Secondary Structure Analysis of Human αS1-Casein by Spectroscopic Methods
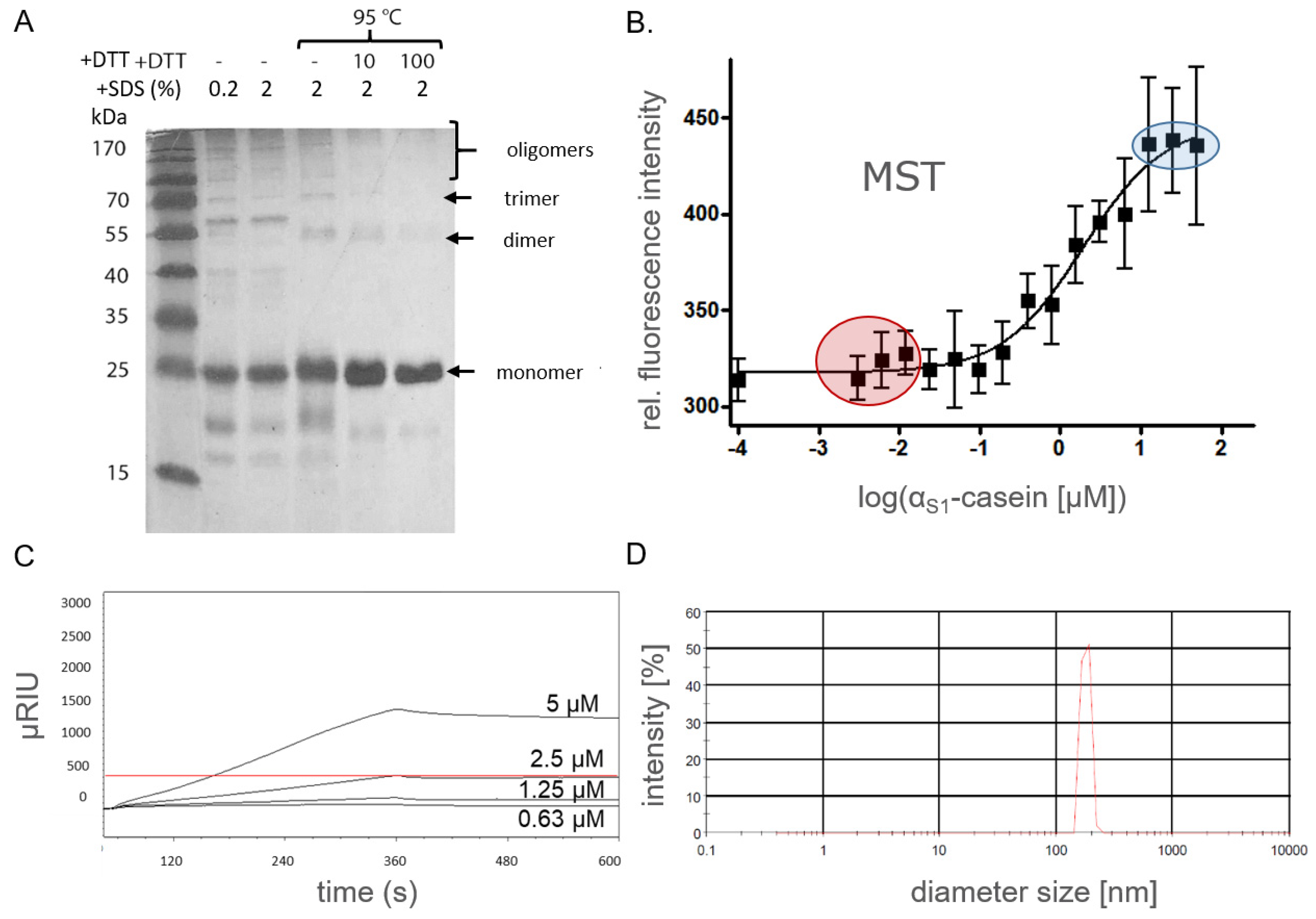
2.2. Oligomerization of Human αS1-Casein
2.3. The Influence of Temperature, pH and Phosphorylation on αS1-Casein Structure
2.4. αS1-Casein Contains a Coiled-Coil Domain
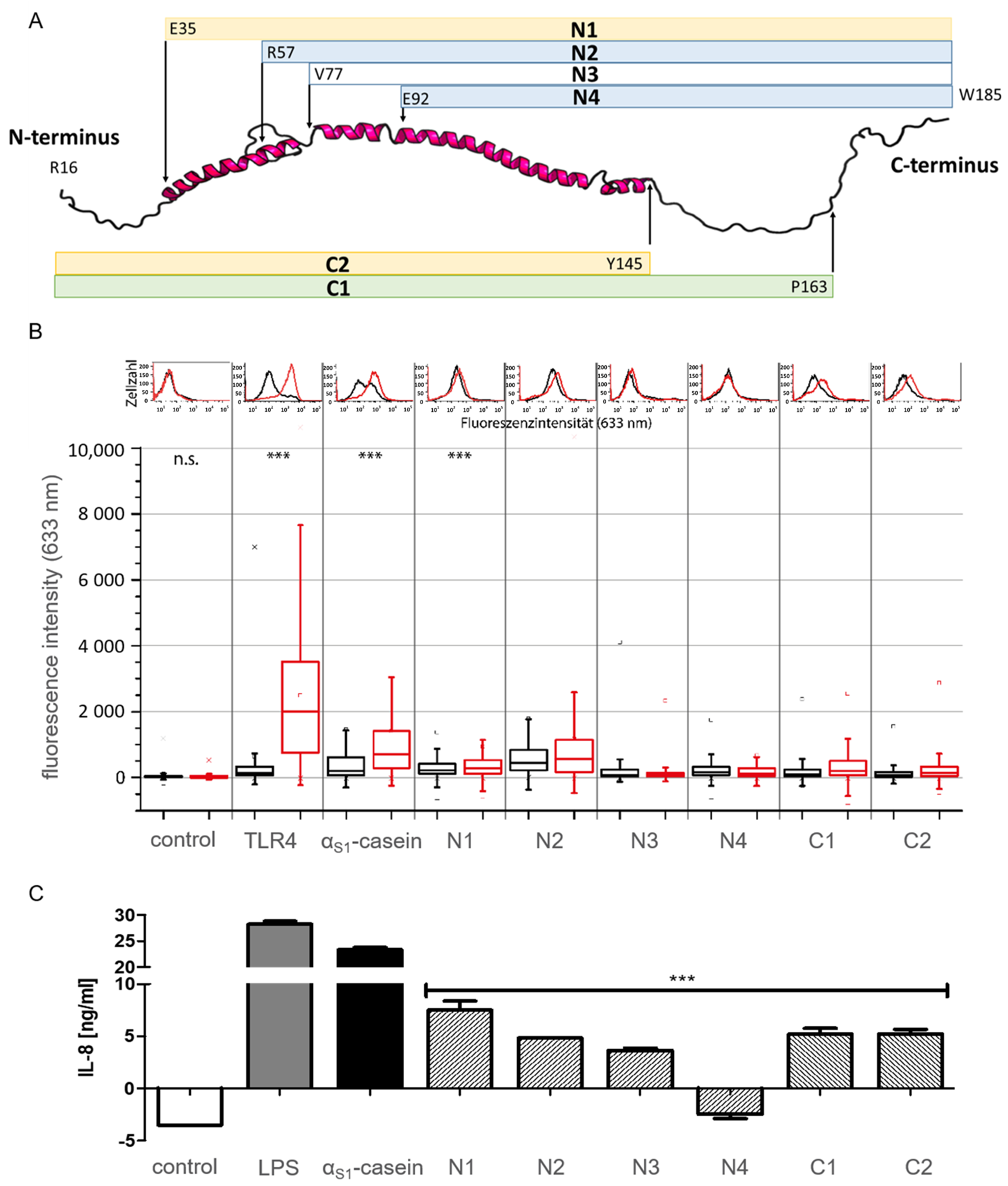
2.5. Identification of a TLR4-Stimulating Peptide Derived from αS1-Casein
3. Materials and Methods
3.1. Purification of Recombinant αS1-Casein, Construction and Isolation of Truncated αS1-Casein Variants
3.2. SDS-PAGE Analysis
3.3. Secondary Structure Analysis by CD- and ATR-FTIR-Spectroscopy
3.4. Microscale Thermophoresis Assay (MST)
3.5. Surface Plasmon Resonance Spectroscopy Assay (SPR)
3.6. Photon Correlation Spectroscopy
3.7. Cell Culture, Stimulation and Flow-Cytometric Binding Experiments
3.8. Studies on Human αS1-Casein Fibrillation (Thioflavin T and Atomic Force Microscopy)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Ungethuem, U.; Haeupl, T.; Witt, H.; Koczan, D.; Krenn, V.; Huber, H.; von Helversen, T.M.; Drungowski, M.; Seyfert, C.; Zacher, J.; et al. Molecular signatures and new candidates to target the pathogenesis of rheumatoid arthritis. Physiol. Genom. 2010, 42A, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, C.; Dehne, T.; Lindahl, A.; Brittberg, M.; Pruss, A.; Sittinger, M.; Ringe, J. Genome-wide expression profiling reveals new candidate genes associated with osteoarthritis. Osteoarthr. Cartil. 2010, 18, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Otaegui, D.; Mostafavi, S.; Bernard, C.C.; Lopez de Munain, A.; Mousavi, P.; Oksenberg, J.R.; Baranzini, S.E. Increased transcriptional activity of milk-related genes following the active phase of experimental autoimmune encephalomyelitis and multiple sclerosis. J. Immunol. 2007, 179, 4074–4082. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Ling, M.T.; Wang, X.; Wong, Y.C. Evidence of a novel biomarker, alpha s1-Casein, a milk protein, in benign prostate hyperplasia. Prostate Cancer Prostatic Dis. 2006, 9, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Bonuccelli, G.; Castello-Cros, R.; Capozza, F.; Martinez-Outschoorn, U.E.; Lin, Z.; Tsirigos, A.; Xuanmao, J.; Whitaker-Menezes, D.; Howell, A.; Lisanti, M.P.; et al. The milk protein alpha-casein functions as a tumor suppressor via activation of STAT1 signaling, effectively preventing breast cancer tumor growth and metastasis. Cell Cycle 2012, 11, 3972–3982. [Google Scholar] [CrossRef] [PubMed]
- Petermann, K.; Vordenbaumen, S.; Maas, R.; Braukmann, A.; Bleck, E.; Saenger, T.; Schneider, M.; Jose, J. Autoantibodies to alphaS1-casein are induced by breast-feeding. PLoS ONE 2012, 7, e32716. [Google Scholar] [CrossRef] [PubMed]
- Vordenbaumen, S.; Braukmann, A.; Altendorfer, I.; Bleck, E.; Jose, J.; Schneider, M. Human casein alpha s1 (CSN1S1) skews in vitro differentiation of monocytes towards macrophages. BMC Immunol. 2013, 14, 46. [Google Scholar] [CrossRef]
- Vordenbaumen, S.; Saenger, T.; Braukmann, A.; Tahan, T.; Bleck, E.; Jose, J.; Schneider, M. Human casein alpha s1 induces proinflammatory cytokine expression in monocytic cells by TLR4 signaling. Mol. Nutr. Food Res. 2016, 60, 1079–1089. [Google Scholar] [CrossRef]
- Saenger, T.; Vordenbäumen, S.; Genich, S.; Haidar, S.; Schulte, M.; Nienberg, C.; Bleck, E.; Schneider, M.; Jose, J. Human αS1-casein induces IL-8 secretion by binding to the ecto-domain of the TLR4/MD2 receptor complex. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2019, 1863, 632–643. [Google Scholar] [CrossRef]
- Cavaletto, M.; Cantisani, A.; Giuffrida, G.; Napolitano, L.; Conti, A. Human alpha S1-casein like protein: Purification and N-terminal sequence determination. Biol. Chem. Hoppe Seyler 1994, 375, 149–151. [Google Scholar]
- Rasmussen, L.K.; Due, H.A.; Petersen, T.E. Human αs1-casein: Purification and characterization. Comp. Biochem. Physiol. B Comp. Biochem. 1995, 111, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Armaforte, E.; Curran, E.; Huppertz, T.; Ryan, C.A.; Caboni, M.F.; O’Connor, P.M.; Ross, R.P.; Hirtz, C.; Sommerer, N.; Chevalier, F.; et al. Proteins and proteolysis in pre-term and term human milk and possible implications for infant formulae. Int. Dairy J. 2010, 20, 715–723. [Google Scholar] [CrossRef]
- Sorensen, E.S.; Moller, L.; Vinther, M.; Petersen, T.E.; Rasmussen, L.K. The phosphorylation pattern of human alphas1-casein is markedly different from the ruminant species. Eur. J. Biochem. 2003, 270, 3651–3655. [Google Scholar] [CrossRef] [PubMed]
- Chatterton, D.E.W.; Rasmussen, J.T.; Heegaard, C.W.; Sorensen, E.S.; Petersen, T.E. In vitro digestion of novel milk protein ingredients for use in infant formulas: Research on biological functions. Trends Food Sci. Technol. 2004, 15, 373–383. [Google Scholar] [CrossRef]
- Peila, C.; Coscia, A.; Bertino, E.; Cavaletto, M.; Spertino, S.; Icardi, S.; Tortone, C.; Visser, G.H.A.; Gazzolo, D. Effects of Holder pasteurization on the protein profile of human milk. Ital. J. Pediatr. 2016, 42, 36. [Google Scholar] [CrossRef] [PubMed]
- Altendorfer, I.; Konig, S.; Braukmann, A.; Saenger, T.; Bleck, E.; Vordenbaumen, S.; Kubiak, A.; Schneider, M.; Jose, J. Quantification of alphaS1-casein in breast milk using a targeted mass spectrometry-based approach. J. Pharm. Biomed. Anal. 2015, 103, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Creamer, L.K.; Richardson, T.; Parry, D.A.D. Secondary structure of bovine αS1- and β-casein in solution. Arch. Biochem. Biophys. 1981, 211, 689–696. [Google Scholar] [CrossRef]
- Carrotta, R.; Canale, C.; Diaspro, A.; Trapani, A.; Biagio, P.L.S.; Bulone, D. Inhibiting effect of αs1-casein on Aβ1–40 fibrillogenesis. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2012, 1820, 124–132. [Google Scholar] [CrossRef]
- Bingham, E.W.; Farrell, H.M., Jr.; Carroll, R.J. Properties of dephosphorylated s1-casein. Precipitation by calcium ions and micelle formation. Biochemistry 1972, 11, 2450–2454. [Google Scholar] [CrossRef]
- Farrell, H.M., Jr.; Jimenez-Flores, R.; Bleck, G.T.; Brown, E.M.; Butler, J.E.; Creamer, L.K.; Hicks, C.L.; Hollar, C.M.; Ng-Kwai-Hang, K.F.; Swaisgood, H.E. Nomenclature of the proteins of cows’ milk—Sixth revision. J. Dairy Sci. 2004, 87, 1641–1674. [Google Scholar] [CrossRef]
- Marchesseau, S.; Mani, J.C.; Martineau, P.; Roquet, F.; Cuq, J.L.; Pugniere, M. Casein interactions studied by the surface plasmon resonance technique. J. Dairy Sci. 2002, 85, 2711–2721. [Google Scholar] [CrossRef]
- Redwan, E.M.; Xue, B.; Almehdar, H.A.; Uversky, V.N. Disorder in milk proteins: Caseins, intrinsically disordered colloids. Curr. Protein Pept. Sci. 2015, 16, 228–242. [Google Scholar] [CrossRef] [PubMed]
- Schulmeister, U.; Hochwallner, H.; Swoboda, I.; Focke-Tejkl, M.; Geller, B.; Nystrand, M.; Harlin, A.; Thalhamer, J.; Scheiblhofer, S.; Keller, W.; et al. Cloning, expression, and mapping of allergenic determinants of alphaS1-casein, a major cow’s milk allergen. J. Immunol. 2009, 182, 7019–7029. [Google Scholar] [CrossRef] [PubMed]
- Stroylova, Y.Y.; Zimny, J.; Yousefi, R.; Chobert, J.-M.; Jakubowski, H.; Muronetz, V.I.; Haertlé, T. Aggregation and structural changes of αS1-, β- and κ-caseins induced by homocysteinylation. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2011, 1814, 1234–1245. [Google Scholar] [CrossRef] [PubMed]
- Slattery, C.W.; Evard, R. A model for the formation and structure of casein micelles from subunits of variable composition. Biochim. Biophys. Acta 1973, 317, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Docena, G.H.; Fernandez, R.; Chirdo, F.G.; Fossati, C.A. Identification of casein as the major allergenic and antigenic protein of cow’s milk. Allergy 1996, 51, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Saenger, T.; Braukmann, A.; Vordenbaumen, S.; Altendorfer, I.; Bleck, E.; Hochwallner, H.; Valenta, R.; Schneider, M.; Jose, J. Development of a surface display ELISA to detect anti-IgG antibodies against bovine alphaS1-casein in human sera. J. Pharm. Biomed. Anal. 2014, 96, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Margulies, E.H.; Cooper, G.M.; Asimenos, G.; Thomas, D.J.; Dewey, C.N.; Siepel, A.; Birney, E.; Keefe, D.; Schwartz, A.S.; Hou, M.; et al. Analyses of deep mammalian sequence alignments and constraint predictions for 1% of the human genome. Genome Res. 2007, 17, 760–774. [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- König, S.; Altendorfer, I.; Saenger, T.; Bleck, E.; Vordenbäumen, S.; Schneider, M.; Jose, J. Ser71 of αS1-Casein is Phosphorylated in Breast Milk—Evidence from Targeted Mass Analysis. Mol. Nutr. Food Res. 2017, 61, 1700496. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Xu, J. RaptorX: Exploiting structure information for protein alignment by statistical inference. Proteins 2011, 79 (Suppl. 10), 161–171. [Google Scholar] [CrossRef] [PubMed]
- Kallberg, M.; Margaryan, G.; Wang, S.; Ma, J.; Xu, J. RaptorX server: A resource for template-based protein structure modeling. Methods Mol. Biol. 2014, 1137, 17–27. [Google Scholar] [PubMed]
- Perdigão, N.; Rosa, A. Dark Proteome Database: Studies on Dark Proteins. High Throughput 2019, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Ferron, F.; Longhi, S.; Canard, B.; Karlin, D. A practical overview of protein disorder prediction methods. Proteins 2006, 65, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; London, E. An amino acid “transmembrane tendency” scale that approaches the theoretical limit to accuracy for prediction of transmembrane helices: Relationship to biological hydrophobicity. Protein Sci. Publ. Protein Soc. 2006, 15, 1987–2001. [Google Scholar] [CrossRef] [PubMed]
- Louis-Jeune, C.; Andrade-Navarro, M.A.; Perez-Iratxeta, C. Prediction of protein secondary structure from circular dichroism using theoretically derived spectra. Proteins 2012, 80, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Greenfield, N.J. Using circular dichroism spectra to estimate protein secondary structure. Nat. Protoc. 2006, 1, 2876–2890. [Google Scholar] [CrossRef]
- Holt, C.; Carver, J.A.; Ecroyd, H.; Thorn, D.C. Invited review: Caseins and the casein micelle: Their biological functions, structures, and behavior in foods. J. Dairy Sci. 2013, 96, 6127–6146. [Google Scholar] [CrossRef]
- Nowakowski, A.B.; Wobig, W.J.; Petering, D.H. Native SDS-PAGE: High resolution electrophoretic separation of proteins with retention of native properties including bound metal ions. Metallomics 2014, 6, 1068–1078. [Google Scholar] [CrossRef]
- Rasmussen, L.K.; Hojrup, P.; Petersen, T.E. Disulphide arrangement in bovine caseins: Localization of intrachain disulphide bridges in monomers of kappa- and alpha s2-casein from bovine milk. J. Dairy Res. 1994, 61, 485–493. [Google Scholar] [CrossRef]
- Farrell, H.M., Jr.; Qi, P.X.; Brown, E.M.; Cooke, P.H.; Tunick, M.H.; Wickham, E.D.; Unruh, J.J. Molten globule structures in milk proteins: Implications for potential new structure-function relationships. J. Dairy Sci. 2002, 85, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Alexander, C.G.; Wanner, R.; Johnson, C.M.; Breitsprecher, D.; Winter, G.; Duhr, S.; Baaske, P.; Ferguson, N. Novel microscale approaches for easy, rapid determination of protein stability in academic and commercial settings. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2014, 1844, 2241–2250. [Google Scholar] [CrossRef] [PubMed]
- Malin, E.L.; Brown, E.M.; Wickham, E.D.; Farrell, H.M., Jr. Contributions of terminal peptides to the associative behavior of alphas1-casein. J. Dairy Sci. 2005, 88, 2318–2328. [Google Scholar] [CrossRef]
- Jakob, U.; Kriwacki, R.; Uversky, V.N. Conditionally and transiently disordered proteins: Awakening cryptic disorder to regulate protein function. Chem. Rev. 2014, 114, 6779–6805. [Google Scholar] [CrossRef]
- Mason, S. Some aspects of gastric function in the newborn. Arch Dis. Child. 1962, 37, 387–391. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Stroopinsky, D.; Yin, L.; Rosenblatt, J.; Alam, M.; Bhargava, P.; Clark, R.A.; Kupper, T.S.; Palmer, K.; Coll, M.D.; et al. Mucin 1 is a potential therapeutic target in cutaneous T-cell lymphoma. Blood 2015, 126, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Mason, J.M.; Arndt, K.M. Coiled coil domains: Stability, specificity, and biological implications. Chembiochem 2004, 5, 170–176. [Google Scholar] [CrossRef]
- Gruber, M.; Soding, J.; Lupas, A.N. Comparative analysis of coiled-coil prediction methods. J. Struct. Biol. 2006, 155, 140–145. [Google Scholar] [CrossRef]
- Alva, V.; Nam, S.Z.; Soding, J.; Lupas, A.N. The MPI bioinformatics Toolkit as an integrative platform for advanced protein sequence and structure analysis. Nucleic Acids Res. 2016, 44, W410–W415. [Google Scholar] [CrossRef]
- Lupas, A. Prediction and analysis of coiled-coil structures. Methods Enzymol. 1996, 266, 513–525. [Google Scholar] [PubMed]
- Fariselli, P.; Molinini, D.; Casadio, R.; Krogh, A. Prediction of Structurally-Determined Coiled-Coil Domains with Hidden Markov Models. In Bioinformatics Research and Development: First International Conference, BIRD 2007, Berlin, Germany, 12–14 March 2007. Proceedings; Hochreiter, S., Wagner, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 292–302. [Google Scholar]
- Sonnichsen, F.D.; Van Eyk, J.E.; Hodges, R.S.; Sykes, B.D. Effect of trifluoroethanol on protein secondary structure: An NMR and CD study using a synthetic actin peptide. Biochemistry 1992, 31, 8790–8798. [Google Scholar] [CrossRef] [PubMed]
- Luidens, M.K.; Figge, J.; Breese, K.; Vajda, S. Predicted and trifluoroethanol-induced alpha-helicity of polypeptides. Biopolymers 1996, 39, 367–376. [Google Scholar] [CrossRef]
- Greenfield, N.J.; Hitchcock-DeGregori, S.E. Conformational intermediates in the folding of a coiled-coil model peptide of the N-terminus of tropomyosin and alpha alpha-tropomyosin. Protein Sci. Publ. Protein Soc. 1993, 2, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- Barth, A. Infrared spectroscopy of proteins. Biochim. Biophys. Acta (BBA)-Bioenerg. 2007, 1767, 1073–1101. [Google Scholar] [CrossRef] [PubMed]
- Jerabek-Willemsen, M.; Wienken, C.J.; Braun, D.; Baaske, P.; Duhr, S. Molecular interaction studies using microscale thermophoresis. Assay Drug Dev. Technol. 2011, 9, 342–353. [Google Scholar] [CrossRef]
- Sood, S.M.; Herbert, P.J.; Slattery, C.W. Structural studies on casein micelles of human milk: Dissociation of beta-casein of different phosphorylation levels induced by cooling and ethylenediaminetetraacetate. J. Dairy Sci. 1997, 80, 628–633. [Google Scholar] [CrossRef]
- Nielsen, L.; Khurana, R.; Coats, A.; Frokjaer, S.; Brange, J.; Vyas, S.; Uversky, V.N.; Fink, A.L. Effect of Environmental Factors on the Kinetics of Insulin Fibril Formation: Elucidation of the Molecular Mechanism. Biochemistry 2001, 40, 6036–6046. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resulting Protein | Corresponding to AAS of αS1-Casein | Plasmid | Oligonucleotide |
|---|---|---|---|
| N1 | 35–185 | pET N1 | Fw: CACCATCACCATCATGAGCCTATACCATTAGAATCAAGAGAGGAA Rv: TAGCASGCCGGATCCGTTACCACTGTAGCATGACGTTATTTTTTTCATA |
| N2 | 57–185 | pET N2 | Fw: CACCATCACCATCATAGAGAAAAACAGACTGATGAAATCAAGGATACTAGG: Rv: TAGCASGCCGGATCCGTTACCACTGTAGCATGACGTTATTTTTTTCATA |
| N3 | 77–185 | pET N3 | Fw: CACCATCACCATCATGAAATGTCTCTCAGTAAGTGTGCGGAACAG Rv: TAGCASGCCGGATCCGTTACCACTGTAGCATGACGTTATTTTTTTCATA |
| N4 | 93–185 | pET N4 | Fw: CACCATCACCATCATTTGTGGCAGAGCCTGAGAAG Rv: TAGCASGCCGGATCCGTTACCACTGTAGCATGACGTTATTTTTTTCATA |
| C1 | 16–163 | pET C1 | Fw: GTGGTGGTGCTCGAGCGGTGGGAAAGGAACATAC Rv: CTCGAGCACCACCACC |
| C2 | 16–145 | pET C2 | Fw: GTGGTGCTCGAGGTAGGCAGCAAGTTGGTT Rv: CTCGAGCACCACCACC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saenger, T.; Schulte, M.F.; Vordenbäumen, S.; Herrmann, F.C.; Bertelsbeck, J.; Meier, K.; Bleck, E.; Schneider, M.; Jose, J. Structural Analysis of Breast-Milk αS1-Casein: An α-Helical Conformation Is Required for TLR4-Stimulation. Int. J. Mol. Sci. 2024, 25, 1743. https://doi.org/10.3390/ijms25031743
Saenger T, Schulte MF, Vordenbäumen S, Herrmann FC, Bertelsbeck J, Meier K, Bleck E, Schneider M, Jose J. Structural Analysis of Breast-Milk αS1-Casein: An α-Helical Conformation Is Required for TLR4-Stimulation. International Journal of Molecular Sciences. 2024; 25(3):1743. https://doi.org/10.3390/ijms25031743
Chicago/Turabian StyleSaenger, Thorsten, Marten F. Schulte, Stefan Vordenbäumen, Fabian C. Herrmann, Juliana Bertelsbeck, Kathrin Meier, Ellen Bleck, Matthias Schneider, and Joachim Jose. 2024. "Structural Analysis of Breast-Milk αS1-Casein: An α-Helical Conformation Is Required for TLR4-Stimulation" International Journal of Molecular Sciences 25, no. 3: 1743. https://doi.org/10.3390/ijms25031743
APA StyleSaenger, T., Schulte, M. F., Vordenbäumen, S., Herrmann, F. C., Bertelsbeck, J., Meier, K., Bleck, E., Schneider, M., & Jose, J. (2024). Structural Analysis of Breast-Milk αS1-Casein: An α-Helical Conformation Is Required for TLR4-Stimulation. International Journal of Molecular Sciences, 25(3), 1743. https://doi.org/10.3390/ijms25031743