
Estrogen Signaling Inhibits the Expression of anti-Müllerian hormone (amh) and gonadal-soma-derived factor (gsdf) during the Critical Time of Sexual Fate Determination in Zebrafish

 ,
,
Abstract
1. Introduction
2. Results
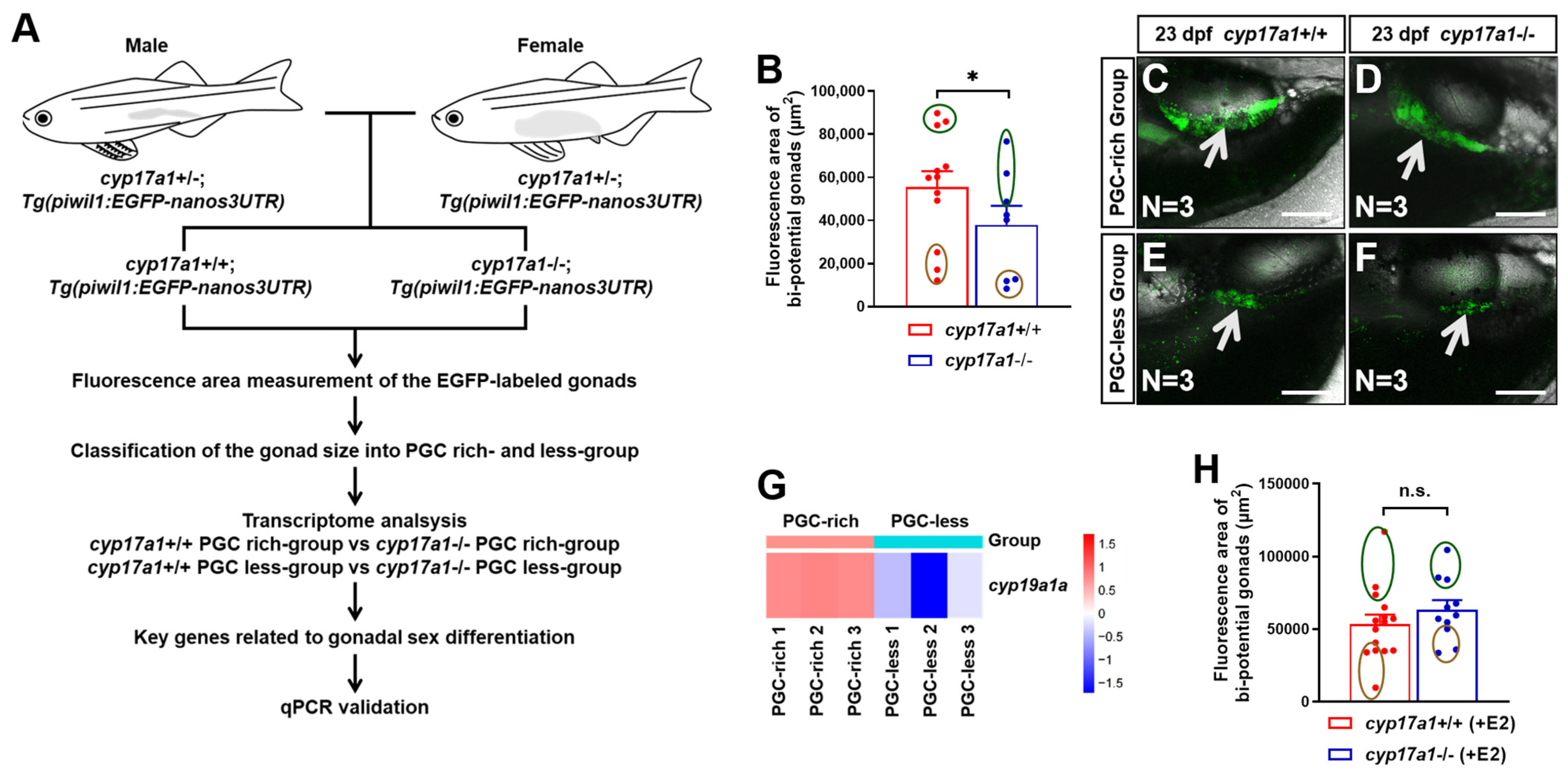
2.1. Estrogen Enlarged the Bi-Potential Gonad in Zebrafish
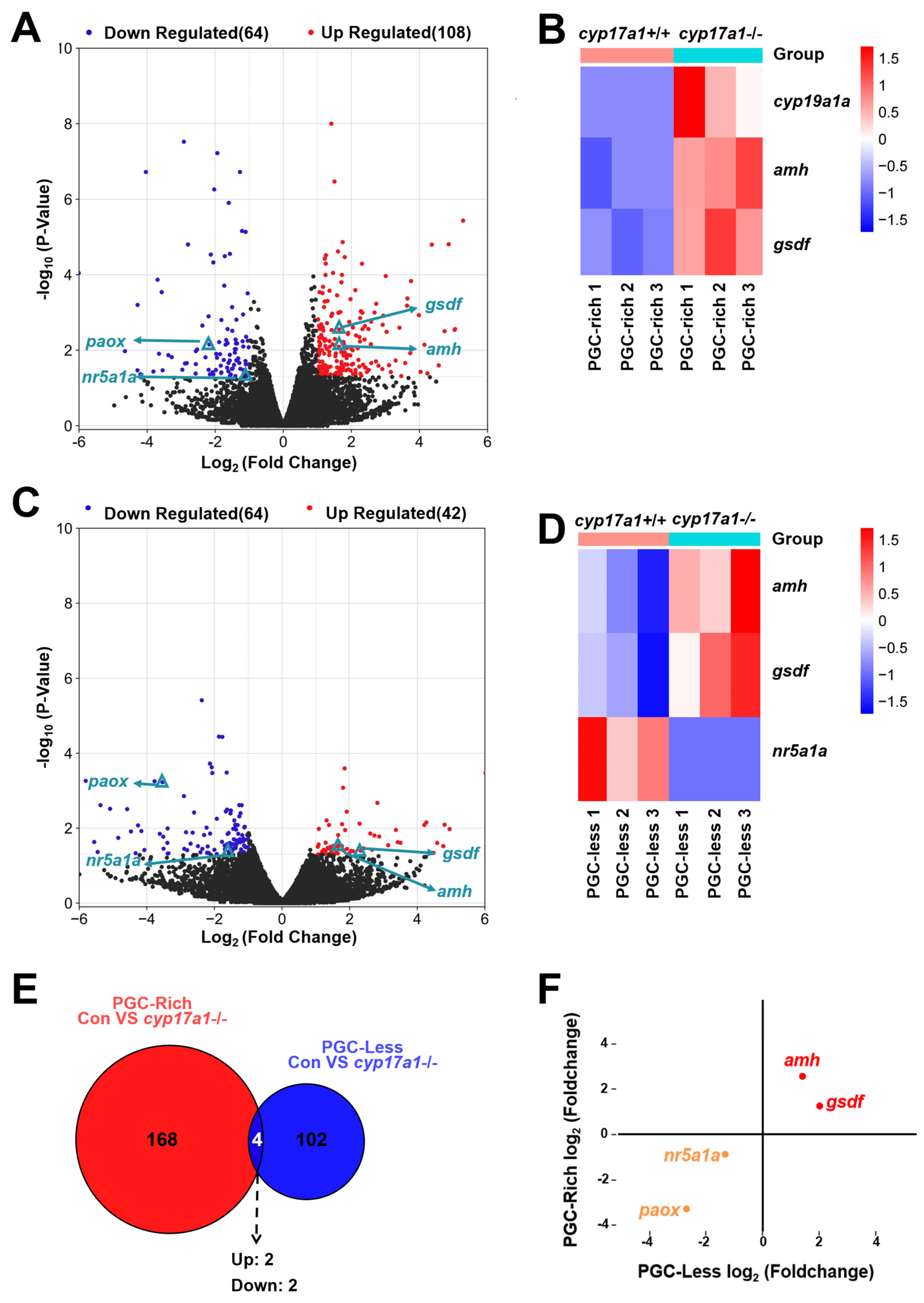
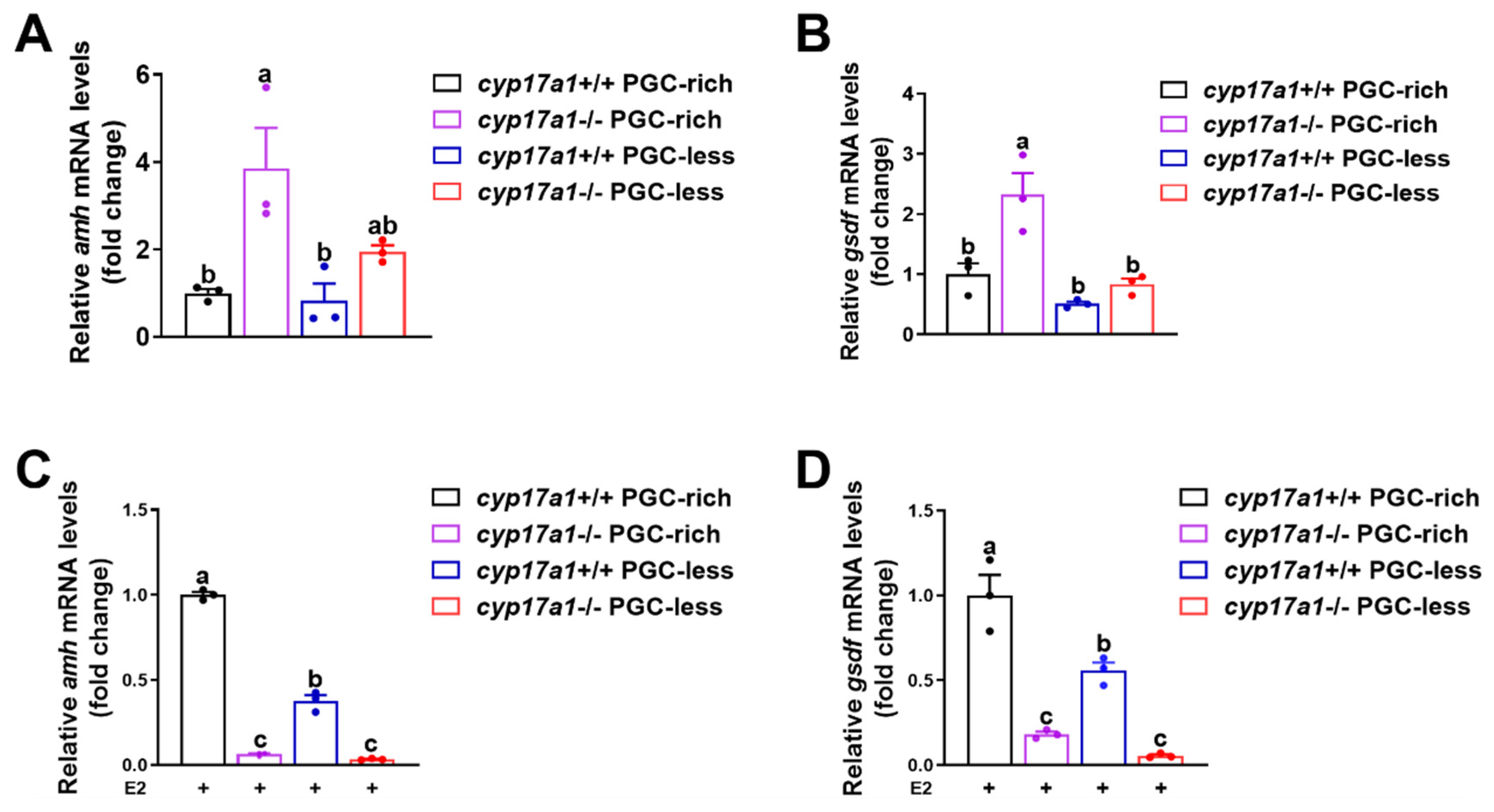
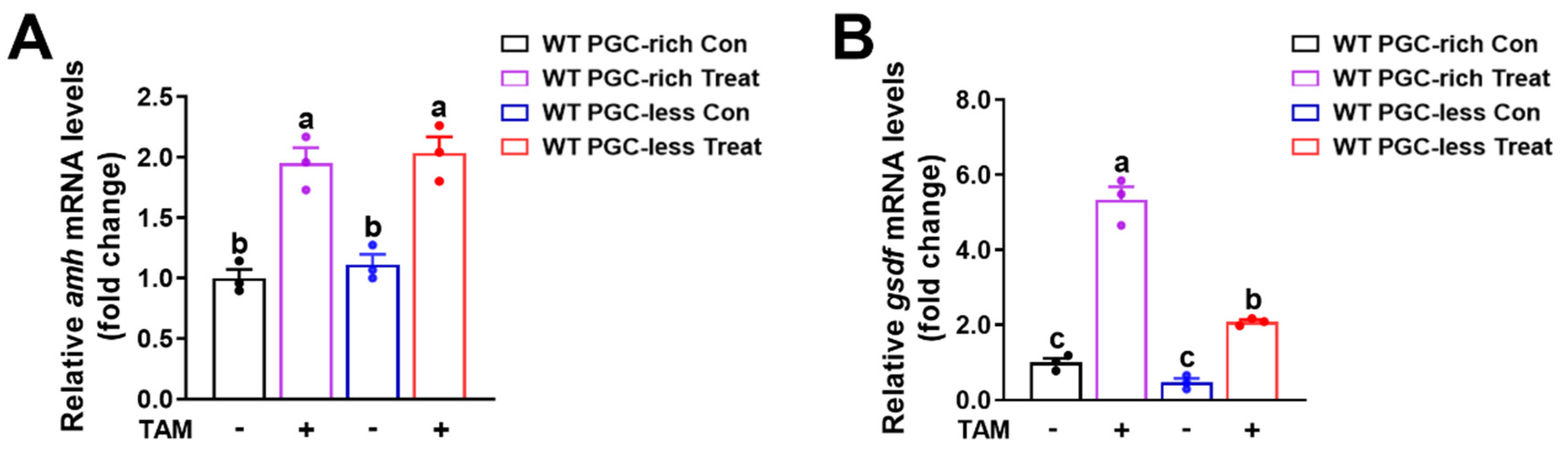
2.2. Upregulated Expression of amh and gsdf Was Observed in cyp17a1−/− Fish of the PGC-Rich Group
2.3. Estrogen Treatment Downregulated the Expression of amh and gsdf in cyp17a1−/− Zebrafish Gonadal Tissues
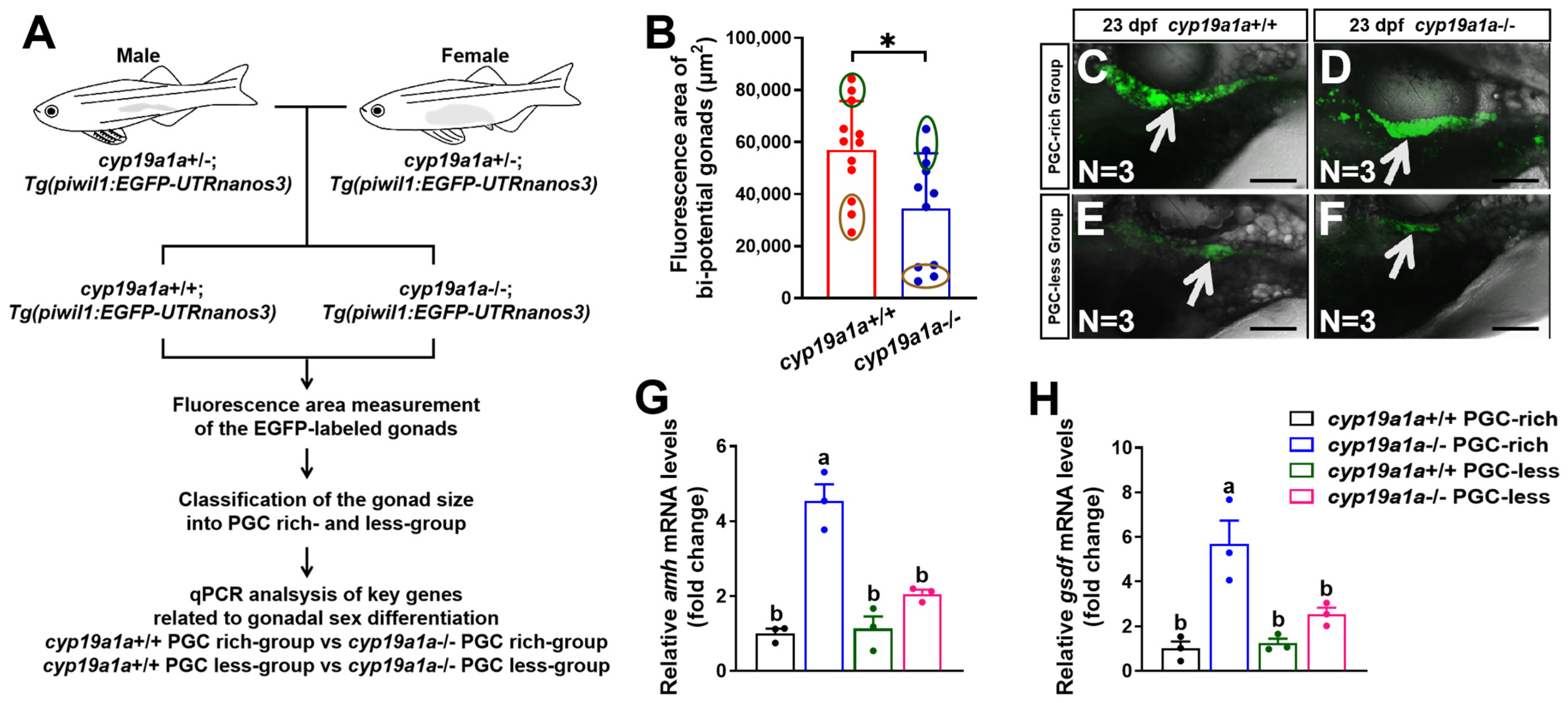
2.4. Upregulated Expression of amh and gsdf Was Observed in cyp19a1a−/− Fish of the PGC-Rich Group
2.5. Estradiol Inhibited the Expression of amh and gsdf in Zebrafish via Estrogen Receptor
3. Discussion
4. Materials and Methods
4.1. Fish Stocks
4.2. Collection of Samples
4.3. Transcriptome Analysis
4.4. Validation of Differential Gene Expression by Quantitative Real-Time PCR
4.5. 17β-Estradiol Administration
4.6. Estrogen Receptor Inhibitor Administration
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dai, S.; Qi, S.; Wei, X.; Liu, X.; Li, Y.; Zhou, X.; Xiao, H.; Lu, B.; Wang, D.; Li, M. Germline sexual fate is determined by the antagonistic action of dmrt1 and foxl3/foxl2 in tilapia. Development 2021, 148, dev199380. [Google Scholar] [CrossRef]
- Bell, G. The Masterpiece of Nature: The Evolution and Genetics of Sexuality; The University of California Press: Berkeley, CA, USA, 1982; pp. 1–635. [Google Scholar]
- Kikuchi, K.; Hamaguchi, S. Novel sex-determining genes in fish and sex chromosome evolution. Dev. Dyn. 2013, 242, 339–353. [Google Scholar] [CrossRef]
- Bull, J.J. Sex determining mechanisms: An evolutionary perspective. Experientia 1985, 41, 1285–1296. [Google Scholar] [CrossRef] [PubMed]
- Liew, W.C.; Orban, L. Zebrafish sex: A complicated affair. Brief. Funct. Genom. 2014, 13, 172–187. [Google Scholar] [CrossRef]
- Vandeputte, M.; Dupont-Nivet, M.; Chavanne, H.; Chatain, B. A polygenic hypothesis for sex determination in the European sea bass Dicentrarchus labrax. Genetics 2007, 176, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Ser, J.R.; Roberts, R.B.; Kocher, T.D. Multiple interacting loci control sex determination in lake Malawi cichlid fish. Evolution 2010, 64, 486–501. [Google Scholar] [CrossRef]
- Guerriero, G. Vertebrate sex steroid receptors: Evolution, ligands, and neurodistribution. Ann. N. Y. Acad. Sci. 2009, 1163, 154–168. [Google Scholar] [CrossRef]
- Wilson, C.A.; High, S.K.; McCluskey, B.M.; Amores, A.; Yan, Y.-l.; Titus, T.A.; Anderson, J.L.; Batzel, P.; Carvan, M.J.; Schartl, M.; et al. Wild sex in zebrafish: Loss of the natural sex determinant in domesticated strains. Genetics 2014, 198, 1291–1308. [Google Scholar] [CrossRef]
- Fenske, M.; Segner, H. Aromatase modulation alters gonadal differentiation in developing zebrafish (Danio rerio). Aquat. Toxicol. 2004, 67, 105–126. [Google Scholar] [CrossRef] [PubMed]
- Uchida, D.; Yamashita, M.; Kitano, T.; Iguchi, T. An aromatase inhibitor or high water temperature induce oocyte apoptosis and depletion of P450 aromatase activity in the gonads of genetic female zebrafish during sex-reversal. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2004, 137, 11–20. [Google Scholar] [CrossRef]
- Zhai, G.; Shu, T.; Xia, Y.; Lu, Y.; Shang, G.; Jin, X.; He, J.; Nie, P.; Yin, Z. Characterization of Sexual Trait Development in cyp17a1-Deficient Zebrafish. Endocrinology 2018, 159, 3549–3562. [Google Scholar] [CrossRef]
- Lau, E.S.-W.; Zhang, Z.; Qin, M.; Ge, W. Knockout of Zebrafish Ovarian Aromatase Gene (cyp19a1a) by TALEN and CRISPR/Cas9 Leads to All-male Offspring Due to Failed Ovarian Differentiation. Sci. Rep. 2016, 6, 37357. [Google Scholar] [CrossRef]
- Dranow, D.B.; Hu, K.; Bird, A.M.; Lawry, S.T.; Adams, M.T.; Sanchez, A.; Amatruda, J.F.; Draper, B.W. Bmp15 Is an Oocyte-Produced Signal Required for Maintenance of the Adult Female Sexual Phenotype in Zebrafish. PLoS Genet. 2016, 12, e1006323. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Tang, H.; Liu, Y.; Chen, Y.; Li, G.; Liu, X.; Lin, H. Targeted Disruption of Aromatase Reveals Dual Functions of cyp19a1a During Sex Differentiation in Zebrafish. Endocrinology 2017, 158, 3030–3041. [Google Scholar] [CrossRef]
- Yu, G.; Zhang, D.; Liu, W.; Wang, J.; Liu, X.; Zhou, C.; Gui, J.; Xiao, W. Zebrafish androgen receptor is required for spermatogenesis and maintenance of ovarian function. Oncotarget 2018, 9, 24320–24334. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Chen, Y.; Wang, L.; Yin, Y.; Li, G.; Guo, Y.; Liu, Y.; Lin, H.; Cheng, C.H.K.; Liu, X. Fertility impairment with defective spermatogenesis and steroidogenesis in male zebrafish lacking androgen receptor. Biol. Reprod. 2018, 98, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Webster, K.A.; Schach, U.; Ordaz, A.; Steinfeld, J.S.; Draper, B.W.; Siegfried, K.R. Dmrt1 is necessary for male sexual development in zebrafish. Dev. Biol. 2017, 422, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.H.; Mei, J.; Li, Z.; Zhang, X.M.; Zhou, L.; Gui, J.F. Distinct and Cooperative Roles of amh and dmrt1 in Self-Renewal and Differentiation of Male Germ Cells in Zebrafish. Genetics 2017, 207, 1007–1022. [Google Scholar] [CrossRef]
- Skaar, K.S.; Nobrega, R.H.; Magaraki, A.; Olsen, L.C.; Schulz, R.W.; Male, R. Proteolytically activated, recombinant anti-mullerian hormone inhibits androgen secretion, proliferation, and differentiation of spermatogonia in adult zebrafish testis organ cultures. Endocrinology 2011, 152, 3527–3540. [Google Scholar] [CrossRef]
- Rodríguez-Marí, A.; Cañestro, C.; BreMiller, R.A.; Nguyen-Johnson, A.; Asakawa, K.; Kawakami, K.; Postlethwait, J.H. Sex reversal in zebrafish fancl mutants is caused by Tp53-mediated germ cell apoptosis. PLoS Genet. 2010, 6, e1001034. [Google Scholar] [CrossRef]
- Zhang, Z.W.; Zhu, B.; Chen, W.T.; Ge, W. Anti-Mullerian hormone (Amh/amh) plays dual roles in maintaining gonadal homeostasis and gametogenesis in zebrafish. Mol. Cell. Endocrinol. 2020, 517, 110963. [Google Scholar] [CrossRef]
- Zhang, Z.W.; Wu, K.; Ren, Z.Q.; Ge, W. Genetic evidence for Amh modulation of gonadotropin actions to control gonadal homeostasis and gametogenesis in zebrafish and its noncanonical signaling through Bmpr2a receptor. Development 2020, 147, dev189811. [Google Scholar] [CrossRef]
- Yan, Y.L.; Desvignes, T.; Bremiller, R.; Wilson, C.; Dillon, D.; High, S.; Draper, B.; Buck, C.L.; Postlethwait, J. Gonadal soma controls ovarian follicle proliferation through Gsdf in zebrafish. Dev. Dynam. 2017, 246, 925–945. [Google Scholar] [CrossRef]
- Clelland, E.; Peng, C. Endocrine/paracrine control of zebrafish ovarian development. Mol. Cell. Endocrinol. 2009, 312, 42–52. [Google Scholar] [CrossRef]
- Orban, L.; Sreenivasan, R.; Olsson, P.E. Long and winding roads: Testis differentiation in zebrafish. Mol. Cell. Endocrinol. 2009, 312, 35–41. [Google Scholar] [CrossRef]
- Kossack, M.E.; Draper, B.W. Genetic regulation of sex determination and maintenance in zebrafish (Danio rerio). Curr. Top. Dev. Biol. 2019, 134, 119–149. [Google Scholar] [PubMed]
- Ye, D.; Zhu, L.; Zhang, Q.; Xiong, F.; Wang, H.; Wang, X.; He, M.; Zhu, Z.; Sun, Y. Abundance of Early Embryonic Primordial Germ Cells Promotes Zebrafish Female Differentiation as Revealed by Lifetime Labeling of Germline. Mar. Biotechnol. 2019, 21, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Carlson, R.W. Scientific review of tamoxifen. Overview from a medical oncologist. Semin. Oncol. 1997, 24, S1-151–S1-157. [Google Scholar] [PubMed]
- Xu, S.; Xie, F.; Tian, L.; Fallah, S.; Babaei, F.; Manno, S.H.C.; Manno, F.A.M.; Zhu, L.; Wong, K.F.; Liang, Y.; et al. Estrogen accelerates heart regeneration by promoting the inflammatory response in zebrafish. J. Endocrinol. 2020, 245, 39–51. [Google Scholar] [CrossRef]
- Yin, N.; Jin, X.; He, J.; Yin, Z. Effects of adrenergic agents on the expression of zebrafish (Danio rerio) vitellogenin Ao1. Toxicol. Appl. Pharmacol. 2009, 238, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Liu, L.; Ge, W. Expression analysis of growth differentiation factor 9 (Gdf9/gdf9), anti-mullerian hormone (Amh/amh) and aromatase (Cyp19a1a/cyp19a1a) during gonadal differentiation of the zebrafish, Danio rerio. Biol. Reprod. 2017, 96, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Uchida, D.; Yamashita, M.; Kitano, T.; Iguchi, T. Oocyte apoptosis during the transition from ovary-like tissue to testes during sex differentiation of juvenile zebrafish. J. Exp. Biol. 2002, 205, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Jin, X.; Chen, X.; He, J.; Yin, Z. Sufficient numbers of early germ cells are essential for female sex development in zebrafish. PLoS ONE 2015, 10, e0117824. [Google Scholar] [CrossRef] [PubMed]
- Tzung, K.W.; Goto, R.; Saju, J.M.; Sreenivasan, R.; Saito, T.; Arai, K.; Yamaha, E.; Hossain, M.S.; Calvert, M.E.K.; Orban, L. Early depletion of primordial germ cells in zebrafish promotes testis formation. Stem Cell Rep. 2015, 4, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Slanchev, K.; Stebler, J.; de la Cueva-Mendez, G.; Raz, E. Development without germ cells: The role of the germ line in zebrafish sex differentiation. Proc. Natl. Acad. Sci. USA 2005, 102, 4074–4079. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, K.R.; Nusslein-Volhard, C. Germ line control of female sex determination in zebrafish. Dev. Biol. 2008, 324, 277–287. [Google Scholar] [CrossRef]
- Ding, Y.; Yixuan, T.; Houpeng, W.; Mudan, H.; Yaqing, W.; Zhengfang, C.; Zhenxia, C.; Yonghua, S. A landscape of differentiated biological processes involved in the initiation of sex differentiation in zebrafish. Water Biol. Secur. 2022, 1, 100059. [Google Scholar]
- Brion, F.; Tyler, C.R.; Palazzi, X.; Laillet, B.; Porcher, J.M.; Garric, J.; Flammarion, P. Impacts of 17beta-estradiol, including environmentally relevant concentrations, on reproduction after exposure during embryo-larval-, juvenile- and adult-life stages in zebrafish (Danio rerio). Aquat. Toxicol. 2004, 68, 193–217. [Google Scholar] [CrossRef]
- Wu, K.; Song, W.Y.; Zhang, Z.W.; Ge, W. Disruption of dmrt1 rescues the all-male phenotype of the cyp19a1a mutant in zebrafish—A novel insight into the roles of aromatase/estrogens in gonadal differentiation and early folliculogenesis. Development 2020, 147, dev182758. [Google Scholar] [CrossRef]
- Wang, Y.; Ye, D.; Zhang, F.; Zhang, R.; Zhu, J.; Wang, H.; He, M.; Sun, Y. Cyp11a2 Is Essential for Oocyte Development and Spermatogonial Stem Cell Differentiation in Zebrafish. Endocrinology 2022, 163, bqab258. [Google Scholar] [CrossRef]
- Yan, Y.-L.; Titus, T.; Desvignes, T.; BreMiller, R.; Batzel, P.; Sydes, J.; Farnsworth, D.; Dillon, D.; Wegner, J.; Phillips, J.B.; et al. A fish with no sex: Gonadal and adrenal functions partition between zebrafish NR5A1 co-orthologs. Genetics 2021, 217, iyaa030. [Google Scholar] [CrossRef] [PubMed]
- Che, H.; Ma, C.; Li, H.; Yu, F.; Wei, Y.; Chen, H.; Wu, J.; Ren, Y. Rebalance of the Polyamine Metabolism Suppresses Oxidative Stress and Delays Senescence in Nucleus Pulposus Cells. Oxidative Med. Cell. Longev. 2022, 2022, 8033353. [Google Scholar] [CrossRef] [PubMed]
- Nakajin, S.; Hall, P.F.; Onoda, M. Testicular microsomal cytochrome P-450 for C21 steroid side chain cleavage. Spectral and binding studies. J. Biol. Chem. 1981, 256, 6134–6139. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. The Zebrafish Book, a Guide for the Laboratory Use of Zebrafish (Danio rerio) OR, 4th ed.; University of Oregon: Eugene, OR, USA, 2020. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhai, G.; Shu, T.; Xia, Y.; Jin, X.; He, J.; Yin, Z. Androgen signaling regulates the transcription of anti-Müllerian hormone via synergy with SRY-related protein SOX9A. Sci. Bull. 2017, 62, 197–203. [Google Scholar] [CrossRef]
- Presslauer, C.; Nagasawa, K.; Dahle, D.; Babiak, J.; Fernandes, J.M.O.; Babiak, I. Induced autoimmunity against gonadal proteins affects gonadal development in juvenile zebrafish. PLoS ONE 2014, 9, e114209. [Google Scholar] [CrossRef]
- Shi, C.; Lu, Y.; Zhai, G.; Huang, J.; Shang, G.; Lou, Q.; Li, D.; Jin, X.; He, J.; Du, Z.; et al. Hyperandrogenism in POMCa-deficient zebrafish enhances somatic growth without increasing adiposity. J. Mol. Cell Biol. 2020, 12, 291–304. [Google Scholar] [CrossRef]
- Xia, L.; Zheng, L.; Zhou, J.L. Transcriptional and morphological effects of tamoxifen on the early development of zebrafish (Danio rerio). J. Appl. Toxicol. 2016, 36, 853–862. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Direction a and Sequence (5′-3′) | Product Size (bp b) | Reference |
|---|---|---|---|
| qPCR | |||
| amh | F: GCTGGGGAACTGGGGAAAAT | 198 | [47] |
| R: CGGTAGGGAATGCTTTGGGA | |||
| gsdf | F: GAACGCTCCTGAATCCACAGAC | 210 | [48] |
| R: AATGACTCCCGCAGATGCTC | |||
| β-actin | F: ACTCAGGATGCGGAAACTGG | 118 | [49] |
| R: AGGGCAAAGTGGTAAACGCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruan, Y.; Li, X.; Zhai, G.; Lou, Q.; Jin, X.; He, J.; Yin, Z. Estrogen Signaling Inhibits the Expression of anti-Müllerian hormone (amh) and gonadal-soma-derived factor (gsdf) during the Critical Time of Sexual Fate Determination in Zebrafish. Int. J. Mol. Sci. 2024, 25, 1740. https://doi.org/10.3390/ijms25031740
Ruan Y, Li X, Zhai G, Lou Q, Jin X, He J, Yin Z. Estrogen Signaling Inhibits the Expression of anti-Müllerian hormone (amh) and gonadal-soma-derived factor (gsdf) during the Critical Time of Sexual Fate Determination in Zebrafish. International Journal of Molecular Sciences. 2024; 25(3):1740. https://doi.org/10.3390/ijms25031740
Chicago/Turabian StyleRuan, Yonglin, Xuehui Li, Gang Zhai, Qiyong Lou, Xia Jin, Jiangyan He, and Zhan Yin. 2024. "Estrogen Signaling Inhibits the Expression of anti-Müllerian hormone (amh) and gonadal-soma-derived factor (gsdf) during the Critical Time of Sexual Fate Determination in Zebrafish" International Journal of Molecular Sciences 25, no. 3: 1740. https://doi.org/10.3390/ijms25031740
APA StyleRuan, Y., Li, X., Zhai, G., Lou, Q., Jin, X., He, J., & Yin, Z. (2024). Estrogen Signaling Inhibits the Expression of anti-Müllerian hormone (amh) and gonadal-soma-derived factor (gsdf) during the Critical Time of Sexual Fate Determination in Zebrafish. International Journal of Molecular Sciences, 25(3), 1740. https://doi.org/10.3390/ijms25031740