
Recent Progress Regarding Jasmonates in Tea Plants: Biosynthesis, Signaling, and Function in Stress Responses


Abstract
1. Introduction
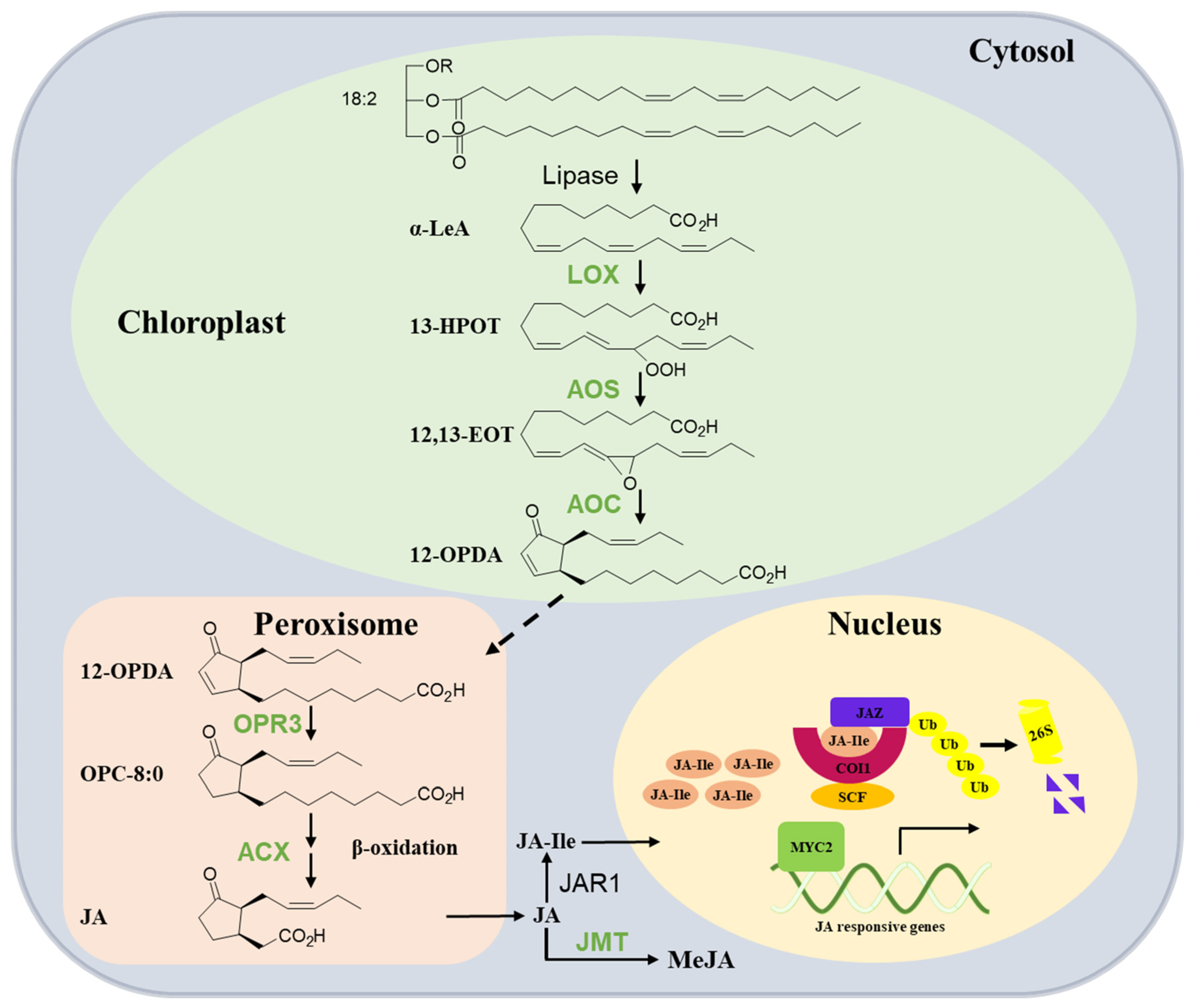
2. Research Advances in JA Biosynthesis

{kind=link}
{kind=link}
| Gene | Gene ID of Tea Plants | Gene Number in Tea Plants | Functions in Tea Plants | Functions in Other Species | References |
|---|---|---|---|---|---|
| LOX | HM440161 (Genbank) | 1 | located in chloroplast; converted LeA into 13-HPOT; induced by wounding, MeJA, SA, and tea geometrid treatments [20] | wound-induced JA biosynthesis [29]; defense against herbivores [30]; resistance to Botrytis cinerea [31] | [20,21,22,23,24,25,26,27,28,29] |
| AOS | 114307969 | 1 | target the chloroplast, wound-induced JA biosynthesis with CsAOS2 transit overexpression [22] | wound-induced JA biosynthesis and male sterility [32]; resistance to Erwinia carotovora [33] | [22,32,33] |
| AOC | 114307969 | 1 | respond to MeJA, SA, tea leaf hopper, and tea geometrid treatment [23] | resistance to the blast fungus [34]; wound-induced JA biosynthesis [35] | [23,34,35] |
| OPR3 | 114299110 | 1 | (+)-cis-OPDA reductase activity [24]; restore wound-induced JA biosynthesis in Arabidopsis opr3 plants [14] | male sterility [36]; JA biosynthesis [37] | [14,24,26,27,28,29,30,31,32,33,34,35,36,37] |
| ACX | 114279706; 114266146 | 2 | induced by wounding, JA, tea geometrid and the tea green leafhopper treatments; restore wound-induced JA biosynthesis in acx1 plants [26] | wound-induced JA biosynthesis; defense against tobacco hornworm [38] | [26,38] |
| JMT | 114288630 | 1 | located in cytoplasm [27]; catalyze JA and SAM into MeJA [28] | resistance against fungus Botrytis cinerea; seed germination and mass [39] | [27,28,39] |
| JAZ1 | 114317541 | 1 | involved in synthesis of flavonoid [40,41] | interact with different proteins to modulate JA regulated physiological process [42,43,44,45,46] | [40,41,42,43,44,45,46] |
| MYC2 | 114266177 | 2 | involved in accumulating multiple JA-regulated defense compounds [23,40,47,48,49,50,51,52] | modulates diverse JA-dependent functions: development, and pathogen/wound response, etc. [6,18,19,53,54] | [6,18,19,23,40,47,48,49,50,51,52,53,54] |
3. Research Advances in JA Signaling Transduction
4. Defense Responses Regulated by JA in Tea Plants
4.1. JA Regulation in Biotic Stress Process
| Compounds | Functions | Regulatory Mechanism by JA | References |
|---|---|---|---|
| Polyphenol oxidases | reduce the larvae growth of E. grisescens | upregulate the CsPPO expression and increase the CsPPO activity | [15] |
| Catechins (EC, C, EGCG) | reduce the larvae growth of E. grisescens | increase the content of EC, C, and EGCG [9]; regulate expression of CsDFR/CsANR/CsLAR with CsJAZ1-CsMYC2.1 signaling module [40] | [9,40] |
| Quercetin glucosides | reduce the larvae growth of E. grisescens | induce the expression of biosynthesis gene CsUGT89AC1 via CsMYC2.1 | [65] |
| Indole | pre-exposure increases tea resistance to E. obliqua larvae | promote the production of indole, regulate CsTSB2 expression with CsJAZ2-CsMYC2.1 signaling module | [47] |
| α-Farnesene | attracting natural enemy wasps of E. grisescens larvae; alter the metabolite profile in the neighboring tea leaves; induce the emission of β-Ocimene in neighboring tea plants | regulate CsAFS expression through CsHDA6-CsMYC2.1 signaling module | [51] |
| Benzyl nitrile | repel E. grisescens larvae and inhibit their growth | CsCYP79 is the target gene of CsMYC2.1 and is positively regulated by CsMYC2.1 | [23] |
| (E)-Nerolidol | induce plant defenses against pathogens and insects [14]; repel mated E. obliqua females [70]; participate in cold stress tolerance [71] | increase the CsNES expression and (E)-nerolidol content through CsMYC2.1 [71] | [14,70,71] |
| β-Ocimene | alter the metabolite profile of neighboring tea plants [50]; repel mated E. obliqua females [70]; induce plant resistance against E. grisescens larvae [71]; | increase the expression of CsOCS and CsBOS1 and β-ocimene content [70,73] | [50,70,71,73] |
| Peroxidase | improve plant tolerance against mannitol stress | regulate expression of CsPER1 and CsPER3 via CsMYC2.1 | [74] |
4.2. JA Regulation in Abiotic Stress Processes
5. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Howe, G.A.; Major, I.T.; Koo, A.J. Modularity in jasmonate signaling for multistress resilience. Annu. Rev. Plant Biol. 2018, 69, 387–415. [Google Scholar] [CrossRef]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef]
- Onkokesung, N.; Baldwin, I.T.; Galis, I. The role of jasmonic acid and ethylene crosstalk in direct defense of Nicotiana attenuata plants against chewing herbivores. Plant Signal. Behav. 2010, 5, 1305–1307. [Google Scholar] [CrossRef]
- Yang, J.; Duan, G.; Li, C.; Liu, L.; Han, G.; Zhang, Y.; Wang, C. The crosstalks between jasmonic acid and other plant hormone signaling highlight the involvement of jasmonic acid as a core component in plant response to biotic and abiotic stresses. Front. Plant Sci. 2019, 10, 1349. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, S.; Wang, Y.; Zeng, W.; Jin, B. Plant responses to herbivory, wounding, and infection. Int. J. Mol. Sci. 2022, 23, 7031. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.; Yan, C.; Li, L.; Xie, D.; Li, C. Jasmonates. In Hormone Metabolism and Signaling in Plants; Li, J., Li, C., Smith, S.M., Eds.; Elsevier Academic Press: London, UK, 2017; pp. 243–272. [Google Scholar]
- Kolupaev, Y.E.; Yastreb, T.O. Jasmonate signaling and plant adaptation to abiotic stressors (Review). Appl. Biochem. Microbiol. 2021, 57, 1–19. [Google Scholar] [CrossRef]
- Zhu, Z. Jasmonate: The Swiss army knife in the plant’s pocket. J. Exp. Bot. 2023, 74, 1159–1161. [Google Scholar] [CrossRef]
- Li, X.; Zhang, J.; Lin, S.; Xing, Y.; Zhang, X.; Ye, M.; Chang, Y.; Guo, H.; Sun, X. (+)-Catechin, epicatechin and epigallocatechin gallate are important inducible defensive compounds against Ectropis grisescens in tea plants. Plant Cell Environ. 2022, 45, 496–511. [Google Scholar] [CrossRef]
- Zeng, L.; Watanabe, N.; Yang, Z. Understanding the biosyntheses and stress response mechanisms of aroma compounds in tea (Camellia sinensis) to safely and effectively improve tea aroma. Crit. Rev. Food Sci. Nutr. 2019, 59, 2321–2334. [Google Scholar] [CrossRef]
- Zeng, L.; Zhou, X.; Liao, Y.; Yang, Z. Roles of specialized metabolites in biological function and environmental adaptability of tea plant (Camellia sinensis) as a metabolite studying model. J. Adv. Res. 2021, 34, 159–171. [Google Scholar] [CrossRef]
- Qiao, D.; Yang, C.; Guo, Y.; Chen, J.; Chen, Z. Transcriptome and co-expression network analysis uncover the key genes mediated by endogenous defense hormones in tea plant in response to the infestation of Empoasca onukii Matsuda. Beverage Plant Res. 2023, 3, 4. [Google Scholar] [CrossRef]
- Gui, L.; Chen, Z.; Liu, S. Effect of exogenous methyl jasmonate-induced tea volatiles on host-selection behavior of insects. J. Tea Sci. 2004, 24, 166–171. [Google Scholar]
- Chen, S.; Zhang, L.; Cai, X.; Li, X.; Bian, L.; Luo, Z.; Li, Z.; Chen, Z.; Xin, Z. (E)-Nerolidol is a volatile signal that induces defenses against insects and pathogens in tea plants. Hortic. Res. 2020, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, X.; Ye, M.; Li, X.-W.; Lin, S.-B.; Sun, X.-L. The jasmonic acid pathway positively regulates the polyphenol oxidase-based defense against tea geometrid caterpillars in the tea plant (Camellia sinensis). J. Chem. Ecol. 2020, 46, 308–316. [Google Scholar] [CrossRef]
- Vick, B.; Zimmerman, D. Biosynthesis of jasmonic acid by several plant species. Plant Physiol. 1984, 75, 458–461. [Google Scholar] [CrossRef] [PubMed]
- Schaller, F. Enzymes of the biosynthesis of octadecanoid-derived signaling molecules. J. Exp. Bot. 2001, 52, 11–23. [Google Scholar] [CrossRef]
- Wasternack, C. Jasmonates: An update on biosynthesis, signal transduction and action in plant stress response, growth and development. Ann. Bot. 2007, 100, 681–697. [Google Scholar] [CrossRef]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- Chen, H. Identification and Characterization of a Herbivore-Induced Lipoxygenase Gene in the Leaves of Camellia sinensis Fed by Ectropis obliqua. Master Thesis, Anhui Agricultural University, Hefei, China, 2011. [Google Scholar]
- Zhu, J.; Wang, X.; Guo, L.; Xu, Q.; Zhao, S.; Li, F.; Yan, X.; Liu, S.; Wei, C. Characterization and alternative splicing profiles of the lipoxygenase gene family in tea plant (Camellia sinensis). Plant Cell Physiol. 2018, 59, 1765–1781. [Google Scholar] [CrossRef]
- Peng, Q.; Zhou, Y.; Liao, Y.; Zeng, L.; Xu, X.; Jia, Y.; Dong, F.; Li, J.; Tang, J.; Yang, Z. Functional characterization of an allene oxide synthase involved in biosynthesis of jasmonic acid and its influence on metabolite profiles and ethylene formation in tea (Camellia sinensis) flowers. Int. J. Mol. Sci. 2018, 19, 2440. [Google Scholar] [CrossRef]
- Wang, M.-X.; Ma, Q.-P.; Han, B.-Y.; Li, X.-H. Molecular cloning and expression of a jasmonate biosynthetic gene allene oxide cyclase from Camellia sinensis. Can. J. Plant Sci. 2016, 96, 109–116. [Google Scholar] [CrossRef]
- Xin, Z.; Zhang, J.; Ge, L.; Lei, S.; Han, J.; Zhang, X.; Li, X.; Sun, X. A putative 12-oxophytodienoate reductase gene CsOPR3 from Camellia sinensis, is involved in wound and herbivore infestation responses. Gene 2017, 615, 18–24. [Google Scholar] [CrossRef]
- Chen, S.; Lu, X.; Ge, L.; Sun, X.; Xin, Z. Wound- and pathogen-activated de novo JA synthesis using different ACX isozymes in tea plant (Camellia sinensis). J. Plant Physiol. 2019, 243, 153047. [Google Scholar] [CrossRef]
- Xin, Z.; Chen, S.; Ge, L.; Li, X.; Sun, X. The involvement of a herbivore-induced acyl-CoA oxidase gene, CsACX1, in the synthesis of jasmonic acid and its expression in flower opening in tea plant (Camellia sinensis). Plant Physiol. Biochem. 2019, 135, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jia, H.; Zhu, B.; Li, J.; Yang, T.; Zhang, Z.Z.; Deng, W.W. Molecular and biochemical characterization of jasmonic acid carboxyl methyltransferase involved in aroma compound production of methyl jasmonate during black tea processing. J. Agric. Food Chem. 2021, 69, 3154–3164. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Wang, J.; Zhao, Y.; Zhang, M.; Zhou, Z.; Li, Y.; Hu, Y.; Wu, Y.; Feng, Z.; Schwab, W.; et al. Biosynthesis of orchid-like volatile methyl jasmonate in tea (Camellia sinensis) leaves in response to multiple stresses during the shaking process of oolong tea. LWT 2021, 143, 111184. [Google Scholar] [CrossRef]
- Bell, E.; Creelman, R.A.; Mullet, J.E. A chloroplast lipoxygenase is required for wound-induced jasmonic acid accumulation in Arabidopsis. Proc. Natl. Acad. Sci. USA 1995, 92, 8675–8679. [Google Scholar] [CrossRef]
- Chauvin, A.; Caldelari, D.; Wolfender, J.; Farmer, E.E. Four 13-lipoxygenases contribute to rapid jasmonate synthesis in wounded Arabidopsis thaliana leaves: A role for lipoxygenase 6 in responses to long-distance wound signals. New Phytol. 2013, 197, 566–575. [Google Scholar] [CrossRef]
- Yan, L.; Zhai, Q.; Wei, J.; Li, S.; Wang, B.; Huang, T.; Du, M.; Sun, J.; Kang, L.; Li, C.; et al. Role of tomato lipoxygenase D in wound-induced jasmonate biosynthesis and plant immunity to insect herbivores. PLoS Genet. 2013, 9, e1003964. [Google Scholar] [CrossRef]
- Park, J.; Halitschke, R.; Kim, H.B.; Baldwin, I.T.; Feldmann, K.A.; Feyereisen, R. A knock-out mutation in allene oxide synthase results in male sterility and defective wound signal transduction in Arabidopsis due to a block in jasmonic acid biosynthesis. Plant J. 2002, 31, 1–12. [Google Scholar] [CrossRef]
- Pajerowska-Mukhtar, K.M.; Mukhtar, M.S.; Guex, N.; Halim, V.A.; Rosahl, S.; Somssich, I.E.; Gebhardt, C. Natural variation of potato allene oxide synthase 2 causes differential levels of jasmonates and pathogen resistance in Arabidopsis. Planta 2008, 228, 293–306. [Google Scholar] [CrossRef]
- Riemann, M.; Müller, A.; Korte, A.; Furuya, M.; Weiler, E.W.; Nick, P. Impaired induction of the jasmonate pathway in the rice mutant hebiba. Plant Physiol. 2003, 133, 1820–1830. [Google Scholar] [CrossRef] [PubMed]
- Riemann, M.; Haga, K.; Shimizu, T.; Okada, K.; Ando, S.; Mochizuki, S.; Nishizawa, Y.; Yamanouchi, U.; Nick, P.; Yano, M.; et al. Identification of rice Allene Oxide Cyclase mutants and the function of jasmonate for defence against Magnaporthe oryzae. Plant J. 2013, 74, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Stintzi, A.; Browse, J. The Arabidopsis male-sterile mutant, opr3, lacks the 12-oxophytodienoic acid reductase required for jasmonate synthesis. Proc. Natl. Acad. Sci. USA 2000, 97, 10625–10630. [Google Scholar] [CrossRef] [PubMed]
- Schaller, A.; Stintzi, A. Enzymes in jasmonate biosynthesis—Structure, function, regulation. Phytochemistry 2009, 70, 1532–1538. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Schilmiller, A.L.; Liu, G.; Lee, G.I.; Jayanty, S.; Sageman, C.; Vrebalov, J.; Giovannoni, J.J.; Yagi, K.; Kobayashi, Y.; et al. Role of β-oxidation in jasmonate biosynthesis and systemic wound signaling in tomato. Plant Cell 2005, 17, 971–986. [Google Scholar] [CrossRef]
- Seo, H.S.; Song, J.T.; Cheong, J.-J.; Lee, Y.-H.; Lee, Y.-W.; Hwang, I.; Lee, J.S.; Choi, Y.D. Jasmonic acid carboxyl methyltransferase: A key enzyme for jasmonate-regulated plant responses. Proc. Natl. Acad. Sci. USA 2001, 98, 4788–4793. [Google Scholar] [CrossRef]
- Zhu, J.; Yan, X.; Liu, S.; Xia, X.; An, Y.; Xu, Q.; Zhao, S.; Liu, L.; Guo, R.; Zhang, Z.; et al. Alternative splicing of CsJAZ1 negatively regulates flavan-3-ol biosynthesis in tea plants. Plant J. 2022, 110, 243–261. [Google Scholar] [CrossRef]
- Lv, W.; Jiang, H.; Cao, Q.; Ren, H.; Wang, X.; Wang, Y. A tau class glutathione S-transferase in tea plant, CsGSTU45, facilitates tea plant susceptibility to Colletotrichum camelliae infection mediated by jasmonate signaling pathway. Plant J. 2023. [Google Scholar] [CrossRef]
- Chini, A.; Fonseca, S.; Fernández, G.; Adie, B.; Chico, J.M.; Lorenzo, O.; García-Casado, G.; López-Vidriero, I.; Lozano, F.M.; Ponce, M.R.; et al. The JAZ family of repressors is the missing link in jasmonate signaling. Nature 2007, 448, 666–671. [Google Scholar] [CrossRef]
- Santner, A.; Estelle, M. The JAZ proteins link jasmonate perception with transcriptional changes. Plant Cell 2007, 19, 3839–3842. [Google Scholar] [CrossRef] [PubMed]
- Thines, B.; Katsir, L.; Melotto, M.; Niu, Y.; Mandaokar, A.; Liu, G.; Nomura, K.; He, S.Y.; Howe, G.A.; Browse, J. JAZ repressor proteins are targets of the SCFCOI1 complex during jasmonate signaling. Nature 2007, 448, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ran, W.; Zhang, J.; Ye, M.; Lin, S.; Li, X.; Sultana, R.; Sun, X. Genome-wide identification of the Tify gene family and their expression profiles in response to biotic and abiotic stresses in tea plants (Camellia sinensis). Int. J. Mol. Sci. 2020, 21, 8316. [Google Scholar] [CrossRef] [PubMed]
- Chini, A.; Boter, M.; Solano, R. Plant oxylipins: COI1/JAZs/MYC2 as the core jasmonic acid-signaling module. FEBS J. 2009, 276, 4682–4692. [Google Scholar] [CrossRef]
- Zhou, Y.; Zeng, L.; Hou, X.; Liao, Y.; Yang, Z. Low temperature synergistically promotes wounding-induced indole accumulation by INDUCER OF CBF EXPRESSION-mediated alterations of jasmonic acid signaling in Camellia sinensis. J. Exp. Bot. 2020, 71, 2172–2185. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Liao, Y.; Jian, G.; Jia, Y.; Zeng, L.; Gu, D.; Li, H.; Yang, Y. Light induces an increasing release of benzyl nitrile against diurnal herbivore Ectropis grisescens Warren attack in tea (Camellia sinensis) plants. Plant Cell Environ. 2023, 46, 3464–3480. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Liu, M.; Erb, M.; Glauser, G.; Zhang, J.; Li, X.; Sun, X. Indole primes defence signaling and increases herbivore resistance in tea plants. Plant Cell Environ. 2021, 44, 1165–1177. [Google Scholar] [CrossRef]
- Zeng, L.; Liao, Y.; Li, J.; Zhou, Y.; Tang, J.; Dong, F.; Yang, Z. alpha-Farnesene and ocimene induce metabolite changes by volatile signaling in neighboring tea (Camellia sinensis) plants. Plant Sci. 2017, 264, 29–36. [Google Scholar] [CrossRef]
- Wu, S.; Yang, Y.; Chen, J.; Li, J.; Jian, G.; Yang, J.; Mao, K.; Zeng, L.; Gu, D. Histone deacetylase CsHDA6 mediates the regulated formation of the anti-insect metabolite alpha-farnesene in tea (Camellia sinensis). Plant Sci. 2023, 326, 111501. [Google Scholar] [CrossRef]
- Han, Z.; Zhang, C.; Zhang, H.; Duan, Y.; Zou, Z.; Zhou, L.; Zhu, X.; Fang, W.; Ma, Y. CsMYB transcription factors participate in jasmonic acid signal transduction in response to cold stress in tea plant (Camellia sinensis). Plants 2022, 11, 2869. [Google Scholar] [CrossRef]
- Kazan, K.; Manners, J.M. MYC2: The master in action. Mol. Plant 2013, 6, 686–703. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Zhao, J.; Tzeng, D.T.; Liu, Y.; Deng, L.; Yang, T.; Zhai, Q.; Wu, F.; Huang, Z.; Zhou, M.; et al. MYC2 orchestrates a hierarchical transcriptional cascade that regulates jasmonate-mediated plant immunity in tomato. Plant Cell 2017, 29, 1883–1906. [Google Scholar] [CrossRef] [PubMed]
- Howe, G.A. Ubiquitin ligase-coupled receptors extend their reach to jasmonate. Plant Physiol. 2010, 154, 471–474. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Zou, Z.; Xing, H.; Duan, Y.; Zhu, X.; Ma, Y.; Wang, Y.; Fang, W. Genome-wide analysis reveals stress and hormone responsive patterns of JAZ family genes in Camellia sinensis. Int. J. Mol. Sci. 2020, 21, 2433. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Chen, X.; Wang, P.; Sun, Y.; Yue, C.; Ye, N. Genome-wide and expression pattern analysis of JAZ family involved in stress responses and postharvest processing treatments in Camellia sinensis. Sci. Rep. 2020, 10, 2792. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Kong, Y.; Zhang, X.; Liao, Z.; He, Y.; Li, L.; Liang, Z.; Sheng, Q.; Hong, G. Structural and functional organization of the MYC transcriptional factors in Camellia sinensis. Planta 2021, 253, 93. [Google Scholar] [CrossRef]
- Yang, Z.-W.; Duan, X.-N.; Jin, S.; Li, X.-W.; Chen, Z.-M.; Ren, B.-Z.; Sun, X.-L. Regurgitant derived from the tea geometrid Ectropis obliqua suppresses wound-induced polyphenol oxidases activity in tea plants. J. Chem. Ecol. 2013, 39, 744–751. [Google Scholar] [CrossRef]
- Dombrecht, B.; Xue, G.P.; Sprague, S.J.; Kirkegaard, J.A.; Ross, J.J.; Reid, J.B.; Fitt, G.P.; Sewelam, N.; Schenk, P.M.; Manners, J.M.; et al. MYC2 differentially modulates diverse jasmonate-dependent functions in Arabidopsis. Plant Cell 2007, 19, 2225–2245. [Google Scholar] [CrossRef]
- Browse, J. Jasmonate passes muster: A receptor and targets for the defense hormone. Annu. Rev. Plant Biol. 2009, 60, 183–205. [Google Scholar] [CrossRef]
- Campos, M.L.; Kang, J.-H.; Howe, G.A. Jasmonate-triggered plant immunity. J. Chem. Ecol. 2014, 40, 657–675. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, S.; Sun, N.; Liu, H.; Zhao, Y.; Liang, Y.; Zhang, L.; Han, Y. Functional diversity of jasmonates in rice. Rice 2015, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Xu, M.; Cai, X.; Han, Z.; Si, J.; Chen, D. Jasmonate signaling pathway modulates plant defense, growth, and their trade-offs. Int. J. Mol. Sci. 2022, 23, 3945. [Google Scholar] [CrossRef] [PubMed]
- Jing, T.; Du, W.; Qian, X.; Wang, K.; Luo, L.; Zhang, X.; Deng, Y.; Li, B.; Gao, T.; Zhang, M.; et al. UGT89AC1-mediated quercetin glucosylation is induced upon herbivore damage and enhances Camellia sinensis resistance to insect feeding. Plant Cell Environ. 2023, 47, 682–697. [Google Scholar] [CrossRef] [PubMed]
- Aljbory, Z.; Chen, M.S. Indirect plant defense against insect herbivores: A review. Insect Sci. 2018, 25, 2–23. [Google Scholar] [CrossRef]
- Yang, H.; Wang, Y.; Li, L.; Li, F.; He, Y.; Wu, J.; Wei, C. Transcriptomic and phytochemicalanalyses reveal root-mediated resource-based defense responseto leaf herbivory by Ectropis oblique in tea plant (Camellia sinensis). J. Agric. Food Chem. 2019, 67, 5465–5476. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Bian, L.; Luo, Z.; Li, Z.; Xiu, C.; Fu, N.; Cai, X.; Chen, Z. Enhanced volatile emissions and anti-herbivore functions mediated by the synergism between jasmonic acid and salicylic acid pathways in tea plants. Hortic. Res. 2022, 9, uhac144. [Google Scholar] [CrossRef]
- Dong, F.; Yang, Z.; Baldermann, S.; Sato, Y.; Asai, T.; Watanabe, N. Herbivore-induced volatiles from tea (Camellia sinensis) plants and their involvement in intraplant communication and changes in endogenous nonvolatile metabolites. J. Agric. Food Chem. 2011, 59, 13131–13135. [Google Scholar] [CrossRef]
- Jing, T.; Qian, X.; Du, W.; Gao, T.; Li, D.; Guo, D.; He, F.; Yu, G.; Li, S.; Schwab, W.; et al. Herbivore-induced volatiles influence moth preference by increasing the beta-Ocimene emission of neighbouring tea plants. Plant Cell Environ. 2021, 44, 3667–3680. [Google Scholar] [CrossRef]
- Gu, D.; Wu, S.; Yu, Z.; Zeng, L.; Qian, J.; Zhou, X.; Yang, Z. Involvement of histone deacetylase CsHDA2 in regulating (E)-nerolidol formation in tea (Camellia sinensis) exposed to tea green leafhopper infestation. Hortic. Res. 2022, 9, uhac158. [Google Scholar] [CrossRef]
- Jian, G.; Jia, Y.; Li, J.; Zhou, X.; Liao, Y.; Dai, G.; Zhou, Y.; Tang, J.; Zeng, L. Elucidation of the regular emission mechanism of volatile beta-ocimene with anti-insect function from tea plants (Camellia sinensis) exposed to herbivore attack. J. Agric. Food Chem. 2021, 69, 11204–11215. [Google Scholar] [CrossRef]
- Chen, S.; Xie, P.; Li, Y.; Wang, X.; Liu, H.; Wang, S.; Han, W.; Wu, R.; Li, X.; Guan, Y.; et al. New insights into stress-induced beta-ocimene biosynthesis in tea (Camellia sinensis) leaves during oolong tea processing. J. Agric. Food Chem. 2021, 69, 11656–11664. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Chen, H.; Liu, L.; Xia, X.; Yan, X.; Mi, X.; Liu, S.; Wei, C. JA-mediated MYC2/LOX/AOS feedback loop regulates osmotic stress response in tea plant (Camellia sinensis). Hortic. Plant J. 2023, in press. [Google Scholar] [CrossRef]
- Lu, Q.; Wang, Y.; Xiong, F.; Hao, X.; Zhang, X.; Li, N.; Wang, L.; Zeng, J.; Yang, Y.; Wang, X. Integrated transcriptomic and metabolomic analyses reveal the effects of callose deposition and multihormone signal transduction pathways on the tea plant-Colletotrichum camelliae interaction. Sci. Rep. 2020, 10, 12858. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, L.; He, Y.; Lang, Z.; Zhao, Y.; Tao, H.; Li, Q.; Hong, G. The CsHSFA-CsJAZ6 module-mediated high temperature regulates flavonoid metabolism in Camellia sinensis. Plant Cell Environ. 2023, 46, 2401–2418. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Yu, Y.; Zhang, J.; Qian, X.; Li, X.; Sun, X. Recent Progress Regarding Jasmonates in Tea Plants: Biosynthesis, Signaling, and Function in Stress Responses. Int. J. Mol. Sci. 2024, 25, 1079. https://doi.org/10.3390/ijms25021079
Zhang X, Yu Y, Zhang J, Qian X, Li X, Sun X. Recent Progress Regarding Jasmonates in Tea Plants: Biosynthesis, Signaling, and Function in Stress Responses. International Journal of Molecular Sciences. 2024; 25(2):1079. https://doi.org/10.3390/ijms25021079
Chicago/Turabian StyleZhang, Xin, Yongchen Yu, Jin Zhang, Xiaona Qian, Xiwang Li, and Xiaoling Sun. 2024. "Recent Progress Regarding Jasmonates in Tea Plants: Biosynthesis, Signaling, and Function in Stress Responses" International Journal of Molecular Sciences 25, no. 2: 1079. https://doi.org/10.3390/ijms25021079
APA StyleZhang, X., Yu, Y., Zhang, J., Qian, X., Li, X., & Sun, X. (2024). Recent Progress Regarding Jasmonates in Tea Plants: Biosynthesis, Signaling, and Function in Stress Responses. International Journal of Molecular Sciences, 25(2), 1079. https://doi.org/10.3390/ijms25021079